

Abstract
Endometriosis, a chronic disease characterized by endometrial tissue located outside the uterine cavity, affects one fourth of young women and is associated with chronic pelvic pain and infertility. However, an in-depth understanding of the pathophysiology and effective treatment strategies of endometriosis is still largely elusive. Inadequate immune and neuroendocrine responses are significantly involved in the pathophysiology of endometriosis, and key findings are summarized in the present review. We discuss here the role of different immune mechanisms particularly adhesion molecules, protein–glycan interactions, and pro-angiogenic mediators in the development and progression of the disease. Finally, we introduce the concept of endometrial dissemination as result of a neuroendocrine-immune disequilibrium in response to high levels of perceived stress caused by cardinal clinical symptoms of endometriosis.
Keywords: Corticotropin-releasing hormone, Ectopic endometrium, Inflammation, Progesterone, Sickness behaviour
Introduction
Endometriosis, first described in 1860 by von Rokitansky, is a chronic disease that is characterized by the occurrence of endometrial glands and stroma outside the uterine cavity [1]. Endometriosis affects up to 22% of women in their reproductive age [2] and is associated with chronic pelvic pain. Additionally, endometriosis is closely linked to severely impaired fertility, as it can be diagnosed in 68% of patients suffering from infertility [3]. To date, insights into the pathophysiology of endometriosis, and thus the development of effective treatment strategies, are surprisingly meager, which is attributed in part to the difficulties in studying the disease in humans.
Various forms of endometriosis have been described, such as endometriosis genitalis interna with endometrium adjacent to the eutopic endometrium, e.g., within the myometrium (adenomyosis) or the fallopian tubes. Further, endometriosis genitalis externa is being referred to if lesions can be located adjoining ovaries (endometriomas, endometriotic cysts), Douglas pouch, uterine ligaments, vagina, vulva, or perineum. Additionally, endometriosis extragenitalis mostly occurs within the pelvic cavity, septum rectovaginale, intestine, and ureter [4]. Deep infiltrating endometriosis, e.g., in the Douglas pouch, the sacrouterine ligaments, or the septum retrovaginale, is a very painful form of endometriosis characterized by fibrosis, smooth muscle proliferation, and fibromuscular nodules; this clinical entity is also called adenomysis externa [5]. According to Redwine [6], the most common sites of pelvic endometriosis are the Douglas pouch and the uterosacral ligaments.
A widely used classification of endometriosis has been introduced by the American Society for Reproductive Medicine (rASRM) and incorporates number, size, and location of endometrial implants, endometriomas, and/or adhesions as well as their morphology (red, white, and black) yielding to a classification from stage 1 (minimal endometriosis) to stage 4 (severe endometriosis) [7]. The unambiguous diagnosis of endometriosis is generally accepted by clinicians only after laparoscopic macroscopic evaluation and subsequent histomorphometric analyses of biopsies. Non-invasive diagnostic tools are not yet available.
Because the clinical symptoms of endometriosis range from severe dysmenorrhea, dyspareunia, dysuria to severe chronic pelvic pain, it is apparent that the disease rigorously interferes with the patients’ quality of life and affects social life, sexuality, and psychological well-being. Accordingly, due to the link between endometriosis and infertility, quality of life is even further reduced if women suffering from the clinical symptoms are surplus childless. It has been proposed that infertility in endometriosis patients might be due to altered folliculogenesis [8], reduced preovulatory steroidogenesis of granulosa cells [9], and decreased capability of fimbrial ovum capture in endometriosis [10]. Furthermore, sperm phagocytosis by peritoneal macrophages [11] and oviductal macrophages [12], anti-sperm antibodies [13], and reduced sperm penetration and velocity [14] have been observed. In addition, an altered egg–sperm interaction [15] and toxicity against early embryonic development [16], as well as defective implantation [17] have been proposed to be responsible for the infertility in endometriosis patients.
Etiology
Different theories have been proposed to explain the etiology of endometriosis: first and foremost, the transplantation theory [18], which is based on observations that retrograde menstruation of vital endometrium results in the implantation of such tissue into the peritoneum. To date, this theory is undeniably the most accepted concept. However, it may be challenged by the fact that retrograde menstruation is physiologically occurring in the majority of women, but endometriosis only occurs in approximately one fourth of women in their reproductive years. In this regard, endometriosis may be the result of a failing immune surveillance in the peritoneal cavity in women susceptible to endometriosis. Thus, immunologists should become far more attentive to this disease, as insights on its susceptibility, pathophysiology, and the identification of therapeutic approaches are likely to arise from immunologically based research. Another premise, the so-called metaplasia theory, is based on the notion that pluripotent coelomic cells differentiate into endometrial cells [19]. This theory may provide explanation for deep adenomyosis and relatively uncommon locations of endometriosis such as the brain or the lung. On the other hand, the denervation–reinnervation theory assumes that damage and denervation during valsalva maneuvers are followed by reinnervation [20] leading to a loss of uterine polar contractility and promoting retrograde menstruation. Spread endometrial tissue then adheres to injured tissue. Here, the extent of denervation and reinnervation is interpreted as primary source for clinical symptoms and their recurrence even after denervatory surgery. This theory is supported by the fact that the majority of patients with pelvic peritoneal defects and a history of pain also suffer from endometriosis [21].
Additionally, various exposure factors are thought to be associated to endometriosis, e.g., dioxin (2,3,7,8-tetrachlorodibenzo-p-dioxin) [22]. Here, the idea is that dioxin promotes the development of endometriosis by interfering with immune-mediated mechanisms, more specifically by stimulating the production of pro-inflammatory cytokines. A genetic etiology has also been proposed due to the high prevalence in relatives of affected women [23]. Although a wealth of genetic factors have been anticipated to be linked to an increased susceptibility to endometriosis [24], generally accepted marker gene(s) for endometriosis could not yet be identified. Emerging evidence points towards differential gene expression in eutopic and ectopic endometrium in humans [25, 26], and future research will be needed to provide a more in-depth understanding of the biological significance of this disease.
Immunological aspects in endometriosis
Because endometrial lesions are frequently present in the peritoneal cavity, they are in direct contact with peritoneal fluid, which bathes the pelvic cavity, uterus, fallopian tubes, and ovaries. However, in endometriosis, ectopic endometrial cells escape pathways involved in immune-mediated surveillance. In search for a better understanding of the pathogenesis of endometriosis, here, we analyzed different immune-mediated mechanisms and mediators that might be involved in disease development and resolution.
Role of innate immune responses in endometriosis
Macrophages
Macrophages are the main population of peritoneal leukocytes and—according to the published evidence so far—comprise up to 90% of peritoneal fluid cells [27]. In women suffering from endometriosis as well as in baboons with spontaneously occurring mild endometriosis, the concentration of peritoneal macrophages is increased as compared to healthy or infertile controls or animals with normal pelvis, respectively [27, 28] (Fig. 1). However, their percentage of total mononuclear cells seems to be decreased in favor of lymphocytes [29]. Further, an increased percentage of peritoneal macrophages is positive for the cell activation marker acid phosphatase in mild endometriosis [30]. Increased cell counts may be attributable to elevated levels of macrophage colony-stimulating factor (M-CSF) [31] and monocyte chemotactic protein (MCP)-1 [32] (Fig. 1). Here, M-CSF and MCP-1 have been proposed to derive from endometrial/endometriotic cells or peritoneal macrophages, respectively.
Fig. 1.
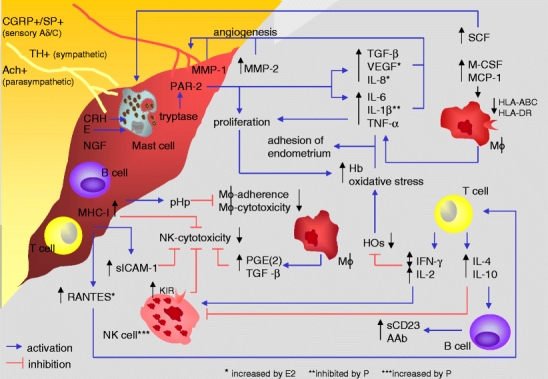
Neuroendocrine–immune disequilibrium in endometriosis. AAb autoantibodies, Ach achetylcholine, CGRP calcitonin-gene-related peptide, CRH corticotropin-releasing hormone, E estrogens, E2 estradiol, Hb hemoglobin, HO heme oxygenases, IL interleukin, HLA human leukocyte antigen, IFN-γ interferon-γ, KIR killer cell inhibitory receptor, M-CSF macrophage colony stimulating factor, MCP-1 monocyte chemotactic protein-1, MHC-I major histocompatibility complex class-I, MMP matrix metalloproteinase, NGF nerve growth factor, NK natural killer, P progesterone, PAR-2 protease-activated receptor-2, PGE(2) prostaglandin E(2), pHp peritoneal haptoglobin, RANTES regulated upon activation normal T cell expressed and secreted, sCD23 soluble CD23, SCF stem cell factor, sICAM-1 soluble intercellular adhesion molecule-1, SP substance P, TGF-β transforming growth factor-β, TH tyrosine hydroxylase, TNF-α tumor necrosis factor-α, VEGF vascular endothelial growth factor. In endometriosis, elevated levels of M-CSF, MCP-1, RANTES, and SCF in peritoneal fluid might lead to increased numbers of macrophages, T cells, and mast cells. Although markers of antigen presentation on macrophages such as HLA-ABC and HLA-DR are decreased in endometriosis, macrophage-derived IL-6, IL-1β, TNF-α, TGF-β, VEGF, and IL-8 are increased in peritoneal fluid, together with MMP-1 and MMP-2 stimulating angiogenesis. IL-6, IL-1β, and TNF-α support adhesion of endometrial cells to the peritoneum, and TNF-α stimulates the proliferation of ectopic tissue, resulting in high levels of Hb. T cell-derived IL-2 and IFN-γ decrease HO, leading to oxidative stress, and would, in sufficient levels, increase NK cell activity. IFN-γ has been inconsistently described as increased or decreased. Increased T cell-derived IL-4 and IL-10 inhibit cellular immunity and stimulate B cells to AAb production. sCD23 is increased in peritoneal fluid in endometriosis and might derive from activated B cells. Lymphocytes are increased in peritoneal fluid and abundantly present in ectopic tissue. Decreased NK cell cytotoxicity might be due to high anti-inflammatory T cytokines, increased KIR, high macrophage-derived PGE(2) and TGF-β, high MHC-I expression on ectopic cells, and high sICAM-1 levels in peritoneal fluid. Nerve fibers found within lesions are positive for CGRP, SP, TH, and Ach, and NGF and CRH were demonstrated. CRH and accumulated E can activate mast cells to release tryptase, activating PAR-2, which leads to increased secretion of VEGF, IL-8, and IL-6 and proliferation of ectopic tissue. pHp, expressed by ectopic tissue, decreases adherence and, in stage 3 and 4, cytotoxicity of peritoneal macrophages. E2 further increases RANTES, IL-8, and VEGF, whereas P inhibits IL-1β secretion from peritoneal macrophages and increases NK cell numbers
In vitro studies revealed that peritoneal macrophages derived from patients with endometriosis produce increased levels of the cytokines interleukin (IL)-6 [33], IL-1β, and tumor necrosis factor (TNF)-α [34], compared to peritoneal macrophages of women with other benign gynecological disorders (Fig. 1). Because IL-6, IL-1β, and TNF-α promote the adhesion of endometrial cells to peritoneum, increased secretion of these cytokines by peritoneal macrophages in patients with endometriosis might contribute to the development and progression of the disease [35] (Fig. 1). It is further noteworthy that IL-6, IL-1, and TNF-α correlate with infertility and embryotoxicity when secreted at high levels [36, 37]. Further, TNF-α induces proliferation of ectopic stromal cells, which would subsequently result in the growth of endometriotic lesions [38] (Fig. 1). Moreover, TNF-α target genes have been proposed to be overexpressed in experimental endometriosis in rats [39].
Elevated prostaglandins (PG), particularly PGE2 in the peritoneal fluid of endometriosis patients [40] (Fig. 1), may result from macrophage activation and have been proposed to subsequently aggravate endometriosis-associated pain by altering uterine and tubal contractility and cause infertility due to a delayed ovum transport [41]. On the other hand, emerging evidence addresses the decrease in human leukocyte antigen (HLA)-ABC and HLA-DR on peritoneal macrophages in endometriosis patients (Fig. 1), suggesting defective antigen presentation [42]. In advanced stages, macrophage-mediated cytotoxicity against endometrial cells is reduced as compared to early stages [43] (Fig. 1). Interestingly, cytotoxic activity could be restored by application of the non-steroidal, PG-inhibiting drug indomethacin, which points towards a dampening effect of PG on macrophage cytotoxicity in endometriosis [43] (Fig. 1). In early stages, however, the cytotoxicity of peritoneal macrophages is increased compared to fertile controls. Thus, hypothetically, activated macrophages in the peritoneal fluid may control the number and size of peritoneal endometriotic lesions. On the other hand, a wealth of mediators derived from peritoneal macrophages such as the abovementioned cytokines may promote adherence and proliferation of endometrial cells and angiogenesis, thus facilitating the dissemination of endometriotic lesions. In conclusion, additional work aiming to dissect the role of peritoneal macrophages in endometriosis is needed.
