

Abstract
Prolactin (PRL) is widely considered to be the juvenile hormone of anuran tadpoles and to counteract the effects of thyroid hormone (TH), the hormone that controls amphibian metamorphosis. This putative function was concluded mainly from experiments in which mammalian PRL was injected into tadpoles or added to cultured tadpole tissues. In this study, we show that overexpression of ovine or Xenopus laevis PRL in transgenic X. laevis does not prolong tadpole life, establishing that PRL does not play a role in the life cycle of amphibians that is equivalent to that of juvenile hormone in insect metamorphosis. However, overexpression of PRL produces tailed frogs by reversing specifically some but not all of the programs of tail resorption and stimulating growth of fibroblasts in the tail. Whereas TH induces muscle resorption in tails of these transgenics, the tail fibroblasts continue to proliferate resulting in a fibrotic tail that is resistant to TH.
Metamorphosis in amphibians and insects is similar in many aspects and is controlled by thyroid hormone (TH) and ecdysone, respectively. The effects of both hormones are mediated by receptors that are ligand-dependent transcription factors and belong to the nuclear receptor superfamily. Another well established hormone that is involved in insect metamorphosis is juvenile hormone (JH). JH counteracts the metamorphic effects of ecdysone. The levels of JH are high during larval stages, and only when the level of JH drops after the last instar does metamorphosis occur (1).
The influence of prolactin (PRL) on amphibian metamorphosis has been the subject of many studies with contradictory results. In particular, administration of mammalian PRL has been reported to have antimetamorphic effects and to prolong the tadpole life of many amphibian species (2–4). It has also been reported that mammalian PRL counteracts the effects of TH when added to cultured tadpole tissues (5, 6). These observations have led to a widely held belief that PRL has the biological role of a “juvenile” hormone in amphibian metamorphosis (7, 8). It was predicted that the levels of endogenous PRL, just like that of JH in insects, should be high at premetamorphosis and prometamorphosis stages and then drop at the climax of metamorphosis (7). However, the direct measurements of both endogenous PRL mRNA (9, 10) and protein levels of PRL in the serum (11, 12) showed them to be low at premetamorphosis and prometamorphosis stages rising substantially only at the climax of metamorphosis, a result that does not support a juvenilizing role for endogenous PRL. Furthermore, the interpretations of earlier experiments were confounded by the questionable purity and unknown specificity of mammalian hormone preparations.
In this paper, we reinvestigate the role of PRL in X. laevis metamorphosis. Transgenic X. laevis tadpoles overexpressing either X. laevis PRL (xPRL) or ovine PRL (oPRL) reached the metamorphic climax at the same time as nontransgenic siblings. Therefore, we conclude that PRL does not play a normal biological role in amphibians that is equivalent to that of JH in insect metamorphosis. However, overexpression of PRL, especially oPRL, produces tailed frogs by inhibiting specifically some but not all of the programs of tail resorption and stimulating growth of fibroblasts in the tail.
Materials and Methods
The xPRL and oPRL cDNA Plasmids.
The coding region of xPRL (10) was amplified by PCR with primers 5′-CGCGGATCCATAATGATTGATCCGATG-3′ and 5′-CAAATGAATTCCTAACAGTTGCTGTCATG-3′ by using pfu polymerase (Stratagene) and cloned into the BamHI and EcoRI sites of the pCS2+ vector (13). The coding region of xPRL was also amplified with primers 5′-GCGCGGATCCACAGCCACCATGATTGATCCGATGGACTC-3′ and 5′-CAAATGAATTCCTAACAGTTGCTGTCATG-3′ to incorporate an optimal Kozak sequence for translation initiation. The coding region of oPRL (14) was amplified with primers 5′-CGACGGATCCACCACCATGGACAGCAAAGG-3′ and 5′-CGCGGAATTCGGGCTTAGCAGTTGTTGTTG-3′ and cloned into the BamHI and EcoRI sites of the pCS2+ vector. The coding region of xPRL receptor A (xPRLR-A) was amplified by PCR with primers 5′-CGGAGAATTCACAGCCACCATGCCGCAGAACTTGGCACC-3′ and 5′-GCGCCTCGAGTTAAGAAAGAAAAGCTGAAGGATCC-3′ and cloned into the EcoRI and XhoI sites of the plasmid pCSGFP3 (a gift of Enrique Amaya, Wellcome/CRC Institute, Cambridge, U.K.) replacing green fluorescent protein.
