

Abstract
A successful procedure for studying imitative behavior in non-humans is the bidirectional control procedure in which observers are exposed to a demonstrator that responds by moving a manipulandum in one of two different directions (e.g., left vs. right). Imitative learning is demonstrated when observers make the response in the direction that they observed it being made. This procedure controls for socially mediated effects (the mere presence of a demonstrator), stimulus enhancement (attention drawn to a manipulandum by its movement), and if an appropriate control is included, emulation (learning how the environment works). Recent research with dogs has found that dogs may not demonstrate imitative learning when the demonstrator is human. In the present research, we found that when odors were controlled for, dogs imitated the direction of a screen push demonstrated by another dog more than in a control condition in which they observed the screen move independently while another dog was present. Furthermore, we found that dogs would match the direction of screen push demonstrated by a human and they were equally likely to match the direction in which the screen moved independently while a human was present.
Keywords: Imitation, dogs, bidirectional control, emulation, cross-species model
Recently, imitative learning in dogs has attracted considerable research interest (Range, Viranyi & Huber, 2007; Topál, Byrne, Miklósi, Csanyi, 2006; Kubinyi, Topál, Miklósi, & Csányi, 2003), This interest can be attributed in part to the fact that dogs are intelligent carnivores that historically lived in complex social groups (Bekoff, 1995) and that during the domestication process, dogs may have been selected for human-like cognitive abilities such as imitation. Imitation is of special interest because the term implies that an observer can watch a demonstrator perform an improbable behavior and then engage in that behavior (Zentall, 1996).
It has been proposed that imitation implies that the observer understands the relationship between its own behavior and the behavior being modeled by a demonstrator (Piaget, 1962). The recent interest in imitation in dogs has been facilitated by the development of new paradigms that allow one to test for imitative learning while controlling for alternative, often less cognitive accounts of behavioral matching.
For example, some animals show species typical behaviors that can be elicited when one member of a group observes another member engage in a certain behavior (Thorpe, 1963). Such reflexive behavior, often called contagion, is triggered automatically and immediately follows the behavior that was observed. Yawning in humans, and herding or flocking in animals are examples of contagious behavior. To control for contagious behavior, research on imitation in animals should study the acquisition of improbable behaviors that are not likely to be reflexive.
It is also possible that the mere presence of another animal can alter the probability of certain behavior. Zajonc (1965) suggested that the mere presence of a conspecific may increase the level of arousal in an observer, and an increase in arousal can affect the general activity of the observer, leading to a change in the probability of performing a response (Levine & Zentall, 1974). Whether mere presence tends to facilitate or retard acquisition may depend on the nature of the response but in general it is important to include a control for such effects.
Perceptual factors can also increase the likelihood of the acquisition of a response. For example, the presence of a conspecific at a particular location can increase the salience of that location (local enhancement), or the movement of an object (e.g., a lever; stimulus enhancement) by a demonstrator can increase the salience of the object. If the behavior of the demonstrator attracts the observer's attention to a location or object, it can increase the probability that the observer makes the appropriate response. For example, Lorenz (1935/1970) found that ducks were more likely to escape from their pen through a hole in the fence if they observed another duck move through the hole. Similarly, there is evidence that dogs that observe a human carrying a target object to the inside of a V-shaped fence, will detour the fence from the apex more quickly than dogs that do not observe the demonstration (Pongrácz et al., 2001; Pongrácz, Miklósi, Kubinyi, Topál & Csányi, 2003; Pongrácz, Miklósi, Tímár-Geng, & Csányi, 2003). However such effects could be produced by drawing the dog's attention to the point at which the dog could access the inner area of the V-shaped fence (local enhancement).
A means of controlling for stimulus and local enhancement called the bidirectional control procedure was developed by Heyes and Dawson (1990). They trained demonstrator rats to push an overhead rod either to the left or to the right. Hungry rats observed the demonstrator push the rod in one direction, and then they were given access to the rod and were rewarded for pushes in either direction. Heyes and Dawson found that observer rats showed a significant tendency to push the rod in the same direction as they observed it pushed. However, Mitchell, Heyes, Gardner, and Dawson (1999) showed that olfactory cues, rather than visual cues, were probably responsible for the matching behavior.