Mast cells
Mast cells play a pivotal role within innate immune responses. In addition, these cells play a critical role in sustaining Th2-mediated responses by secreting high levels of IL-4. Because mast cells are predominantly resident cells of loose connective tissue, mast cells’ presence and function have been investigated in endometriotic tissue [44]. Here, Kempuraj et al. observed increased numbers of highly activated mast cells in the stroma of peritoneal endometriotic lesions as compared to eutopic endometrium (Fig. 1). Mast cell migration and proliferation may be related to increased levels of stem cell factor (SCF) in the peritoneal fluid of women with early stage endometriosis and the expression of its cognate receptor on ectopic endometrium [45], whereby SCF may derive from fibroblasts, endothelial, and granulosa cells [46]. Activated mast cells release enzymes, such as tryptase, which stimulate protease-activated receptor (PAR)-2. Strikingly, PAR-2 agonist induces proliferation of purified endometriotic stromal cells and the release of IL-6 and IL-8 in vitro [47] (Fig. 1). Especially in deep infiltrating endometriosis, mast cells were found near nerve fibers. This led to the suggestion that mast cells might play a pivotal role in endometriosis-related pain [48].
Natural killer cells
Natural killer (NK) cells play a pivotal role at the crossroads of innate and adaptive immunity not only through their ability to lyse infected or tumor cells but also by the secretion of cytokines that contribute to direct selective adaptive responses. Published data indicate that NK cells derived from peripheral blood, characterized by markers such as CD16 and CD57, are involved in the cytotoxicity against endometrial cells in vitro [49]. The cytotoxic activity of peritoneal NK cells derived from endometriosis patients against NK-cell-sensitive K562 target cells was shown to be reduced [50, 51] (Fig. 1). The literature varies with regards to the percentages of NK cells within the peritoneal fluid among different patients. Nevertheless, the alterations in cytotoxicity seem to depend on a functional defect of NK cell activity in endometriosis rather than differences in the total cell number [50]. One explanation for such a decrease in NK cell activity might rely on the increased levels of transforming growth factor (TGF)-β [52] and PGE2 [40] in peritoneal fluid of women with endometriosis, which both inhibit NK cell activity and are derived from macrophages (Fig. 1). A second explanation for the reduced function of these cells in endometriosis involves a group of receptors referred to as killer cell inhibitory receptors (KIR). These receptors recognize major histocompatibility complex class I (MHC-I) antigens, which inhibit cytotoxicity against MHC-I-expressing target cells. Interestingly, the expression of KIRs on peritoneal NK cells of women with endometriosis is increased [53] (Fig. 1). Insights on MHC expression of endometriotic lesions are surprisingly meager; however, in a recent study using cDNA microarray technique, an up-regulation of genes encoding MHC antigens could be detected on ovarian endometrial cysts [54], which would support the notion of an escape of NK-cell-mediated cytotoxicity induced by the endometriotic lesion itself. Strikingly, in severe endometriosis, endometriotic cells are also more resistant to lysis by heterologous NK cells derived from peripheral blood [49], which further supports the immune escape theory.
Role of adaptive immune responses in endometriosis
The absolute number as well as the relative percentage of lymphocytes in peritoneal fluid of endometriosis patients have been shown to be significantly augmented [29].
T cells
Endometriotic lesions may be considered as an ‘autologous transplant’; nonetheless, one might expect T cells in the peritoneal fluid to be a candidate population in the rejection of endometric tissue. Indeed, T cells expressing markers such as CD3, CD4, and CD8 have been described to be elevated in endometriotic tissue [55, 56] (Fig. 1). RANTES (short for ‘regulated upon activation normal T cell expressed and secreted’), chemoattractant, e.g., to T cells, is expressed in ectopic endometrium [57] and elevated in the peritoneal fluid in endometriosis [58] (Fig. 1), which might explain the high number of T cells within the lesion. However, an in-depth analysis of the frequency of T cells and, in particular, their functional differentiation into cytotoxic (CD8+) or helper (Th1, Th2, or Th17 cells) cell subsets in peritoneal fluids of endometriosis patients still remains elusive.
Surprisingly, no published evidence is currently available whether or not regulatory T cells (Treg) [59] are involved in the undesired ‘immunological tolerance’ of endometriotic lesions. Autoreactive T cells are capable of encountering self-peptide/MHC complexes. Hence, the basic importance of Tregs in the maintenance of immune cell homeostasis is to suppress such autoreactive T cells. One might speculate that, in the context of endometriosis, it may be desirable to have autoreactive T cell-like populations in the peritoneal fluid, which may be capable of targeting the autologous endometriotic transplant. Hence, the presence of peritoneal Treg—which could suppress such autoagressive cells—would be undesired. However, no published data indicate the existence of autoreactive T cell-like populations in the peritoneal cavity of women without endometriosis, which could provide an explanation for the successful deletion of endometrial tissue entering the peritoneal cavity by retrograde menstruation in these women.
Natural killer T (NKT) cells are also cells that bridge innate and adaptive immune mechanisms. They produce a wide range of cytokines, mainly IL-4, driving the development of Th2-mediated responses and recognize glycolipids associated to CD1d molecules. Therefore, it appears rather puzzling that no published evidence is currently available indicating the presence or function of such cells in peritoneal fluid in the context of endometriosis.
Various authors have addressed the issue of Th1/Th2 cytokine balance in the peritoneal fluid of patients with endometriosis. Several of these cytokines may be produced by cells of the adaptive immune response, and especially the ratio of pro-inflammatory Th1-like and anti-inflammatory Th2-like cytokines has been in the center of scientific attention. In endometriosis, the production of pro-inflammatory interferon (IFN)-γ and IL-2 as well as the anti-inflammatory cytokines IL-4 and IL-10 was found to be increased in peritoneal fluids, although controversial data have been published especially with regard to IFN-γ [29, 60, 61] (Fig. 1). Th1 cytokines such as IFN-γ and IL-2 are well known to lead to the activation of T cell-mediated and delayed-type hypersensitivity (Fig. 1). On the other hand, Th2 cytokines such as IL-4 and IL-5 stimulate antibody-mediated immunity via B cell differentiation to plasma cells and inhibit Th1-mediated responses (Fig. 1). Whether endometriosis is a typical Th1- or Th2-mediated disease is not clear. In addition, future studies are warranted on the role of the recently identified pro-inflammatory Th17 cells, which secrete high levels of IL-17A, IL-17F, IL-6, and TNF-α and sustain tissue damage.
A pro-inflammatory status has been proposed to result in the down-regulation of heme oxygenases (HOs) [62] (Fig. 1). HOs are required to degrade heme into biliverdin and carbon monoxide (CO) to avoid toxic heme effects such as oxidative stress; hence, HOs have been proposed to be involved in tissue protection. Increased proliferation of endometrial tissue leads to high levels of hemoglobin (Fig. 1) as well as heme, which requires adequate HO activity to sustain tissue protection. Hemoglobin [63] as well as markers of oxidative stress such as lipid peroxides [64] are elevated in patients with endometriosis (Fig. 1). Thus, it has been suggested that the HO system might be insufficient to detoxify heme in women with endometriosis. Such insufficiency of the HO system may result from the continuous pro-inflammatory environment in the peritoneal cavity of patients with endometriosis. As a consequence of the insufficient HO system, high levels of oxidative stress may further enhance adhesion of more refluxed endometrial cells onto the peritoneum during menstruation and perpetuate the progression of endometriosis (Fig. 1).
Interestingly, recombinant human TNF-binding protein-1 reduces experimental endometriosis in rats [65], supporting the notion of an adverse effect of this pro-inflammatory cytokine. In contrast, intraperitoneal injection of the Th1-like cytokine IL-12 also reduces endometriotic lesions in vivo [66], probably by enhancing cytotoxic activity. It is, thus, not finally resolved whether pro- or anti-inflammatory responses or both of them should be suspected to contribute to the development of endometriosis.
B cells
B cells are involved in antibody-mediated adaptive immune responses. While B2 cells can give rise to classical plasma cells producing many distinct immunoglobulin (Ig) isotypes (IgG, IgA), other B cell types including B1 cells, which are present in the peritoneal cavity, and BZM cells, which are located in the marginal zone of the spleen, produce only high levels of IgM to T-independent antigens. An increase in activated CD20+ B cells could be demonstrated in ectopic endometrium [67] (Fig. 1). Soluble CD23, which may be derived from mature B cells and also from activated macrophages, eosinophils, follicular dendritic cells, and platelets, is increased in serum and peritoneal fluid of endometriosis patients, which may be suggestive of an exacerbated B cell activation in endometriosis [68] (Fig. 1). In addition, an increase in autoantibodies (AAb) such as anti-phospholipid and anti-histone IgG could be detected in the peritoneal fluid of endometriosis patients [69] (Fig. 1). Serum and cervical secretions of women with endometriosis contain organ-specific anti-endometrial and anti-ovarian specific IgG and IgA, suggesting the activation and plasma cell differentiation of B2 cells [70].
Some of the antigens from endometriotic tissue, which are recognized by serum AAb, have been identified; these include mainly glycoproteins including the human chorionic gonadotropin (hCG) receptor [71], carbonic anhydrase isoforms I and II [72, 73], transferrin, and α2-Heremans Schmidt glycoprotein (α2-HSG) [74]. Interestingly, Lang and Yeaman [73] demonstrated that removal of carbohydrate moieties from endometrial antigens prevented antibody binding via Thomsen–Friedenreich disaccharide-dependent pathways. Thomsen–Friedenreich antigens, which are expressed on epithelial cells of the uterus, bind to a variety of glycan-binding proteins including galectin (Gal)-1 and Gal-3, which are also expressed in endometrial tissue. Gal-1 and Gal-3 are evolutionarily conserved glycan-binding proteins that have been shown to contribute to cell–cell and cell–matrix interactions, cell migration, and angiogenesis [75]. In addition, galectins have been shown to modulate T cell apoptosis and immune privilege in vivo [76], thus contributing to autoimmunity by dampening antigen-specific immune responses. Because levels of AAb against Thomsen–Friedenreich antigen are significantly up-regulated in endometriosis tissue, it is possible that these AAb might block or mimic some of the biological functions of galectins. However, to date, no data are available indicating the regulated expression of Gal-1 and Gal-3 in endometriotic tissue.
In conclusion, AAb might be directly involved in the pathogenesis of endometriosis supporting an autoimmune etiology. However, AAb against Thomsen–Friedenreich antigen could also be an autoimmune epiphenomenon due to aberrantly glycosylated endometrial antigens. If the latter assumption is correct, it would be of great interest to investigate cells of the adaptive immune response with regard to the recognition of such altered self-antigens.
Adhesion molecules and protein–glycan interactions in endometriosis
Due to the prevailing inflammation in endometriosis, adhesion molecules have been suggested to be involved, e.g., by mediating the migration of leukocytes into the peritoneal cavity. In this context, published data point towards altered cell adhesion mechanisms, which might interfere with leukocyte function. The following scenarios might—at least in part—provide an explanation for the failing immune surveillance in endometriosis. First, leukocyte function antigen (LFA)-1+ effector cells interact with intercellular adhesion molecule (ICAM)-1 expressed by target cells, whereby soluble ICAM-1 (sICAM-1) may interfere with such interactions. In endometriosis, the severe decrease in NK cell-mediated lysis of endometrial cells could be manifested when endometrial supernatants contained high levels of sICAM-1 (Fig. 1), whereby it remains to be fully elucidated whether peritoneal NK cells express LFA-1 [66]. Strikingly, the expression of sICAM-1 by ectopic endometrium is increased as compared to eutopic endometrium [77], and the levels of sICAM-1 are elevated in peritoneal fluid of endometriosis patients [78] (Fig. 1). Hence, endometriotic lesions may ‘neutralize’ peritoneal LFA-1+ effector leukocytes via sICAM-1, which results in impaired immune surveillance of such effector cells.
A second putative scenario involves the glycoprotein peritoneal haptoglobin (pHp) or endometriosis protein-I (Endo-I). This glycoprotein is highly expressed in peritoneal endometriotic lesions [79] (Fig. 1) and interacts with various lectins or glycan-binding proteins [80]. Increased interaction of pHp with different lectins might be due to variations in the ratios of α(2–3) to α(2–6) sialic acid and fucose [80]. These alterations could be connected to increased expression of glycans mainly involved in carbohydrate–selectin interactions and cell–cell adhesion. Some studies report increased expression of E- and P-selectins in endometriosis [39, 81]. Because several reports are controversial [82], one might hypothesize that the function of selectins, determined by the binding to their epitopes, is more important than their quantitative levels. Sharpe-Timms et al. [83] showed that peritoneal macrophages from women with endometriosis bind more pHp in vivo than those from women without the disease, although the concentration of pHp is similar in both groups. The authors speculate that this effect may be due to altered forms of haptoglobin. Glycoproteins with increased numbers of glycans containing Sialyl-Lewis X structures are able to block adhesions of immune cells [84, 85], which may provide an explanation for the decreased ability of peritoneal macrophages to mediate cytolysis of misplaced endometrial tissues, as detectable in the peritoneal cavity (Fig. 1), which is further associated with an increased resistance of these cells to apoptosis in women with endometriosis [83].