Cloning of xPRLR cDNAs, Northern Blots, and Cell Culture Assays.
Reverse transcription–PCR was performed by using RNA isolated from tadpole tail and degenerate primers corresponding to well conserved regions of PRLR between mammalian, avian, and fish species. Primers used were 5′-CCHGANAARGARACHTTYACHTGYTGGTGG-3′, which corresponds to the conserved peptide PE(D)KETFTCWW, and 5′-CCTTTDATYTTWGGNCCNGGNACTGGNGG-3′, which corresponds to the conserved peptide PPVPGPKIKG. The PCR condition was 3 cycles of 94°C for 30s, 55°C for 1 min, and 72°C for 3 min, followed by 30 cycles of 94°C for 30 s, 60°C for 1 min, and 72°C for 2 min. The hot-start PCR technique was applied with the AmpliWax PCR Gem 100 wax bead (Perkin–Elmer). The amplified fragment was cloned, sequenced, and used to screen a tail cDNA phage library as described (10). Two copies of xPRLR cDNA (xPRLR-A and xPRLR-B) were identified. The xPRLR-A cDNA was used in these experiments. The methods for Northern blot, cell culture, and transient transfection have been described (15).
Transgenesis.
Procedures of transgenesis, the identification of transgenic animals, and tadpole-raising conditions have been described (16, 17). X. laevis tadpoles were staged according to the criteria of Nieuwkoop and Faber (18). Stage 59 is the onset of metamorphic climax.
Histology.
Sections of tails (19) were stained with hematoxylin and eosin.
Results
Cloning of xPRLR cDNA.
To compare the specificity of xPRL and oPRL, the hormone that has been administered most frequently to tadpoles in earlier studies, we cloned the xPRLR cDNA. A DNA fragment was amplified by PCR from tadpole tail cDNA by using degenerate primers corresponding to conserved regions of PRLR between mammalian and avian species. The amplified fragment was cloned, sequenced, and used to screen a tadpole tail cDNA library. Two 2.3-kilobase full-length cDNA clones (xPRLR-A and xPRLR-B) were identified that encode 611 amino acid residues. They are 87% identical to each other and 44–52% identical to mammalian PRLRs. Sequences of xPRLR that are most highly conserved include positionally conserved cysteines, the WSxWS motif, and the proline-rich box 1 region (20). The intracellular domain of the receptor is more diverged than its extracellular domain. xPRLR mRNAs were expressed uniformly at all stages of tadpole development in the whole body (Fig. 1A). However, expression of xPRLR mRNA rises in the tail late in climax to about five times the level in the head (Fig. 1B). Just as the developmental expression of xPRL by the pituitary gland was shown not to correlate with an expected role as a JH (10), neither does the developmental expression pattern of xPRLR during metamorphosis.
Figure 1.

Northern blot of xPRLR mRNA expression at different developmental stages. (A) Whole tadpole poly(A)-RNA; (B) Tail total RNA. The blots were hybridized first with the xPRLR probe and then the PR28 probe, which is used to standardize RNA loading (29). The marks on the left indicate the positions of the ribosomal RNAs. The numbers on top of the lanes are tadpole stages (18).
