

Abstract
Several experimental studies have shown that human grasping behavior exhibits a transition from one-handed to two-handed grasping when to-be-grasped objects become larger and larger. The transition point depends on the relative size of objects measured in terms of human body-scales. Most strikingly, the transitions between the two different behavioral ‘modes’ of grasping exhibit hysteresis. That is, one-to-two hand transitions and two-to-one hand transitions occur at different relative object sizes when objects are scaled up or down in size. In our study we approach body-scaled hysteresis and mode transitions in grasping by exploiting the notion that human behavior in general results from self-organization and satisfies appropriately-defined order parameter equations. To this end, grasping transitions and grasping hysteresis are discussed from a theoretical perspective in analogy to cognitive processes defined by Haken’s neural network model for pattern recognition. In doing so, issues such as the exclusivity of grasping modes, biomechanical constraints, mode-mode interactions, single subject behavior and population behavior are explored.
Keywords: Order parameters, Hysteresis, Grasping
Introduction
Many objects in every-day situations (books, pencils, bags and so on) can be grasped with one or two hands. However, not all large objects can easily be grasped with one hand, and objects that exceed certain critical limit sizes are usually grasped with two hands. That is, the ratio between characteristic object- and body-scales determines whether objects are grasped with one hand or with two hands. In general, if objects are large with respect to characteristic body scales (e.g. the hand size), humans tend to use two hands for grasping. In contrast, if objects are relatively small, humans show the tendency to handle objects with one hand only, as is schematically illustrated in Fig. 1. Consequently, at critical object-body scale ratios, transitions between one-handed and two-handed grasping occur. Several studies have been devoted to examining these so-called body-scaled transitions. For example, body-scaled transitions in grasping have been studied in the field of motor development [1–6]. Body-scaled transitions have also been studied in the context of cognitive processes [7] and social behavior [7, 8]. Studies on body-scaled transitions in general [1–4] have addressed issues regarding the ecological approach to perception and action [9] and in particular [5–7] the actualization of perceived action possibilities, often referred to as affordences [9–12].
Fig. 1.

Schematic illustration (not real data) of a body-scaled transition in grasping objects of different sizes. The control parameter α is typically the ratio between the object size and a characteristic body-scale (e.g. hand size). Y-axis: ‘1H’= one-handed grasping, ‘2H’ = two-handed grasping
The transitions between different behavioral modes of grasping are examples of the behavioral transitions that have been extensively studied in motor control systems [13–29]. Behavioral transitions in motor control systems in turn are examples of transitions that naturally occur in complex non-equilibrium systems [30], as is shown in Fig. 2. Such transitions can be understood using the principles of dynamical systems theory and self-organization [30]. A key ingredient of self-organizing systems are control parameters. Control parameters are certain parameters that induce transitions when they are scaled across critical values, but do not specify the spatio-temporal patterns or behavioral modes that emerge due to these transitions. For self-organized motor-control systems, the control parameters of particular relevance capture the informational variables that specify the different action possibilities available to an actor [9, 10, 12, 31–33]. Indeed, there is a relationship between the behavior of a human actor in a given environment and the information that is available to that human actor in that environment. In some instances, this relationship may assume the form of a mapping [31, 34, 35]. The information variable is mapped to different types of behavioral modes. For behavioral transitions that involve hysteresis (such as grasping transitions, as discussed below), however, such a map must involve a dynamic element (see Section 3) and the relationship between behavior and information can be described in terms of the relationship between behavioral modes and control parameters of dynamical self-organizing systems.
Fig. 2.

Fundamental theoretical framework that will be used in Section 2 to study body-scaled transitions in grasping
Due to the omnipresence of grasping behavior in every-day activities, understanding the dynamics and self-organization of such behavior is important for an understanding of motor control systems in general. However, a dynamical systems approach that studies explicit forms of evolution equations to describe the phenomenon of body-scaled transitions in human grasping has not yet been developed. The aim of the current paper is to provide some initial steps in this direction. To this end, we will present a dynamic system that accounts for three key properties of grasping transitions: hysteresis, exclusivity of behavioral modes, and the distinction between biomechanically induced transitions and transitions induced by interacting behavioral modes.
