

Abstract
Legionella pneumophila is an intracellular parasite of protozoa that differentiates late in infection into metabolically dormant cysts that are highly infectious. Regulation of this process is poorly understood. Here we report that the small DNA binding regulatory proteins integration host factor (IHF) and HU are reciprocally expressed over the developmental cycle, with HU expressed during exponential phase and IHF expressed postexponentially. To assess the role of these regulatory proteins in development, chromosomal deletions were constructed. Single (ihfA or ihfB) and double deletion (Δihf) IHF mutants failed to grow in Acanthamoeba castellanii unless complemented in trans when expressed temporally from the ihfA promoter but not under Ptac (isopropyl-β-d-thiogalactopyranoside). In contrast, IHF mutants were infectious for HeLa cells, though electron microscopic examination revealed defects in late-stage cyst morphogenesis (thickened cell wall, intracytoplasmic membranes, and inclusions of poly-β-hydroxybutyrate), and were depressed for the developmental marker MagA. Green fluorescent protein promoter fusion assays indicated that IHF and the stationary-phase sigma factor RpoS were required for full postexponential expression of magA. Finally, defects in cyst morphogenesis noted for Δihf mutants in HeLa cells correlated with a loss of both detergent resistance and hyperinfectivity compared with results for wild-type cysts. These studies establish IHF and HU as markers of developmental stages and show that IHF function is required for both differentiation and full virulence of L. pneumophila in natural amoebic hosts.
The gram-negative intracellular pathogen Legionella pneumophila is responsible for most cases of Legionnaires' disease, an atypical pneumonia that often develops following inhalation of Legionella-laden aerosols (17, 23). In natural environments, L. pneumophila has evolved as an intracellular parasite of protozoa, in which it progresses through stages of a distinct dimorphic developmental cycle, alternating between vegetative replicative forms and planktonic cyst forms, also referred to as MIFs (19, 26, 33). The characteristic features of the metabolically dormant cyst forms include the following: (i) thickened cell walls, (ii) laminations of intracytoplasmic membranes, and (iii) cytoplasmic inclusions of poly-β-hydroxybutyrate (PHBA). These forms are highly resistant to lysis by detergents and antibiotics and are hyperinfectious in cell-based infection models (14, 19, 26, 27). Cysts are not appreciably produced in U937 macrophages (26) or under in vitro conditions, since morphogenesis aborts in stationary phase. The developmental program is activated upon ingestion of vegetative bacteria by Tetrahymena species, a process by which the bacteria ablate digestion and in the absence of bacterial replication rapidly differentiate into cysts, which are excreted in fecal pellets (8, 20). The underlying regulatory cascades and environmental cues controlling this dimorphic life cycle are poorly understood.
A common feature of aquatic and soil microorganisms is their ability to form cysts or spores that promote survival during periods of desiccation, starvation, or other extreme conditions (52). In contrast to our knowledge of endospores, the regulatory control of cyst biogenesis has not received much attention. In Azotobacter vinelandii, cyst morphogenesis occurs postexponentially with synergistic synthesis of alginate and PHBA under the control of a regulatory cascade involving the stationary-phase sigma factor RpoS, the two-component regulatory system GacA/S, and the transcriptional regulator AlgR (13). Similarly, cyst formation by L. pneumophila in natural hosts occurs postexponentially and coordinately with the appearance of transmission phenotypes previously described for stationary-phase bacteria that include increased infectivity, motility, sodium sensitivity, osmotic resistance, and cytotoxicity (3, 12, 26, 61). Our studies have identified several developmentally regulated proteins, including MagA (MIF-associated gene of unknown function) and the alkyl hydroperoxide reductases AhpC1, AhpC2, and AphD, that serve as stage-specific markers (38, 41, 42). Genetic studies indicate that the postexponential transmission traits and MagA expression are partly controlled by a regulatory cascade involving, among other regulatory factors, RpoS and the two-component system LetA/S (an ortholog of GacA/S) (4, 5, 22, 25, 34, 45, 46, 47, 65). OxyR, a non-redox-active LysR variant, is induced in late exponential phase and directly represses ahpC2D (41). Tight regulation of developmental stages appears to be important, since rpoS and letA mutants show defects in both cyst morphogenesis and host range (20). Moreover, several studies have indicated that the major virulence system (Dot/Icm type IVB secretion system) is not developmentally regulated (20, 54).
Analysis of the promoter region of magA revealed a regulatory motif resembling the Escherichia coli consensus integration host factor (IHF) binding site (30). In the aquatic dimorphic organism Caulobacter crescentus, IHF is required for temporal activation of flagellar genes and promotes efficient chromosomal replication (29, 49, 50, 56). Likewise, IHF appears to be involved in transcription activation of developmentally regulated genes in Chlamydia trachomatis (63). IHF was originally identified and characterized for E. coli as a heterodimeric protein (α and β subunits, encoded by himA and himD, respectively) that recognizes a 13-bp core consensus DNA sequence which upon binding bends the DNA up to an angle of 180° (2, 15, 30, 32, 51). By bending the DNA, IHF acts as an accessory factor that influences regulatory processes such as replication and transcription (31). In addition, IHF has been shown to be involved in virulence gene expression in a wide range of bacterial pathogens (37, 43, 44, 56, 57), and loss-of-function single (himA) or double (himA himD) mutations in IHF subunits are often associated with a loss of virulence (43, 49, 58, 62).
Here we show that the IHF proteins of L. pneumophila are most abundant postexponentially and become concentrated in the transmissive cyst form. In contrast, HU, a heat-stable nucleoid protein paralogue of IHF (1, 16), is undetectable in the cyst and is one of the most abundant proteins, like AhpC2 and AhpD expressed by replicative forms. A loss of IHF function results in pleiotropic phenotypes, including depressed levels of MagA and of PHBA-containing inclusions and incomplete differentiation into the cyst form. Moreover, IHF function, while not required for infectivity of HeLa cells, is required for infection of its natural host, Acanthamoeba castellanii.
MATERIALS AND METHODS
Bacterial strains, oligonucleotides, media, and general methods.
Bacterial strains and oligonucleotides used in this study are listed in Table 1 and in Table S1 in the supplemental material, respectively. All L. pneumophila strains were grown in BYE broth or BCYE agar as described previously (21), which when required was supplemented with thymidine (100 μg/ml). E. coli DH5α and E. coli BL21(DE3) CodonPlusRIL, used as host strains for cloning strategies and recombinant protein expression, respectively, were grown in LB medium. Antibiotics (streptomycin [100 μg/ml], kanamycin [40 μg/ml], gentamicin [10 μg/ml], ampicillin [100 μg/ml], metronidazole [20 μg/ml], and chloramphenicol [4 or 20 μg/ml]) were added for selection where appropriate. All oligonucleotides were synthesized by Integrated DNA Technologies (Coralville, IA), Invitrogen (Frederick, MD), or Qiagen (Mississauga, Ontario, Canada). All restriction enzymes were purchased from New England Biolabs. DNA manipulations followed general protocols (53). All protein concentrations were determined using Bradford's protein assay reagent (Bio-Rad).
TABLE 1.