Klein and Zentall (2003) used the bidirectional control procedure to test for imitative learning in pigeons, a species with good visual acuity but less likely to be influenced by odor cues. In their design, observer pigeons viewed one of two different demonstrations. They either observed a demonstrator push a feeder-blocking screen to the left or right of the feeder opening, or they observed the screen move unobtrusively by the experimenter with another pigeon present (a control for emulation learning and social facilitation). The control group was included because under these conditions the observer could learn the relationship between the object movement and the salient outcome and instead of matching the behavior of the demonstrator, the observer might produce the outcome via object movement reenactment (copying the way object move), a type of emulation learning (Whiten & Ham, 1992). Klein and Zentall found that following the conspecific demonstration, observer pigeons showed a significant tendency to push the screen in the same direction as they had observed it being pushed. However, observer pigeons that viewed the screen move unobtrusively by the experimenter with another pigeon present did not match the direction of screen movement. Thus, it appears that the bidirectional control procedure, with appropriate control for emulation and social facilitation, is an effective procedure for assessing imitative learning. Similar results have been found using Japanese quail as subjects (Akins, Klein, & Zentall, 2002).
In spite of this strong evidence for imitation in birds using the bidirectional control procedure, Kubinyi et al. (2003) did not report similar findings using this procedure with dogs. In their design, dogs observed one of several demonstrations. In one condition, dogs observed a human (owner) push a handle to the left or right to release a ball from a box. The owner then initiated play with the dog and the ball. Dogs in another condition observed the owner touch the handle of the box without releasing the ball, and in two other conditions dogs observed their owners either touch the top of the box or not interact with the box at all. The latter three conditions controlled for the possible effects of social facilitation and stimulus enhancement as they did not produce the ball. Odor cues were controlled by having the owner rub the handle prior to demonstrating. After the demonstration, the dogs were given test trials in which they were allowed to manipulate the handle of the box. The authors found that dogs that observed their owners manipulate the handle tended to manipulate the handle more than the other groups, however, in general, they did not move the handle in the direction demonstrated. Thus, these dogs did not show evidence of imitative learning. Rather, the results suggest that the demonstration enhanced the salience of the handle (stimulus enhancement). When manipulation of the handle was associated with a rewarding event, the dogs were more attracted to it and were more likely to manipulate it.
To account for the fact that pigeons are more likely to imitate using the bidirectional control procedure than rats or dogs one could propose that it is because they have a more highly developed visual system that enables them observe the behavior of others. Alternatively, there are several differences in the procedure used by Klein and Zentall (2003) and those used by Heyes and Dawson (1990) and by Kubinyi et al. (2003) that might account for the differences in results found. In the case of the rats, Heyes and Dawson (1990) confounded the visual observation cues with olfactory cues and when the confound was resolved by pitting the two against each other (Mitchell et al. 1999), the rats behavior appeared to be controlled by the olfactory cues.
In the case of the Kubinyi et al. (2003) experiment, it is possible that dogs do not naturally imitate humans. Alternatively, the difference in findings between Klein and Zentall (2003) with pigeons and those of Kubinyi et al. (2003) with dogs may be the fact that Klein and Zentall used a response that was easier to acquire (pushing aside an obstacle to get to reinforcement).
In contrast to the Kubinyi et al. (2003) results, there is evidence that dogs may be able to acquire the presumably more complex concept of imitation in the form of a do-as-I-do task (Topál et. al, 2006). In this study, Philip, a service dog, was trained to reproduce several actions performed by his trainer when followed by a “do it” command. When Philip was then shown several new actions followed by the “do it” command he performed them at a reasonably high level of accuracy.
There is also evidence that dogs have the remarkable ability to imitate selectively based on an inference about the necessity of the matching response. When dogs could make a response to obtain food using their paw or their mouth, they showed a preference for using their mouth. However, when a demonstrator dog used its paw to obtain food rather than its mouth, observers too used their paw. Remarkably, when the demonstrator dog's mouth was occupied with a ball and it used its paw, observers used their mouth. That is, when the demonstrator appeared to have the option of using its mouth or its paw but used its paw, observers matched the demonstrator's behavior. However, when the demonstrator appeared to be unable to use its mouth because it was occupied, the observer presumably inferred that it could use its mouth because it was not occupied.