Specificity of xPRL and oPRL.
xPRL, oPRL, and X. laevis growth hormone (xGH) were tested for their interaction with xPRLR by a transient transfection assay (21). xPRL and xGH were prepared by transfecting human embryonic kidney fibroblast 293 cells with DNA constructs expressing xPRL and xGH, respectively. The hormone was secreted into the culture medium, and its concentration was quantified by Western blotting (data not shown). The relative affinity of xPRL and oPRL for xPRLR was compared by a dilution series of both hormones (Fig. 2). At concentrations of 50 ng/ml or higher, oPRL and xPRL induced luciferase activity equally well. At concentrations of 10 ng/ml or lower, xPRL induces the reporter much better than oPRL, suggesting that xPRL has a higher affinity for xPRLR than does oPRL. xGH does not activate xPRLR even at 500 ng/ml (Fig. 2). Interestingly, oPRL but not xPRL cross-reacts with xGH receptor significantly at a concentration of 250 ng/ml or higher (15).
Figure 2.

Specificity of different hormones for xPRLR as determined by a transient transfection assay (15). Luciferase (Luc) reporter activity was assayed and normalized to β-galactosidase activity. Each column represents the mean of triplicate samples, and the error bar represents standard deviation. The hormone concentrations used are no hormone/control (Con; bar 1), 500 ng/ml (bar 2), 50 ng/ml (bar 3), 5 ng/ml (bar 4), 50 ng/ml (bar 5), and 5 ng/ml (bar 6).
Overexpression of PRL.
To test the effect of high levels of PRL on metamorphosis and compare the effects of xPRL and oPRL, transgenic tadpoles were produced overexpressing xPRL or oPRL driven by a cytomegalovirus promoter. There was no obvious difference between transgenic and nontransgenic tadpoles before stage 50. After stage 50, transgenic tadpoles of both xPRL and oPRL grew bigger and longer tails than their nontransgenic siblings. At stage 59, the onset of the climax of metamorphosis, the tails of both xPRL and oPRL transgenic tadpoles were about 15% longer than control tails. oPRL transgenic tadpoles weighed about 30–50% more than controls at stage 59 (15). There was no significant difference between the weights of xPRL transgenics and controls, suggesting that the increased weight of oPRL transgenics could reflect the activation of xGH receptor by oPRL (15). Both xPRL and oPRL transgenics reached stage 59 within the same time range as controls (Fig. 3). Thus, tadpole life was not prolonged in transgenics overexpressing either xPRL or oPRL. After stage 59, gill resorption occurred normally in transgenics, producing a stage 63 tadpole with a long tail. In control tadpoles, tail resorption is completed in the next 3–5 days, and the froglets begin to eat worms. These transgenic tadpoles begin eating worms at the same time as controls, but they do not resorb their tails (Fig. 4 A–D). In normal tadpoles at climax, the dorsal and ventral fins resorb first (Fig. 5A). Next, tail muscle loses its characteristic repeated chevron structure, and the muscle cells die as the notochord collapses allowing the tail to shorten (19). In tadpoles overexpressing oPRL, the muscle resorbs at the same time as it does in controls, and there is some localized collapse of the notochord often at the junction of the tail and body. Remarkably, the fins do not resorb. In fact, they continue to grow in the tailed frog (Figs. 4 E and F and 5 B and C).
Figure 3.

The time it takes tadpoles to reach stage 59. Transgenic tadpoles overexpressing xPRL or oPRL compared with sibling controls (Con) raised together in four separate experiments. Animals must be raised together in the same tank because of the effect of even slight differences in rearing conditions on the rate of tadpole development. Each point represents an individual animal.
Figure 4.

Transgenic froglets overexpressing PRL. (A and B) Representative oPRL transgenic froglets at the end of metamorphosis. (C and D) Representative xPRL transgenic froglets at the end of metamorphosis. (E) Dorsal view of an oPRL tailed frog 3 months after metamorphosis. (F) Side view of the animal shown in E. Note the huge fibrotic fins that have grown since metamorphosis. The scale bars equal 1 cm. A–D have the same magnification; E and F have the same magnification.
Figure 5.