Bacterial strains and plasmids used in this study
| Strain | Description | Reference or source |
|---|---|---|
| L. pneumophila | ||
| SVir | Spontaneous Strr isolate of Philadelphia-1 strain | 39 |
| Lp02 | Strr, Thy−, HsdR− derivative of Philadelphia-1 strain | 7 |
| JV918 | Lp02 ΔdotB mutant strain | 55 |
| Lp02ihfA | Lp02 Strr, Thy−, HsdR−ihfA::Genr | This study |
| Lp02ihfB | Lp02 Strr, Thy−, HsdR−ihfA::Kanr | This study |
| Lp02Δihf | Lp02 Strr, Thy−, HsdR−ihfA::genihfB::kan | This study |
| Lp02ΔrpoS | Lp02 Strr, Thy−, HsdR−rpoS::kan | 3 |
| MGM001 | pMMB206 into Lp02 | This study |
| MGM002 | pMGM007 into Lp02 | This study |
| MGM003 | pMMB206 into Lp02Δihf | This study |
| MGM004 | pMGM007 into Lp02Δihf | This study |
| KB120 | pBH6119 into Lp02; Strr, Thy+ | This study |
| KB123 | pBH6119 into Lp02Δihf; Strr Kanr Genr Thy+ | This study |
| KB130 | pKB127 into Lp02; Strr, Thy+ | This study |
| KB131 | pKB127 into Lp02ΔrpoS; Strr Kanr, Thy+ | This study |
| KB135 | pBH6119 into Lp02 rpoS; Strr Kanr, Thy+ | This study |
| KB151 | pKB127 into Lp02Δihf; Strr Kanr Genr, Thy+ | This study |
| E. coli | ||
| DH5α | F′ endA1 hsdR17 (rk− mk+) supE44 thi-1 recA1 gyrA (Nalr) relA1 Δ(lacIZYA-argF)U169 deoR (φ80dlacΔ(lacZ)M15) | New England Biolabs |
| BL21(DE3) CodonPlus RIL | B F−ompT hsdS(rB− mB−) dcm+ Tetrgal λ(DE3) endA Hte [argU ileY leuW Camr] | Stratagene |
| Plasmids | ||
| pET16b | N-terminal 10-histidine-tagged fusion protein expression vector; Ampr | Novagen |
| pBluescriptII KS(+) | Cloning vector; Ampr | Stratagene |
| pUC18 | Cloning vector; Ampr | Invitrogen |
| pMMB206 | Derivative of pRSF1010; Ptac promoter and IPTG-inducible lacIQ system; Ampr Camr, Thy+ | Michele Swanson |
| pBH6119 | Promoterless GFP vector; Ampr, Thy+ | 35 |
| pKB127 | pBH6119::magA 245-bp promoter region cloned into BamHI and XbaI sites | This study |
| pRDX | Dual suicide vector pBOC20 with Bacillus subtilis sacB and Helicobacter pylorirdxA (nitroreducase) as counterselectable markers; Camr | This study |
| pKB58 | pET16b::himA (IHFα) into NdeI and XhoI sites; Ampr | This study |
| pKB59 | pET16b::himD (IHFβ) into NdeI and XhoI sites; Ampr | This study |
| pKB129 | pET16b::hupB (HU) into NdeI and XhoI sites; Ampr | This study |
| pMGM001 | pBS ihfA | This study |
| pMGM002 | pBS ihfA::Genr | This study |
| pMGM003 | pRDX ihfA::Genr | This study |
| pMGM004 | pUC18 ihfB | This study |
| pMGM005 | pUC18 ihfB::Kmr | This study |
| pMGM006 | pRDX ihfB::Kmr | This study |
| pMGM007 | pMMB206::ihfA+ihfB+ | This study |
Protein expression and purification of IHFα and IHFβ subunits and HU.
L. pneumophila orthologs of the E. coli IHFα (himA) and -β (himD) subunit genes and the HU gene (hupB) were found via bioinformatic analyses of the L. pneumophila Philadelphia-1 genome sequence (http://genolist.pasteur.fr/LegioList/) and henceforth are referred to as ihfA (Lpg2709), ihfB (Lpg2955), and hupB (Lpg1858), respectively. L. pneumophila ihfA (primers IHFαNdeIF and IHFαXhoIR), ihfB (primers IHFβNdeIF and IHFβXhoIR), hupB (primers HUNdeIF and HUXhoIR), and rpoS (primers rpoSNdeIF and rpoSXhoIR) were PCR amplified, directionally cloned into NdeI and XhoI restriction sites to produce His6-tagged fusion proteins in the expression vector pET16b, and overexpressed in E. coli BL21(DE3) CodonPlus RIL (Stratagene) by using 1 mM (final concentration) isopropyl-β-d-thiogalactopyranoside (IPTG) induction at an optical density at 660 nm (OD660) of ∼0.500 in 1 liter of Luria-Bertani medium for 60 min. Extraction and Ni interaction purification were performed as described previously (41). In order to attain the IHFαβ heterodimer, IHFα and IHFβ eluted protein preparations were mixed at a 1:1 ratio before dialysis to facilitate formation of heterodimers and verified by gel filtration (64).
Gel retardation assays.
The binding reaction mix contained 1.5 mg/ml sheared fish sperm DNA (Sigma), 1 mg/ml bovine serum albumin (New England BioLabs), 5% glycerol, 40 mM Tris-HCl (pH 7.5), 30 mM MgCl2, 0.1 mM EDTA, 0.1 mM dithiothreitol, 0.1 M NaCl, and ∼10,000 cpm of radiolabeled 245-bp PmagA promoter region DNA generated by PCR using the primers magABamHIF and magAXbaIR. The radioisotope [α-32P]dCTP (3,000 Ci/mmol) was obtained from Amersham BioSciences (Baie D'Urfé, Quebec, Canada) and used in PCR-based labeling as described previously (41). The purified recombinant proteins IHFαβ, E. coli IHFαβ (IHFαβEc), and RpoS were added over a range of 0 to 14 μM. The total volume of the reaction mix was then adjusted to 20 μl with sterile distilled water. Protein was omitted from the control tube in order to determine the electrophoretic mobility of free probe DNA. All of the reaction mixtures were incubated for 30 min at room temperature, and 2 μl of loading buffer (0.3% glycerol and 0.01% bromophenol blue in 10× Tris-borate buffer) was added to each of the reaction mixtures, which were run in prerun (1 h at 20 mA) 6% 0.5× Tris-borate-EDTA nondenaturing polyacrylamide gel electrophoresis at 20 mA, dried, and exposed to Kodak BioMax MR autoradiographic film. Consistent results were obtained from three replicates.
Construction of L. pneumophila Lp02Δihf mutant strain.
IHF chromosomal deletions were made by utilizing an in-house gene replacement suicide vector (pRDX) strategy (41). Briefly, ∼500-bp sequences flanking ihfA were amplified by PCR (primer pairs IHFαP1 and IHFαP2 and IHFαP3 and IHFαP4) and ligated into pBluescript (pBS), creating pBS ihfA (pMGM001), after which the gentamicin cassette was inserted into the BamHI site within the insert, creating pBS ihfA::Genr (pMGM002). The ihfA::Genr construct was then excised from pMGM002 and ligated into the suicide vector pRDX, creating pRDX ihfA::Genr (pMGM003). Likewise, the same strategy was employed to create the pRDX ihfB::Kanr (pMGM006) construct except that the cloning vector pUC18 (pMGM004) and a kanamycin resistance cassette (pMGM005) were utilized (see Table S1 in the supplemental material for primers). Approximately 10 μg of pMGM006 was electroporated into electrocompetent L. pneumophila Lp02, plated on BCYE medium supplemented with streptomycin, kanamycin, and thymidine, and incubated for 3 to 4 days. The resulting transformants were replica plated and screened for loss of metronidazole sensitivity on BCYE supplemented with 20 μg/ml of metronidazole (loss of the plasmid vector). The selected Lp02ihfB (Kmr) and Lp02ihfA (Genr) chromosomal deletion strains were confirmed by PCR amplification for the absence of ihfB and ihfA, respectively. To create the Lp02 ihfA::Genr ihfB::Kanr (henceforth referred to as Lp02Δihf) chromosomal deletion double mutant strain, approximately 10 μg of pMGM003 was electroporated into Lp02ihfB cells and plated on BCYE supplemented with streptomycin, gentamicin, kanamycin, and thymidine. After replica plating and screening for metronidazole resistance, the Lp02Δihf chromosomal double-deletion mutant strain was confirmed for the absence of ihfA and ihfB by PCR amplification and by immunoblotting with rabbit polyclonal anti-E. coli IHF (IHFEc) (supplied by S. D. Goodman).