Given the results of the more complex form of the imitation task used by Range et al. (2007) and by Topál et al. (2006), as well as the failure to find imitation using the bidirectional procedure reported by Kubinyi et al. (2003), the purpose of the present experiment was to assess imitative learning in dogs by using a bidirectional control procedure with an apparatus similar to the one used with pigeons. A mesh screen that covered an opening in a panel of wood could be pushed to the left or right. Each dog observed a demonstration of the screen moving to the left or to the right to obtain food from an experimenter sitting behind the screen. The dog imitation group observed a conspecific demonstrate the required action. This group was included to determine whether dogs could imitate another dog (a conspecific). The social-facilitation/emulation control group observed a conspecific sit in front of the screen and wait for the screen to be moved unobtrusively by the experimenter located behind the apparatus, who made the screen move by pulling on the fishing line attached to either side of the screen. This group was included to control for the (arousal or motivational) effects that the presence of a conspecific may have had on emulation learning associated with the movement of the screen. The human imitation group observed a human demonstrator move the screen with her hand and was included to determine whether dogs can imitate humans. The emulation group observed the screen being moved inconspicuously by the experimenter located behind the apparatus. A second human (the “demonstrator”) was present but she did not touch the screen. This last group was included as a control for the human imitation group and to determine whether dogs can emulate the movement of the screen (without a conspecific present). Following the demonstration, the demonstrator was removed (if present) and the observer was released and rewarded for pushing the screen in either direction.
Method
Subjects
Sixty seven dogs (Canis familiaris), 23 males and 31 females, which belonged to private owners (ages ranging from 3 to 122 months, M = 44.7 months) were recruited. All dog owners were given a short questionnaire. Owners needed to confirm that their dogs matched several selection criteria. All dogs needed to be highly motivated by the opportunity to interact with the experimenters. Additionally, they needed to be highly motivated by food reinforcers. Finally, the owners had to be willing to deprive the dog of food for at least 4 hours prior to participating.
After owners affirmed that their dogs met the qualifications, plans were made to test the dog at a location familiar to them. The experimenters found that some dogs (N=13) did not meet the qualifications. Some dogs were frightened (avoided the experimenters and/or the apparatus) and others were disinterested in food reinforcers (they did not immediately consume food offered by the experimenter). If the opinions of the owner and the experimenters were in conflict, the dog was not included in the experiment. Of the dogs that participated in the experiment 25 came from breeds classified by the American Kennel Club as herding dogs (14 Belgian Tervuren, 4 Australian Shepherds, 4 border collies, 2 German shepherds and a Belgian Sheepdog), 2 were sporting dogs (a Portuguese water dog, and a Labrador retriever), 2 were non-sporting dogs (2 poodles) one was a hound dog (Rhodesian Ridgeback), 10 were working dogs (Akitas), one was a terrier (Parson Russell terrier) one was a toy dog (Yorkshire terrier) and 12 were of mixed breeding. All of these dogs had been trained to sit and to walk next to their owners on command.
Apparatus
The apparatus consisted of a plywood panel (94 cm high × 124 cm wide × 1.27 cm thick) that was attached by hinges on both sides to two additional panels of wood (94 cm high × 63.5 cm wide × 1.27 cm thick). Both the panel attached to the right and left of the central panel could be moved so that the apparatus was supported in an upright position. All three panels were painted flat black. A square hole (35.5 cm high × 30.5 cm wide) was cut out of the center of the middle panel. A drawer slide (KV model 1129) was attached to the central panel above the square hole. An aluminum screen (35.5 cm high × 30.5 cm wide × 1.27 cm thick) was mounted to the drawer slide such that it covered the hole completely and could be moved an equal distance (16.5 cm) to the left or right of the hole. Transparent fishing line (5.5 kg test, 0.3 mm diameter) was attached to each side of the screen to allow the screen to be moved by the experimenter in either direction without approaching the screen. Reinforcement was provided in a blue bowl (15 cm diameter and 7 cm deep). For dogs the reinforcement consisted of Pet Botanics® semi-moist Chicken & Brown Rice dinner dog food that was cut into portions of approximately 2 gm. The human demonstrator was reinforced with a Dorito® from a separate blue bowl.