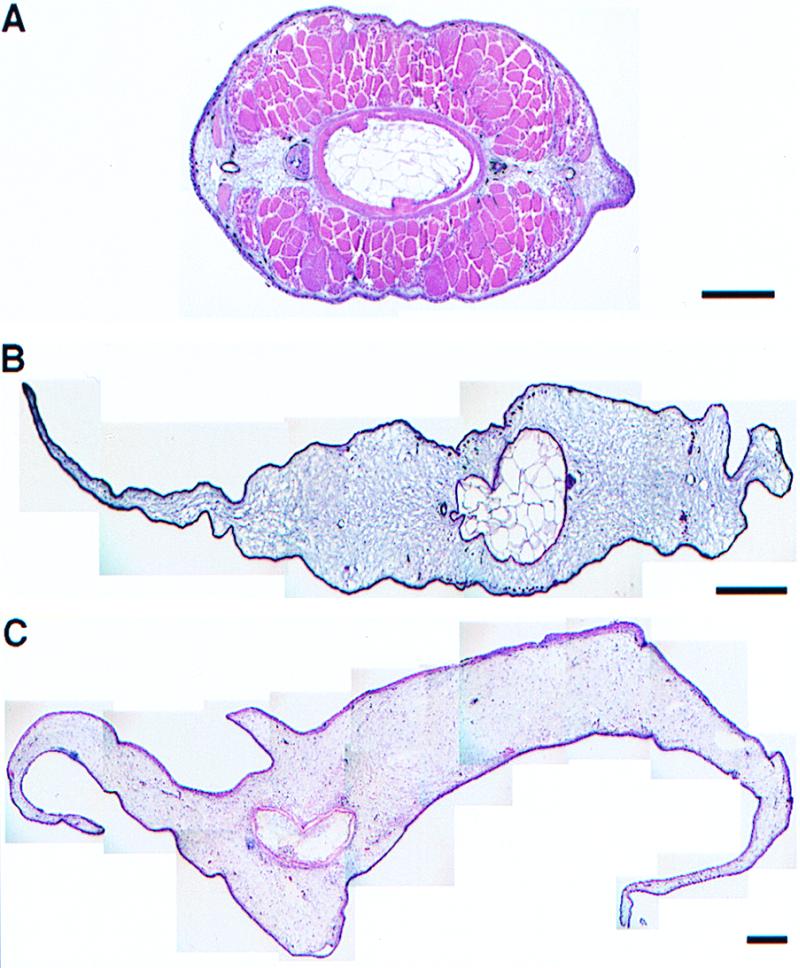
Some but not all of the programs of tail resorption are inhibited. (A) Image of a control tadpole tail at stage 64 showing that the fins are already resorbed and that the muscle is dying as the tail starts to resorb quickly. (B) An oPRL transgenic tail at the end of metamorphosis. The fins have grown but most of the muscle has resorbed. (C) An oPRL transgenic tail 3 months after metamorphosis. Note the growth of the retained tail. All sections are cross sections stained with hematoxylin-eosin. The scale bars equal 1 mm.
The permanent tail that develops in these otherwise normal frogs maintains a notochord through most of its length. There is a spinal cord, huge fibrotic fins, and a dominant dorsal and ventral blood vessel. The only remaining muscle is the pairs of dorsal and ventral repeating bundles of slow muscle referred to as “cords” (22). The tail is full of collagen-producing cells as detected by collagen-specific stains and by in situ hybridization of known collagen genes (data not shown). Tails of tadpoles overexpressing either oPRL or xPRL were about 15% longer than those of controls, but oPRL was much more potent in inhibiting tail resorption than xPRL. Of 38 tadpoles transgenic for oPRL, 35 became tailed frogs, whereas only 8 of 49 tadpoles transgenic for xPRL retained their tails. The tails retained by xPRL frogs are smaller than those of oPRL frogs, although they share the same structural features described above.
