

Abstract
Direct cultivation of the first filial generation of gametophyte clones from different Laminaria species is a highly effective way of utilizing kelp heterozygous vigor (heterosis). A male gametophyte clone of L. longissima Miyabe and a female one of L. japonica Areschoug were hybridized, generating Dongfang No. 2 hybrid kelp. This hybrid kelp was used directly in trial cultivation, and its agronomical traits were evaluated. L. longissima and L. japonica are obviously different and complement each other in their morphological characteristics and ecological performances. The hybrid of their gametophyte clones, Dongfang No. 2, showed 56.8% heterozygous vigor in yield. It also showed increased yields of 41.0 and 76.4% compared to the widely used commercial kelps Variety 1 and Variety 2, respectively. In large-scale cultivation trials at different locations and in different years, Dongfang No. 2 attained significantly higher yields than Varieties 1 and 2, increasing yield by 26.4% on average over Variety 1 and by 65.0% over the other. Dongfang No. 2 has a robust holdfast and a wide, long and deep-brown uniform blade, which shows a distinct middle groove. In addition to yield, Dongfang No. 2 also demonstrates obvious heterozygous vigor in other agronomic traits. It is resistant to strong irradiance, as the two commercial varieties are, has an appropriate vegetative maturation time, and adapts well to a range of different culture conditions. The parentage analysis using AFLP of total DNA and SNP of the ITS region of ribosomal RNA transcription unit showed that Dongfang No. 2 is the real hybrid of L. japonica and L. longissima.
Key words: Dongfang No. 2, hybrid kelp, gametophyte clone, heterosis, hybridization
Introduction
Kelp (Laminaria japonica) was introduced into China in 1927 (Tseng 2001). Its cultivation, breeding and biological research were initiated in China at the end of 1950s. With the development of a summer sporeling-raising method (Tseng et al. 1955) and the implementation of a raft cultivation method (Li 1990), kelp cultivation has evolved rapidly into the largest maricultural industry in China with China’s total yield ranked first of the world (Tseng 2001; Zemke-White and Ohno 1999). The breeding through selection of a set of elite kelp varieties including Haiqing No. 1 (Fang et al. 1962) and Zhaohoucheng No. 1 (Tian and Yuan 1989) has promoted the development of Chinese kelp cultivation.
In order to breed varieties with high yield potential and desirable qualities, physical mutation methods have been used by Chinese breeders, and varieties such as 1170 were developed in the 1970s in this way (Algal Breeding Team of Qingdao Maricultural Institute 1976).
At the end of the 1970s, gametophyte cloning and gametophyte clone hybridizing methods were developed, bringing breeders the chances of developing hybrid kelps (Fang et al. 1978, 1983; Li et al. 2002, 1999; Zhou and Wu 1998). Diverse varieties were developed through hybridization of L. longissima and L. japonica gametophyte clones, giving breeders the opportunity to develop hybrid kelps with the introduction of alien Laminaria species like L. longissima (Zhang et al. 1998, 2000). These included Danhai No. 1 (Ou et al. 1983) and 901 (Zhang et al. 2001). Each was developed through a time-consuming and labor-intensive process including gametophyte clone hybridization, self-crossing, and desirable trait-targeted selection (Zhang et al. 1999). Apart from being used as starting materials for cross-breeding gametophyte clones, hybrids can also be used directly for cultivation. In 1985, Danza No. 10, the hybrid of gametophyte clones of L. longissima and L. japonica, was developed and used in trial cultivation, demonstrating the direct use of kelp heterosis in production (Fang et al. 1985). Because a large-scale hybrid sporeling-raising technique was not available at that time, this hybrid kelp was not commonly used.
Laminaria longissima and L. japonica show distinct differences in their morphological characteristics and eco-physiological requirements but complement each other in their agronomic traits. It was expected that their hybrids would hold significant heterozygous vigor (heterosis). To our knowledge, L. japonica, a Laminaria species well adapted to Chinese coastal waters, has always been used as the maternal parent in previous hybridization breedings of elite kelp varieties, and L. longissima, recently imported from Japan, as the paternal one. The objective of this study was to develop a method for exploiting the kelp heterosis effect directly in cultivation. The feasibility of such a strategy was tested by hybridizing a male gametophyte clone of L. longissima with a female one of L. japonica in 2001. The resulting hybrid, Dongfang No. 2, clearly showed heterosis. By combining the gametophyte cloning, hybridizing and hybrid sporeling-raising methods available currently, Dongfang No. 2 was cultured from 2002–2004, reaching a total trial cultivation area of ca. 333 hectares.
High biomass yield is always the target of genetic improvement of kelp. Important factors for improving yield are fast growth, reduced tissue abrasion and tolerance to high irradiance and water temperature. Due to its high yield potential and excellent economic characteristics, Dongfang No. 2 has been approved by the Chinese Approving Committee of Aquacultural Elite Varieties and Stock Seeds in 2005 with approval number GS02-001–2004, with its traits evaluated and its performance in trial cultivation reported in this paper.
Materials and methods
Gametophytes and their cultivation
The male gametophyte clone of Laminaria longissima and the female gametophyte clone of L. japonica used in this study were maintained at the Aquacultural Elite Variety Factory, Shandong Oriental Ocean Sci-tec, Yantai, Shandong Province, China. The gametophyte clones were cultivated in seawater supplemented with 11.76 μmol L−1 NaNO3 and 7.35 μmol L−1 KH2PO4 at 10–15°C and under 40–60 μmol photons m−2 s−1 irradiance. The culture container was aerated (Wang 1994; Li et al. 2003a, b, c ). The filamentous gametophytes grew gradually into ball-like shapes. After reaching a diameter of 2 mm, the balls were fragmented into pieces of about 200 μm in length using a tissue homogenizer set at 200 W and 10,000–12,000 rpm for 10 s. These filamentous gametophyte fragments were further cultivated until a sufficient amount of biomass was obtained. The enriched seawater was changed every week. Biomass was reduced when it reached 20 g L−1 to maintain high growth rates of gametophytes (Li et al. 2003a, b, c).