Glycosylation and morphological changes were further investigated in a baboon model of endometriosis, and an increased lectin binding to fucosylated N-acetylglucosamine residues could be detected in early stages of the disease, whereas such binding—accompanied by a late secretory phenotype of endometrial glands—was decreased in later stages of the disease [86]. A clear evidence of an asynchrony between the estimated day of the menstrual cycle and the observed histological/ultrastructural appearance of the glands could be identified. Fucosylated N-acetylglucosamine residues are essential for the adhesion of the hatched blastocyst to the endometrium [87]. Glycosylation is important in the regulation of embryo attachment, and differential glycosylation can be regulated in the endometrium by the action of several hormones mainly during the secretory phase [88]. Therefore, one might speculate that abnormal glycosylation in endometriosis may be one of the factors leading to reduced fertility and other clinical and pathological manifestations of the disease.
Angiogenesis in endometriosis
The establishment of a new blood supply is essential for the survival of endometrium attached to the peritoneum and the maintenance of endometriosis. Increased microvessel density in endometriotic tissue with high proliferative activity has been demonstrated compared to lesions with low proliferative activity [89], which could be inhibited by blocking vascular endothelial growth factor (VEGF) [90]. High levels of VEGF and more specifically VEGF-A are present in peritoneal fluid of endometriosis patients [89] (Fig. 1). Other macrophage- or mast cell-derived factors, which either directly or indirectly induce angiogenesis and are up-regulated in endometriosis, include IL-1, IL-6, TNF-α, and TGF-β [91–93] (Fig. 1). In addition, IL-8 is increased in peritoneal fluid in endometriosis [94] and contributes to angiogenesis [95] (Fig. 1). Finally, matrix metalloproteinases (MMP) are involved in tissue remodeling and angiogenesis. The expression of MMP-1 by endometriotic cells is increased as compared to eutopic endometrium of patients and controls [96], and levels of MMP-2 were found to be elevated in peritoneal fluid of endometriosis patients [97] (Fig. 1). In contrast, another study reported MMP-1 and MMP-2 to be highly expressed in eutopic but not ectopic endometrium of women with endometriosis and a decrease in the MMP-2 inhibitor (TIMP-2) [98], suggesting a role of MMP in the very early development of the disease.
It may be concluded that immune cells in the peritoneal fluid and probably within the lesions do not only fail to reject endometriotic lesions but also support their growth by promoting vascularization and thus nourishing of the tissue with their secreted factors such as cytokines.
The role of steroid hormones in endometriosis
Circumstantial and laboratory evidences are indicative of critical roles of steroid hormones in the establishment and maintenance of endometriosis. A lower incidence of the disease is noted in women with decreased endogenous estrogen production due to extensive exercise or smoking [99]. Although the development of endometriosis has historically been viewed as an estrogen-dependent disease, recent studies suggest that a failure of progesterone to appropriately regulate the expression of genes during endometrial differentiation might be a critical component of the disease process. It is well accepted that progesterone inhibits estradiol (E2)-dependent proliferation in the uterine epithelium. In humans and other vertebrates, the biological activities of progesterone are mediated by interaction with specific progesterone receptors (PRs) that are members of nuclear receptor superfamily of transcription factors. In contrast to healthy endometrium, hormone receptors remain at constant levels during the menstrual cycle in endometriosis [100], which points at a disruption of the normal cyclic responsiveness to hormonal changes with regular apoptosis of the epithelial stratum functionalis in endometriotic lesions.
The following molecular observations support a role of progesterone in endometriosis. First, a PR gene polymorphism was reported to be associated with endometriosis [101] (Fig. 2b). Second, only low levels of PR isoform A (PR-A) and no PR-B are detectable in extraovarian endometriosis [102] (Fig. 2b). Third, progesterone-dependent regulation of target genes was found to be perturbed in endometriosis. In the normal secretory endometrium, progesterone indirectly induces the 17β-hydroxysteroid dehydrogenase type 2 (17β-HSD-2), which converts estradiol (E2) to estrone (E1) [103] (Fig. 2a). In endometriotic tissue, 17β-HSD-2 is undetectable [104] (Fig 2b). As a consequence, E2 accumulates and probably induces proliferation of endometrial tissue. Moreover, the enzyme aromatase is exclusively found in endometriotic stroma [105] and provides E1, which is further converted to E2 by 17β-HSD type 1 (17β-HSD-1), contributing to the accumulation of E2 (Fig. 2b). The importance of aromatase for the endometriotic growth has been demonstrated by its genetic or enzymatic disruption [106].
Fig. 2.
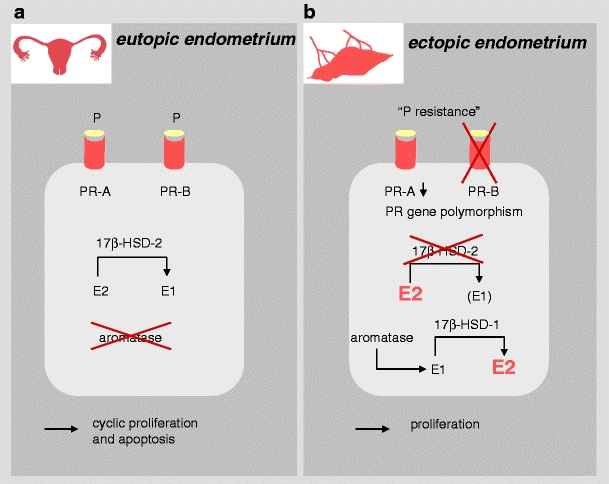
Progesterone resistance and estradiol accumulation in endometriosis. E1 estrone, E2 estradiol, P progesterone, PR progesterone receptor, 17β-HSD-1 17β-hydroxysteriod dehydrogenase type 1, 17β-HSD-2 17β-hydroxysteriod dehydrogenase type 2. a In normal endometrium, P actions are mediated by PR; its isoforms PR-A and PR-B are both expressed on the tissue. P indirectly induces 17β-HSD-2, which converts E2 to E1; aromatase is not expressed. Tissue proliferation is followed by cyclic apoptosis. b In endometriotic tissue, only low expression of PR-A and the lack of PR-B have been observed, which leads to a partial resistance to P actions. Additionally, a PR gene polymorphism has been described to be more frequent in patients with endometriosis, which may be additionally involved in the partial resistance to P actions. Low levels of 17β-HSD-2 along with the enzymatic activity of aromatase—which is exclusively found in endometriotic stroma—lead to an accumulation of E2. Hence, physiological cyclic regulation of proliferation and apoptosis is disrupted in endometriosis
Besides, E2 might also be involved in endometriosis-related inflammation, as it stimulates several mediators of inflammation such as IL-8 and RANTES [57, 107] (Fig. 1). Levels of MMP-2 in peritoneal fluid positively correlate with E2, but inversely with progesterone [97], and estrogens can also co-stimulate peritoneal mast cell activation in rats [108] (Fig. 1). Further, both E2 and progesterone increase the release of VEGF from peritoneal macrophages [109] and may hereby promote angiogenesis in endometriotic lesions. Given the high levels of E2 within the lesion, E2 might be more relevant in this context.
In contrast to estrogens, progesterone or synthetically produced progestogens called progestins exert potent immunosuppressive properties, and mechanisms of dampening inflammation have been demonstrated for the progestin dydrogesterone [110–112]. Using other progestins available to date, a reduction in transplant growth, a degeneration of transplants, an increase in NK cells in peritoneal fluid, and a decrease in IL-1β secretion from peritoneal macrophages could be demonstrated [113–115] (Fig. 1). It would be interesting to investigate the cross talk between hormones and regulatory T cells in this process.
One might conclude from these observations that E2 promotes endometriosis and progesterone/progestins dampen it, which is the rationale why progestins are widely used to treat endometriosis. In general, they act as agonists on the PR and have a functional anti-proliferative effect on endometrial tissue and endometriotic lesions. In addition, progestin treatment is generally better tolerated than other endometriosis therapies, which are associated with rather severe side effects [116]. Importantly, progestins are known to be effective in the control of pain symptoms in general or related to endometriosis [112, 116, 117]. However, not all patients respond to this regimen [118]. This clinical observation may be explicable by the partial resistance of endometriosis tissue to progesterone action due to the down-regulation of its receptors, as outlined earlier.
Neurological aspects in endometriosis
The female reproductive organs are innervated by sympathetic, parasympathetic, and sensory afferent nerves. Their distribution and the immunoreactivity to various peptides point at a role in hemodynamics and smooth muscle contraction. Uteri of patients with advanced endometriosis—similar to chronic pelvic pain—show increased numbers of nerve fibers and perivascular nerve fiber proliferation [119]. In addition, small nerve fibers were detected in the functional layer of the eutopic endometrium in endometriosis patients but not in controls [120].
Compared to normal peritoneum, nerve fibers in human peritoneal endometriotic lesions are increased [121]. In addition, artificially induced endometriotic cysts in rats are robustly innervated by fibers accompanying blood vessels and extending in patterns similar to the uterus [122]. Because endometriotic tissue may be considered as a ‘vascularized autologous transplant’, its innervation has been suggested to occur via sprouting of para- and perivascular nerve fibers. This coordination of vascularization and innervation has also been observed in other transplanted organs such as the parathyroid glands [123] and skin [124]. Immunostaining of endometriotic tissue revealed calcitonin-gene-related peptide (CGRP)+ and substance P (SP)+ sensory C and Aδ [121, 122], acetylcholine (Ach)+ parasympathetic as well as tyrosine hydroxylase (TH)+ sympathetic [121] fibers (Fig. 1). CGRP+ and SP+ fibers even reach the epithelium of the ectopic tissue [122]. Endometriosis-related pain might be mediated by these pro-inflammatory neuroactive agents. They are increased by nerve growth factor (NGF) [125, 126], which also sensitizes terminals of sensory nerve fibers [127]. Interestingly, intense immunoreactivity for NGF and its receptor NGFRp75 was demonstrated near endometriotic glands [121] (Fig. 1). Whether and how this might be connected with immunological alterations in endometriosis will be discussed in the course of the following section.
Stress perception, endometriosis, and sickness response
As previously outlined, cardinal symptoms of endometriosis are chronic pain and infertility, which may severely interfere with the patient’s quality of life and be perceived as a persistent stressor. To date, it is well established that high perception of stress may trigger or aggravate the incidence or exacerbation of diseases such as inflammatory bowel disease [128, 129], immune dermatoses [130, 131], or pregnancy complications such as spontaneous abortion and pre-eclampsia [132]. Further, high levels of perceived stress have been proposed to contribute to the progression of endometriosis [133].
Nowadays, it is well established that high levels of perceived stress activate the release of neurohormones such as corticotropin-releasing hormone (CRH) as well as adrenocorticotropic hormone (ACTH) and glucocorticoids (GCs) largely via the hypothalamus–pituitary–adrenal (HPA) axis [134–136]. Via these stress-related hormones, accompanied by additional stress response mediators like neuropeptides or neurotrophins [137, 138], immune responses are profoundly altered [135, 136]. For example, GCs dampen the release of IL-12, IFN-γ, and TNF-α by antigen-presenting cells and Th1 cells, but up-regulate the production of IL-4, IL-10, and IL-13 by Th2 cells [139, 140]. Thus, it has been proposed that the release of CRH, ACTH, and subsequently GCs induces the selective suppression of the Th1-mediated cellular immunity and elicits a skew in the direction of Th2-dominated immunity. Indeed, it has also been postulated that such Th2 shift may actually protect the organism from systemic surpassing of pro-inflammatory cytokines, which could have severe tissue-injuring features [141].
However, besides such well-described immunosuppressive effects of GCs, relevant examples of pro-inflammatory actions of peripheral CRH have recently been introduced, e.g., in the synovia of rheumatoid arthritis patients [142]. This peripheral CRH leads to proliferation of immunocytes [143, 144] and production of IL-1, IL-2, and IL-6 [145, 146]. Moreover, CRH antagonizes some of the immunoregulatory effects of serotonin (5-HT), e.g., the suppression of TNF-α production [147]. Inflammation can be dampened by blocking CRH at the periphery [148], as CRH receptors occur on immune cells, e.g., on T cells, monocytes [149], and mast cells [150]. Peripheral CRH may, thus, be referred to as “immune” CRH. Immune cells such as T cells, B cells [151], and mast cells [152] may be the source of peripheral CRH. Besides, CRH-like immunoreactivity was demonstrated in splenic nerve fibers [153], in the dorsal horn of the spinal cord, and dorsal root ganglia as well as sympathetic nerve fibers [154] pointing at a neuronal source of “immune” CRH in descending nerve fibers.
Intriguingly, CRH is highly expressed in endometriotic lesions [44] and has been proposed to stimulate mast cells to secrete VEGF [150] (Fig. 1). This would facilitate angiogenesis in endometriotic lesions and perpetuate the dissemination of the disease. Thus, it may be proposed that high levels of perceived stress promote the dissemination of endometriosis via CRH-dependent pathways, as peripheral CRH is found increased in response to psychological stress [155]. It remains to be elucidated whether and how CRH can be made attributable for the peritoneal inflammation present in the peritoneal cavity of patients suffering from endometriosis. In contrast to endometriotic tissue, CRH levels in peritoneal fluid of endometriosis patients were not different from controls; however, CRH-binding protein (CRH-BP) is increased in advanced stages of endometriosis, compared to early stages or healthy controls [156], which may be interpreted as an attempt of the body to down-regulate high CRH levels.