The retained tails are remarkably resistant to TH. On day 11 after tailed frogs began to eat worms, they were treated with different concentrations of TH. A concentration of 10 nM T3 had no effect on tails in 3 weeks; 50 nM T3 caused tails to shorten in half in 10 days. However, the remaining tail did not shorten further. It was fixed for histology 3.5 weeks after the addition of T3. The TH-induced shortening is probably due to the partial loss of the notochord sheath, which is a required step in normal tail resorption (19). The muscle “cords,” which are normally the last muscle elements to be resorbed in the tail (22), are the only muscles that remain in these fibrotic tails. They actually are larger as a result of TH-induction perhaps because of synthesis of new cord muscle (data not shown).
Discussion
These results show that PRL overexpression does not influence the time it takes tadpoles to reach metamorphic climax (Fig. 3). This experiment contradicts those studies that led to the original conclusion that PRL is a JH. These were mainly “overexpression” studies in which mammalian (usually ovine) PRL was injected into tadpoles (2) or added to cultured tadpole tissues (5, 6). However, it has been reported that the injection of ovine PRL (oPRL) antiserum accelerates TH-induced metamorphosis in Rana catesbeiana tadpoles (23). However, these authors pointed out that oPRL antiserum did not cross-react with any protein in a Rana tadpole pituitary homogenate, questioning the interpretation of these antiserum injection experiments. We have confirmed that at least one excellent oPRL antiserum obtained from the National Institute of Diabetes and Digestive and Kidney Diseases National Hormone and Pituitary Program (AFP-C3581016789) does not cross-react with xPRL (data not shown). Subsequently, it was reported that a homologous bullfrog PRL antiserum could also accelerate bullfrog metamorphosis, although it was not shown whether that antiserum could neutralize bullfrog PRL (24). The growth rate of mice has been reported to decrease when they are treated with PRL antiserum (25). However, transgenic mice lacking either the PRLR (26) or PRL gene (27) grow at the same rate as controls.
We were unable to detect xPRL protein in the serum of control or xPRL transgenic froglets by Western blot, whereas oPRL protein could be detected in the serum of oPRL transgenic froglets (data not shown). Therefore, a PRL concentration difference might account for the different severity of the phenotype of oPRL and xPRL transgenics. The reason that xPRL protein level was lower in serum is not clear, although the xPRL transgene mRNA was abundant and easily detected by Northern blotting (data not shown). xPRL cDNA with an optimal Kozak consensus sequence and one with the endogenous sequence for translation initiation were used for transgenesis and produced the same percentage of tailed frogs. To date, the tailed frogs overexpressing oPRL are 7 months old and healthy. The males are sexually mature, and one of them has mated successfully. These F0 frogs grow slightly faster than control frogs, but their growth rate is much slower than that of the frogs overexpressing xGH (15). As the frogs grow, most of the tail is lost, probably by necrosis caused by inadequate blood supply, leaving a small fibrotic tail stump.
The tail fibroblasts normally are induced by TH at climax to change their synthetic program from the production of collagen as part of a growth program to one that includes proteolytic enzymes as part of the resorption program (19). Overexpressing PRL clearly stimulates a fibroblast growth program, agreeing with the earlier observation that excess PRL stimulates collagen synthesis in tadpole tail fins (28). The tails of these transgenic frogs are remarkably resistant to added TH, an observation that has been made repeatedly on in vitro cultures of tail fins (6). The persistent notochord sheath can be induced to resorb by adding TH. The tails of transgenic frogs persist presumably because of the fibrotic fins and the continued and augmented synthetic activity of the fibroblasts. The normal tadpole tail has a higher level of PRLR mRNA at the climax of metamorphosis than at premetamorphosis (Fig. 1B), but the combination of this increased receptor mRNA level and the rise in endogenous PRL that occurs at climax (10) is unlikely to play a role in the timing of tail resorption. The fins are normally the first tail structures to resorb at the climax of metamorphosis. We have prepared transgenic tadpoles overexpressing both oPRL and xPRLR with the idea that the metamorphic changes of tissues and organs other than the tail could be inhibited by excess oPRL when they express higher levels of xPRLR. However, in those animals, only tail resorption was inhibited (data not shown).