IHF complementation in Lp02Δihf.
Complementing constructs were created by PCR, with ihfB positioned behind ihfA (containing its own Shine-Delgarno sequence) and under the control of both the ihfA and vector Ptac promoters (including a Shine-Delgarno sequence upstream of ihfA) in pMMB206. The resulting plasmid, pMMB206::ihfA+ ihfB+ (pMGM007), was electroporated into Lp02Δihf to create the complemented strain MGM004, and expression of both IHF subunits was confirmed by reverse transcription-PCR and immunoblotting with rabbit polyclonal anti-IHFEc. The overexpression strain MGM002 was created by electroporating the plasmid pMGM007 into Lp02. For negative controls, pMMB206 was electroporated into Lp02 and Lp02Δihf to create MGM001 and MGM003, respectively.
Immunoblots.
Approximately 1 μg each of purified L. pneumophila IHFα, IHFβ, and HU proteins was loaded onto a 4-to-12%-gradient Bis-Tris NuPage gel (Invitrogen). L. pneumophila cells were harvested from 48-h BCYE plate growth, normalized to an OD620 of 0.20, pelleted, and resuspended in 50 μl of 50 mM Tris-Cl (pH 7.5). L. pneumophila SVir cyst forms were isolated from infected HeLa cells as described elsewhere (26). Pelleted cysts with an OD620 of 1.0 (1 ml) were resuspended in 5 ml of warmed BYE broth (no antibiotics) and incubated in a 37°C shaker. At various time points, cell samples at an OD620 of 0.20 were taken, pelleted, and resuspended in 50 μl of 50 mM Tris-Cl (pH 7.5). Samples were normalized for protein loading and run on a 10% sodium dodecyl sulfate-polyacrylamide gel or on a precast 4 to 12% NuPAGE Bis-Tris gel (Invitrogen), transferred to nitrocellulose (BioLynx) as per the laboratory protocol, and immunoblotted with anti-IHFEc antibody that was prepared as described elsewhere (30) at a 1:1,000 dilution.
Green fluorescent protein (GFP) promoter fusion assays.
The magA 245-bp promoter region (PmagAgfp) was PCR amplified and ligated into the appropriate restriction sites in pBH6119 (35), creating pKB127. The control plasmid pBH6119 and the construct pKB127 were then electroporated into electrocompetent Lp02, Lp02Δihf, and Lp02ΔrpoS cells. Strains KB120, KB123, KB130, KB131, KB135, and KB151 were streaked on BCYE plates (supplemented with appropriate antibiotics) from frozen stock and incubated at 37°C for 48 h. Bacteria were suspended in BYE broth and normalized to a final OD620 of 0.01 to initiate growth. For fluorescence analysis at each time point, aliquots of cells were washed twice with 1× phosphate-buffered saline (PBS), resuspended in 1× PBS, and normalized to an OD620 of 0.1, and fluorescence was measured in 2-ml samples in a VeraFluor fluorometer (Bio-Rad) at excitation and emission wavelengths of 488 nm and 510 nm, respectively. Means and standard deviations were determined from triplicate assays, and data were analyzed by using a t test.
Acanthamoeba castellani infection.
A. castellanii (ATCC 30010) was maintained in ATCC medium 354 at 25°C. For the infection experiments, a 48-h culture of A. castellanii was washed and resuspended in Tris-buffered salt solution containing 2 mM KCl, 1 mM CaCl2, 0.5 mM MgCl2, and 1 mM Tris, with the pH adjusted to 6.8 to 7.2 (9). For infections with Lp02 strains, 100 μg/ml of thymidine was added to the medium. Approximately 105 cells per ml of A. castellanii were infected with ∼103 CFU/ml of L. pneumophila. All experiments were performed in triplicate at 25°C, and at each time point (0, 24, 48, and 72 h postinfection), amoeba cells were disrupted by passage through a 27-gauge needle. Intracellular bacteria were enumerated following decimal dilution and plating on BCYE agar plates, and CFU per ml are reported as the means and standard deviations of triplicates.
HeLa cell infection.
HeLa cells were grown in minimal essential medium (MEM) as described previously (28). HeLa cells were harvested, counted in a Neubauer hemocytometer, and resuspended in MEM without antibiotics to a concentration of 106 cells/ml. Cells from spinner bottles were dispensed into 12-well plates at 106 cells per well (1 ml) and left to adhere and spread for 1 to 2 h. Plate-grown L. pneumophila was harvested in double-distilled water, and the bacterial suspension was standardized to an OD600 of 1.0. One hundred microliters of the standardized bacterial suspensions were added to triplicate wells to a final inoculum of 108 bacteria/106 HeLa cells (28). Plates were centrifuged at 500 × g for 10 min at room temperature to maximize contact of bacteria with the HeLa cell monolayer and then incubated for 3 h to facilitate infection. For attachment-and-invasion assays, plates were washed six times with PBS, the monolayers were lysed, and dilutions were plated for bacterial enumeration. For invasion assays, monolayers were washed three times with PBS and then treated for 1 h with MEM containing 100 μg/ml gentamicin and similarly enumerated for viable bacteria. For studies with the Lp02 strain, MEM was supplemented with thymidine and 2 mM IPTG. For cyst purification, monolayers were harvested using a cell scraper, resuspended in ddH2O, and passaged through a 26-gauge needle to lyse HeLa cells as described previously (26).
Cell cycle experiments.
Cyst germination studies were performed with BYE broth in which freshly isolated Lp02 cysts were suspended to an OD660 of 0.1 and allowed to grow over a period of 48 h. Samples were taken at early exponential phase (OD660 = 0.2 to 0.4), mid-exponential phase (OD660 = 0.6 to 0.8), late exponential phase (OD660 = 0.9 to 1.1), and post-exponential phase (OD660 = 1.2 to 1.5). Samples were normalized to an OD660 of 1.0 following suspension in sodium dodecyl sulfate sample buffer and subjected to electrophoresis and immunoblotting to track IHF and HU protein production as previously outlined. All experiments were performed in triplicate to ensure reproducibility.
Transmission electron microscopy (TEM).
At 24, 48, and 72 h postinfection, light microscopic images of Legionella-infected HeLa cells were captured at magnification ×40 using an Olympus IX-71 (Olympus) inverted microscope equipped with a 12-bit Evolution QET Monochrome camera (Media Cybernetics) to confirm the stage of intracellular growth. Thin sections of infected HeLa cells were prepared from specimens fixed in glutaraldehyde and embedded in epoxy resin as reported previously (19). Multiple sections from each sample were photographed, and of 20 to 50 cells examined, one typical photograph was selected to represent the group. To ensure conformity, multiple replicates on different days were examined. Statistical analysis was generally unnecessary since photographs represent 90% or more of forms observed for each sample.
RESULTS
Analysis of upstream sequences of the developmentally regulated magA gene revealed a putative integration host factor binding consensus sequence (TATCCAAAAATAA), suggesting that IHF might participate in postexponential regulation. Potential IHF binding motifs are also found in upstream sequences of ∼200 L. pneumophila genes (based on BLAST searches of the L. pneumophila genome on the Legiolist web server), which includes several regulators of postexponentially expressed genes (letA, letE, fleQ, rpoS, and ihfA). Since IHF proteins are highly conserved, we found that a polyclonal IHFEc antibody cross-reacted with purified IHFα and IHFβ subunits and with a paralogous protein, HU (see Fig. 1A). L. pneumophila HU shares 78% identity and 84% similarity with E. coli HU, as well as ∼37% identity and 57% similarity to IHFα and IHFβ of L. pneumophila (data not shown). The IHF immunoblot of cellular extracts of L. pneumophila Lp02 cells revealed two bands close together in the 10- to 12-kDa range: the upper band corresponded to IHF subunits based on the absence of these bands in the Lp02Δihf mutant strain, and the lower band migrated similarly to purified HU (Fig. 1B). For verification purposes, L. pneumophila SVir, harvested at late and post-exponential phases, were compared with Lp02 and Lp02Δihf mutant strains to confirm these identities. Note that the IHFα and IHFβ subunits are not distinguishable in Fig. 1B and C due to a lower resolving gel percentage than that viewed in Fig. 1A. Similar allelic replacement strategies used to knock out the ihfA and ihfB genes were used to knock out hupB, but these proved unsuccessful and were not further pursued (data not presented).