In several experimental studies it has been shown that grasping behavior shows hysteresis [1, 5, 7]. If object size is increased gradually (ascending condition), the transition from one-handed grasping to two-handed grasping occurs at a particular critical value of a control parameter α representing an object-size to body-scale ratio. If object size is decreased gradually (descending condition), the transition from two-handed grasping to one-handed grasping typically occurs at a different critical value of the control parameter. That is, grasping transitions typically show hysteresis as illustrated schematically in Fig. 3. Typically, the critical control parameter of the one-to-two hand transition in the ascending condition is larger than the critical parameter of the two-to-one hand transition in the descending condition (for exceptions, see, for example, [7, 36]). The question arises as to what can be inferred from the observation of hysteresis? To what extent does the phenomenon of hysteresis provide us with insights into the structure of the dynamical systems underlying grasping transitions?
Fig. 3.

Illustration of hysteresis in transitions between unimanual and bimanual grasping (not real data). (o) Ascending condition (object size is gradually increased) and (+) descending condition (object size is gradually decreased)
Grasping transitions involve two mutually exclusive behavioral modes: grasping with one hand and grasping with two hands. That is, there is not a continuous spectrum of grasping modes that is bounded on either side by the one-handed and two-handed grasps, respectively. For example, there does not exist a 1.5-handed grasping behavior. Grasping transitions confront us with the question of how to approach the exclusivity of grasping modes from a dynamical systems perspective.
Grasping objects with one hand becomes impossible for objects that are simply too large to be picked up with one hand. Consequently, grasping transitions from one-handed grasps to two-handed grasps may be induced by biomechanical limitations. The argument of bio-mechanical limitations, however, does not necessarily apply to grasping transitions from two-handed to one-handed grasps as observed under the aforementioned descending object presentation condition. Relatively small objects such as pencils can be picked up with two hands although adult participants in an experimental study might feel uncomfortable doing so. The two-to-one hand transitions observed in experimental studies suggest that there is a second process that triggers grasping transitions. We will argue in Sections 2 and 3 that this second process reflects interactions between unimanual and bimanual grasping modes. In short, when discussing grasping transitions from a theoretical perspective, we need to take two different types of processes into consideration. First, one of the two grasping modes may become unavailable due to biomechanical constraints. In this context, we would deal with a transition due to biomechanical limitations. Second, both behavioral modes are available and interact with each other. In this context, we would deal with transitions involving a mode selection. How can these two processes be rephrased in the language of dynamical systems theory? How can these two types of transitions be related to attractors and repellers of a dynamical system underlying grasping transitions?
In the subsequent sections, we will study grasping transitions from a dynamical systems perspective by adopting a model proposed by [37] (see also [38–40]). Haken’s model is tailored to describe pattern recognition processes (e.g. the recognition of faces and scenes). In particular, it has been applied to study oscillatory perceptual processes centered on ambiguous patterns [41]. The model proposed by Haken has often been interpreted as a neural network model (see, for example [42–44]). In this context, states of neural activity are determined by attractors and repellers of the neural system of humans and animals. In our context, we need to take a slightly different perspective. That is, the dynamical system involved in grasping transitions must incorporate both the individual and the environment. More specifically, the dynamical system involved in grasping transitions must incorporate the complementary relation that exists between the relevant properties of the to-be-grasped objects (i.e. object-length or size) and the relevant properties of the individual’s action system (i.e. hand size). It is this animal-environment relation (more precisely, the information that specifies this animal-environment relation) that constrains and guides the grasping behavior of an individual [9, 10, 32]. The attractors and the repellers that we will discuss in our study do not exist exclusively within the body or brain of an individual; rather, they exist in the animal-environment system composed of the individual and its environment [45, 46].