The inhibition of tail resorption by excess PRL is a “pharmacological” effect that might even be transmitted through a receptor other than the xPRLR. However, the remarkable specificity of PRL's effect on the tail as a target for TH provides a powerful tool to pursue the molecular basis of tail resorption.
Acknowledgments
We are grateful to J. Rosen for STAT5A1 construct, P. A. Kelly for β-casein/luciferase construct, T. Adams for oPRL cDNA, S. Kikuyama for xPRL antiserum, C. M. Fan for 293 cells and pCEP4 and pREP4 plasmids, A. F. Parlow and the National Hormone and Pituitary Program of the National Institute of Diabetes, Digestive, and Kidney Diseases for various mammalian hormones and antisera. We thank D. L. Berry for the stage 64 tail section (Fig. 5A), A. M. Schreiber for help with statistical analysis, and B. F. Remo and T. Liu for excellent technical support. Our colleagues and J. R. Tata provided helpful comments on the manuscript. This research has been performed by H.H. in partial fulfillment of the requirements for the Ph.D. degree at the Johns Hopkins University. This work was supported by grants to D.D.B. from the National Institutes of Health and the G. Harold and Leila Y. Mathers Charitable Trust.
Abbreviations
- TH
thyroid hormone
- JH
juvenile hormone
- PRL
prolactin
- oPRL
ovine PRL
- xPRL
X. laevis PRL
- PRLR
prolactin receptor
- xGH
X. laevis growth hormone
Footnotes
References
- 1.Gilbert L I, Rybczynski R, Tobe S S. In: Metamorphosis: Postembryoic Reprogramming of Gene Expression in Amphibian and Insect Cells. Gilbert L I, Tata J R, Atkinson B G, editors. San Diego: Academic; 1996. pp. 59–107. [Google Scholar]
- 2.Dodd M H I, Dodd J M. In: Physiology of the Amphibia. Lofts B, editor. Vol. 3. New York: Academic; 1976. pp. 467–599. [Google Scholar]
- 3.White B A, Nicoll C S. In: Metamorphosis: A Problem in Developmental Biology. Gilbert L I, Frieden E, editors. New York: Plenum; 1981. pp. 363–396. [Google Scholar]
- 4.Denver R J. In: Metamorphosis: Postembryoic Reprogramming of Gene Expression in Amphibian and Insect Cells. Gilbert L I, Tata J R, Atkinson B G, editors. San Diego: Academic; 1996. pp. 433–464. [Google Scholar]
- 5.Derby A, Etkin W. J Exp Zool. 1968;169:1–8. doi: 10.1002/jez.1401690102. [DOI] [PubMed] [Google Scholar]
- 6.Tata J R, Kawahara A, Baker B S. Dev Biol. 1991;146:72–80. doi: 10.1016/0012-1606(91)90447-b. [DOI] [PubMed] [Google Scholar]
- 7.Etkin W, Gona A G. J Exp Zool. 1967;165:249–258. doi: 10.1002/jez.1401650209. [DOI] [PubMed] [Google Scholar]
- 8.Bern H A, Nicoll C S, Strohan R C. Proc Soc Exp Biol Med. 1967;126:518–520. doi: 10.3181/00379727-126-32493. [DOI] [PubMed] [Google Scholar]
- 9.Takahashi N, Yoshihama K, Kikuyama S, Yamamoto K, Wakabayashi K, Kato Y. J Mol Endocrinol. 1990;5:281–287. doi: 10.1677/jme.0.0050281. [DOI] [PubMed] [Google Scholar]