FIG. 1.

Polyclonal IHFEc immunoblots. (A) Polyclonal IHFαβEc antibody detects purified histidine-tagged IHFα and IHFβ proteins and cross-reacts with histidine-tagged HU protein. Molecular masses of the proteins (including the His10 tag) are indicated. (B) Determination of IHF and HU bands in L. pneumophila SVir time point samples (LE and PE are identical to those samples used in panel C) and in strains Lp02 and Lp02Δihf. (C) IHF production varies throughout the L. pneumophila SVir cell cycle (bottom panel), and Ponceau Red staining of the immunoblot (top panel) indicates equal protein loading of all time point samples. Note that the LE and PE time point samples are identical to the samples used in panel B. Also note that the gel percentage is higher in panel A than in panels B and C, and hence there is no discrimination between the IHFα and IHFβ subunits in L. pneumophila cell samples in panels B and C. EE, early exponential phase; ME, mid-exponential phase; LE, late exponential phase; PE, post-exponential phase. Details of the phases of growth are presented in the text. Experiments depicted are typical of at least three replicates for two independent experiments.
To track IHF and HU protein levels during the in vitro growth cycle, bacterial growth was synchronized by utilizing purified cysts harvested from 48-h-infected HeLa cells. As seen in Fig. 1C, cysts are enriched for IHF proteins, whereas the HU protein is undetectable. Following germination of cysts into vegetative bacteria (∼18 h) in BYE broth and proceeding through early-, mid-, and late-exponential phases (growth curve), IHF protein levels were found to decrease coordinately with increasing levels of HU. During late log phase, IHF levels were visible and increased to near cyst levels during stationary phase while HU levels decreased. The trajectory of IHF production followed those of other developmentally regulated protein markers, MagA and AhpC1 (data not presented).
Complementation of Lp02Δihf.
To restore IHF production in Lp02Δihf, the coding sequences for ihfA+ and ihfB+ under the control of both the ihfA promoter and the Ptac promoter of pMMB206 and both subunits were verified by PCR (Fig. 2A) and by immunoblotting for MGM004 (Fig. 2B).
FIG. 2.
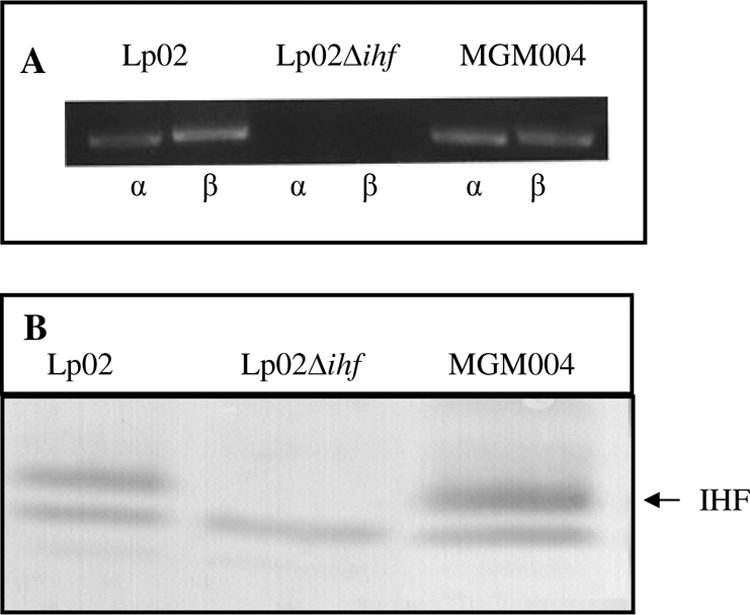
Deletion, complementation, and expression of IHF. (A) Presence or absence of ihfA and ihfB subunit genes as determined by PCR amplification. (B) Expression of IHF determined by polyclonal IHFEc immunoblotting of in vitro growth (postexponential) of Lp02 (lane 1), Lp02Δihf (lane 2), or MGM004 (pMGM007; ihfA+ ihfB+) complement (lane 3).
In vitro and in vivo growth.
Since IHF may be involved in controlling the transcription of many genes, some of which might have growth-enfeebling effects, we next examined the in vitro growth characteristics of the Lp02Δihf strain. As is shown in Fig. 3A, the growth rates of Lp02 and Lp02Δihf were indistinguishable, indicating that the Lp02Δihf mutant was not defective for in vitro growth. To assess the role of IHF in the intracellular multiplication of L. pneumophila, infectivity studies with in vitro-grown bacteria were performed with Acanthamoeba and HeLa cells. As shown in Fig. 3B, single- and double-knockout mutants of L. pneumophila failed to grow in A. castellani, with both Lp02Δihf and Lp02ihfA mutants being cleared while Lp02ihfB mutants persisted but failed to grow. Complementation of the Lp02Δihf mutant with pMGM007 (the MGM004 strain) restored infectivity and intracellular growth, but only from the PihfA (endogenous) promoter. Constitutive expression from pMGM007 (IPTG) failed to restore infectivity (Fig. 3B), suggesting that inappropriate expression in early exponential phase likely results in dysregulation of developmental systems or factors required for infectivity of amoebae.
FIG. 3.

Growth curves. (A) In vitro growth in BYE broth of Lp02 (▪) or Lp02Δihf (•). (B) Intracellular growth in A. castellanii of Lp02 (⧫), Lp02ihfA (▴), Lp02ihfB (▵), MGM004 (Lp02Δihf/pMGM007 complement)(▪), Lp02Δihf (○), or MGM004 plus IPTG (□). (C) Intracellular growth in HeLa cells of Lp02 (▪), MGM002 (Lp02/pMGM007) with 2 mM IPTG (⧫), MGM001 (Lp02/pMMB206 vector control) with 2 mM IPTG (□), Lp02Δihf (•), or MGM002 (Lp02/pMGM007) (○). All experiments were performed in triplicate, and mean and standard deviation are depicted.
In contrast to the results with natural hosts, all of the mutants displayed comparable growth rates in HeLa cells (Fig. 3C). Microscopic examination revealed that there were extracellular bacteria present in the culture medium after 24 h, indicating no obvious defect in escape from the host cell (data not shown). To determine if constitutive overexpression of complementing IHF from pMGM007 (the MGM004 strain) also resulted in intracellular growth defects, infections were carried out in the presence of IPTG. Overexpression of both IHF subunits in the MGM004 strain resulted in no measurable differences from results for the Lp02 and MGM002 strains. In addition, we found no appreciable differences between Lp02Δihf and Lp02 in efficiency of attachment or invasiveness for HeLa cells (data not shown). Thus, IHF mutants show infectivity defects only in natural hosts, a phenotype also reported with rpoS mutants (34).
MagA expression.