Gametophyte clone hybridization
Gametophyte clones were collected by filtering through a sieve cloth and weighing. Male and female gametophyte clones were mixed at a ratio of 2:1, homogenized (see above) and cultured under short-day regime (10 h light/14 h dark) for about 10 days. Before the oocytes were released, the mixed gametophyte clones were homogenized again, filtered using sieve cloth with the pore size of ca. 50 μm, and sprayed onto curtains kept in cooled seawater, with 4.5 g of the gametophyte clone mix to each curtain.
Sporelings used for trait evaluation were raised indoors at a constant temperature of 8–10°C. The seawater used was sterilized using double sand filtering and an ultraviolet sterilizer set at 40 W and 15 cm3 s−1 and cooled to 8–10°C before use. Fluorescent lamps were used as the light source. Irradiance and nutrient supplements in seawater used at different developmental stages are listed in Table 1. Until eight rows of cells developed, the sporelings were kept under 10 h of irradiance; after that stage, sporelings were cultivated under 12 h irradiance per day. The water was replaced completely every 3 days until eight rows of cells had developed, and every day after this stage. After 50–60 days, when sporelings reached a length of 1.5–2.0 cm, they were transferred from the hatchery to the open sea usually in the first or second 10 days of October. The palm curtains with sporelings were placed horizontally in the top 100 cm layer of the water in the direction of seawater current. The young sporelings were fertilized using fertilizer bags containing 150–250 g NH4NO3, with 10 of these fixed to each floating rope. The bags were replaced every second day.
Table 1.
Irradiance and nutrition supplement for cultivation of hybrid sporelings
| Growth phase | Irradiance (μmole photons m−2 s−1) | Nutrient supplementa |
|---|---|---|
| Sporophyte formation to 4 cell rows | 50 | A |
| 4 cell rows to 8 cell rows | 60 | A |
| 8 cell rows to 0.5 mm sporophyte | 70 | B |
| 0.5–2 mm sporophyte | 80 | B |
| 2–3 mm sporophyte | 90 | B |
| 3–4 mm sporophyte | 90 | C |
| 4–8 mm sporophyte | 90 | C |
| 8–20 mm sporophyte | 100 | C |
aA 23.53 μmol L−1 NaNO3, 1.47 μmol L−1 KH2PO4 and 2.99 μmol L−1 FeC6H5O7·5H2O; B 35.29 μmol L−1 NaNO3, 2.21 μmol L−1 KH2PO4 and 2.99 μmol L−1 FeC6H5O7·5H2O; C 47.06 μmol L−1 NaNO3, 2.94 μmol L−1 KH2PO4 and 2.99 μmol L−1 FeC6H5O7·5H2O
In about 50 days, the sporelings increased in length to approximately 40 cm. Sporelings of such size were used for transplanting. Sporelings used for large-scale trial cultivation were raised in a commercial sporeling-raising workshop. Instead of changing water by hand and illuminating using fluorescent lamps, these sporelings were cultured under a plastic membrane and/or net-roofed natural light and in seawater current (current width 2.2 m, ca. 3 L.s-1). All other conditions were identical to the sporeling raising carried out for trait evaluation. The times of transferring young sporelings to the open sea and transplanting of sporelings were the same as commercial production (Li et al. 2003a, b, c; Scoggan et al. 1989; Tseng et al. 1955). The sporelings of L. longissima and L. japonica were raised by hybridizing about 15 randomly selected male gametophyte clones with 15 female ones cultured separately but mixed before hybridization. The sporelings of Varieties 1 and 2 were raised as is done for commercial purposes.
Parentage analysis of Dongfang No. 2
DNA was isolated from about 100 mg young sporophytes following the protocol described by Patwary and Ron (1993). The parentage of Dongfang No. 2 was determined with amplified fragment length polymorphism (AFLP) analysis of total DNA and single nucleotide polymorphism (SNP) of the internal transcribed spacer region of ribosomal RNA genes (ITS region). AFLP analysis was done following the protocol described by Vos et al. (1995). Instead of isotope labeling and autoradiography, PCR products were separated using PAGE and envisaged by silver staining. The adaptors used included the Eco RI regenerating 5′-CTC GTA GAC TGC GTA CCA-3′, and the Mse I regenerating 5′-GAC GAT GAG TCC TGA G-3′. The pre-amplification primers were EP 5′-GAC TGC GTA CCA ATT C-3′ at the Eco RI side and MP 5′-GAT GAG TCC TGA GTA A-3′ at the Mse I side. Selective amplification was achieved using a set of primers derived from EP and MP by adding 2- or 3-nucleotide overhangs at their 3′ ends. These primers are designed as F1 (EP + CA), F2 (EP + ACC), F3 (EP + CAC), F4 (EP + AGG), R1 (MP + CTA), R2 (MP + CAA), R3 (MP + CGA) and R4 (MP + CTT). Sixteen primer combinations were generated by pairing forward and reverse primers; they were used for selective amplification, and 10 yielded clear and even band patterns.
The ITS region of ribosomal RNA transcribed unit was amplified using primers LB1 (5′-CGC GAG TCA TCA GCT CGC ATT-3′) and LB2 (5′-AGC TTC ACT CGC CCG TAC TGG-3′) previously used by Yoon et al. (2001). PCR was carried out by pre-denaturing at 94°C for 5 min followed by 30 cycles of denaturing at 94°C for 1 min, annealing at 57°C for 1 min and extending at 72°C for 1 min, and an extra extension at 72°C for 10 min. In the 25 μL reaction mixture, there was 50 ng template DNA, 2.0 mmol L-1 MgCl2, 0.2 mmol L-1 dNTP (each kind), 0.2 μmol L−1 primers (each direction), 1 U Taq DNA polymerase and 1× buffer. The PCR product was checked with 1.0% agarose gel electrophoresis and EtBr staining, ligated into pMD-T vector (TaKaRa Biotechnology, Dalian, China) and transferred into competent E. coli JM109. Recombinants with appropriate inserts were selected with their inserts sequenced using ABI 3730 sequencer.