With Haken’s model at hand, we will address in detail the three key issues mentioned above. In Section 2.1 we will show that the observation of hysteresis suggests that the dynamical system involved in grasping transitions exhibits a bistable parameter domain. We will show that the exclusivity of behavioral modes naturally results from a winner-takes-all dynamics as described by Haken’s model for pattern recognition. We will study fixed point attractors that vanish entirely and discuss attractors that become repellers when control parameters are varied. We will show to what extent these two types of bifurcations are related to the two above-mentioned processes that may result in grasping transitions. While Section 2.1 is concerned with individual behavior, in Section 2.2, predictions for participant populations will be derived. In particular, we will demonstrate how to derive, from our dynamical systems approach, the hysteresis curves for participant populations that have been reported in several studies [4, 5, 7].
Haken’s model of pattern recognition applied to grasping transitions
In our study, we will restrict our analysis to the order parameter dynamics of Haken’s model for pattern recognition [37].
Individual participant behavior
Let ξ1 and ξ2 denote the generalized amplitudes of one-handed and two-handed grasping movements, respectively. Consequently, if ξ1 is finite and ξ2 equals zero, then one-handed grasps are performed. In contrast, if we have ξ2 > 0 and ξ1 = 0 then two-handed grasps are performed. In general, ξ1 and ξ2 are so-called order parameters that emerge close to bifurcation points and evolve with time. Accordingly, grasping behavior is determined by the evolution of the order parameters ξ1 and ξ2. In line with Haken’s model for pattern recognition, we assume that the order parameter dynamics satisfies the coupled set of nonlinear first-order dynamical equations [37, Section 5.2, Eq. 5.33]:
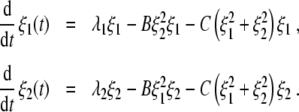 |
1 |
Note that in what follows we will consider only positive order parameters ξ1,ξ2 ≥ 0. The linear terms involve the parameters λ1,λ2 that can assume both positive and negative values. λ1 and λ2 correspond to the growth rates of the amplitudes ξ1 and ξ2, if ξ1 and ξ2 are close to zero. Consequently, the order parameters can only grow from a state ξ1,2 ≈ 0 to ξ1 > 0 or ξ2 > 0 if at least one of the λ parameters is positive. In the context of Haken’s model for pattern recognition the parameters λ1 and λ2 have been called attention parameters because a pattern can be recognized only if its corresponding λ parameter is positive [37, Section 5.2]. Accordingly, λ > 0 means that attention is directed to a pattern, whereas λ < 0 means that no attention is paid to the pattern. In our study, we will refer to λ1 and λ2 as attention parameters as well, in order to be consistent with the literature. However, we will not necessarily interpret them as parameters that express a degree of attention (see Section 2.1.8). In line with the study by [41] on oscillatory perceptual instabilities, we assume that in general the attention parameters can differ from each other. The nonlinear terms involve two parameters B, C > 0. Equation 1 can equivalently be expressed in terms of the potential dynamics
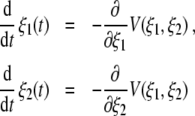 |
2 |
involving the two-dimensional potential
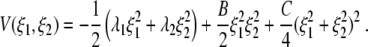 |
3 |
Note that in our study we will not evaluate the magnitudes of ξ1 and ξ2. For example, the states (ξ1,ξ2) = (1,0) and (ξ1,ξ2) = (2,0) both describe one-handed grasping. We will focus on the issue of whether a behavioral mode is present (e.g. ξ1 > 0) or absent (e.g. ξ1 = 0). We will briefly return to the relevance of the magnitudes of the order parameters ξ1 and ξ2 in Section 3.