Procedure
Demonstrator training
A single dog (48 month old female Belgian sheepdog) was trained to serve as the demonstrator. She was familiar (had previously interacted) with half of the observers. The demonstrator was shaped to push the screen with its muzzle left and right by rewarding successive approximations. The discriminative cue (that served to indicate which direction to push the screen) was a single finger point to one side of the screen from the experimenter located behind the apparatus. The cue was given before demonstration trials were initiated. The demonstrator sitting in front of the screen was able to notice this subtle cue despite the fact that it was given from behind the screen (but the cue could not be seen by the observer that sat to the side). The demonstrator was also trained to sit and wait for the screen to move left or right. A successful screen-push response was defined as a push that moved the screen far enough for the dog to insert its head into the opening in the center of the middle panel. Each successful response was rewarded by an experimenter who knelt behind the screen. The experimenter moved a blue bowl (that contained a piece of food) within the reach of the dog and offered verbal praise.
In the human demonstrator condition, one of the experimenters (a 25 yr old female) served as the demonstrator. She was familiar with three of the dogs. The demonstrator crouched several feet in front of the screen and waited until the experimenter gave the release signal. The demonstrator then moved forward and pushed the screen with her hand. The experimenter moved the blue bowl (which contained a Dorito®) within the demonstrators reach and offered verbal praise. The demonstrator consumed the reinforcer. Every attempt was made to make the demonstration given by the human demonstrator as similar to that of the dog demonstrator, and the reinforcement given to the demonstrator similar to that given to the dog demonstrator. All demonstration trials were conducted prior to testing.
Odor control
Odor cues were controlled in several ways. First, the dog demonstrator's saliva and the observer's saliva were applied to both sides of the screen before each demonstration. This was accomplished by having the experimenter rub her hands along the dogs head and muzzle and then rubbing both sides of the screen with her hands. To further control for odor cues, the side to which the screen was pushed alternated between observers. Thus, if the first dog observed the screen move left, the next dog observed it move to the right.
Observer training
Observers were given a treat from the experimenter who was standing behind the apparatus. This experimenter reached over the apparatus so that the dog could obtain the treat without interacting with the screen. This was intended to demonstrate to the observer that food rewards were available from the experimenter who was located behind the apparatus and to familiarize the observer with the apparatus and the experimenter.
In the dog imitation group, observers observed the conspecific demonstrator successfully push the screen in one direction to obtain a food reinforcer. In the social-facilitation/emulation group, the experimenter sitting behind the screen moved the screen in one direction by pulling on the fishing line that was attached to either side of the screen. Thus, the observers observed the screen move (apparently by itself) while the conspecific demonstrator sat and waited. Once the screen had moved, the demonstrator approached the opening in the panel and received reinforcement. In the human imitation group, observers observed a human demonstrator push the screen in one direction to obtain a food reinforcer. In the emulation group, the experimenter sitting behind the screen moved the screen in one direction by pulling on the fishing line that was attached to either side of the screen. Thus, the observers observed the screen move (apparently by itself) without a conspecific present. After the screen moved, the experimenter offered verbal praise and displayed the blue bowl at the opening.
Observers were randomly assigned to the four groups. The sample size was equal for all of the groups (N=12), except the emulation group (N=18). An equal number of dogs observed the screen move to the left or the right in each group. Observers were placed at a 45° angle to the center panel and at a distance of about 90 cm from the screen. Half of the dogs observed the first 6 demonstration trials from a position to the right of the screen and the last 6 demonstration trials from a position to the left of the screen. The remaining dogs experienced the opposite order. Each session consisted of 12 observation trials.
Testing
After completing the 12 demonstration trials, the demonstrator was removed (if present) and the observer was placed about 1.25 m from the front of the apparatus. The owner stood directly behind the observer and held onto to the dog by a leash. The owner was then told to look directly ahead, avoid eye contact with the dog, to avoid looking at the screen, and to remain silent. This was done to avoid having the owner provide any inadvertent cues as to the direction the screen should be pushed. The experimenter then arranged a release signal such as “Okay!” with the owner. Upon giving the release signal, the owner allowed the dog to approach the apparatus by releasing the tension on the leash that they held or by dropping it. Once the observer pushed the screen in either direction, the dog was rewarded with food from the blue bowl and was given verbal praise. Each observer dog received 6 test trials. If an observer did not push the screen within 120 sec the session was terminated.
Data analyses
Data from each group was compared to chance on the first trial using a two-tailed binomial test and pooled over the 6 test trials using a two-tailed t test. Between group comparisons were made on the first trial data using a χ2 test and pooled over the 6 test trials using a two-tailed t test.