It was assumed that DNA fragments of a specific length found in the different materials used (L. japonica, L. longissima, Dongfang No. 2 and Varieties 1 and 2) would present one specific DNA locus. Number “1” was assigned to a band present at a locus and “0” to the absence of a band, yielding a matrix consisting of numbers 1 and 0. Such a matrix was used to calculate the genetic distances between any two of the four materials except for Dongfang No. 2 (Nei and Li 1979). Such a matrix was used also to find the bands in Dongfang No. 2 with L. japonica or L. longissima origins or both. The sequences of ITS regions of five materials (1–10, depending on materials) were aligned using MEGA2 program (Version 2.1, http://www.megasoftware.net/).
Daily growth and tissue loss by abrasion
The traits of Dongfang No. 2 were compared with those of L. longissima, L. japonica and Varieties 1 and 2 in the kelp cultivation area of Zhifu Bay, Yantai, Shandong Province. Seawater temperature and transparency were measured frequently. The traits compared included daily growth, daily tissue loss by abrasion, average individual fresh weight, the ratio of fresh-to-dry weight and yield. In total, 24 individuals were measured for growth rate and tissue loss by abrasion.
Fresh weight and ratio of fresh-to-dry weight
Average individual fresh weight was measured with daily growth and daily tissue loss by abrasion. For the calculation of growth rates, the ropes were weighed. Average individual fresh weight was calculated by dividing the total weight of four ropes by the total number of individuals counted each time. One additional rope was selected randomly, and the holdfasts were weighed after removing all blades at the base. Average individual fresh weight=[total weight of four whole ropes − (the weight of the rope with holdfasts × 4)]/the number of total individuals.
Four hanging ropes different from those used for daily growth measuring were sampled, six samples per rope, two each at upper, middle and lower water levels. The blades were air-dried and then dried at 50°C to constant weight. Individual dry weight and individual commercial weight were calculated. For the calculation of the individual commercial dry weight, a water content of 25% in air-dried Laminaria blades was assumed.
Blade length, width, thickness and yield
Three unmarked hanging ropes were selected randomly for each material, and all blades were measured for blade length, width and thickness. Blade width was the maximum width of the blade. For measurements of thickness, discs were taken at three of the five points (except for the tip and blade basis) that divide the blade into four fragments of equal lengths, and all discs from each material were piled up into several stacks to determine their thicknesses; the blade is not thick enough for accurate individual measurements. Blade thickness was the average of all discs of a material. All blades with the discs taken for blade thickness measurement were used for dry weight determination as done for ratio of fresh-to-dry weight described above. Yield = individual dry weight × 1.25 × number of individuals per hectare. In practice, the number of individuals per hectare is assumed to be 180,000. Heterozygous vigor of Dongfang No. 2 = (its trait value − the average of L. longissima and L. japonica)/the average of L. longissima and L. japonica × 100%. Increase in yield = (the yield of Dongfang No. 2 − that of the control)/the yield of the control × 100%. These parameters describe the harvesting characteristics.
Trial cultivation
From 2002 to 2004, large-scale cultivation trials of Dongfang No. 2 were carried out in different sea areas including Zhifu Bay and Taozi Bay of Yantai, Shandong Province, the surrounding sea area of Tuoji Island of Changdao County and Chudao Bay of Rongcheng City of Shandong Province, and Longwangtang sea area of Lushun, Dalian, Liaoling Province. The yield of Dongfang No. 2 obtained in these cultivation trials was then compared to the yield of commercially cultivated Varieties 1 and 2. Sporelings of these three materials were raised in Yantai and also in the hatcheries that cultivate kelp sporelings commercially. Yield was estimated when the blades were ready for harvest. The location of trial cultivation sea areas, method and times of sporeling culturing in open sea and young sporeling transplanting are listed in Table 2.
Table 2.
Time and method of sporeling cultivation and sporeling transplantation to different sea areas
| Location | Date of cultivation | Cultivation method | Date of transplantation | |
|---|---|---|---|---|
| Zhifu Bay | 37°35′N, 120°25′E | 15 Oct | Horizontally placed | 10 Nov |
| Taizi Bay | 37°41′N, 121°09′E | 15 Oct | Horizontally placed | 10 Nov |
| Chudao Bay | 37°01′N, 122°34′E | 20 Oct | Vertically placed | 20 Nov |
| Tuoji Island | 38°08′N, 120°45′E | 8 Oct | Vertically placed | 11–20 Nov |
| Longwangtang | 38°42′N, 121°12′E | 8 Oct | Vertically placed | 11–20 Nov |
Results
Parentage of Dongfang No. 2
Dongfang No. 2 was fingerprinted together with Laminaria longissima, L. japonica and Varieties 1 and 2 with AFLP of total genomic DNA and SNP of the ITS region of rRNA gene transcription unit. Of 440 different bands generated by 16 pairs of selective amplification primers, 167 were shared by Dongfang No. 2, L. japonica and L. longissima, and 48, 38 and 51 were specific to these materials, respectively. Among the remaining 136 bands, 70 were shared by Dongfang No. 2 and L. japonica, 55 by Dongfang No. 2 and L. longissima, and only 11 by L. japonica and L. longissima (Table 3). There were significantly fewer bands shared by L. japonica and L. longissima than by Dongfang No. 2 and L. japonica (70) and Dongfang No. 2 and L. longissima (55), proving that the hybrid kelp originated from these two species. The two species contributed genetic materials almost equally to the hybrid kelp.
Table 3.
Number of bands specific to or shared by different raw materials
| Dongfang No. 2 | L. japonica | L. longissima | |
|---|---|---|---|
| Dongfang No. 2 | 48 | 70 | 55 |
| L. japonica | 38 | 11 | |
| L. longissima | 51 | ||
| No. of bands shared by all | 167 | ||
| Total no. of bands | 440 | ||
We found that nucleotide T at position 847 was specific to L. japonica, while C at this position was specific to L. longissima. Although Variety 1 was related to L. longissima, it held the L. japonica-specific T nucleotide at this position. This SNP-fingerprinted Dongfang No. 2 as the hybrid of L. japonica and L. longissiuma absolutely (Table 4); Dongfang No. 2 holds both nucleotides at this position at the ratio of 1:1.
Table 4.