Neurohormonal responses to stress also include an activation of the sympathetic nervous system with the subsequent increase in catecholamines [137], and published evidence strongly supports that the immune response can be regulated via the sympathetic nervous system/catecholamines at regional, local, and systemic levels [134]. For example, lymphocytes express adrenergic receptors and respond to catecholamine stimulation with the development of stress-induced lymphocytosis, inflammation, and distinct changes in lymphocyte trafficking, circulation, proliferation, and cytokine production [139, 157, 158]. Because sympathetic nerve fibers are present in endometriotic tissue, it appears likely that—besides CRH—catecholamines contribute to the peritoneal inflammation in endometriosis, and an increased release of catecholamines in response to high levels of stress could additionally perpetuate local inflammation.
Besides the classical stress-related neurohormones—the players of the HPA axis and catecholamines—NGF and the neuropeptide SP are now recognized as pivotal mediators of the stress responses [131, 159–161]. SP co-functions as neurotransmitter in the central and peripheral nervous system in pain, anxiety, and emotional centers besides stress [131]. Additionally, SP is a potent pro-inflammatory mediator, as it induces the production and release of cytokines such as IL-1, IL-6, and TNF-α and NGF [162, 163] and promotes angiogenesis during inflammation [164]. NGF also enhances pro-inflammation and has been proposed to increase the release of SP [161, 165, 166]. SP and NGF derived from nerve endings induce leukocyte recruitment and release of pro-inflammatory cytokines, which is generally referred to as neurogenic inflammation. Vice versa, pro-inflammatory cytokines may induce the expression of neuropeptides such as SP and NGF and their cognate receptors [167, 168], which might provide an explanation for the up-regulation of NGF expression in inflamed areas, e.g., in peritoneal endometriosis [121]. Interestingly, the activation of PAR-2 on afferent neurons, e.g., by proteases such as mast cell tryptase, leads to subsequent release of pro-inflammatory CGRP and SP [169], which might provide explanation for pain during inflammation. To date, published evidence on the expression of SP in endometriosis is very limited; some authors report no changes of SP in endometriosis, which could be due to the difficulties in detecting SP because it is a rather small peptide of no more than 11 amino acids [170]. Hence, future studies employing novel technologies such as gene or protein arrays may allow to investigate the expression of pain-related neurotransmitters as well as their receptors, e.g., SP high affinity receptor neurokinin (NK)-1R [112] on endometrial tissue. However, as outlined earlier, Tokushige et al. [121] recently identified an increased number of nerve fibers in peritoneal endometriotic lesions using immunohistochemistry; in fact, these nerve fibers were immunoreactive for SP, CGRP, Ach, NGF, and TH (Fig. 1).
In conclusion, a hypothetical scenario on how high stress perception might be related to endometriosis is depicted in Fig. 3. It may be postulated, in terms of a “brain-body cross talk”, that psychologically stressful conditions promote the aggravation of inflammatory processes, angiogenesis, pain, and infertility via pathways that most likely involve catecholamines, “immune” CRH, NGF, SP, and CGRP (Fig. 3 right), whereby the dependancy between NGF/SP and CRH should be addressed in future experimental settings.
Fig. 3.
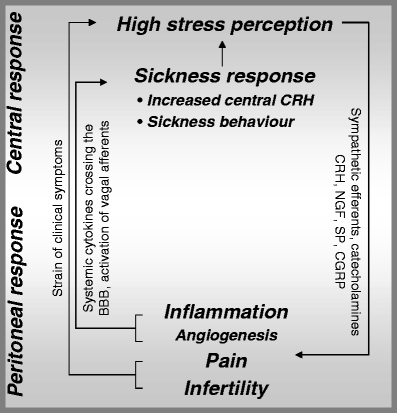
Hypothetical scenario for endometriosis: the ‘brain–body–brain cross talk’. BBB blood–brain barrier, CGRP calcitonin-gene-related peptide, CRH corticotropin-releasing hormone, NGF nerve growth factor, SP substance P. Perceived stress aggravates peritoneal inflammation and angiogenesis, chronic pain, and infertility in patients with endometriosis via neuronal pathways involving catecholamines, CRH, NGF, SP, and CGRP; this may be referred to as ‘brain–body cross talk’ because it involves central stress response pathways. Clearly, the aspect of infertility—a frequent clinical symptom in patients with endometriosis—may be perceived as an additional stressor besides stressors such as daily hassles. Peripheral ‘inflammatory stress’—as described in endometriosis—and the strain of clinical symptoms such as chronic pain may in turn induce a sickness response, which can be entitled ‘body–brain cross talk’. Such ‘body–brain cross talk’ may subsequently perpetuate stress perception and trigger the release of central CRH and the onset of behavioral alterations, i.e., sickness behavior, via elevated circulating pro-inflammatory cytokines bypassing/crossing the BBB or via stimulation of vagal afferents by peritoneal pro-inflammatory cytokines. As a result, a vicious circle of ‘brain–body–brain cross talk’ is closed in patients suffering from endometriosis
Peripheral immunological events in turn signal certain brain regions to induce the so-called sickness response including (neuro)hormonal and behavioral changes [171]. These profound behavioral alterations are referred to as sickness behavior, which comprises depressive-like behavior and is characterized by reduced locomotor activity, fatigue, hypophagia or anorexia, diminished social interactions, and female sexual behavior [171–173]. Interestingly, such sickness behavior can be experimentally provoked by peripheral administration of IL-1 or TNF-α [172, 173]. In healthy humans, levels of anxiety, depressed mood, and memory function correlate with levels of TNF-α, its soluble receptors, and IL-6 [174]. Brain regions identified to date comprise structures associated with the central stress system, namely, the hypothalamus, hippocampus, frontal/prefrontal structures, and the limbic system including the amygdala [175].
Intravenous as well as intraperitoneal applications of IL-1 result in an increased CRH production in the hypothalamus [176, 177]. Because elevated levels of central CRH have been associated to the abovementioned behavioral changes during sickness [178], non-cognitive ‘inflammatory stress’ can induce behavioral and emotional alterations via increase in central CRH. As mentioned earlier in this review, IL-1, IL-6, and TNF-α are elevated in peritoneal fluid of endometriosis patients. Evidently, due to the up-regulation of pro-inflammatory cytokines not only in the peritoneal fluid but also in the peripheral blood of patients with endometriosis [179], this ‘inflammatory stress’ may increase stress perception in affected patients due to the onset of the sickness response, which includes an increase in central CRH as well as behavioral changes and which would aggravate the strain generated by symptoms such as pain and infertility in terms of a ‘body–brain cross talk’ (Fig. 3, left).
Undoubtedly, the question that arises is how peripheral ‘inflammatory stress’ can reach the central nervous system. Here, it has been proposed that peripheral blood cytokines bypass the blood–brain barrier (BBB) entering in the circumventricular organs [180]. Secondly, these cytokines might cross the BBB, especially when its permeability is enhanced, e.g., in stressful conditions [181]. Thus, perceived stress might predispose certain individuals to develop sickness behavior during illness. Finally, vagal nervous afferents might be involved, as suggested by experiments in rats where behavior has been investigated before and after vagotomy and upon intraperitoneal application of IL-1β [182], and IL-1β-induced inhibition of social exploration was attenuated in vagotomized rats. Because Ach+ nerve fibers were found in endometriotic tissue, as illustrated above, behavioral changes mediated by vagal afferents might play a relevant role in endometriosis (Fig. 3, left).
In conclusion, dissemination of endometriosis may be sustained and advanced by high levels of perceived stress accompanying neuroendocrine-immune disequilibrium in affected patients. This, in turn, fosters additional stressors such as pain and infertility and induces sickness response. As a result, stress perception is even higher, which induces a vicious cycle of ‘brain–body–brain cross talk’ in affected women (Fig. 3).
Animal models of endometriosis
As evident from the previous paragraphs, an in-depth knowledge on the pathophysiology of endometriosis is limited, mostly due to the difficulty in studying the disease in the human [(a) need for laparoscopy, (b) painful lesions are often very small in size and invisible from the peritoneal cavity, and (c) high individual patient variation]. To overcome this gap, convincing animal models are needed in addition to clinical studies. Endometriosis is a primate-specific disease, but primate studies can only be performed adequately in a limited number of laboratories [183] and are rather time-consuming. To date, the rat is one of the most thoroughly studied laboratory animal species, and in spite of its phylogenetic distance to the human, surgically generated transplants of uterine fragments at different sites of the peritoneal cavity have been shown to resemble human endometriotic lesions in many respects [184]. Advantages and disadvantages of such rat models have been vividly discussed, e.g., the absence of menstruation into the peritoneal cavity in rats and consequently the lack of retrograde menstruation as one of the key prerequisites for the development of endometriosis. On the other hand, rat endometrium undergoes hormone-dependent tissue remodeling; surgically transplanted uterine tissue in the rat mirrors human endometriotic lesions, and endometrial lesions in rats are responsive to steroid treatment. The benefits of using the rat model are also supported by observations that, in rats with surgically induced endometriosis—similar to humans—fertility and fecundity are also impaired [41, 185].
Similar to rats, mice do not menstruate and—similar to rats—murine endometrium undergoes hormone-dependent tissue remodeling. Further, mouse models are commonly used in immunological research. Hence, based on the growing evidence of a failing immune surveillance in endometriosis, the need for a murine mouse model to study immunological aspects of endometriosis is becoming more and more evident. To date, a few mouse models have already been introduced and are summarized in Table 1. In some of these models, human or murine endometrium is injected into the peritoneal cavity [66, 186, 187] or fixed on the mesentery or the peritoneum [188, 189] of either intact or ovariectomized mice of different strains. In other murine models, human endometrioma tissue was either injected subcutaneously in nude mice [190] or injection is performed upon transducing whole fragments of human endometrium in vitro by adenoviral infection with the green fluorescent protein (GFP) cDNA before transplantation into nude mice. Implantation and growth of endometriotic-like lesions could be here non-invasively followed and repeatedly documented by in vivo imaging [191]. Such rodent models may provide tools for drug testing and/or gene target validation in endometriosis.
Table 1.
Recently published mouse models of endometriosis
| Mouse strain | Ovariectomy | Initial tissue placement | Reference # |
|---|---|---|---|
| ncr/nude (athymic) | + | Subcutaneous injection of human endometrioma biopsies | [190] |
| C57BL/6 Balb/c | + | Intraperitoneal inoculation of murine endometrial fragments | [66] |
| ncr/nude | −/+ | Intraperitoneal injection of human endometrioma biopsies | [186] |
| Progesterone receptor −/− | + | Intraperitoneal fixation of murine endometrium on mesentery | [188] |
| C57BL/6 | − | Intraperitoneal fixation of murine endometrial punches on peritoneum and mesentery | [189] |
| C57BL/6 | + | Intraperitoneal injection of GFP-labeled murine endometrial fragments | [187] |
GFP Green fluorescent protein, PR progesterone receptor
Clinical considerations
Current approaches to therapy for endometriosis are surgical, directed at resecting endometriotic tissue, and pharmacologically, aiming at inhibiting estrogenic stimulation, thus reducing dissemination and disease burden. However, despite such therapeutical approaches, endometriosis is still a progressive disease, and recurrences are observed in up to 74.4% after medical treatment depending on the stage of disease [192].
Hormonal therapeutic approaches currently used for amelioration of endometriosis-associated pelvic pain include GnRH agonists and daily oral contraceptives used in a cyclic or continuous regimen [118]. Because these interventions inhibit ovulation and interfere with the cyclic remodeling of the endometrium, the onset of pregnancy is impossible, and one potential stressor of endometriosis, the infertility, is not targeted. In line with such hormonal approach in endometriosis treatment, clinical studies suggest an apparent resolution of endometriosis-related symptoms during pregnancy, supporting the rationale of treatments aiming at a pseudo-pregnancy regime [193]. As mentioned earlier in this review, published evidence suggests that progestins are effective therapies in the treatment of pain symptoms associated with endometriosis and ameliorate symptoms of endometriosis, as the pro-inflammatory setting in the peritoneal cavity may be dampened and pain receptors such as the NK1 receptor are down-regulated [112, 194]. Clearly, the future holds promise for highly specific therapeutic agents aiming at targeting the pro-inflammatory setting in endometriosis. Such approaches may include aromatase inhibitors, progestins, and progesterone receptor modulators. Therapeutic approaches aiming at diminishing inflammation also include cytokine antagonists such as the aforementioned application of recombinant human TNF-α-binding protein-1 [65]. Further, MMP inhibitors and mast cell stabilizers such as the flavonoid quercetin may have the potential to ameliorate symptoms in endometriosis, as they have been shown to inhibit the release of pro-inflammatory mediators [195]. Interestingly, progesterone is known to suppress MMPs and to enhance the expression of MMP inhibitors (TIMPs) during endometrial differentiation [196]. Hence, a combinational therapy of progestins and MMP inhibitors may be envisioned to be highly effective in endometriosis.