Results
All observers pushed the screen at least once during testing. Most of the dogs (47 out of 54) pushed the screen on all 6 test trials: all of the observers in the social-facilitation/emulation group, 11 of the 12 observers in the dog imitation group, 8 of the 12 observers in the human imitation group, and 16 of the 18 observers in the emulation group. Thus, each observer dog received a proportion score consisting of the number of pushes that matched the direction demonstrated, divided by the number of total responses made.
Dog imitation group
On the first test trial, 11 of the 12 dogs in the dog imitation group pushed the screen in the same direction that they saw the screen-push demonstrated (92%), a number that was statistically different from chance according to a binomial test, p = .003. When pooled over the 6 test trials the percentage of matching screen pushes was 77.1%, a percentage that was also significantly different from chance, t = 2.69, p = .02.
Social-facilitation/emulation group
On the first test trial, 6 of the 12 dogs in the social-facilitation/emulation group pushed the screen in the same direction that they saw the screen-push demonstrated (50%). When pooled over the 6 test trials the percentage of matching screen pushes was also 50%.
Human imitation group
On the first test trial, 9 of the 12 dogs in the human imitation group pushed the screen in the same direction that they saw the screen-push demonstrated (75%), a number that was marginally different from chance according to a binomial test, p = .073. When pooled over the 6 test trials the percentage of matching screen pushes was 68.1%, a percentage that was not significantly different from chance, t = 1.46, p = .17.
Emulation group
When pooled over the 6 test trials the percentage of matching screen pushes was 68.5%, a percentage that was significantly different from chance, t = 2.10, p = .05. On the first test trial, most of the dogs in the emulation group (12 of 18) pushed the screen in the same direction that they saw the screen-push demonstrated (67%), however, this difference was not significantly different from chance as indicated by a binomial test, p = .12.
Imitation
To test for imitation of a conspecific, we compared the performance of observers in the dog imitation group to that of observers in the social-facilitation/emulation group. A chi-square (nonparametric) test was used to compare correspondence of the first response by each observer with the direction of the demonstration. Observers in the dog imitation group pushed the screen in the direction demonstrated on the first trial more often (11 of 12) than the dogs in the social-facilitation/emulation group (6 of 12), χ2 = 5.04, p < .05. When performance pooled over the 6 test trials was compared using an independent sample t-test, the difference in performance approached statistical significance t(22) = 1.77, p = .09. As performance by the dog imitation group was significantly above chance and performance by the social-facilitation/emulation group was exactly at chance, the results of this group comparison can be attributed to the high variability in the performance of the social-facilitation/emulation group (SD = ±44.4%; eight of the dogs always pushed the screen in one direction, four in the same directions as that observed, four in the opposite direction).
To test for imitation of a human, we compared the performance of observers in the human imitation group to that of observers in the emulation group. A chi-square analysis performed on the first trial scores for dogs in the human imitation group (8 of 10) and in the emulation group (12 of 18) indicated that the difference between groups was not statistically significant, χ2 < 1. Furthermore, performance pooled over the 6 test trials for the human imitation group (73.3%) and the emulation group (68.5%) did not differ significantly, t(28) < 1.
Familiarity
Familiarity with the demonstrator did not significantly affect the performance of dogs in the imitation conditions. The six dogs that were familiar to the dog demonstrator matched the direction demonstrated (87.5%) more than the six dogs that were unfamiliar (66.7%), but not significantly so, t(10) = 1.18, p = .26. The three dogs that were familiar to the human demonstrator were equally likely to match the direction demonstrated as the nine dogs that were unfamiliar to the demonstrator, t(10) < 1, (66.7% and 68.1%, respectively).
Discussion
Observers in the dog imitation condition were more likely to match their first response by pushing in the direction demonstrated than observers in the social-facilitation/emulation control condition. Thus, there is evidence for imitation of the screen push by these dogs.
The results of the human imitation group are a bit more difficult to interpret. The performance of this group was similar to the performance of the dog imitation group and even more similar to the performance of the emulation group. However, because of the variability of performance by dogs in this group, the imitation effect was not statistically reliable.
On the other hand, the results suggest that dogs are able to learn from emulating the direction in which the screen moved. That is, in the absence of a human model or the mere presence of another dog, observers appear to match the direction that they observe the screen move. This result is important in its own right because the mechanism responsible for emulation is not obvious.