Variable nucleotides in the ITS region of materials used
| Materials | Nucleotide position at the ITS region | Frequency | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 4 | 7 | 7 | 8 | 8 | 8 | 9 | |||||
| 6 | 6 | 9 | 2 | 6 | 7 | 8 | 0 | 4 | 9 | 4 | ||
| 4 | 5 | 5 | 6 | 7 | 3 | 1 | 3 | 7 | 8 | 9 | ||
| Dongfang No.2 | C | C | T | G | T | T | C | T | T | T | C | 4/10 |
| . | . | . | . | . | . | . | . | C | . | . | 3/10 | |
| . | . | . | . | . | . | G | C | . | . | . | 1/10 | |
| . | . | . | . | . | C | . | . | C | . | . | 1/10 | |
| . | . | C | . | C | . | . | . | C | . | . | 1/10 | |
| L. japonica | . | . | . | . | . | . | . | . | . | . | . | 4/6 |
| T | T | . | . | . | . | . | . | . | . | . | 1/6 | |
| . | . | . | A | . | . | . | . | . | . | . | 1/6 | |
| L. longissima | . | . | . | . | . | . | . | . | C | . | . | 5/6 |
| . | . | . | . | . | . | . | . | C | . | A | 1/6 | |
| Variety 1 | . | . | . | . | . | . | . | . | . | A | . | 1/1 |
| Variety 2 | . | . | . | . | . | . | . | . | . | . | . | 1/1 |
Sequences of ITS region are retrievable from GenBank with accession nos. DQ143048 - DQ143057 (Dongfang No.2), DQ143058 - DQ143063 (L. japonica), DQ143064 - DQ143069 (L. longissima), DQ143070 (Variety 1) and DQ143071 (Variety 2)
In the ITS region, 11 nucleotide positions were found to be polymorphic in Dongfang No. 2, L. japonica, L. longissima and Varieties 1 and 2 (Table 4). L. japonica and L. longissima were not highly divergent; three and two variants were identified, respectively, in them and there existed dominant variants. These variants may come from the gametophytes with different sequences of ITS region; the sporophytes of the two species originated from a collection of gametophytes. It was interesting to note that five variants were found in Dongfang No. 2.
Morphological characteristics
With respect to morphological characteristics, Dongfang No. 2 showed greater similarity to L. japonica than to L. longissima. It combined the desirable characteristics of L. longissima and L. japonica, and showed novel traits they do not have (Table 5).
Table 5.
Morphological characteristics of Dongfang No. 2, L. longissima and L. japonica
| L. longissima | L. japonica | Dongfang No. 2 | |
|---|---|---|---|
| Blade length | Long | Short | Intermediate |
| Blade width | Narrow | Intermediate | Wide |
| Blade thickness | Thin | Intermediate | Thick |
| Color | Deep brown | Deep brown | Deep brown |
| Robustness of holdfast | Not robust | Robust | Very robust |
| Shape of stipe | Thin and flat | Thick and flat | Thick and flat |
| Shape of blade basis | Wedge shaped | Spherical | Imperfectly spherical |
| Vertical groove | Not obvious | Obvious | Obvious |
Daily growth and loss of tissue by abrasion
The rates of daily growth and daily loss of tissue by abrasion of five plant materials and the changes in seawater temperature and seawater transparency are illustrated in Fig. 1. Dongfang No. 2 grew faster than L. japonica and two commercial varieties but slower than L. longissima before 8 April, and faster than L. japonica and Variety 2 but slower than L. longissima and Variety 1 after 8 April. As the fastest, daily growth of Dongfang No. 2 reached 4.7 cm, which was significantly higher than 4.2 cm for Variety 1 and 2.7 cm for Variety 2 (P<0.05). If a daily increase of more than 2.0 cm was considered fast growth, Dongfang No. 2 hybrid maintained fast growth until 14 April, about 20 days less than Variety 1 and 7 days more than Variety 2. The losses of tissue by abrasion of the five plant materials used were lower and were similar to each other before 14 April. The loss of tissue by abrasion of Dongfang No. 2 was similar to the two commercial varieties but significantly lower than L. japonica and L. longissima for the period from 14 April to 25 May. From 14 April to 11 June, seawater transparency increased quickly from 2.7 to 7.0 m and then stayed above 5.7 m, while seawater temperature increased gradually from 8.6 to 18.4°C. The loss of tissue of Dongfang No. 2 by abrasion was close to the two commercial varieties when irradiance became more intense.
Fig. 1.
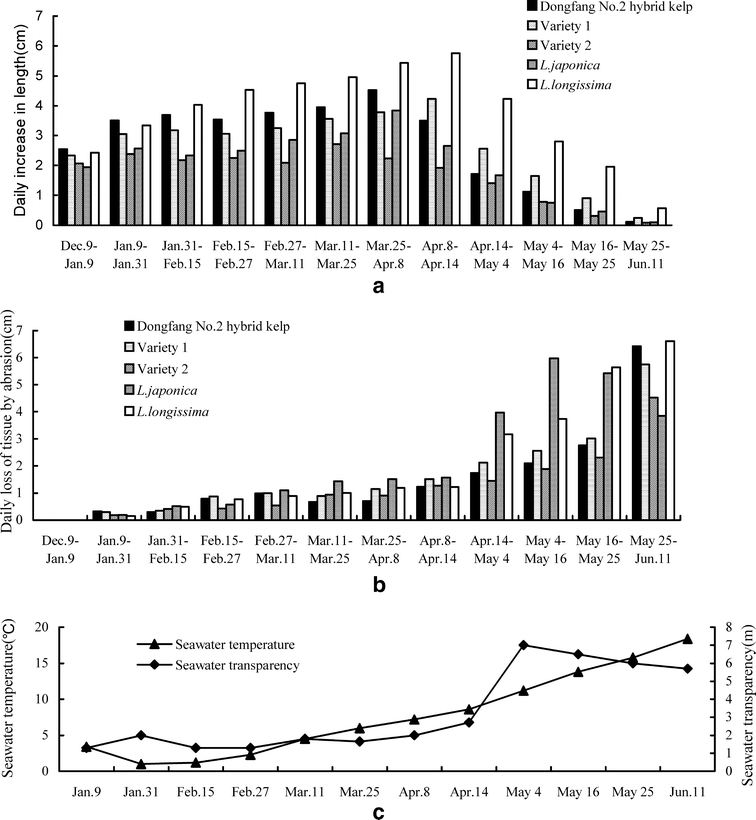
Daily increase in length (a) and daily loss of tissue by abrasion (b) of Dongfang No. 2 hybrid kelp, L. longissima, L. japonica and Varieties 1 and 2, and seawater temperature and transparency changes (c). Data were obtained at Zhifu Bay in 2004
Average fresh weight and fresh-to-dry weight ratio
The dynamics of average fresh weights and fresh-to-dry weight ratios of Dongfang No. 2, Varieties 1 and 2, L. longissima and L. japonica are illustrated in Fig. 2. The average fresh weight of L. japonica decreased after 14 April. After 4 May, the average fresh weights of all the different materials decreased except that of Variety 1. After 25 May, the average fresh weights of all materials decreased. From 4 to 25 May, the average fresh weights of Dongfang No. 2 decreased slowly. From 14 April to 4 May, seawater transparency increased significantly. In parallel, the average fresh weights of L. longissima and L. japonica decreased quickly. In mid-May, at temperatures below 14°C, the blade of Dongfang No. 2 was tough, uniform in thickness at the top and bottom parts and spherical at the base with a fresh-to-dry weight ratio of 6.5:1, implying it had reached its harvesting time. Its harvesting can extend to 16°C (last 10 days of May). Beyond this temperature, fast loss of tissue by abrasion will cause the loss of yield, although fresh-to-dry weight ratio decreases further.