Fixed points, attractors and repellers
Equation 1 exhibits, in general, up to four fixed points (ξ1,st,ξ2, st) that will be denoted by I, II, III, and IV (here ‘st’ refers to ‘stationary’). Fixed point I is given by
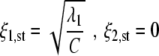 |
4 |
and describes one-handed grasping activity. Fixed point II assumes the form
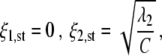 |
5 |
and describes two-handed grasps. Fixed point III satisfies
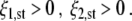 |
6 |
This fixed point describes the co-existence of the one- and two-handed grasping modes. Since the grasping modes are mutually exclusive, such a co-existence is forbidden. We will show below to what extent the dynamical system (1) indeed does not exhibit such states of co-existence, although, under particular parameter conditions, fixed point III is a solution of Eq. 1. Fixed point IV is the origin, given by
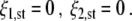 |
7 |
This fixed point describes the absence of grasping activity. Let us discuss next the existence and stability of the fixed points I − IV. To this end, we will proceed along the lines of the discussion carried out in [37].
Case λ1 > 0, λ2 ≤ 0
In this case fixed points I and IV exist. Fixed points II and III do not exist. A linear stability analysis shows that fixed point I represents a stable node. That is, there is an attractor of the one-handed grasping mode. The attractor corresponds to a minimum of the potential V as illustrated in Fig. 4. Using linear stability analysis it can be shown that the origin (fixed point IV) corresponds to an unstable node. That is, the no-action-at-all mode corresponds to a repeller. As can be seen from Fig. 4 this repeller corresponds to a maximum of the potential V. Consequently, for any initial condition ξ1(0) > 0, ξ2(0) > 0 the dynamical system (1) converges to the fixed point I, indicating that under the condition λ1 > 0, λ2 ≤ 0 only one-handed grasps are performed.
Fig. 4.

Case λ1 > 0, λ2 ≤ 0. The potential (3) exhibits a minimum at the fixed point I defined by Eq. 4. The amplitude (order parameter) of the one-handed grasping mode is finite, whereas the amplitude (order parameter) of the two-handed grasping mode vanishes. Note that in this figure and in all subsequent figures we will present the potential V in the full two-dimensional plane spanned by ξ1 and ξ2 which improves the overall visibility of the potential function. In our study the focus is on the part of this plane given by the sub-domain ξ1 ≥ 0 and ξ2 ≥ 0
Case λ1 ≤ 0, λ2 > 0
Under these conditions, fixed points II and IV exist, whereas fixed points I and III do not exist. By means of linear stability analysis we can show that fixed point II is a stable node. Fixed point IV corresponds to an unstable node. The dynamical system (1) converges for any initial condition ξ1(0) > 0 and ξ2(0) > 0 to the fixed point II. Consequently, only two-handed grasps are performed.
Case λ1 > 0, λ2 > 0
The fixed points I, II and IV exist for any pair (λ1,λ2) with λ1 > 0 and λ2 > 0. The existence of fixed point III will be discussed separately below. Linear stability analysis shows that fixed point IV is an unstable node. In order to examine fixed point I by means of linear stability analysis we put ξ1 = ξ1,st + ε1, ξ2 = ε2, where ε1 and ε2 correspond to small perturbations. Substituting these relations into Eq. 1, we see that the perturbations evolve as
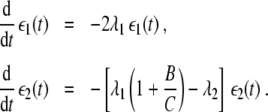 |
8 |
At this stage we introduce the parameter
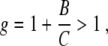 |
9 |
that will be used frequently in subsequent steps of our analysis. From Eqs. 8 and 9 it follows that for λ1g > λ2 we are dealing with a stable node I, whereas for λ1g < λ2 fixed point I is unstable and corresponds to a saddle point. Likewise, let us examine the stability of fixed point II by introducing the perturbations ε1 and ε2 defined by ξ1 = ε1 and ξ2 = ξ2,st + ε2. From Eq. 1 we obtain
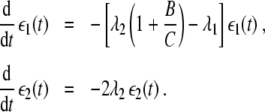 |
10 |
Consequently, for λ2g > λ1 we find that fixed point II is a stable node, whereas for λ2g < λ1 fixed point II corresponds to a saddle point. Figure 5 illustrates this latter case: λ2 is larger than zero, but the dynamical system (1) is still monostable because we have λ2g < λ1.