Conclusions
Profound and comprehensive insights on neuroendocrine-immunological disequilibrium are required to fully understand the pathophysiology of endometriosis and to develop primary and secondary prevention strategies. Clearly, multidisciplinary research activities are needed, and future basic science and clinical trials must refrain from analyses of single pathways, but aim at identifying currently known key protagonists in endometriosis. In this regard, markers and mediators introduced and summarized in the present review may just reflect the often quoted ‘tip of the iceberg’, but foster vivid scientific discussions and mutual research endeavors between basic scientists and physicians aiming at successfully targeting symptoms of this complex and multifactorial disease that affects up to 22% of women in their reproductive age. Obviously, the effect of psychosocial counseling or psychotherapy of patients with endometriosis—which has noticeably been underrated in previous studies—may be beneficial because an improved stress management may disrupt the vicious cycle of cross talk between peritoneal inflammation, sickness response, pain perception, and stress.
Acknowledgment
The writing of this review and the authors’ own experimental work cited herein were made possible by grants provided by the German Research Foundation and the Charité to PCA. NT, SMB, and PCA are part of the EMBIC Network of Excellence, co-financed by the European Commission throughout the FP6 framework program ‘Life Science, Genomics and Biotechnology for Health. NT is a scholar of the German National Merit Foundation, and SMB is supported by the Habilitation Program of the Charité. GAR is a member of the research career of CONICET and fellow of the John Simon Guggenheim Memorial Foundation. The authors are grateful for the support provided by Sarah Tariverdian to improve the quality of the graphics.
References
- 1.Rokitansky K (1860) Über Uterusdrüsen-Neubildung. Z Gesellschaft Aerzte (Wien) 16:577–581
- 2.Moen MH, Muus KM (1991) Endometriosis in pregnant and non-pregnant women at tubal sterilization. Hum Reprod 6:699–702 [DOI] [PubMed]
- 3.Koninckx PR, Meuleman C, Demeyere S, Lesaffre E, Cornillie FJ (1991) Suggestive evidence that pelvic endometriosis is a progressive disease, whereas deeply infiltrating endometriosis is associated with pelvic pain. Fertil Steril 55:759–765 [DOI] [PubMed]
- 4.Bruckner C (2004) Symptoms and diagnosis. In: Steck T, Felberbaum R, Küpker W, Bruckner C, Finas D (eds) Endometriosis. Development, diagnosis, progression, therapy. Springer, New York, pp 81–101
- 5.Steck T (2004) Manifestations and progression. In: Steck T, Felberbaum R, Küpker W, Bruckner C, Finas D (eds) Endometriosis. Development, diagnosis, progression, therapy. Springer, Berlin Heidelberg New York, pp 37–79
- 6.Redwine DB (1999) Ovarian endometriosis: a marker for more extensive pelvic and intestinal disease. Fertil Steril 72:310–315 [DOI] [PubMed]
- 7.ASRM (1997) Revised American Society for Reproductive Medicine classification of endometriosis: 1996. Fertil Steril 67:817–821 [DOI] [PubMed]
- 8.Tummon IS, Maclin VM, Radwanska E, Binor Z, Dmowski WP (1988) Occult ovulatory dysfunction in women with minimal endometriosis or unexplained infertility. Fertil Steril 50:716–720 [PubMed]
- 9.Harlow CR, Cahill DJ, Maile LA, Talbot WM, Mears J, Wardle PG, Hull MG (1996) Reduced preovulatory granulosa cell steroidogenesis in women with endometriosis. J Clin Endocrinol Metab 81:426–429 [DOI] [PubMed]
- 10.Suginami H, Yano K, Watanabe K, Matsuura S (1986) A factor inhibiting ovum capture by the oviductal fimbriae present in endometriosis peritoneal fluid. Fertil Steril 46:1140–1146 [DOI] [PubMed]
- 11.Jha P, Farooq A, Agarwal N, Buckshee K (1996) In vitro sperm phagocytosis by human peritoneal macrophages in endometriosis-associated infertility. Am J Reprod Immunol 36:235–237 [DOI] [PubMed]
- 12.Haney AF, Misukonis MA, Weinberg JB (1983) Macrophages and infertility: oviductal macrophages as potential mediators of infertility. Fertil Steril 39:310–315 [DOI] [PubMed]
- 13.Nip MM, Taylor PV, Rutherford AJ, Hancock KW (1995) Autoantibodies and antisperm antibodies in sera and follicular fluids of infertile patients; relation to reproductive outcome after in-vitro fertilization. Hum Reprod 10:2564–2569 [DOI] [PubMed]
- 14.Aeby TC, Huang T, Nakayama RT (1996) The effect of peritoneal fluid from patients with endometriosis on human sperm function in vitro. Am J Obstet Gynecol 174:1779–1783 (discussion 1783–1785) [DOI] [PubMed]
- 15.Qiao J, Yeung WS, Yao YQ, Ho PC (1998) The effects of follicular fluid from patients with different indications for IVF treatment on the binding of human spermatozoa to the zona pellucida. Hum Reprod 13:128–131 [DOI] [PubMed]
- 16.Damewood MD, Hesla JS, Schlaff WD, Hubbard M, Gearhart JD, Rock JA (1990) Effect of serum from patients with minimal to mild endometriosis on mouse embryo development in vitro. Fertil Steril 54:917–920 [DOI] [PubMed]
- 17.Matson PL, Yovich JL (1986) The treatment of infertility associated with endometriosis by in vitro fertilization. Fertil Steril 46:432–434 [DOI] [PubMed]
- 18.Sampson J (1927) Peritoneal endometriosis due to the menstrual dissemination of endometrial tissue into the peritoneal cavity. Am J Obstet Gynecol 14:422
- 19.Meyer R (1919) Über den Stand der Frage der Adenomyositis und Adenomyome im allgemeinen und insbesondere über Adenomyositis seroepithelialis und Adenomyometritis sarcomatosa. Zentralbl Gynäkol 43:745–750
- 20.Quinn M (2004) Endometriosis: the consequence of neurological dysfunction? Med Hypotheses 63:602–608 [DOI] [PubMed]
- 21.Chatman DL (1981) Pelvic peritoneal defects and endometriosis: Allen-Masters syndrome revisited. Fertil Steril 36:751–756 [DOI] [PubMed]
- 22.Rier SE, Martin DC, Bowman RE, Dmowski WP, Becker JL (1993) Endometriosis in rhesus monkeys (Macaca mulatta) following chronic exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Fundam Appl Toxicol 21:433–441 [DOI] [PubMed]
- 23.Moen MH, Magnus P (1993) The familial risk of endometriosis. Acta Obstet Gynecol Scand 72:560–564 [DOI] [PubMed]
- 24.Giudice LC, Kao LC (2004) Endometriosis. Lancet 364:1789–1799 [DOI] [PubMed]
- 25.Kao LC, Germeyer A, Tulac S, Lobo S, Yang JP, Taylor RN, Osteen K, Lessey BA, Giudice LC (2003) Expression profiling of endometrium from women with endometriosis reveals candidate genes for disease-based implantation failure and infertility. Endocrinology 144:2870–2881 [DOI] [PubMed]
- 26.Hu WP, Tay SK, Zhao Y (2006) Endometriosis-specific genes identified by real-time reverse transcription-polymerase chain reaction expression profiling of endometriosis versus autologous uterine endometrium. J Clin Endocrinol Metab 91:228–238 [DOI] [PubMed]
- 27.Haney AF, Muscato JJ, Weinberg JB (1981) Peritoneal fluid cell populations in infertility patients. Fertil Steril 35:696–698 [DOI] [PubMed]
- 28.D’Hooghe TM, Hill JA, Oosterlynck DJ, Koninckx PR, Bambra CS (1996) Effect of endometriosis on white blood cell subpopulations in peripheral blood and peritoneal fluid of baboons. Hum Reprod 11:1736–1740 [DOI] [PubMed]
- 29.Sukhikh GT, Sotnikova NY, Antsiferova YS, Posiseeva LV, Veryasov VN, Van’ko LV (2004) Cytokine production by immunocompetent cells of peritoneal fluid in women with external genital endometriosis. Bull Exp Biol Med 137:568–571 [DOI] [PubMed]
- 30.Halme J, Becker S, Hammond MG, Raj MH, Raj S (1983) Increased activation of pelvic macrophages in infertile women with mild endometriosis. Am J Obstet Gynecol 145:333–337 [DOI] [PubMed]
- 31.Fukaya T, Sugawara J, Yoshida H, Yajima A (1994) The role of macrophage colony stimulating factor in the peritoneal fluid in infertile patients with endometriosis. Tohoku J Exp Med 172:221–226 [DOI] [PubMed]
- 32.Akoum A, Lemay A, McColl S, Turcot-Lemay L, Maheux R (1996) Elevated concentration and biologic activity of monocyte chemotactic protein-1 in the peritoneal fluid of patients with endometriosis. Fertil Steril 66:17–23 [PubMed]
- 33.Keenan JA, Chen TT, Chadwell NL, Torry DS, Caudle MR (1994) Interferon-gamma (IFN-gamma) and interleukin-6 (IL-6) in peritoneal fluid and macrophage-conditioned media of women with endometriosis. Am J Reprod Immunol 32:180–183 [DOI] [PubMed]
- 34.Keenan JA, Chen TT, Chadwell NL, Torry DS, Caudle MR (1995) IL-1 beta, TNF-alpha, and IL-2 in peritoneal fluid and macrophage-conditioned media of women with endometriosis. Am J Reprod Immunol 34:381–385 [DOI] [PubMed]
- 35.Beliard A, Noel A, Goffin F, Frankenne F, Foidart JM (2003) Adhesion of endometrial cells labeled with 111Indium-tropolonate to peritoneum: a novel in vitro model to study endometriosis. Fertil Steril 79(Suppl 1):724–729 [DOI] [PubMed]
- 36.Taketani Y, Kuo TM, Mizuno M (1992) Comparison of cytokine levels and embryo toxicity in peritoneal fluid in infertile women with untreated or treated endometriosis. Am J Obstet Gynecol 167:265–270 [DOI] [PubMed]
- 37.Gomez-Torres MJ, Acien P, Campos A, Velasco I (2002) Embryotoxicity of peritoneal fluid in women with endometriosis. Its relation with cytokines and lymphocyte populations. Hum Reprod 17:777–781 [DOI] [PubMed]
- 38.Iwabe T, Harada T, Tsudo T, Nagano Y, Yoshida S, Tanikawa M, Terakawa N (2000) Tumor necrosis factor-alpha promotes proliferation of endometriotic stromal cells by inducing interleukin-8 gene and protein expression. J Clin Endocrinol Metab 85:824–829 [DOI] [PubMed]
- 39.Rojas-Cartagena C, Appleyard CB, Santiago OI, Flores I (2005) Experimental intestinal endometriosis is characterized by increased levels of soluble TNFRSF1B and downregulation of Tnfrsf1a and Tnfrsf1b gene expression. Biol Reprod 73:1211–1218 [DOI] [PubMed]
- 40.Morita M, Yano Y, Otaka K, Kojima E, Momose K (1990) Minimal and mild endometriosis. Nd:YAG laser treatment and changes in prostaglandin concentrations in peritoneal fluid. J Reprod Med 35:621–624 [PubMed]
- 41.Golan A, Dargenio R, Winston RM (1986) The effect of treatment on experimentally produced endometrial peritoneal implants. Fertil Steril 46:954–958 [DOI] [PubMed]
- 42.Kusume T, Maeda N, Izumiya C, Yamamoto Y, Hayashi K, Oguri H, Nishimori Y, Fukaya T (2005) Human leukocyte antigen expression by peritoneal macrophages from women with pelvic endometriosis is depressed but coordinated with costimulatory molecule expression. Fertil Steril 83(Suppl 1):1232–1240 [DOI] [PubMed]
- 43.Braun DP, Gebel H, Rotman C, Rana N, Dmowski WP (1992) The development of cytotoxicity in peritoneal macrophages from women with endometriosis. Fertil Steril 57:1203–1210 [PubMed]
- 44.Kempuraj D, Papadopoulou N, Stanford EJ, Christodoulou S, Madhappan B, Sant GR, Solage K, Adams T, Theoharides TC (2004) Increased numbers of activated mast cells in endometriosis lesions positive for corticotropin-releasing hormone and urocortin. Am J Reprod Immunol 52:267–275 [DOI] [PubMed]