Fig. 2.
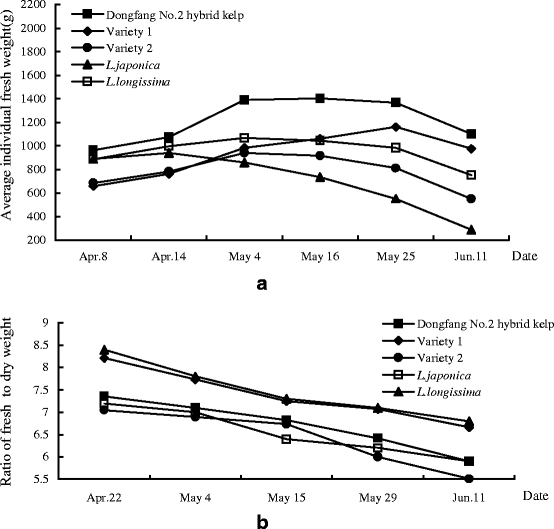
Dynamics of the average individual fresh weight (a) and the ratio of fresh-to-dry weight (b) of Dongfang No. 2 hybrid kelp, L. longissima, L. japonica and Varieties 1 and 2
Yield
The average fresh weight and yield of DongfangNo. 2 and Varieties 1 and 2, respectively, were the highest in 2003, followed by those in 2004 and 2002. These annual changes could be correlated to seawater temperature and transparency, two major environmental factors determining the performance of kelp varieties. In contrast, annual changes in blade length, width and thickness did not follow such a trend. The blade thickness actually changed inversely: the lower the yield, the thicker the blade. In general, average individual fresh weight and yield of Dongfang No. 2 were significantly higher than those of Varieties 1 and 2. The blade width and thickness were also significantly larger than those of Varieties 1 and 2, while the blade length was intermediate between them, significantly shorter than Variety 1 but longer than Variety 2. The yield of Dongfang No. 2 also increased significantly, and displayed significant heterozygous vigor (Table 6).
Table 6.
Performances of Dongfang No. 2 hybrid kelp, L. longissima, L. japonica and Variety 1 and 2 in different economical traits at harvesting time
| Blade lengtha (cm) | Blade width (cm) | Blade thickness (mm) | Individual fresh weight (g) | Yield (kg/ha) | Yield increase by (%) | |
|---|---|---|---|---|---|---|
| L. longissima | 503.8±43.8 | 15.8±1.5 | 1.7 | 1,056 | 26,100 | 41.0 |
| L. japonica | 172.7±15.5 | 21.5±2.0 | 2.1 | 744 | 20,850 | 76.4 |
| Dongfang No.2 hybrid kelp | 395.4±35.2a | 31.9±2.7a | 2.5 | 1,396 | 36,750 | NA |
| 487.8±39.01b (2002) | 32.9±2.7b | 2.5 | 1,173 | 31,200 | NA | |
| 423.9±34.0a, b (2003) | 34.8±3.1a, b | 2.3 | 1,811 | 47,400 | NA | |
| Average | 408.5±51.4e | 33.5±2.9e | 2.4a | 1,575a | 41,400a | NA |
| Variety 1 | 367.6±50.4c | 28.0±3.8c | 1.8 | 1,093 | 27,150 | 35.4 |
| 454.0±64.0c (2002) | 21.1±2.9c | 2.2 | 868 | 22,950 | 36.2 | |
| 486.6±66.5c (2003) | 31.8±4.1c | 1.7 | 1,751 | 39,150 | 21.3 | |
| Average | 441.8±74.3e | 29.1±3.7e | 1.8a | 1,403a | 32,400a | 26.4 |
| Variety 2 | 284.1±48.0d | 25.9±3.4d | 2.1 | 902 | 24,150 | 52.6 |
| 214.7±33.9 d (2003) | 24.3±3.5d | 2.6 | 569 | 15,600 | 99.9 | |
| 273.6±40.8d (2003) | 30.5±4.0d | 2.1 | 990 | 28,350 | 66.9 | |
| Average | 268.7±49.1e | 28.0±4.1e | 2.2a | 900a | 25,050a | 65.0 |
| Heterozygous vigor (%) | 16.9 | 71.0 | 31.6 | 55.1 | 56.8 | NA |
NA Not applicable
Averages are of all individuals (blade length and width) or those of annual measurements (thickness, fresh weight and yield). Data in the same column are different significantly each other if they are marked with the same letter (P<0.05)
a 2002 Data obtained 13 May 2002, 2003 15 May 2003, all others 15 May 2004
In trial cultivation, Dongfang No. 2 increased the yield significantly compared to two commercial varieties. A total of 15, 35 and 75 million sporelings were raised in 2002, 2003, and 2004, respectively, using gametophyte clone-hybridizing and sporeling-raising methods, and used in large-scale cultivation trials in different sea areas scattered throughout Shandong and Liaoling Provinces, China. The total trial cultivation area reached ca. 333 hectares. Unfortunately the yields were not measured in 2002. On average, the yield of Dongfang No. 2 was greater by 26.0 and 62.2% than the two commercial varieties (Table 7).