Fig. 5.

Illustration of the potential V(ξ1,ξ2) for λ1, λ2 > 0 and λ2g < λ1. Fixed point I representing the one-handed grasping mode corresponds to a potential minimum, whereas fixed point II representing the two-handed grasping mode corresponds to a saddle point. The shape of the potential function implies that according to the potential dynamics (1) only one-handed grasps can be performed
We may summarize the results that we have derived so far by assuming that λ1 > 0 is fixed and that λ2 is increased gradually. Then, we see that the dynamical system (1) defined on ξ1 ≥ 0 and ξ2 ≥ 0 is monostable for λ2/λ1 < 1/g and exhibits a unique stable attractor given by fixed point I. The system (1) is bistable for 1/g < λ2/λ1 < g and exhibits two attractors related to the fixed points I and II. The dynamic system becomes monostable again for λ2/λ1 > g and exhibits a unique stable attractor given by the fixed point II. In the monostable parameter domains the system converges to its uniquely defined stable fixed point for arbitrary initial conditions ξ1(0), ξ2(0) > 0. This implies that only one of the two grasping modes can be performed. In the bistable parameter domain the system converges to either of the two attractors. That is, the system may converge to the fixed point I reflecting the one-handed grasping mode or to the fixed point II reflecting the two-handed grasping mode. Both grasping modes are available. Which mode is selected depends on the initial conditions. Figure 6 illustrates the potential V(ξ1,ξ2) for the bistable case. The attractors at (ξ1,st > 0,ξ2,st = 0) (fixed point I) and (ξ1,st = 0,ξ2,st > 0) (fixed point II) correspond to potential minima of V.
Fig. 6.

Potential function V(ξ1,ξ2) in the bistable parameter domain characterized by the inequalities 1/g < λ2/λ1 < g. The dynamics settles down in either of the two attractors at (ξ1,st > 0,ξ2,st = 0) or (ξ1,st = 0, ξ2,st > 0) depending on the initial condition (ξ1(0),ξ2(0)). Consequently, the system (1) predicts that in the bistable parameter domain both one-handed grasps and two-handed grasps can be performed
Co-existence in the bistable parameter domain?
In the preceding analysis we neglected the fixed point III. From Eq. 1 it follows that fixed point III is defined by
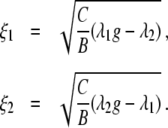 |
11 |
Equation 11 reveals that fixed point III exits only in the bistable parameter domain. In the parameter space ξ1-ξ2 this bistable domain is given by the hatched area shown in Fig. 7.
Fig. 7.

Illustration of the bistable domain (hatched area) defined by 1/g < λ2/λ1 < g in the two-dimensional parameter space ξ1-ξ2. Note that the size of the area depends on the parameter g defined by Eq. 9, which in turn depends on the ratio B/C. For B/C →0 the hatched area vanishes and reduces to the diagonal. In contrast, for B/C ≫ 1 the hatched area would basically correspond to the entire first and third quadrants
In order to determine the character of the fixed point III we first note that within the bistable parameter domain the number of fixed points does not change. Likewise, the stability of the fixed points I, II, and IV does not change. From these two properties and from the fact that the potential V depends in a continuous way on the attention parameters λ1 and λ2, we conclude that the stability of the fixed point III cannot change within the bistable parameter domain. In other words, the topology of the potential V does not depend on the parameters λ1 and λ2 as long as they are selected from the bistable parameter domain (i.e. as long as they satisfy the inequality 1/g < λ2/λ1 < g). From this argument it follows that the fixed point III for the special case λ1 = λ2 exhibits the same stability as for all other attention parameters satisfying 1/g < λ2/λ1 < g. For the special case λ1 = λ2 it has been shown that the fixed point III corresponds to a saddle point [37]. Consequently, if fixed point III exists, then it is an unstable fixed point that corresponds to a saddle point of the potential V(ξ1,ξ2). Fixed point III corresponds to the saddle between the potential minima at (ξ1 > 0,ξ2 = 0) and (ξ1 = 0,ξ2 > 0) as illustrated in Fig. 6. With this result at hand, we can conclude our discussion on the existence and stability of the fixed points of Eq. 1. Table 1 summarizes our results.