Some have proposed that emulation might result from a Pavlovian association between the direction that the screen moves (e.g., a screen moving right) and food. Such an association has sometimes been referred to as observational conditioning (see Zentall, 1996). Such an explanation might account for a preference for a right-moving screen (the stimulus associated with the sight of food which should be a conditioned reinforcer), but it does not explain how the observer understands that it should move the screen to the right.
The present results parallel the results of a similar bidirectional control experiment with pigeons (Klein & Zentall, 2003). Klein and Zentall found that pigeons that observed the screen push by a pigeon demonstrator tended to push the screen in the direction they observed it pushed, whereas pigeons in a social facilitation emulation group did not. They also found that pigeons in a pure emulation group tended to push the screen in the direction that they observed it move.
The results of the present experiment add to the growing literature on the cognitive abilities of dogs (see e.g., Udell & Wynne, 2008). Not only can dogs learn to imitate on command – to “do as I do” (Topál et al. 2006), and appear to “understand” when a demonstrator dog must use its paw to operate a lever because its mouth is occupied, rather than chooses to use its paw because its mouth is free (Range et al., 2007), but they also can match the direction of a demonstrated screen push. Given the abundance of dogs that live with humans and the various tasks that they have been trained to perform, it is surprising that they have not been the subject of nearly as much experimentation as the more favored experimental animals, rats and pigeons.
Figure 1.

(a) Apparatus with dog at the start of a test trial. (b) Apparatus as dog is starting to make a screen push response.
Figure 2.
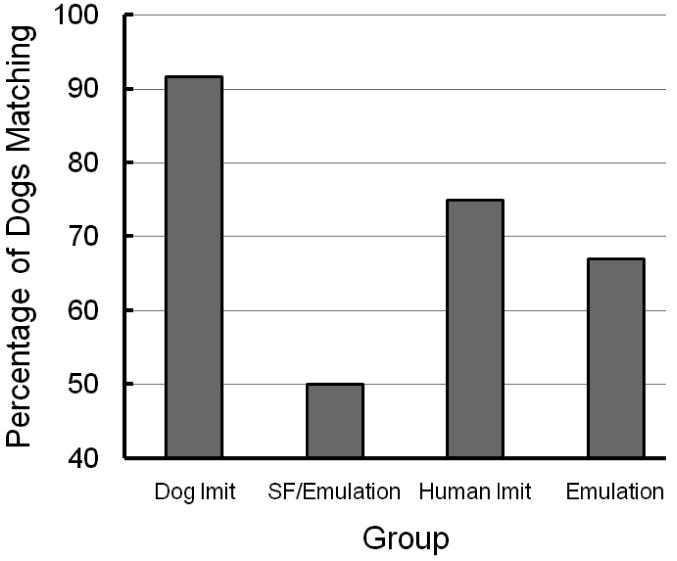
Percentage of dogs in each group that made a matching screen-push response on the first test trial.
Figure 3.
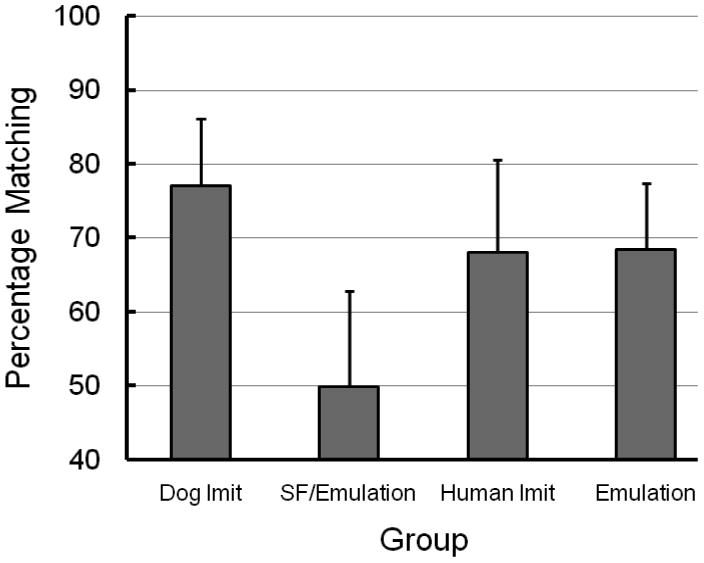
Percentage of matching screen-push responses pooled over the 6 test trials (error bars = ±SEM) for dogs in each group.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Akins CK, Klein ED, Zentall TR. Imitative learning in Japanese quail (Coturnix japonica) using the bidirectional control procedure. Animal Learning & Behavior. 2002;30:275–281. doi: 10.3758/bf03192836. [DOI] [PubMed] [Google Scholar]
- Bekoff M. Play signals as punctuation: the structure of social play in canids. Behaviour. 1995;132:419–429. [Google Scholar]
- Heyes CM, Dawson GR. A demonstration of observational learning using a bidirectional control. Quarterly Journal of Experimental Psychology: Comparative and Physiological Psychology. 1990;42(B):59–71. [PubMed] [Google Scholar]
- Klein ED, Zentall TR. Imitation and affordance learning by pigeons (Columba livia) Journal of Comparative Psychology. 2003;117:414–419. doi: 10.1037/0735-7036.117.4.414. [DOI] [PubMed] [Google Scholar]