Table 7.
Yield (kg ha−1) of Dongfang No. 2 from different sea areas in comparison to Varieties 1 and 2
| Zhifu Bay | Taozi Bay | Tuoji Islands | Chudao Bay | Longwangtang | ||||
|---|---|---|---|---|---|---|---|---|
| 2003 | 2004 | 2003 | 2004 | 2003 | 2004 | 2004 | 2004 | |
| Dongfang No. 2 hybrid | 37,500 | 33,000 | 33,000 | 27,000 | 42,000 | 40,500 | 39,000 | 42,000 |
| Variety 1 | 30,750 | 24,750 | 27,000 | 21,000 | 34,500 | 30,750 | 31,500 | 33,750 |
| Variety 2 | 22,500 | 21,000 | 19,500 | 17,250 | 25,500 | 25,500 | 22,500 | 27,750 |
| Increase over Variety 1 (%) | 22.0 | 33.3 | 22.2 | 28.6 | 21.7 | 31.7 | 23.8 | 24.4 |
| Increase over Variety 2 (%) | 66.7 | 57.1 | 69.2 | 56.5 | 64.7 | 58.8 | 73.3 | 51.4 |
Trial cultivation area of Dongfang No. 2 varied depending on location and year, ranging from 6.67–33.33 ha
Variation in Dongfang No. 2 self-bred progenies
Blade lengths and widths of the individuals of L. longissima, L. japonica, and Dongfang No. 2 and its self-bred progenies were measured on 15 May 2004, with the variation indexes [(standard error/average) × 100] listed in Table 8. The index of individual length of Dongfang No. 2 was slightly lower than those of Varieties 1 and 2, but similar to those of L. japonica and L. longissima; this was also true for blade width. In contrast, the variation index in blade length of Dongfang No. 2 self-bred progenies was 4.76-fold higher than that of Dongfang No. 2, 2.77-fold higher than the average of Varieties 1 and 2 and 4.79-fold higher than the average of L. longissima and L. japonica.
Table 8.
Variation indexes of blade length and width of Dongfang No. 2 hybrid kelp and its self-bred progenies, L. longissima, L. japonica and Varieties 1 and 2
| Blade length | Blade width | |
|---|---|---|
| L. longissima | 8.7 | 9.5 |
| L. japonica | 9.0 | 9.3 |
| Variety 1 | 13.7 | 13.6 |
| Variety 2 | 16.9 | 13.1 |
| Dongfang No. 2 hybrids | 8.9 | 8.5 |
| Self-bred progenies | 42.4 | 31.8 |
Comparison of the variation indexes for blade width showed that Dongfang No. 2 self-bred progenies were 3.74-fold higher than Dongfang No. 2, 2.38-fold higher than the average of Varieties 1 and 2, and 3.38-fold higher than the average of L. longissima and L. japonica. It should be noted that the sporelings of L. longissima and L. japonica were raised using a very limited number of gametophyte clones, which may reduce the individual variation in the sporophytes leading to relatively low variation indexes.
The two commercial varieties have been used for many years. Their obvious individual trait variation was reflected in the relatively higher variation indexes compared with Dongfang No. 2, L. longissima and L. japonica. Dongfang No. 2 was the hybrid of two haploid gametophytes and therefore uniform in its genetic background, morphological characteristics and performance. The percentages of individuals of Dongfang No. 2 and its self-bred progenies with different length are illustrated in Fig. 3. As crop hybrids, only the first filial generation can be used in cultivation; the traits of self-bred progenies segregate significantly.
Fig. 3.
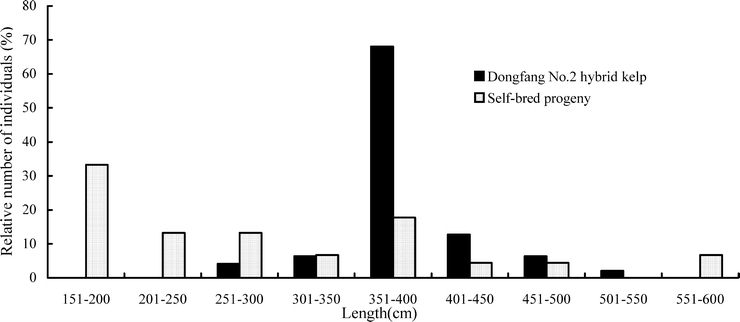
Distributions of blade length of Dongfang No. 2 hybrid kelp and its self-bred progenies
Discussion
Dongfang No. 2, the hybrid of the gametophyte clones from Laminaria japonica and L. longissima, inherited the good characteristics of these two species and expressed novel ones that are desirable for cultivation. It demonstrated very obvious heterozygous vigor (heterosis). The success of breeding this hybrid kelp indicated the feasibility of the direct use of heterosis in economic algae, and also the possibility of significantly improving the yield and other traits of kelp. Its trial cultivation showed that the techniques currently available are effective, although endeavors have to be made to implement these techniques in kelp cultivation.
Varieties 1 and 2 are two representatives of the varieties bred with hybridization and pedigree selection methods. Although Variety 1 was bred through the hybridization of L. japonica and L. longissima, continuous self-crossing, and desirable trait-targeted selection, it may not the best of the possible genotypes. In addition, continuous backcrossing with a recurrent parent, L. japonica, may cause the loss of the genes that determine the high performance of the variety to some extent. At least, the genetic basis of heterosis formation does not exist in Variety 1; it is a genetically pure line selected out from L. japonica. In contrast, Dongfang No. 2 is the hybrid of L. japonica and L. longissima, with its good characteristics distinguishing it from about 50 combinations. It can be expected that Dongfang No. 2 hybrid is better than Variety 1 and Variety 2.