We have previously reported that MagA is a marker for postexponential development in L. pneumophila (38). To confirm that IHF function is associated with the expression of magA, PmagAgfp reporter constructs were introduced into Lp02Δihf and into an Lp02ΔrpoS mutant strain, and fluorescence intensity was tracked over the in vitro growth cycle. Figure 4A shows the upstream region of magA, including the putative IHF binding motif and inferred RpoS promoter sequences. Figure 4B shows the relative fluorescence levels for Lp02 PmagA::GFP (KB130), Lp02Δihf PmagA::GFP (KB151), and Lp02ΔrpoS PmagA::GFP (KB131), along with the vector controls Lp02 vector GFP (KB120), Lp02Δihf vector GFP (KB123), and Lp02ΔrpoS vector GFP (KB135) measured at mid-, late-, and post-exponential growth phases. No significant differences were observed in the GFP expression levels in Lp02, Lp02Δihf, and Lp02ΔrpoS at mid-exponential growth phase (Fig. 4B). However, statistically significant differences in GFP expression levels (P = 0.001 by t test) became evident throughout the late- and post-exponential growth phases in that the GFP expression levels in the Lp02Δihf and Lp02ΔrpoS strains were similar to one another and both were below the GFP expression level observed for Lp02. Thus, the observed decreased fluorescence suggested that RpoS and IHF contribute to the post-exponential expression of this developmental marker.
FIG. 4.

Expression of magA-gfp fusions in mutants of L. pneumophila. (A) Promoter sequence of magA, depicting the −10 and −35 hexamers and the putative IHF and RpoS binding sites (bold). (B) MagA promoter GFP fusions were monitored at different points of in vitro growth (ME, midexponential; LE, late exponential; PE, postexponential). GFP vector control, Lp02/pBH6119::PmagAgfp, Lp02Δihf/pBH6119::PmagAgfp, and Lp02ΔrpoS/pBH6119::PmagAgfp) are analyzed. For simplicity, a single GFP vector control is depicted, which is representative of controls for Lp02, Lp02Δihf, and Lp02ΔrpoS. (C) Gel retardation analysis of the 245-bp PmagA promoter region with IHFαβ, IHFαβEc, and RpoS. The radiolabeled 245-bp PmagA amplicon was incubated with increasing concentrations of IHFα (2.95, 8.95, and 14.9 μM), IHFβ (2.8, 8.5, and 14.2 μM), IHFαβ (1.5, 4.6, and 7.65 μM), the IHFαβEc protein (1.7, 5.1, and 8.5 μM), or RpoS (7.4, 22.1, and 36.8 μM). The control lane (0) does not include the IHF or RpoS protein in the binding reaction mix. Arrows denote band shifts.
EMSA.
To confirm that the IHF and RpoS proteins indeed bound the magA promoter region (direct regulatory role), we performed DNA binding electrophoretic mobility shift assay (EMSA) studies. As shown in Fig. 4C, only the IHFαβ and IHFαβEc heterodimers produced a gel shift, and in a concentration-dependent manner. Since mixing of the IHFα and IHFβ subunits produces heterodimers, we cannot rule out the possibility that some of the remaining monomers might form dimers through interaction with the IHF DNA motif. Similar studies with RpoS provided some evidence for binding of RpoS to magA promoter sequences. Since sigma factors generally bind poorly to DNA in the absence of RNA polymerase, our findings are only suggestive of an interaction. These studies support a direct role for IHF and perhaps RpoS in the postexponential regulation of magA gene expression.
Lp02Δihf mutants display altered morphology in HeLa cells.
As seen in Fig. 5, HeLa cell vacuoles containing Lp02 (panel A) and those containing Lp02Δihf (panel B) were similar at 48 h postinfection, as described previously (28). In contrast to Lp02, Lp02Δihf was conspicuous for an absence of PHBA-containing inclusions (Fig. 5B), suggesting that postexponential production of PHBA may be dependent on IHF function.
FIG. 5.

Electron microscopic examination of altered vacuoles. Transmission electron microscopy of HeLa cells 48 h postinfection with Lp02 (A), Lp02Δihf (B), complemented MGM004 (Lp02Δihf/pMGM007) strain (C), mock-complemented MGM003 (lp02Δihf/pMMB206) strain (D), overexpression MGM002 (Lp02/pMGM007) strain (E), or mock-overexpression MGM001 (Lp02/pMMB206) strain (F). Scale bar = 2 μm for all images. Images are representative of some 25 fields examined.
Attempts to restore PHBA inclusions through IHF complementation yielded mixed results, as has been reported for letA and rpoS complements (20). As seen in Fig. 5C and D, respectively, neither the complemented mutant strain MGM004 (IPTG induced) nor the mock-complemented mutant MGM003 contained PHBA inclusions. TEM examination of HeLa cells infected with the IPTG-induced strain MGM002 (Lp02 constitutively expressing ihfA+ and infB+) also revealed an absence of inclusions (Fig. 5E), while the mock-infected and IPTG-induced strain MGM001 was essentially wild type for PHBA-containing inclusions. Thus, it would appear that either the lack of IHF or its constitutive overexpression severely affects PHBA production within the infected HeLa cells.
Lp02Δihf mutant strain is defective in cyst morphogenesis.
The observed defects in morphogenesis, as well as the apparent decrease in PHBA inclusions, led us to more closely examine the maturation process of Lp02Δihf to determine its ability to form cysts. The intracellular maturation process of L. pneumophila in HeLa cells and the characteristic features of cysts (i.e., multilayered, tightly associated internal membranes, cytoplasmic inclusions, and irregular shapes) have been described previously (19, 20, 26). For confirmatory purposes, the Lp02 strain (Fig. 6A) used in this study was determined to undergo differentiation similar to that described for the SVir strain (Fig. 6B). Ultrastructural analysis of Lp02Δihf within HeLa cells (panel C) revealed defects in the late stages of differentiation; Lp02Δihf typically did not possess the same level of internal membrane layering as was seen with the wild-type strains (Fig. 6A and B) but instead possessed more loosely associated bilayers and fewer cytoplasmic inclusions and exhibited a more typical gram-negative envelope organization than wild-type strains (Fig. 6A and B). These defects were only partly restored by trans complementation using strain MGM004 (Fig. 6D). An examination of five randomly selected fields revealed only 6 to 10% of bacteria contained inclusions or showed evidence of cyst maturation. The mock-complemented MGM003 strain was similar in ultrastructure to Lp02Δihf (data not shown). Ultrastructural analysis of the overexpressing MGM002 strain within HeLa cells also showed defects in maturation, particularly in the formation of cytoplasmic inclusions (Fig. 6E). Of the five randomly selected fields containing replicative vacuoles of the MGM002-infected HeLa cells, only 26% of the bacterial population contained any cytoplasmic inclusions whereas 6% contained inclusions comparable to those observed with wild-type strains. In comparison to HeLa cells infected with the mock overexpression strain MGM001, nearly 100% of the bacterial population within the replicative vacuoles contained cytoplasmic inclusions (Fig. 6F). Thus, the absence of or constitutive overexpression of IHF affects not only PHBA production but also the ultrastructure of the cyst cell wall.
FIG. 6.

MIF morphogenesis phenotypes. Transmission electron microscopy of HeLa cells 48 h postinfection with Lp02 (A), Svir (B), Lp02Δihf (C), complemented MGM004 (Lp02Δihf/pMGM007) (D) strain, overexpression MGM002 (Lp02/pMGM007) strain (E), or mock-overexpression MGM001 (Lp02/pMMB206) strain (F). Scale bar = 100 nm for all images. The images are representative of 30 fields examined for each strain.
Resilience properties of defective Lp02Δihf cysts.
We previously demonstrated that cysts from Lp02 and Svir strains are highly resistant to the effects of detergents and antibiotics, and they were hyperinfectious in cell-based assays (26). Using these methods, we compared in vitro stationary-phase grown bacteria and the cyst forms harvested from HeLa cells. As seen in Table 2, there were no differences in detergent tolerance or in infectivity between the Lp02Δihf strain and Lp02 in stationary growth phase. However, the cyst forms of Lp02Δihf were much more susceptible to lysis by detergents than were Lp02 cysts. Moreover, reinfection efficiencies for the Lp02Δihf cysts were 100-fold lower than with Lp02 cysts. These findings are consistent with differences in cell wall morphology noted between these cyst forms by TEM.