- 45.Osuga Y, Koga K, Tsutsumi O, Igarashi T, Okagaki R, Takai Y, Matsumi H, Hiroi H, Fujiwara T, Momoeda M, Yano T, Taketani Y (2000) Stem cell factor (SCF) concentrations in peritoneal fluid of women with or without endometriosis. Am J Reprod Immunol 44:231–235 [DOI] [PubMed]
- 46.Vliagoftis H, Worobec AS, Metcalfe DD (1997) The protooncogene c-kit and c-kit ligand in human disease. J Allergy Clin Immunol 100:435–440 [DOI] [PubMed]
- 47.Hirota Y, Osuga Y, Hirata T, Harada M, Morimoto C, Yoshino O, Koga K, Yano T, Tsutsumi O, Taketani Y (2005) Activation of protease-activated receptor 2 stimulates proliferation and interleukin (IL)-6 and IL-8 secretion of endometriotic stromal cells. Hum Reprod 20:3547–3553 [DOI] [PubMed]
- 48.Anaf V, Chapron C, El Nakadi I, De Moor V, Simonart T, Noel JC (2006) Pain, mast cells, and nerves in peritoneal, ovarian, and deep infiltrating endometriosis. Fertil Steril 86:1336–1343 [DOI] [PubMed]
- 49.Oosterlynck DJ, Cornillie FJ, Waer M, Vandeputte M, Koninckx PR (1991) Women with endometriosis show a defect in natural killer activity resulting in a decreased cytotoxicity to autologous endometrium. Fertil Steril 56:45–51 [DOI] [PubMed]
- 50.Oosterlynck DJ, Meuleman C, Waer M, Vandeputte M, Koninckx PR (1992) The natural killer activity of peritoneal fluid lymphocytes is decreased in women with endometriosis. Fertil Steril 58:290–295 [DOI] [PubMed]
- 51.Iwasaki K, Makino T, Maruyama T, Matsubayashi H, Nozawa S, Yokokura T (1993) Leukocyte subpopulations and natural killer activity in endometriosis. Int J Fertil Menopausal Stud 38:229–234 [PubMed]
- 52.Oosterlynck DJ, Meuleman C, Waer M, Koninckx PR (1994) Transforming growth factor-beta activity is increased in peritoneal fluid from women with endometriosis. Obstet Gynecol 83:287–292 [PubMed]
- 53.Wu MY, Yang JH, Chao KH, Hwang JL, Yang YS, Ho HN (2000) Increase in the expression of killer cell inhibitory receptors on peritoneal natural killer cells in women with endometriosis. Fertil Steril 74:1187–1191 [DOI] [PubMed]
- 54.Arimoto T, Katagiri T, Oda K, Tsunoda T, Yasugi T, Osuga Y, Yoshikawa H, Nishii O, Yano T, Taketani Y, Nakamura Y (2003) Genome-wide cDNA microarray analysis of gene-expression profiles involved in ovarian endometriosis. Int J Oncol 22:551–560 [PubMed]
- 55.Witz CA, Montoya IA, Dey TD, Schenken RS (1994) Characterization of lymphocyte subpopulations and T cell activation in endometriosis. Am J Reprod Immunol 32:173–179 [DOI] [PubMed]
- 56.Jones RK, Bulmer JN, Searle RF (1996) Immunohistochemical characterization of stromal leukocytes in ovarian endometriosis: comparison of eutopic and ectopic endometrium with normal endometrium. Fertil Steril 66:81–89 [DOI] [PubMed]
- 57.Akoum A, Lemay A, Maheux R (2002) Estradiol and interleukin-1beta exert a synergistic stimulatory effect on the expression of the chemokine regulated upon activation, normal T cell expressed, and secreted in endometriotic cells. J Clin Endocrinol Metab 87:5785–5792 [DOI] [PubMed]
- 58.Khorram O, Taylor RN, Ryan IP, Schall TJ, Landers DV (1993) Peritoneal fluid concentrations of the cytokine RANTES correlate with the severity of endometriosis. Am J Obstet Gynecol 169:1545–1549 [DOI] [PubMed]
- 59.Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M (1995) Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol 155:1151–1164 [PubMed]
- 60.Szyllo K, Tchorzewski H, Banasik M, Glowacka E, Lewkowicz P, Kamer-Bartosinska A (2003) The involvement of T lymphocytes in the pathogenesis of endometriotic tissues overgrowth in women with endometriosis. Mediators Inflamm 12:131–138 [DOI] [PMC free article] [PubMed]
- 61.Podgaec S, Abrao MS, Dias JA Jr, Rizzo LV, de Oliveira RM, Baracat EC (2007) Endometriosis: an inflammatory disease with a Th2 immune response component. Hum Reprod 22(5):1373–1379 [DOI] [PubMed]
- 62.Zenclussen AC, Joachim R, Hagen E, Peiser C, Klapp BF, Arck PC (2002) Heme oxygenase is downregulated in stress-triggered and interleukin-12-mediated murine abortion. Scand J Immunol 55:560–569 [DOI] [PubMed]
- 63.Van Langendonckt A, Casanas-Roux F, Dolmans MM, Donnez J (2002) Potential involvement of hemoglobin and heme in the pathogenesis of peritoneal endometriosis. Fertil Steril 77:561–570 [DOI] [PubMed]
- 64.Szczepanska M, Kozlik J, Skrzypczak J, Mikolajczyk M (2003) Oxidative stress may be a piece in the endometriosis puzzle. Fertil Steril 79:1288–1293 [DOI] [PubMed]
- 65.D’Antonio M, Martelli F, Peano S, Papoian R, Borrelli F (2000) Ability of recombinant human TNF binding protein-1 (r-hTBP-1) to inhibit the development of experimentally-induced endometriosis in rats. J Reprod Immunol 48:81–98 [DOI] [PubMed]
- 66.Somigliana E, Vigano P, Rossi G, Carinelli S, Vignali M, Panina-Bordignon P (1999) Endometrial ability to implant in ectopic sites can be prevented by interleukin-12 in a murine model of endometriosis. Hum Reprod 14:2944–2950 [DOI] [PubMed]
- 67.Antsiferova YS, Sotnikova NY, Posiseeva LV, Shor AL (2005) Changes in the T-helper cytokine profile and in lymphocyte activation at the systemic and local levels in women with endometriosis. Fertil Steril 84:1705–1711 [DOI] [PubMed]
- 68.Odukoya OA, Bansal A, Wilson P, Lim K, Weetman AP, Cooke ID (1996) Soluble CD23 protein in the peritoneal fluid of patients with endometriosis. Hum Reprod 11:2018–2021 [DOI] [PubMed]
- 69.Confino E, Harlow L, Gleicher N (1990) Peritoneal fluid and serum autoantibody levels in patients with endometriosis. Fertil Steril 53:242–245 [DOI] [PubMed]
- 70.Mathur S, Peress MR, Williamson HO, Youmans CD, Maney SA, Garvin AJ, Rust PF, Fudenberg HH (1982) Autoimmunity to endometrium and ovary in endometriosis. Clin Exp Immunol 50:259–266 [PMC free article] [PubMed]
- 71.Moncayo H, Moncayo R, Benz R, Wolf A, Lauritzen C (1989) Ovarian failure and autoimmunity. Detection of autoantibodies directed against both the unoccupied luteinizing hormone/human chorionic gonadotropin receptor and the hormone–receptor complex of bovine corpus luteum. J Clin Invest 84:1857–1865 [DOI] [PMC free article] [PubMed]
- 72.D’Cruz OJ, Wild RA, Haas GG Jr, Reichlin M (1996) Antibodies to carbonic anhydrase in endometriosis: prevalence, specificity, and relationship to clinical and laboratory parameters. Fertil Steril 66:547–556 [DOI] [PubMed]
- 73.Lang GA, Yeaman GR (2001) Autoantibodies in endometriosis sera recognize a Thomsen–Friedenreich-like carbohydrate antigen. J Autoimmun 16:151–161 [DOI] [PubMed]
- 74.Mathur SP, Holt VL, Lee JH, Jiang H, Rust PF (1998) Levels of antibodies to transferrin and alpha 2-HS glycoprotein in women with and without endometriosis. Am J Reprod Immunol 40:69–73 [DOI] [PubMed]
- 75.Liu FT, Rabinovich GA (2005) Galectins as modulators of tumour progression. Nat Rev Cancer 5:29–41 [DOI] [PubMed]
- 76.Rubinstein N, Alvarez M, Zwirner NW, Toscano MA, Ilarregui JM, Bravo A, Mordoh J, Fainboim L, Podhajcer OL, Rabinovich GA (2004) Targeted inhibition of galectin-1 gene expression in tumor cells results in heightened T cell-mediated rejection; a potential mechanism of tumor-immune privilege. Cancer Cell 5:241–251 [DOI] [PubMed]
- 77.Vigano P, Gaffuri B, Somigliana E, Busacca M, Di Blasio AM, Vignali M (1998) Expression of intercellular adhesion molecule (ICAM)-1 mRNA and protein is enhanced in endometriosis versus endometrial stromal cells in culture. Mol Hum Reprod 4:1150–1156 [DOI] [PubMed]
- 78.Fukaya T, Sugawara J, Yoshida H, Murakami T, Yajima A (1999) Intercellular adhesion molecule-1 and hepatocyte growth factor in human endometriosis: original investigation and a review of literature. Gynecol Obstet Invest 47(Suppl 1):11–16 (discussion 16–17) [DOI] [PubMed]
- 79.Piva M, Sharpe-Timms KL (1999) Peritoneal endometriotic lesions differentially express a haptoglobin-like gene. Mol Hum Reprod 5:71–78 [DOI] [PubMed]
- 80.Piva M, Moreno JI, Sharpe-Timms KL (2002) Glycosylation and over-expression of endometriosis-associated peritoneal haptoglobin. Glycoconj J 19:33–41 [DOI] [PubMed]
- 81.Daniel Y, Geva E, Amit A, Eshed-Englender T, Baram A, Fait G, Lessing JB (2000) Do soluble cell adhesion molecules play a role in endometriosis? Am J Reprod Immunol 43:160–166 [DOI] [PubMed]
- 82.Schmidt M, Regidor PA, Engel K, Regidor M, Winterhager E, Scotti S, Schindler AE (2000) E- and P-selectin expression in endometriotic tissues and the corresponding endometria. Gynecol Endocrinol 14:111–117 [DOI] [PubMed]
- 83.Sharpe-Timms KL, Zimmer RL, Ricke EA, Piva M, Horowitz GM (2002) Endometriotic haptoglobin binds to peritoneal macrophages and alters their function in women with endometriosis. Fertil Steril 78:810–819 [DOI] [PubMed]
- 84.Stahn R, Goletz S, Stahn R, Wilmanowski R, Wang X, Briese V, Friese K, Jeschke U (2005) Human chorionic gonadotropin (hCG) as inhibitior of E-selectin-mediated cell adhesion. Anticancer Res 25:1811–1816 [PubMed]
- 85.Jeschke U, Wang X, Briese V, Friese K, Stahn R (2003) Glycodelin and amniotic fluid transferrin as inhibitors of E-selectin-mediated cell adhesion. Histochem Cell Biol 119:345–354 [DOI] [PubMed]
- 86.Jones CJ, Denton J, Fazleabas AT (2006) Morphological and glycosylation changes associated with the endometrium and ectopic lesions in a baboon model of endometriosis. Hum Reprod 21:3068–3080 [DOI] [PubMed]
- 87.Aplin JD, Meseguer M, Simon C, Ortiz ME, Croxatto H, Jones CJ (2001) MUC1, glycans and the cell-surface barrier to embryo implantation. Biochem Soc Trans 29:153–156 [DOI] [PubMed]
- 88.Aplin JD (1997) Adhesion molecules in implantation. Rev Reprod 2:84–93 [DOI] [PubMed]
- 89.Bourlev V, Volkov N, Pavlovitch S, Lets N, Larsson A, Olovsson M (2006) The relationship between microvessel density, proliferative activity and expression of vascular endothelial growth factor-A and its receptors in eutopic endometrium and endometriotic lesions. Reproduction 132:501–509 [DOI] [PubMed]
- 90.Laschke MW, Elitzsch A, Scheuer C, Holstein JH, Vollmar B, Menger MD (2006) Rapamycin induces regression of endometriotic lesions by inhibiting neovascularization and cell proliferation. Br J Pharmacol 149:137–144 [DOI] [PMC free article] [PubMed]
- 91.Lin YJ, Lai MD, Lei HY, Wing LY (2006) Neutrophils and macrophages promote angiogenesis in the early stage of endometriosis in a mouse model. Endocrinology 147:1278–1286 [DOI] [PubMed]
- 92.Jeon SH, Chae BC, Kim HA, Seo GY, Seo DW, Chun GT, Kim NS, Yie SW, Byeon WH, Eom SH, Ha KS, Kim YM, Kim PH (2007) Mechanisms underlying TGF-{beta}1-induced expression of VEGF and Flk-1 in mouse macrophages and their implications for angiogenesis. J Leukoc Biol 81:557–566 [DOI] [PubMed]
- 93.Sokolov DI, Solodovnikova NG, Pavlov OV, Niauri DA, Volkov NN, Sel’kov SA (2005) Study of cytokine profile and angiogenic potential of peritoneal fluid in patients with external genital endometriosis. Bull Exp Biol Med 140:541–544 [DOI] [PubMed]
- 94.Ryan IP, Tseng JF, Schriock ED, Khorram O, Landers DV, Taylor RN (1995) Interleukin-8 concentrations are elevated in peritoneal fluid of women with endometriosis. Fertil Steril 63:929–932 [PubMed]
- 95.Li A, Dubey S, Varney ML, Dave BJ, Singh RK (2003) IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J Immunol 170:3369–3376 [DOI] [PubMed]