The gametophyte clone hybrids demonstrating heterozygous vigor can be used also as the starting materials of self-crossing and desirable trait-targeted selection (Zhang et al. 1999). As shown, diverse varieties were developed through hybridization of L. longissima and L. japonica gametophyte clones including Danhai No. 1 (Ou et al. 1983) and 901 (Zhang et al. 2001). Due to deep trait segregation of self-bred progenies, only hybrid sporelings can be used in cultivation. However, the large-scale sporeling-raising technique could use more improvement and is not implemented among sporeling raisers. We are a long way from implementing gametophyte-cloned hybrid kelp in cultivation. As an alternative, the hybrid could be used as the starting material for desirable trait-targeted selection as has been done in past breeding practices. Large-scale trial cultivation of Dongfang No. 2 has showed that the protocol used currently for the cultivation of commercial varieties works for the cultivation of Dongfang No. 2.
The distance between sporelings on the hanging rope should be greater than that for other commercially used varieties: the individuals of DongfangNo. 2 are wider than commercial ones. It would be advantageous to change the distance to 8–10 cm. In addition, increased space between sporelings would reduce the number of sporelings needed for seeding the ropes, which, consequently, would reduce overall sporeling-raising cost. It has to be investigated if an increased space between sporelings of Dongfang No. 2 would have a positive effect on yield. In order to promote vegetative maturation, the kelp should be transferred to shallower seawater when seawater temperature rises above 10°C.
Temperature and light are two key factors affecting kelp growth. Relatively low irradiance caused by low transparency in 2004 reduced the growth of Dongfang No. 2. It is certain that a higher yield of Dongfang No. 2 can be achieved under normal conditions as shown in other years. The fast increase in temperature and transparency in the late growth phase of Dongfang No. 2 led to the decrease in growth rate and increase in tissue loss by abrasion, and consequently the loss of yield. The correlation between culturing area conditions and yield was also observed in other varieties cultured in 2004. Dongfang No. 2 showed significant heterozygous vigor in yield, evidence that it is more resistant to stress than other commercial varieties. Heterozygous vigor in yield has been proved also by its performances in large-scale cultivation trials in different sea areas and in different years.
As shown by genetic distance (Table 9), L. japonica and L. longossima are relatively more divergent, and Varieties 1 and 2 are closer to L. japonica than to L. longissima although Variety 1 is related to L. longissima. It is interesting to note that Varieties 1 and 2 are closely related to each other, although their origins are different. This may imply that L. japonica holds more genes adapted to the culturing environments today.
Table 9.
Genetic distances between L. japonica, L. longissima and Varieties 1 and 2
| L. japonica | L. longissima | Variety 1 | |
|---|---|---|---|
| L. japonica | |||
| L. longissima | 0.5936 | ||
| Variety 1 | 0.3771 | 0.5606 | |
| Variety 2 | 0.2667 | 0.5788 | 0.2068 |
AFLP and SNP analyses unambiguously showed that Dongfang No. 2 is a hybrid of L. japonica and L. longissima. AFLP bands of hybrids should theoretically be common to either L. japonica or L. longissima. It is interesting to note that 48 bands were identified as specific to Dongfang No. 2. It can be expected that L. japonica and L. longissima will have a certain number of specific bands; their sporophytes came from a collection of gametophytes. Dongfang No. 2 is from two gametophyte clones, one of each sex. The bands specific to it may originate from mutation or somatic recombination. However, the real mechanism by which these bands formed remains to be studied further.
More variants were found in Dongfang No. 2 in the ITS region of the ribosomal RNA transcription unit, and more interestingly, most of these variants do not exist in L. japonica and L. longissima. Theoretically, the hybrid should have only one or two variants; it is the offspring of two monoploid gametophytes. The reason for the occurance of these variants is not clear. Two possible explanations have been considered: (1) mutation in the vegetative amplification of gametophyte clones, and (2) silencing of the related mutation when two types of ribosomal RNA gene complexes co-exist in a hybrid.
In nature, gametophytes are of microscopic size. Consequently, the production of seeding material requires a high multiplication rate of the gametophyte cells and therefore the potential for mutations may increase and they may be quickly fixed through concerted evolution (Aguilar et al. 1999; Ganley and Scott 2002; Schlotterer and Tautz 1994). Usually, a cell has a thousand copies of ribosomal RNA gene transcription units—the mutation in one of these copies can extend to all members quickly. In addition, the two ITSs in this region may allow mutation to take place at high frequency. All these possible reasons make the mutation accumulate to the extent that it can easily be detect ed as DNA sequence variation. As an alternative, ribosomal RNA gene complex from one parent or from both may be silenced in the hybrid completely or partially, which allows very frequent mutation as occurs in duplicated genes (Pikaard 1999, 2000). These mutations could also be homogenized fast through concerted evolution mechanism. A mutation in the vegetative amplification of gametophyte clones may also explain the appearance of a high proportion of hybrid-specific AFLP bands.
Mutation rate may be high when gametophytes are forced into continuous fast growth. However, this has not been proved yet. Mutation in the vegetative amplification of gametophytes will be investigated at different stages of gametophyte clone amplification. Even if it is true, the breeding and trial cultivation of Dongfang No. 2 demonstrated that the mutation generated by forced growth of gametophytes will not affect the use of the hybrid.
Acknowledgments
This work was supported by The Popularization Project of Agricultural Elite Varieties of Shandong Province (2004113) and National Natural Science Foundation of China (30471318). The authors appreciate the reviewers and editors for their kind and detailed corrections and constructive comments. We thank also editors and reviewers for their patience in processing and reviewing our manuscript and their encouragements. Without their endeavors, this paper would not have come out as it is.