TABLE 2.
Resilience properties of cyst formsa
| Bacterial strain | Cell type | Time (min) to 50% turbidity with detergent
|
Reinfection efficiency (%) | |
|---|---|---|---|---|
| Sarkosyl | Triton X-100 | |||
| Lp02 | SP | 90 | 1,170 | |
| Cyst | ∞ | ∞ | 0.256 | |
| Lp02Δihf | SP | 90 | 1,170 | |
| Cyst | 4,440 | 4,020 | 0.216 × 10−2 | |
Cyst-like forms were harvested at 72 h postinfection from HeLa cells and compared with in vitro growth stationary-phase bacteria (SP).
DISCUSSION
Developmental cycles have been described for aquatic and soil bacteria originating from a variety of prokaryotic genera with the common element of formation of cyst-like progeny tailored for survival in nutrient-poor environments (48, 52). Accordingly, the transition from vegetative growth to terminally differentiated cysts (and germination) must be tightly controlled through developmental programs composed of stage-specific regulatory elements. Here we show that IHF and HU are reciprocally expressed across the developmental cycle of L. pneumophila, following trajectories similar to those previously reported for other developmentally regulated proteins, including the alkyl hydroperoxide reductases AhpC2D and AhpC1 (42), MagA (38), and the developmental regulator OxyR (41). The nucleoid protein HU and AhpC2 and AhpD are most abundant during exponential phase, whereas IHF, AhpC1, and MagA are most abundant postexponentially and become concentrated in the developing cyst. The LysR regulator OxyR is expressed postexponentially and specifically represses the ahpC2D operon and possibly other genes associated with differentiation (41). As with LetA and RpoS, IHF function is required for infection of natural amoebic hosts but dispensable for infection of mammalian cells, and both are required for full expression of the developmentally regulated gene magA. EMSA studies confirmed that both IHF and to a lesser extent RpoS directly bind to the promoter region of magA.
While the role of HU appears to be essential for cell division, IHF is not essential, possibly being compensated for by other small DNA binding proteins, such as Fis (H-NS is absent in Legionella spp.). However, the absence of IHF function correlates with several postexponential phenotypes, including altered terminal vacuole morphology in HeLa cells and an absence of typical cyst morphology (e.g., absence of a thick cell wall, laminations of intracytoplasmic membranes, and PHBA inclusions). The latter phenotypes also correlate with increased susceptibility to detergents and a 100-fold decrease in infectiousness compared with results for cysts from wild-type strains (26). These phenotypic differences have also been noted with letA and rpoS mutant strains in HeLa cells (20), suggesting a requirement for several independent signaling pathways in order to activate the developmental program. Allelic complementation of the IHF mutant from the ihfA endogenous promoter partially restored infectivity for A. castellani, but induction with IPTG (Ptac promoter) and constitutive expression resulted in a loss of infectivity. In the HeLa cell infection model, constitutive overexpression of IHF proteins did not reverse phenotypic changes noted for the Lp02Δihf strain and caused mutant phenotypic changes to strain Lp02, including the absence of PHBA-containing inclusions. Thus, timing of IHF production appears to be crucial for the proper initiation of the postexponential developmental program. Our studies are consistent with the general view of postexponential regulatory cascades (3, 4, 12, 34, 35, 36, 41, 60, 65), in which upregulation of IHFαβ postexponentially contributes broadly to regulation of as many as 200 genes, based on consensus IHF binding sequences within 300 bp upstream of the translational start sites. Accordingly, either overexpression or inappropriate expression of IHF appears to disrupt the developmental program, affecting cyst morphogenesis as well as infectivity for natural hosts.
In E. coli, IHFEc is recognized as a global regulatory factor associated with the transcription of many genes (2, 40). As in E. coli (2, 18), expression of IHF and HU in L. pneumophila is growth phase dependent, with maximal protein levels in postexponential and exponential growth phases, respectively. However, the basis for developmental regulation of IHF in L. pneumophila remains to be determined. Synthesis of IHF has been shown to be upregulated in other species, such as C. crescentus (29), Shigella flexneri (49), Neisseria gonorrhoeae (24), and C. trachomatis (63) upon entry into exponential phase. More importantly, the C. trachomatis IHF protein is also developmentally expressed and controls the expression of late genes associated with morphogenesis of the resilient infectious elementary bodies (63). The Lp02Δihf strain showed no obvious growth defects in vitro or in vivo in HeLa cells. However, IHF function seems to be more important late in intracellular infection as vegetative bacteria differentiate into resilient cysts. Electron micrographs of Lp02Δihf show early stages of cyst morphogenesis, including some intracytoplasmic membranes and cell wall modifications that correlate with some postexponential phenotypes (sodium sensitivity). The paucity of PHBA inclusions in the Lp02Δihf mutant suggests that IHF must regulate some step in the biosynthetic pathway, since none of the PHBA synthase genes contains obvious consensus IHF binding sites. Further study is required to identify additional genes or regulatory factors associated with late stages of cyst morphogenesis which are dependent on IHF function.
The role of HU in controlling gene expression in L. pneumophila is not as apparent as for IHF. In enteric bacteria, HU is a heterodimer composed of the hupA- and hupB-encoded subunits (6, 10). In L. pneumophila, HU is a homodimer and the product of a single gene, hupB. HU, a structural homologue of IHF (1, 2), does not exhibit DNA binding sequence specificity (10, 15) and preferentially binds distorted DNA (i.e., single-stranded gaps, cruciforms, kinks, bends, etc.) to provide architectural assistance in various DNA transactions (59). Recently it has been shown for E. coli that the efficiency of stationary-phase sigma factor rpoS translation is reduced in cells lacking HU; thus, rpoS transcription or RpoS stability is affected by HU (6). Furthermore, HU specifically bound an RNA fragment containing the translational initiation region of rpoS mRNA, strongly suggesting that HU regulates rpoS translation (6). In L. pneumophila, rpoS transcripts appear mainly during exponential phase, when HU levels are high, and are translated postexponentially as HU levels begin to diminish (5). While control of RpoS would not be considered an essential function (since RpoS is not essential), we speculate that HU essentiality may reflect functions unique to the exponential growth stage, much as LpOxyR is essential for postexponential transitions (41).
The postexponential role of RpoS in controlling stationary-phase events is well established for L. pneumophila (3, 34). As shown in this study, RpoS and IHF participate in regulating expression of the developmental stage marker MagA, either independently of each or in a cooperative manner. However, we cannot rule out the possibility of epistatic effects that might be caused by IHF modulation of rpoS expression (the rpoS promoter region contains an IHF consensus binding motif, as do those of the ihfA and ihfB genes). Clearly there are additional levels of regulation, since cyst morphogenesis is aborted in stationary phase in vitro and these bacteria express both IHF and RpoS (20, 26). In contrast, our studies indicate that IHF function is necessary for cyst maturation and perhaps activation of genes associated with PHBA production. The enrichment of IHF in mature cysts may also play an important role in germination, perhaps through selective activation of early genes whose function is required for subversion of host cells in order to produce a replication permissive endosome. In summary, HU and IHF are reciprocally expressed across the developmental cycle of L. pneumophila and appear to be uniquely adapted to the control of growth-stage-specific genes.
Supplementary Material
Acknowledgments
We are grateful to Michele Swanson and Joseph Vogel for their kind gifts of the promoterless GFP vectors and the Lp02ΔrpoS and ΔdotB mutant strains. We also thank Audrey Chong, Jennifer Chase, and Priscilla Frenette for technical assistance and Elizabeth Garduño for providing isolated SVir cysts.