Table 1.
| λ2/λ1 < 1/g | 1/g < λ2/λ1 < g | λ2/λ1 > g | ||
|---|---|---|---|---|
| I | stable | stable | unstable | 1H grasping mode |
| II | unstable | stable | stable | 2H grasping mode |
| III | – | unstable | – | |
| IV | unstable | unstable | unstable | no action |
Bistability and hysteresis
Since Eq. 1 exhibits a bistable parameter domain and satisfies a potential dynamics involving the potential function V(ξ1,ξ2) we conclude that Eq. 1 features hysteresis. If we gradually increase the ratio λ2/λ1, then from Table 1 we can read off that for λ2/λ1 < g the dynamical system (1) will settle down to the fixed point I irrespective of its initial condition. Accordingly, participants perform one-handed grasping movements for λ2/λ1 < g. If the ratio λ2/λ1 is increased further such that the system operates in its bistable parameter domain, then the system dynamics will not leave the attractor I. Only if λ2/λ1 is increased beyond the threshold λ2/λ1 = g, the system will settle down to the attractor of fixed point II. Accordingly, the one-handed grasping mode vanishes and the two-handed grasping mode emerges. The transition from one- to two-handed grasps occurs at λ2/λ1 = g. We can identify this transition with the grasping transition in the ascending condition (see Fig. 3). In contrast, if we gradually decrease the ratio λ2/λ1 starting with a value λ2/λ1 > g, then the system will settle down initially to the attractor of fixed point II and will remain there until the critical threshold λ2/λ1 = 1/g is reached. That is, Eq. 1 predicts that the two-handed grasping mode is active during the descending condition as long as λ2/λ1 > 1/g. At λ2/λ1 = 1/g the two-handed grasping mode vanishes and the one-handed grasping mode emerges (see descending condition in Fig. 3). In short, Eq. 1 predicts that the one-to-two hand grasping transition occurs at λ2/λ1 = g in the ascending condition, whereas the two-to-one hand grasping transition occurs at λ2/λ1 = 1/g in the descending condition.
Winner-takes-all dynamics and exclusivity of grasping modes
From our previous discussion it follows that the dynamical system (1) does not exhibit a stable fixed point III. That is, Eq. 1 does not exhibit a state in which the amplitudes ξ1 and ξ2 of the two grasping modes are both finite. In other words, one mode dominates the other mode. We may refer to the dominant mode as the ‘winner’ mode. The winner mode takes all activity and ‘inhibits’ the other mode completely. Consequently, the winner-takes-all dynamics of Eq. 1 reflects the exclusivity of grasping modes.