Footnotes
X. Li and Y. Cong contributed equally to this work.
References
- Algal Genetic Research and Breeding Team of Oceanography Institute of Chinese Academy of Science and Algal Culturing Team of Qingdao Maricultural Institute (1976) Breeding of a novel variety with high yield potential and iodine content (in Chinese with English abstract). Sci China 5:512–517
- Aguilar JF, Rossello JA, Feliner GN (1999) Nuclear ribosomal DNA (nrDNA) concerted evolution in natural and artificial hybrids of Armeria (Plumbaginaceae). Mol Biol 8:1341–1346 [DOI] [PubMed]
- Fang TC, Wu CY, Jiang Y, Li TT, Ren GZ (1962) The breeding of a new breed of Haidai (Laminaria japonica Aresch.) and its preliminary genetic analysis (in Chinese with English abstract). Acta Bot Sinica 10(3):197–209
- Fang Z, Ou Y, Cui J, Dai J (1978) Success in culturing clones of the gametophytes of Laminaria japonica (in Chinese with English abstract). Transactions of Science 2:115–116
- Fang Z, Ou Y, Cui J (1983) Experiments on heterosis of Laminaria (in Chinese with English abstract). Mar Sci Bull (China) 2(6):57–60
- Fang Z, Ou Y, Cui J (1985) Breeding of hybrid Laminaria “Danza No.10”: An application of the Laminarian haploid cell clones (in Chinese with English abstract). J Shangdong Coll Oceanol 15(1):64–72
- Ganley ARD, Scott B (2002) Concerted evolution in the ribosomal RNA genes of an Epichloe endophyte hybrid: Comparison between tandemly arranged rDNA and dispersed 5S rRNA genes. Fung Genet Biol 35:39–51 [DOI] [PubMed]
- Li HJ (1990) Notes on the Laminaria raft cultivation method (in Chinese). Mariculture 1990(1/2):41–48
- Li D, Zhou Z, Liu H, Wu C (1999) A new method of Laminaria japonica strain selection and sporeling raising by the use of gametophyte clones. Hydrobiologia 398/399:473–476 [DOI]
- Li D, Lu Y, Wu C (2002) History and current status of kelp breeding and sporeling raising technology (in Chinese). Biol Bull 37(8):1–3
- Li D, Wu C, Liu W, Zang K, Xia J (2003a) A new technology of Laminaria japonica sporeling culture by the use of gametophyte clones (in Chinese with English abstract). Acta Oceanol Sinica 25(5):141–145
- Li M, Yu B, Hu W, Chu C, Xie H, Cui Y (2003b) Study on the large scale raising technique of Laminaria japonica sporelings (in Chinese). Shandong Fisheries 20(9):25–27
- Li Z, Zhang Q, Yang Y, Xie J, Luo S (2003c) Study on the technique of large-scale culture of the clone of Laminaria gametophyte (in Chinese). Shandong Fisheries 20(5):1–3
- Nei M, Li WH (1979) Mathematical model for studying genetic variation for restriction endonuclease. Proc Natl Acad Sci USA 76:5269–5273 [DOI] [PMC free article] [PubMed]
- Ou Y, Cui J, Fang Z (1983) Experiment of Laminaria heterosis (in Chinese with English abstract). Mar Sci Bull 2(6):57–61
- Patwary MU, Ron MM (1993) Revealing genetic markers on Gelidium vagum (Rhodophyta) through the random amplified polymorphic DNA (RAPD) technique. J Phycol 19:216–222 [DOI]
- Pikaard CS (1999) Nucleolar dominance and silencing of transcription. Trends Plant Sci 4(12):478–483 [DOI] [PubMed]
- Pikaard CS (2000) The epigenetics of nucleolar dominance. TIG 16(11):495–500 [DOI] [PubMed]
- Schlotterer C, Tautz D (1994) Chromosomal homogeneity of Drosophila ribosomal DNA arrays suggests intrachromosomal exchanges drive concerted evolution. Curr Biol 4:777–783 [DOI] [PubMed]
- Scoggan S, Zhuang Z, Wang F (1989) Laminaria Seafarming in China: training manual 89/5 (RAS/86/024). Qingdao, China
- Tian Z, Yuan B (1989) Report of the breeding of a novel kelp variety, “Zaohoucheng No.1” (in Chinese). Mariculture 1:7–17
- Tseng CS (2001) Algal biotechnology industries and research activities in China. J Appl Phycol 13:375–380 [DOI]
- Tseng CK, Sun KY, Wu CY (1955) On the cultivation of Haidai (Laminaria japonica Aresch) by summering young sporophytes at low temperature (in Chinese with English abstract). Acta Bot Sinica 4(3):255–264
- Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Homes M, Frijters A, Pot J, Peleman J, Kuiper M, Zabeau M (1995) AFLP: a new technique for DNA fingerprinting. Nucl Acids Res 23:4407–4414 [DOI] [PMC free article] [PubMed]
- Wang S (1994) Seaweed biotechnology (in Chinese). Shanghai Scientific & Technical Publishers, Shanghai, China, pp. 104–111
- Yoon HS, Lee JY, Boo SM, Bhattacharya D (2001) Phylogeny of Alariaceae, Laminariaceae, and Lessoniaceae (Phaeophyceae) based on plastid-encoded RuBisCo spacer and nuclear-encoded ITS sequence comparison. Mol Phylogenet Evol 21:231–243 [DOI] [PubMed]
- Zemke-White WL, Ohno M (1999) World seaweed utilization: An end-of-century summary. J Appl Phycol 11:369–376 [DOI]
- Zhang Z, Fan C, Cao S, You X (1998) Study on the indoor culture and cultivation of Laminaria longissima Mijabe. J Dalian Fish Univ 13(4):1–6
- Zhang Z, Fan C, Cao S, You X, Su Y (1999) Study on interspecific crossing of Laminaria species (in Chinese with English abstract). J Dalian Fish Univ 14(4):13–17
- Zhang Q, Qu S, Xie J, Liu J, Cui F (2000) Experiment of culturing Laminaria longissima in Zhifu Bay. Shandong Fisheries 17(1):10–11
- Zhang Q, Liu S, Qu S, Tian Z, Guo Z, Cong Y, Li Z (2001) Studies on rearing new variety of kelp “901” (in Chinese with English abstract). Trans Oceanol Limnol 2:46–53
- Zhou Z, Wu C (1998) Clone culture of Laminaria japonica and induction of its sporophytes (in Chinese with English abstract). Chinese J Biotechnol 14(1):109–111