This work was funded by a Canadian Institutes of Health Research (CIHR) postdoctoral fellowship awarded to A.K.C.B., by CIHR operating grant MT11318 and NIH grant R01AI066058 to P.S.H., and by National Institutes of Health grant R01GM55392 to S.D.G.
Footnotes
Published ahead of print on 5 February 2009.
Supplemental material for this article may be found at http://aem.asm.org/.
REFERENCES
- 1.Aki, T., and S. Adhya. 1997. Repressor induced site-specific binding of HU for transcription regulation. EMBO J. 16:3666-3674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Azam, T. A., A. Iwata, A. Nishimura, S. Ueda, and A. Ishihama. 1999. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 181:6361-6370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bachman, M. A., and M. S. Swanson. 2001. RpoS co-operates with other factors to induce Legionella pneumophila virulence in the stationary phase. Mol. Microbiol. 40:1201-1214. [DOI] [PubMed] [Google Scholar]
- 4.Bachman, M. A., and M. S. Swanson. 2004. Genetic evidence that Legionella pneumophila RpoS modulates expression of the transmission phenotype in both the exponential phase and the stationary phase. Infect. Immun. 72:2468-2476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bachman, M. A., and M. S. Swanson. 2004. The LetE protein enhances expression of multiple LetA/LetS-dependent transmission traits by Legionella pneumophila. Infect. Immun. 72:3284-3293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Balandina, A., L. Claret, R. Hengge-Aronis, and J. Rouvière-Yaniv. 2001. The Escherichia coli histone-like protein HU regulates rpoS translation. Mol. Microbiol. 39:1069-1079. [DOI] [PubMed] [Google Scholar]
- 7.Berger, K. H., and R. R. Isberg. 1993. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol. Microbiol. 7:7-19. [DOI] [PubMed] [Google Scholar]
- 8.Berk, S. G., G. Faulkner, E. Garduño, M. C. Joy, M. A. Ortiz-Jimenez, and R. A. Garduño. 2008. Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a Dot/Icm-mediated survival mechanism. Appl. Environ. Microbiol. 74:2187-2199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Berk, S. G., R. S. Ting, G. W. Turner, and R. J. Ashburn. 1998. Production of respirable vesicles containing live Legionella pneumophila cells by two Acanthamoeba spp. Appl. Environ. Microbiol. 64:279-286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bonnefoy, E., and J. Rouvière-Yaniv. 1991. HU and IHF, two homologous histone-like proteins of Escherichia coli, form different protein-DNA complexes with short DNA fragments. EMBO J. 10:687-696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reference deleted.
- 12.Byrne, B., and M. S. Swanson. 1998. Expression of Legionella pneumophila virulence traits in response to growth conditions. Infect. Immun. 66:3029-3034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Castañeda, M., J. Sánchez, S. Moreno, C. Núñez, and G. Espín. 2001. The global regulators GacA and σS form part of a cascade that controls alginate production in Azotobacter vinelandii. J. Bacteriol. 183:6787-6793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cirillo, J. D., S. Falkow, and L. S. Tompkins. 1994. Growth of Legionella pneumophila in Acanthamoeba castellani enhances invasion. Infect. Immun. 62:3254-3261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Craig, N. L., and H. A. Nash. 1984. E. coli integration host factor binds to specific sites in DNA. Cell 39:707-716. [DOI] [PubMed] [Google Scholar]
- 16.Dame, R. T., and N. Goosen. 2002. HU: promoting or counteracting DNA compaction. FEBS Lett. 529:151-156. [DOI] [PubMed] [Google Scholar]
- 17.Davis, G. S., and W. C. Winn, Jr. 1987. Legionnaires' disease: respiratory infections caused by Legionella bacteria. Clin. Chest Med. 8:419-439. [PubMed] [Google Scholar]
- 18.Ditto, M. D., D. Roberts, and R. A. Weisberg. 1994. Growth phase variation of integration host factor level in Escherichia coli. J. Bacteriol. 176:3738-3748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Faulkner, G., and R. A. Garduño. 2002. Ultrastructural analysis of differentiation in Legionella pneumophila. J. Bacteriol. 184:7025-7041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Faulkner, G., S. G. Berk, E. Garduno, M. A. Ortiz-Jimenez, and R. A. Garduno. 2008. Passage through Tetrahymena tropicalis triggers a rapid morphological differentiation in Legionella pneumophila. J. Bacteriol. 190:7728-7738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Feeley, J. C., R. J. Gibson, G. W. Gorman, N. C. Langford, J. K. Rasheed, D. C. Mackel, and W. B. Baine. 1979. Charcoal-yeast extract agar: primary isolation medium for Legionella pneumophila. J. Clin. Microbiol. 10:437-441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fettes, P. S., V. Forsbach-Birk, D. Lynch, and R. Marre. 2001. Overexpression of a Legionella pneumophila homologue of the E. coli regulator csrA affects cell size, flagellation, and pigmentation. Int. J. Med. Microbiol. 291:353-360. [DOI] [PubMed] [Google Scholar]
- 23.Fields, B. S., R. F. Benson, and R. E. Besser. 2002. Legionella and Legionnaires' disease: 25 years of investigation. Clin. Microbiol. Rev. 15:506-526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fyfe, J. A., and J. K. Davies. 1998. An AT-rich tract containing an integration host factor-binding domain and two UP-like elements enhances transcription from the pilEp1 promoter of Neisseria gonorrhoeae. J. Bacteriol. 180:2152-2159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gal-Mor, O., and G. Segal. 2003. The Legionella pneumophila GacA homolog (LetA) is involved in the regulation of icm virulence genes and is required for intracellular multiplication in Acanthamoeba castellanii. Microb. Pathog. 34:187-194. [DOI] [PubMed] [Google Scholar]
- 26.Garduño, R. A., E. Garduño, M. Hiltz, and P. S. Hoffman. 2002. Intracellular growth of Legionella pneumophila gives rise to a differentiated form dissimilar to stationary-phase forms. Infect. Immun. 70:6273-6283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Garduño, R. A., E. Garduño, and P. S. Hoffman. 1998. Surface-associated Hsp60 chaperonin of Legionella pneumophila mediates invasion in a HeLa cell model. Infect. Immun. 66:4602-4610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Garduño, R. A., F. D. Quinn, and P. S. Hoffman. 1998. HeLa cells as a model to study the invasiveness and biology of Legionella pneumophila. Can. J. Microbiol. 44:430-440. [PubMed] [Google Scholar]
- 29.Gober, J. W., and L. Shapiro. 1990. Integration host factor is required for the activation of developmentally regulated genes in Caulobacter. Genes Dev. 4:1494-1504. [DOI] [PubMed] [Google Scholar]
- 30.Goodman, S. D., N. J. Velten, Q. Gao, S. Robinson, and A. M. Segall. 1999. In vitro selection of integration host factor binding sites. J. Bacteriol. 181:3246-3255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Goosen, N., and P. van de Putte. 1995. The regulation of transcription initiation by integration host factor. Mol. Microbiol. 16:1-7. [DOI] [PubMed] [Google Scholar]
- 32.Granston, A. E., and H. A. Nash. 1993. Characterization of a set of integration host factor mutants deficient for DNA binding. J. Mol. Biol. 234:45-59. [DOI] [PubMed] [Google Scholar]
- 33.Greub, G., and D. Raoult. 2003. Morphology of Legionella pneumophila according to their location within Hartmanella vermiformis. Res. Microbiol. 154:619-621. [DOI] [PubMed] [Google Scholar]