- Kubinyi E, Topál J, Miklósi Á, Csányi V. Dogs learn from their owner via observation in a manipulation task. Journal of Comparative Psychology. 2003;117:156–165. doi: 10.1037/0735-7036.117.2.156. [DOI] [PubMed] [Google Scholar]
- Levine JM, Zentall TR. Effect of conspecific's presence on deprived rats performance: social facilitation vs. distraction/imitation. Animal Learning and Behavior. 1974;2:119–122. [Google Scholar]
- Lorenz K. Der kumpanin der umvelt des vogels: die artgenosse als ausloesendesmoment socialer verhaltenweisen. Journal fur Ornithologie. 1935;83:289–413. [Google Scholar]
- Mitchell CJ, Heyes CM, Dawson GR. Limitations of a bidirectional control procedure for the investigation of imitation in rats: odour cues on the manipulandum. Quarterly Journal of Experimental Psychology. 1999;52:193–202. [Google Scholar]
- Piaget J. Play, Dreams, and Imitation in Childhood. New York: W. W. Norton; 1962. [Google Scholar]
- Pongrácz P, Miklósi Á, Kubinyi E, Gurobi K, Topál J, Csányi V. Social learning in dogs: the effect of a human demonstrator on the performance of dogs, Canis familiaris, in a detour task. Animal Behaviour. 2001;62:1109–1117. [Google Scholar]
- Pongrácz P, Miklósi Á, Kubinyi E, Topál J, Csányi V. Interaction between individual experience and social learning in dogs. Animal Behaviour. 2003;65:595–603. [Google Scholar]
- Pongrácz P, Miklósi Á, Timár-Geng K, Csányi V. Preference for copying unambiguous demonstrations in dogs (Canis familiaris) Journal of Comparative Psychology. 2003;117:337–343. doi: 10.1037/0735-7036.117.3.337. [DOI] [PubMed] [Google Scholar]
- Range F, Viranyi Z, Huber L. Selective Imitation in Domestic Dogs. Current Biology. 2007;17:868–872. doi: 10.1016/j.cub.2007.04.026. [DOI] [PubMed] [Google Scholar]
- Thorpe WH. Learning and instinct in animals. 2nd. Cambridge, MA: Harvard University Press; 1963. [Google Scholar]
- Topál J, Byrne R, Miklósi Á, Csányi V. Reproducing human actions and action sequences: “do as I do!” in a dog. Animal Cognition. 2006;9:355–367. doi: 10.1007/s10071-006-0051-6. [DOI] [PubMed] [Google Scholar]
- Udell MA, Wynne CDL. A review of domestic dogs' (Canis familiaris) human-like behaviors: Or why behavior analysts should stop worrying and love their dogs. Journal of the Experimental Analysis of Behavior. 2008;89:247–261. doi: 10.1901/jeab.2008.89-247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whiten A, Ham R. On the nature and evolution of imitation in the animal kingdom: Reappraisal of a century of research. In: Slater PJB, Rosenblatt JS, Beer C, Milinski M, editors. Advances in the study of behaviour. Vol. 21. New York: Academic Press; 1992. pp. 239–283. [Google Scholar]
- Zajonc RB. Social facilitation. Science. 1965;149:269–274. doi: 10.1126/science.149.3681.269. [DOI] [PubMed] [Google Scholar]
- Zentall TR. An analysis of imitative learning in animals. In: Heyes CM, Galef BG, editors. Social learning in animals: The roots of culture. San Diego, CA: Academic Press; 1996. pp. 221–243. [Google Scholar]