- 96.Hudelist G, Lass H, Keckstein J, Walter I, Wieser F, Wenzl R, Mueller R, Czerwenka K, Kubista E, Singer CF (2005) Interleukin 1alpha and tissue-lytic matrix metalloproteinase-1 are elevated in ectopic endometrium of patients with endometriosis. Hum Reprod 20:1695–1701 [DOI] [PubMed]
- 97.Huang HF, Hong LH, Tan Y, Sheng JZ (2004) Matrix metalloproteinase 2 is associated with changes in steroid hormones in the sera and peritoneal fluid of patients with endometriosis. Fertil Steril 81:1235–1239 [DOI] [PubMed]
- 98.Chung HW, Lee JY, Moon HS, Hur SE, Park MH, Wen Y, Polan ML (2002) Matrix metalloproteinase-2, membranous type 1 matrix metalloproteinase, and tissue inhibitor of metalloproteinase-2 expression in ectopic and eutopic endometrium. Fertil Steril 78:787–795 [DOI] [PubMed]
- 99.Cramer DW, Wilson E, Stillman RJ, Berger MJ, Belisle S, Schiff I, Albrecht B, Gibson M, Stadel BV, Schoenbaum SC (1986) The relation of endometriosis to menstrual characteristics, smoking, and exercise. JAMA 255:1904–1908 [PubMed]
- 100.Fujishita A, Nakane PK, Koji T, Masuzaki H, Chavez RO, Yamabe T, Ishimaru T (1997) Expression of estrogen and progesterone receptors in endometrium and peritoneal endometriosis: an immunohistochemical and in situ hybridization study. Fertil Steril 67:856–864 [DOI] [PubMed]
- 101.Wieser F, Schneeberger C, Tong D, Tempfer C, Huber JC, Wenzl R (2002) PROGINS receptor gene polymorphism is associated with endometriosis. Fertil Steril 77:309–312 [DOI] [PubMed]
- 102.Attia GR, Zeitoun K, Edwards D, Johns A, Carr BR, Bulun SE (2000) Progesterone receptor isoform A but not B is expressed in endometriosis. J Clin Endocrinol Metab 85:2897–2902 [DOI] [PubMed]
- 103.Casey ML, MacDonald PC, Andersson S (1994) 17 beta-hydroxysteroid dehydrogenase type 2: chromosomal assignment and progestin regulation of gene expression in human endometrium. J Clin Invest 94:2135–2141 [DOI] [PMC free article] [PubMed]
- 104.Zeitoun K, Takayama K, Sasano H, Suzuki T, Moghrabi N, Andersson S, Johns A, Meng L, Putman M, Carr B, Bulun SE (1998) Deficient 17beta-hydroxysteroid dehydrogenase type 2 expression in endometriosis: failure to metabolize 17beta-estradiol. J Clin Endocrinol Metab 83:4474–4480 [DOI] [PubMed]
- 105.Fazleabas AT, Brudney A, Chai D, Langoi D, Bulun SE (2003) Steroid receptor and aromatase expression in baboon endometriotic lesions. Fertil Steril 80(Suppl 2):820–827 [DOI] [PubMed]
- 106.Fang Z, Yang S, Gurates B, Tamura M, Simpson E, Evans D, Bulun SE (2002) Genetic or enzymatic disruption of aromatase inhibits the growth of ectopic uterine tissue. J Clin Endocrinol Metab 87:3460–3466 [DOI] [PubMed]
- 107.Sakamoto Y, Harada T, Horie S, Iba Y, Taniguchi F, Yoshida S, Iwabe T, Terakawa N (2003) Tumor necrosis factor-alpha-induced interleukin-8 (IL-8) expression in endometriotic stromal cells, probably through nuclear factor-kappa B activation: gonadotropin-releasing hormone agonist treatment reduced IL-8 expression. J Clin Endocrinol Metab 88:730–735 [DOI] [PubMed]
- 108.Vliagoftis H, Dimitriadou V, Boucher W, Rozniecki JJ, Correia I, Raam S, Theoharides TC (1992) Estradiol augments while tamoxifen inhibits rat mast cell secretion. Int Arch Allergy Immunol 98:398–409 [DOI] [PubMed]
- 109.McLaren J, Prentice A, Charnock-Jones DS, Millican SA, Muller KH, Sharkey AM, Smith SK (1996) Vascular endothelial growth factor is produced by peritoneal fluid macrophages in endometriosis and is regulated by ovarian steroids. J Clin Invest 98:482–489 [DOI] [PMC free article] [PubMed]
- 110.Joachim R, Zenclussen AC, Polgar B, Douglas AJ, Fest S, Knackstedt M, Klapp BF, Arck PC (2003) The progesterone derivative dydrogesterone abrogates murine stress-triggered abortion by inducing a Th2 biased local immune response. Steroids 68:931–940 [DOI] [PubMed]
- 111.Blois SM, Joachim R, Kandil J, Margni R, Tometten M, Klapp BF, Arck PC (2004) Depletion of CD8+ cells abolishes the pregnancy protective effect of progesterone substitution with dydrogesterone in mice by altering the Th1/Th2 cytokine profile. J Immunol 172:5893–5899 [DOI] [PubMed]
- 112.Orsal AS, Blois S, Labuz D, Peters EM, Schaefer M, Arck PC (2006) The progesterone derivative dydrogesterone down-regulates neurokinin 1 receptor expression on lymphocytes, induces a Th2 skew and exerts hypoalgesic effects in mice. J Mol Med 84:159–167 [DOI] [PubMed]
- 113.Schor E, Baracat EC, Simoes MJ, de Freitas V, Giannotti Filho O, de Lima GR (1999) Effects of conjugated estrogens and progestogen in surgically induced endometriosis in oophorectomized rats. Clin Exp Obstet Gynecol 26:158–161 [PubMed]
- 114.Quereda F, Barroso J, Acien P (1996) Individual and combined effects of triptoreline and gestrinone on experimental endometriosis in rats. Eur J Obstet Gynecol Reprod Biol 67:35–40 [DOI] [PubMed]
- 115.Katsuki Y, Takano Y, Futamura Y, Shibutani Y, Aoki D, Udagawa Y, Nozawa S (1998) Effects of dienogest, a synthetic steroid, on experimental endometriosis in rats. Eur J Endocrinol 138:216–226 [DOI] [PubMed]
- 116.Vercellini P, Fedele L, Pietropaolo G, Frontino G, Somigliana E, Crosignani PG (2003) Progestogens for endometriosis: forward to the past. Hum Reprod Update 9:387–396 [DOI] [PubMed]
- 117.Crosignani P, Olive D, Bergqvist A, Luciano A (2006) Advances in the management of endometriosis: an update for clinicians. Hum Reprod Update 12:179–189 [DOI] [PubMed]
- 118.Winkel CA, Scialli AR (2001) Medical and surgical therapies for pain associated with endometriosis. J Womens Health Gend Based Med 10:137–162 [DOI] [PubMed]
- 119.Atwal G, du Plessis D, Armstrong G, Slade R, Quinn M (2005) Uterine innervation after hysterectomy for chronic pelvic pain with, and without, endometriosis. Am J Obstet Gynecol 193:1650–1655 [DOI] [PubMed]
- 120.Tokushige N, Markham R, Russell P, Fraser IS (2006) High density of small nerve fibres in the functional layer of the endometrium in women with endometriosis. Hum Reprod 21:782–787 [DOI] [PubMed]
- 121.Tokushige N, Markham R, Russell P, Fraser IS (2006) Nerve fibres in peritoneal endometriosis. Hum Reprod 21:3001–3007 [DOI] [PubMed]
- 122.Berkley KJ, Dmitrieva N, Curtis KS, Papka RE (2004) Innervation of ectopic endometrium in a rat model of endometriosis. Proc Natl Acad Sci USA 101:11094–11098 [DOI] [PMC free article] [PubMed]
- 123.Luts L, Sundler F (1998) Autotransplantation of rat parathyroid glands: a study on morphological changes. Transplantation 66:446–453 [DOI] [PubMed]
- 124.Ferretti A, Boschi E, Stefani A, Spiga S, Romanelli M, Lemmi M, Giovannetti A, Longoni B, Mosca F (2003) Angiogenesis and nerve regeneration in a model of human skin equivalent transplant. Life Sci 73:1985–1994 [DOI] [PubMed]
- 125.Skoff AM, Adler JE (2006) Nerve growth factor regulates substance P in adult sensory neurons through both TrkA and p75 receptors. Exp Neurol 197:430–436 [DOI] [PubMed]
- 126.Bracci-Laudiero L, Aloe L, Caroleo MC, Buanne P, Costa N, Starace G, Lundeberg T (2005) Endogenous NGF regulates CGRP expression in human monocytes, and affects HLA-DR and CD86 expression and IL-10 production. Blood 106:3507–3514 [DOI] [PubMed]
- 127.Dmitrieva N, McMahon SB (1996) Sensitisation of visceral afferents by nerve growth factor in the adult rat. Pain 66:87–97 [DOI] [PubMed]
- 128.Qiu BS, Vallance BA, Blennerhassett PA, Collins SM (1999) The role of CD4+ lymphocytes in the susceptibility of mice to stress-induced reactivation of experimental colitis. Nat Med 5:1178–1182 [DOI] [PubMed]
- 129.Levenstein S (2003) Stress and ulcerative colitis: convincing the doubting Thomases. Am J Gastroenterol 98:2112–2115 [DOI] [PubMed]
- 130.Gil KM, Keefe FJ, Sampson HA, McCaskill CC, Rodin J, Crisson JE (1987) The relation of stress and family environment to atopic dermatitis symptoms in children. J Psychosom Res 31:673–684 [DOI] [PubMed]
- 131.Paus R, Theoharides TC, Arck PC (2006) Neuroimmunoendocrine circuitry of the ‘brain–skin connection’. Trends Immunol 27:32–39 [DOI] [PubMed]
- 132.Arck P, Knackstedt MK, Blois SM (2006) Current insights and future perspectives on neuro-endocrine–immune circuitry challenging pregnancy maintenance and fetal health. J Reprod Med Endo 3:98–102
- 133.Lima AP, Moura MD, Rosa e Silva AA (2006) Prolactin and cortisol levels in women with endometriosis. Braz J Med Biol Res 39:1121–1127 [DOI] [PubMed]
- 134.Cacioppo JT, Berntson GG, Malarkey WB, Kiecolt-Glaser JK, Sheridan JF, Poehlmann KM, Burleson MH, Ernst JM, Hawkley LC, Glaser R (1998) Autonomic, neuroendocrine, and immune responses to psychological stress: the reactivity hypothesis. Ann N Y Acad Sci 840:664–673 [DOI] [PubMed]
- 135.Blalock JE (1994) The syntax of immune–neuroendocrine communication. Immunol Today 15:504–511 [DOI] [PubMed]
- 136.Glaser R, Kiecolt-Glaser JK (2005) Stress-induced immune dysfunction: implications for health. Nat Rev Immunol 5:243–251 [DOI] [PubMed]
- 137.Benschop RJ, Rodriguez-Feuerhahn M, Schedlowski M (1996) Catecholamine-induced leukocytosis: early observations, current research, and future directions. Brain Behav Immun 10:77–91 [DOI] [PubMed]
- 138.Webster JI, Tonelli L, Sternberg EM (2002) Neuroendocrine regulation of immunity. Annu Rev Immunol 20:125–163 [DOI] [PubMed]
- 139.Dhabhar FS (2000) Acute stress enhances while chronic stress suppresses skin immunity. The role of stress hormones and leukocyte trafficking. Ann N Y Acad Sci 917:876–893 [DOI] [PubMed]
- 140.Wonnacott KM, Bonneau RH (2002) The effects of stress on memory cytotoxic T lymphocyte-mediated protection against herpes simplex virus infection at mucosal sites. Brain Behav Immun 16:104–117 [DOI] [PubMed]
- 141.Elenkov IJ, Webster EL, Torpy DJ, Chrousos GP (1999) Stress, corticotropin-releasing hormone, glucocorticoids, and the immune/inflammatory response: acute and chronic effects. Ann N Y Acad Sci 876:1–11 (discussion 11–13) [DOI] [PubMed]
- 142.Crofford LJ, Sano H, Karalis K, Friedman TC, Epps HR, Remmers EF, Mathern P, Chrousos GP, Wilder RL (1993) Corticotropin-releasing hormone in synovial fluids and tissues of patients with rheumatoid arthritis and osteoarthritis. J Immunol 151:1587–1596 [PubMed]
- 143.van Tol EA, Petrusz P, Lund PK, Yamauchi M, Sartor RB (1996) Local production of corticotropin releasing hormone is increased in experimental intestinal inflammation in rats. Gut 39:385–392 [DOI] [PMC free article] [PubMed]
- 144.Singh VK (1989) Stimulatory effect of corticotropin-releasing neurohormone on human lymphocyte proliferation and interleukin-2 receptor expression. J Neuroimmunol 23:257–262 [DOI] [PubMed]
- 145.Singh VK, Leu SJ (1990) Enhancing effect of corticotropin-releasing neurohormone on the production of interleukin-1 and interleukin-2. Neurosci Lett 120:151–154 [DOI] [PubMed]
- 146.Angioni S, Petraglia F, Gallinelli A, Cossarizza A, Franceschi C, Muscettola M, Genazzani AD, Surico N, Genazzani AR (1993) Corticotropin-releasing hormone modulates cytokines release in cultured human peripheral blood mononuclear cells. Life Sci 53:1735–1742 [DOI] [PubMed]