- 34.Hales, L. M., and H. A. Shuman. 1999. The Legionella pneumophila rpoS gene is required for growth within Acanthamoeba castellanii. J. Bacteriol. 181:4879-4889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hammer, B. K., and M. S. Swanson. 1999. Coordination of Legionella pneumophila virulence with entry into stationary phase by ppGpp. Mol. Microbiol. 33:721-731. [DOI] [PubMed] [Google Scholar]
- 36.Hammer, B. K., E. S. Tateda, and M. S. Swanson. 2002. A two-component regulator induces the transmission phenotype of stationary-phase Legionella pneumophila. Mol. Microbiol. 44:107-118. [DOI] [PubMed] [Google Scholar]
- 37.Hill, S. A., D. S. Samuels, C. Nielsen, S. W. Knight, F. Pagotto, and J. A. R. Samuels. 2002. Integration host factor interactions with Neisseria gene sequences: correlation between predicted binding sites and in vitro binding of Neisseria-derived IHF protein. Mol. Cell Probes 16:153-158. [DOI] [PubMed] [Google Scholar]
- 38.Hiltz, M. F., G. R. Sisson, A. K. Brassinga, E. Garduño, R. A. Garduño, and P. S. Hoffman. 2004. Expression of magA in Legionella pneumophila Philadelphia-1 is developmentally regulated and a marker of formation of mature intracellular forms. J. Bacteriol. 186:3038-3045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hoffman, P. S., C. A. Butler, and F. D. Quinn. 1989. Cloning and temperature-dependent expression in Escherichia coli of a Legionella pneumophila gene coding for a genus-common 60-kilodalton antigen. Infect. Immun. 57:1731-1739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hwang, D. S., and A. Kornberg. 1992. Opening of the replication origin of Escherichia coli by DnaA protein with protein HU or IHF. J. Biol. Chem. 267:23083-23086. [PubMed] [Google Scholar]
- 41.LeBlanc, J. J., A. K. Brassinga, F. Ewann, R. J. Davidson, and P. S. Hoffman. 2008. An ortholog of OxyR in Legionella pneumophila is expressed postexponentially and negatively regulates the alkyl hydroperoxide reductase (ahpC2D) operon. J. Bacteriol. 190:3444-3455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.LeBlanc, J. J., R. J. Davidson, and P. S. Hoffman. 2006. Compensatory functions of two alkyl hydroperoxide reductases in the oxidative defense system of Legionella pneumophila. J. Bacteriol. 188:6235-6244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Li, M., I. Rosenshine, S. L. Tung, X. H. Wang, D. Friedberg, C. L. Hew, and K. Y. Leung. 2004. Comparative proteomic analysis of extracellular proteins of enterohemorrhagic and enteropathogenic Escherichia coli strains and their ihf and ler mutants. Appl. Environ. Microbiol. 70:5274-5282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Marshall, D. G., B. J. Sheehan, and C. J. Dorman. 1999. A role for the leucine-responsive regulatory protein and integration host factor in the regulation of the Salmonella plasmid virulence (spv) locus in Salmonella typhimurium. Mol. Microbiol. 34:134-145. [DOI] [PubMed] [Google Scholar]
- 45.McNealy, T. L., V. Forsbach-Birk, C. Shi, and R. Marre. 2005. The Hfq homologue in Legionella pneumophila demonstrates regulation by LetA and RpoS and interacts with the global regulator CsrA. J. Bacteriol. 187:1527-1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Molofsky, A. B., and M. S. Swanson. 2003. Legionella pneumophila CsrA is a pivotal repressor of transmission traits and activator of replication. Mol. Microbiol. 50:445-461. [DOI] [PubMed] [Google Scholar]
- 47.Molofsky, A. B., and M. S. Swanson. 2004. Differentiate to thrive: lessons from the Legionella pneumophila life cycle. Mol. Microbiol. 53:29-40. [DOI] [PubMed] [Google Scholar]
- 48.Poindexter, J. S. 1964. Biological properties and classification of the Caulobacter group. Bacteriol. Rev. 28:231-295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Porter, M., and C. J. Dorman. 1997. Positive regulation of Shigella flexneri virulence genes by integration host factor. J. Bacteriol. 179:6537-6550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Quon, K. C., G. T. Marczynski, and L. Shapiro. 1996. Cell cycle control by an essential bacterial two-component signal transduction protein. Cell 84:83-93. [DOI] [PubMed] [Google Scholar]
- 51.Rice, P. A., S. Yang, K. Mizuuchi, and H. A. Nash. 1996. Crystal structure of an IHF-DNA complex: a protein-induced DNA U-turn. Cell 87:1295-1306. [DOI] [PubMed] [Google Scholar]
- 52.Sadoff, H. L. 1973. Comparative aspects of morphogenesis in three prokaryotic genera. Annu. Rev. Microbiol. 27:133-153. [DOI] [PubMed] [Google Scholar]
- 53.Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 54.Segal, G., M. Feldman, and T. Zusman. 2005. The Icm/Dot type-IV secretion systems of Legionella pneumophila and Coxiella burnetii. FEMS Microbiol. Rev. 29:65-81. [DOI] [PubMed] [Google Scholar]
- 55.Sexton, J. A., J. S. Pinkner, R. Roth, J. E. Heuser, S. J. Hultgren, and J. P. Vogel. 2004. The Legionella pneumophila PilT homologue DotB exhibits ATPase activity that is critical for intracellular growth. J. Bacteriol. 186:1658-1666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Siam, R., A. K. C. Brassinga, and G. T. Marczynski. 2003. A dual binding site for integration host factor and the response regulator CtrA inside the Caulobacter crescentus replication origin. J. Bacteriol. 185:5563-5572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sierra, R., D. J. Comerci, L. I. Pietrasanta, and R. A. Ugalde. 2004. Integration host factor is involved in transcriptional regulation of the Brucella abortus virB operon. Mol. Microbiol. 54:808-822. [DOI] [PubMed] [Google Scholar]
- 58.Stonehouse, E., G. Kovacikova, R. K. Taylor, and K. Skorupski. 2008. Integration host factor positively regulates virulence gene expression in Vibrio cholerae. J. Bacteriol. 190:4736-4748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Swinger, K. K., and P. A. Rice. 2004. IHF and HU: flexible architects of bent DNA. Curr. Opin. Struct. Biol. 14:28-35. [DOI] [PubMed] [Google Scholar]
- 60.Tiaden, A., T. Spirig, S. S. Weber, H. Brüggemann, R. Bosshard, C. Buchrieser, and H. Hilbi. 2007. The Legionella pneumophila response regulator LqsR promotes host cell interactions as an element of the virulence regulatory network controlled by RpoS and LetA. Cell. Microbiol. 9:2903-2920. [DOI] [PubMed] [Google Scholar]
- 61.Vogel, J. P., C. Roy, and R. R. Isberg. 1996. Use of salt to isolate Legionella pneumophila mutants unable to replicate in macrophages. Ann. N. Y. Acad. Sci. 797:271-272. [DOI] [PubMed] [Google Scholar]
- 62.Yona-Yadler, C., T. Umanski, S. I. Aizawa, D. Friedberg, and I. Rosenshine. 2003. Integration host factor (IHF) mediates repression of flagella in enteropathogenic and enterohaemorrhagic Escherichia coli. Microbiology 149:877-884. [DOI] [PubMed] [Google Scholar]
- 63.Zhong, J., A. L. Douglas, and T. P. Hatch. 2001. Characterization of integration host factor (IHF) binding upstream of the cysteine-rich protein operon (omcAB) promoter of Chlamydia trachomatis LGV serovar L2. Mol. Microbiol. 41:451-462. [DOI] [PubMed] [Google Scholar]
- 64.Zulianello, L., E. de la Gorgue de Rosny, P. van Ulsen, P. van de Putte, and N. Goosen. 1994. The HimA and HimD subunits of integration host factor can specifically bind to DNA as homodimers. EMBO J. 13:1534-1540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zusman, T., O. Gal-Mor, and G. Segal. 2002. Characterization of a Legionella pneumophila relA insertion mutant and roles of RelA and RpoS in virulence gene expression. J. Bacteriol. 184:67-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.