- 10.Buckbinder L, Brown D D. Proc Natl Acad Sci USA. 1993;90:3820–3824. doi: 10.1073/pnas.90.9.3820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Clemons G K, Nicoll C S. Gen Comp Endocrinol. 1977;32:531–535. doi: 10.1016/0016-6480(77)90237-4. [DOI] [PubMed] [Google Scholar]
- 12.Yamamoto K, Kikuyama S. Endocrinol Jpn. 1982;29:159–167. doi: 10.1507/endocrj1954.29.159. [DOI] [PubMed] [Google Scholar]
- 13.Turner D L, Weintraub H. Genes Dev. 1994;8:1434–1447. doi: 10.1101/gad.8.12.1434. [DOI] [PubMed] [Google Scholar]
- 14.Adams T E, Baker L, Brandon M R. Nucleic Acids Res. 1989;17:440. doi: 10.1093/nar/17.1.440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Huang H, Brown D D. Proc Natl Acad Sci USA. 2000;97:190–194. doi: 10.1073/pnas.97.1.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kroll K L, Amaya E. Development (Cambridge, UK) 1996;122:3173–3183. doi: 10.1242/dev.122.10.3173. [DOI] [PubMed] [Google Scholar]
- 17.Huang H, Marsh-Armstrong N, Brown D D. Proc Natl Acad Sci USA. 1999;96:962–967. doi: 10.1073/pnas.96.3.962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nieuwkoop P D, Faber J. Normal Table of Xenopus laevis (Daudin) Amsterdam: Elsevier/North-Holland; 1956. [Google Scholar]
- 19.Berry D L, Schwartzman R A, Brown D D. Dev Biol. 1998;203:12–23. doi: 10.1006/dbio.1998.8974. [DOI] [PubMed] [Google Scholar]
- 20.Kelly P A, Ali S, Rozakis M, Goujon L, Nagano M, Pellegrini I, Gould D, Djiane J, Edery M, Finidori J, et al. Recent Prog Horm Res. 1993;48:123–164. doi: 10.1016/b978-0-12-571148-7.50009-9. [DOI] [PubMed] [Google Scholar]
- 21.Lesueur L, Edery M, Ali S, Paly J, Kelly P A, Djiane J. Proc Natl Acad Sci USA. 1991;88:824–828. doi: 10.1073/pnas.88.3.824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ellinson R P, Remo B F, Brown D D. Dev Biol. 1999;215:243–252. doi: 10.1006/dbio.1999.9481. [DOI] [PubMed] [Google Scholar]
- 23.Eddy L, Lipner H. Gen Comp Endocrinol. 1975;25:462–466. doi: 10.1016/0016-6480(75)90157-4. [DOI] [PubMed] [Google Scholar]
- 24.Clemons G K, Nicoll C S. Gen Comp Endocrinol. 1977;31:495–497. doi: 10.1016/0016-6480(77)90042-9. [DOI] [PubMed] [Google Scholar]
- 25.Sinha Y N, Vanderlaan W P. Endocrinology. 1982;110:1871–1878. doi: 10.1210/endo-110-6-1871. [DOI] [PubMed] [Google Scholar]
- 26.Ormandy C J, Camus A, Barra J, Damotte D, Lucas B, Buteau H, Edery M, Brousse N, Babinet C, Binart N, et al. Genes Dev. 1997;11:167–178. doi: 10.1101/gad.11.2.167. [DOI] [PubMed] [Google Scholar]
- 27.Horseman N D, Zhao W, Montecino-Rodriguez E, Tanaka M, Nakashima K, Engle S J, Smith F, Markoff E, Dorshkind K. EMBO J. 1997;16:6926–6935. doi: 10.1093/emboj/16.23.6926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yoshizato K, Yasumasu I. Dev Growth Differ. 1970;11:305–317. doi: 10.1111/j.1440-169x.1970.00305.x. [DOI] [PubMed] [Google Scholar]
- 29.Shi Y B, Liang V C. Biochim Biophys Acta. 1994;1217:227–228. doi: 10.1016/0167-4781(94)90042-6. [DOI] [PubMed] [Google Scholar]