- 147.Maes M, Kenis G, Kubera M (2003) In humans, corticotropin releasing hormone antagonizes some of the negative immunoregulatory effects of serotonin. Neuro Endocrinol Lett 24:420–424 [PubMed]
- 148.Karalis K, Sano H, Redwine J, Listwak S, Wilder RL, Chrousos GP (1991) Autocrine or paracrine inflammatory actions of corticotropin-releasing hormone in vivo. Science 254:421–423 [DOI] [PubMed]
- 149.Singh VK, Fudenberg HH (1988) Binding of [125I]corticotropin releasing factor to blood immunocytes and its reduction in Alzheimer’s disease. Immunol Lett 18:5–8 [DOI] [PubMed]
- 150.Cao J, Papadopoulou N, Kempuraj D, Boucher WS, Sugimoto K, Cetrulo CL, Theoharides TC (2005) Human mast cells express corticotropin-releasing hormone (CRH) receptors and CRH leads to selective secretion of vascular endothelial growth factor. J Immunol 174:7665–7675 [DOI] [PubMed]
- 151.Stephanou A, Jessop DS, Knight RA, Lightman SL (1990) Corticotrophin-releasing factor-like immunoreactivity and mRNA in human leukocytes. Brain Behav Immun 4:67–73 [DOI] [PubMed]
- 152.Kempuraj D, Papadopoulou NG, Lytinas M, Huang M, Kandere-Grzybowska K, Madhappan B, Boucher W, Christodoulou S, Athanassiou A, Theoharides TC (2004) Corticotropin-releasing hormone and its structurally related urocortin are synthesized and secreted by human mast cells. Endocrinology 145:43–48 [DOI] [PubMed]
- 153.Bileviciute I, Ahmed M, Bergstrom J, Ericsson-Dahlstrand A, Kreicbergs A, Lundeberg T (1997) Expression of corticotropin-releasing factor in the peripheral nervous system of the rat. Neuroreport 8:3127–3130 [DOI] [PubMed]
- 154.Merchenthaler I, Hynes MA, Vigh S, Shally AV, Petrusz P (1983) Immunocytochemical localization of corticotropin releasing factor (CRF) in the rat spinal cord. Brain Res 275:373–377 [DOI] [PubMed]
- 155.Saunders PR, Santos J, Hanssen NP, Yates D, Groot JA, Perdue MH (2002) Physical and psychological stress in rats enhances colonic epithelial permeability via peripheral CRH. Dig Dis Sci 47:208–215 [DOI] [PubMed]
- 156.Florio P, Busacca M, Vignali M, Vigano P, Woods RJ, Lowry PJ, Genazzani AR, Luisi S, Santuz M, Petraglia F (1998) Peritoneal fluid levels of immunoreactive corticotropin-releasing factor (CRF) and CRF-binding protein (CRF-BP) in healthy and endometriosic women. J Endocrinol Invest 21:37–42 [DOI] [PubMed]
- 157.Benschop RJ, Nieuwenhuis EE, Tromp EA, Godaert GL, Ballieux RE, van Doornen LJ (1994) Effects of beta-adrenergic blockade on immunologic and cardiovascular changes induced by mental stress. Circulation 89:762–769 [DOI] [PubMed]
- 158.Sanders VM, Baker RA, Ramer-Quinn DS, Kasprowicz DJ, Fuchs BA, Street NE (1997) Differential expression of the beta2-adrenergic receptor by Th1 and Th2 clones: implications for cytokine production and B cell help. J Immunol 158:4200–4210 [PubMed]
- 159.Arck PC, Merali FS, Stanisz AM, Stead RH, Chaouat G, Manuel J, Clark DA (1995) Stress-induced murine abortion associated with substance P-dependent alteration in cytokines in maternal uterine decidua. Biol Reprod 53:814–819 [DOI] [PubMed]
- 160.Aloe L, Alleva E, Fiore M (2002) Stress and nerve growth factor: findings in animal models and humans. Pharmacol Biochem Behav 73:159–166 [DOI] [PubMed]
- 161.Peters EM, Handjiski B, Kuhlmei A, Hagen E, Bielas H, Braun A, Klapp BF, Paus R, Arck PC (2004) Neurogenic inflammation in stress-induced termination of murine hair growth is promoted by nerve growth factor. Am J Pathol 165:259–271 [DOI] [PMC free article] [PubMed]
- 162.Massaad CA, Safieh-Garabedian B, Poole S, Atweh SF, Jabbur SJ, Saade NE (2004) Involvement of substance P, CGRP and histamine in the hyperalgesia and cytokine upregulation induced by intraplantar injection of capsaicin in rats. J Neuroimmunol 153:171–182 [DOI] [PubMed]
- 163.Dallos A, Kiss M, Polyanka H, Dobozy A, Kemeny L, Husz S (2006) Effects of the neuropeptides substance P, calcitonin gene-related peptide, vasoactive intestinal polypeptide and galanin on the production of nerve growth factor and inflammatory cytokines in cultured human keratinocytes. Neuropeptides 40:251–263 [DOI] [PubMed]
- 164.Seegers HC, Hood VC, Kidd BL, Cruwys SC, Walsh DA (2003) Enhancement of angiogenesis by endogenous substance P release and neurokinin-1 receptors during neurogenic inflammation. J Pharmacol Exp Ther 306:8–12 [DOI] [PubMed]
- 165.Barouch R, Kazimirsky G, Appel E, Brodie C (2001) Nerve growth factor regulates TNF-alpha production in mouse macrophages via MAP kinase activation. J Leukoc Biol 69:1019–1026 [PubMed]
- 166.Tometten M, Blois S, Kuhlmei A, Stretz A, Klapp BF, Arck PC (2006) Nerve growth factor translates stress response and subsequent murine abortion via adhesion molecule-dependent pathways. Biol Reprod 74:674–683 [DOI] [PubMed]
- 167.Blasing H, Hendrix S, Paus R (2005) Pro-inflammatory cytokines upregulate the skin immunoreactivity for NGF, NT-3, NT-4 and their receptor, p75NTR in vivo: a preliminary report. Arch Dermatol Res 296:580–584 [DOI] [PubMed]
- 168.Hattori A, Iwasaki S, Murase K, Tsujimoto M, Sato M, Hayashi K, Kohno M (1994) Tumor necrosis factor is markedly synergistic with interleukin 1 and interferon-gamma in stimulating the production of nerve growth factor in fibroblasts. FEBS Lett 340:177–180 [DOI] [PubMed]
- 169.Steinhoff M, Vergnolle N, Young SH, Tognetto M, Amadesi S, Ennes HS, Trevisani M, Hollenberg MD, Wallace JL, Caughey GH, Mitchell SE, Williams LM, Geppetti P, Mayer EA, Bunnett NW (2000) Agonists of proteinase-activated receptor 2 induce inflammation by a neurogenic mechanism. Nat Med 6:151–158 [DOI] [PubMed]
- 170.Sanfilippo JS, Williams RS, Yussman MA, Cook CL, Bissonnette F (1992) Substance P in peritoneal fluid. Am J Obstet Gynecol 166:155–159 [DOI] [PubMed]
- 171.Watkins LR, Maier SF (2000) The pain of being sick: implications of immune-to-brain communication for understanding pain. Annu Rev Psychol 51:29–57 [DOI] [PubMed]
- 172.Yirmiya R, Weidenfeld J, Barak O, Avitsur R, Pollak Y, Gallily R, Wohlman A, Ovadia H, Ben-Hur T (1999) The role of brain cytokines in mediating the behavioral and neuroendocrine effects of intracerebral mycoplasma fermentans. Brain Res 829:28–38 [DOI] [PubMed]
- 173.Avitsur R, Cohen E, Yirmiya R (1997) Effects of interleukin-1 on sexual attractivity in a model of sickness behavior. Physiol Behav 63:25–30 [DOI] [PubMed]
- 174.Reichenberg A, Yirmiya R, Schuld A, Kraus T, Haack M, Morag A, Pollmacher T (2001) Cytokine-associated emotional and cognitive disturbances in humans. Arch Gen Psychiatry 58:445–452 [DOI] [PubMed]
- 175.Charmandari E, Tsigos C, Chrousos G (2005) Endocrinology of the stress response. Annu Rev Physiol 67:259–284 [DOI] [PubMed]
- 176.Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W (1987) Interleukin-1 stimulates the secretion of hypothalamic corticotropin-releasing factor. Science 238:522–524 [DOI] [PubMed]
- 177.Harbuz MS, Stephanou A, Sarlis N, Lightman SL (1992) The effects of recombinant human interleukin (IL)-1 alpha, IL-1 beta or IL-6 on hypothalamo–pituitary–adrenal axis activation. J Endocrinol 133:349–355 [DOI] [PubMed]
- 178.Schobitz B, Reul JM, Holsboer F (1994) The role of the hypothalamic–pituitary–adrenocortical system during inflammatory conditions. Crit Rev Neurobiol 8:263–291 [PubMed]
- 179.Braun DP, Gebel H, House R, Rana N, Dmowski NP (1996) Spontaneous and induced synthesis of cytokines by peripheral blood monocytes in patients with endometriosis. Fertil Steril 65:1125–1129 [PubMed]
- 180.Banks WA (2005) Blood–brain barrier transport of cytokines: a mechanism for neuropathology. Curr Pharm Des 11:973–984 [DOI] [PubMed]
- 181.Esposito P, Gheorghe D, Kandere K, Pang X, Connolly R, Jacobson S, Theoharides TC (2001) Acute stress increases permeability of the blood–brain-barrier through activation of brain mast cells. Brain Res 888:117–127 [DOI] [PubMed]
- 182.Luheshi GN, Bluthe RM, Rushforth D, Mulcahy N, Konsman JP, Goldbach M, Dantzer R (2000) Vagotomy attenuates the behavioural but not the pyrogenic effects of interleukin-1 in rats. Auton Neurosci 85:127–132 [DOI] [PubMed]
- 183.Fazleabas AT (2006) A baboon model for inducing endometriosis. Methods Mol Med 121:95–99 [DOI] [PubMed]
- 184.Sharpe-Timms KL (2002) Using rats as a research model for the study of endometriosis. Ann N Y Acad Sci 955:318–327 (discussion 340–312, 396–406) [DOI] [PubMed]
- 185.Vernon MW, Wilson EA (1985) Studies on the surgical induction of endometriosis in the rat. Fertil Steril 44:684–694 [PubMed]
- 186.Harris HA, Bruner-Tran KL, Zhang X, Osteen KG, Lyttle CR (2005) A selective estrogen receptor-beta agonist causes lesion regression in an experimentally induced model of endometriosis. Hum Reprod 20:936–941 [DOI] [PubMed]
- 187.Hirata T, Osuga Y, Yoshino O, Hirota Y, Harada M, Takemura Y, Morimoto C, Koga K, Yano T, Tsutsumi O, Taketani Y (2005) Development of an experimental model of endometriosis using mice that ubiquitously express green fluorescent protein. Hum Reprod 20:2092–2096 [DOI] [PubMed]
- 188.Fang Z, Yang S, Lydon JP, DeMayo F, Tamura M, Gurates B, Bulun SE (2004) Intact progesterone receptors are essential to counteract the proliferative effect of estradiol in a genetically engineered mouse model of endometriosis. Fertil Steril 82:673–678 [DOI] [PubMed]
- 189.Efstathiou JA, Sampson DA, Levine Z, Rohan RM, Zurakowski D, Folkman J, D’Amato RJ, Rupnick MA (2005) Nonsteroidal antiinflammatory drugs differentially suppress endometriosis in a murine model. Fertil Steril 83:171–181 [DOI] [PubMed]
- 190.Bruner-Tran KL, Eisenberg E, Yeaman GR, Anderson TA, McBean J, Osteen KG (2002) Steroid and cytokine regulation of matrix metalloproteinase expression in endometriosis and the establishment of experimental endometriosis in nude mice. J Clin Endocrinol Metab 87:4782–4791 [DOI] [PubMed]
- 191.Fortin M, Lepine M, Page M, Osteen K, Massie B, Hugo P, Steff AM (2003) An improved mouse model for endometriosis allows noninvasive assessment of lesion implantation and development. Fertil Steril 80(Suppl 2):832–838 [DOI] [PubMed]
- 192.Waller KG, Shaw RW (1993) Gonadotropin-releasing hormone analogues for the treatment of endometriosis: long-term follow-up. Fertil Steril 59:511–515 [DOI] [PubMed]
- 193.Prentice A, Deary AJ, Bland E (2000) Progestagens and anti-progestagens for pain associated with endometriosis. Cochrane Database Syst Rev 2:CD002122 [DOI] [PubMed]
- 194.Overton CE, Lindsay PC, Johal B, Collins SA, Siddle NC, Shaw RW, Barlow DH (1994) A randomized, double-blind, placebo-controlled study of luteal phase dydrogesterone (Duphaston) in women with minimal to mild endometriosis. Fertil Steril 62:701–707 [DOI] [PubMed]
- 195.Kempuraj D, Madhappan B, Christodoulou S, Boucher W, Cao J, Papadopoulou N, Cetrulo CL, Theoharides TC (2005) Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. Br J Pharmacol 145:934–944 [DOI] [PMC free article] [PubMed]
- 196.Osteen KG, Bruner-Tran KL, Eisenberg E (2005) Reduced progesterone action during endometrial maturation: a potential risk factor for the development of endometriosis. Fertil Steril 83:529–537 [DOI] [PubMed]