Sign of attention parameters and two types of grasping transitions
Figures 4 and 5 illustrate that there are two types of monostable parameter domains. There are two monostable parameter domains (defined by λ1 ≤ 0 or λ2 ≤ 0) for which the dynamical system (1) exhibits only one fixed point different from the origin, as exemplified by Fig. 4. In addition, there are two monostable parameter domains (defined by λ1,λ2 > 0 and either λ1 > gλ2 or λ2 > gλ1 ) for which the system (1) exhibits two fixed points different from the origin, but only one of the two corresponds to an attractor (see, for example, Fig. 5). This implies that we can distinguish between two cases. In the first case there is only one grasping mode available at all. In the second case there are two grasping modes available. However, only one amplitude of the two mode amplitudes has an attractor. The other mode amplitude has a saddle point. We can identify the first case with experimental conditions under which a particular grasping mode cannot be performed at all. As mentioned in the introduction, there are instances in which larger objects simply can not be grasped with one hand due to biomechanical limitations. Monostability in the second case arises from an interaction between the one- and two-handed grasping modes. In terms of the attention parameters λ1 and λ2, we may say that if a grasping mode is physically forbidden (i.e. can not be performed due to biomechanical constraints), then the mode exhibits a negative attention parameter. For example, if we continously increase the size of an object then the attention parameter λ1 of the one-handed grasping mode as a function of the object size will become negative at a certain critical object size. However, if potentially both grasping modes are available (if they are consistent with biomechanical constraints) but only one grasping mode is selected for all possible initial conditions, then both attention parameters are positive and one attention parameter is critically larger than the other (i.e. we have either λ1 > gλ2 or λ2 > gλ1) such that the underlying dynamics (1) is in a monostable parameter domain. Consequently, the two processes mentioned in the introduction that can induce transitions between grasping modes can be classified in terms of attention parameters. A transition from one grasping mode to another mode due to biomechanical limitations is related to an attention parameter that becomes negative. A transition from one grasping mode to another mode due to mode-mode interactions occurs when one attention parameter becomes critically larger than the other in the sense that one of the inequalities λ1 > gλ2 and λ2 > gλ1 is satisfied.
Object-body ratio (control parameter) and attention parameters
In order to relate Haken’s model (1) closer to experimental findings, we need to introduce the control parameter α typically used in experimental studies on grasping transitions. As mentioned earlier, the control parameter α reflects typically the ratio between the size of an object and a characteristic body-scale. For example, α was defined as the ratio of the size of the to-be-grasped object and the hand size of a participant (see e.g. [4, 7]). In general, all parameters λ1,λ2, B, C occurring in Eq. 1 may depend on this control parameter α. In this section, we will assume that the parameters B and C describing the magnitudes of the system nonlinearities do not depend on α (we will briefly address the more general case in the discussion, see Section 3). We are left to discuss the functions λ1(α) and λ2(α). From the empirically observed transition in the ascending and descending conditions, we conclude that the attention parameter λ1 of the one-handed grasping mode decreases with α, whereas the attention parameter λ2 of the two-handed mode increases with α. The most parsimonious approach is to linearize the functions λ1(α) and λ2(α) such that we obtain
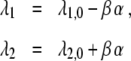 |
12 |
with β > 0 and λ1,0 > λ2,0. From Eq. 12 and the critical thresholds λ2/λ1 = 1/g and λ2/λ1 = g, we can derive the critical control parameters:
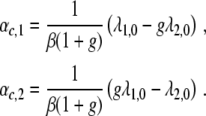 |
13 |
The critical control parameter αc,1 corresponds to the case λ2/λ1 = 1/g, whereas αc,2 reflects the case λ2/λ1 = g. It can be shown that αc,2 > αc,1. Figure 8 illustrates the linear functions λ1(α) and λ2(α) as well as the critical control parameters αc,1 and αc,2.
Fig. 8.

Attention parameters λ1 (thin decreasing line) and λ2 (thick increasing line) as functions of α, computed from Eq. 12. Critical control parameters αc,1 and αc,2 (see Eq. 13) are indicated by vertical lines (o) and (+), respectively. Parameters: B = C = 1, λ1,0 = 1, λ2,0 = 0, β = 1 (which implies g = 2, αc,1 = 1/3, αc,2 = 2/3)
Using the linear functions λ1(α) and λ2(α) defined by Eq. 12 in combination with Eq. 1, we can derive the stationary order parameters ξ1,st and ξ2,st as functions of α for the ascending and descending conditions. Figure 9 shows the functions ξ1,st(α) and ξ2,st(α) as obtained by solving Eqs. 1 and 12 numerically. In line with our analytical discussion, we see that in the ascending condition the amplitude ξ1,st of the one-handed grasping mode is finite and the amplitude ξ2,st of the two-handed grasping mode vanishes for α < αc,2, whereas for α > αc,2 we have ξ1,st = 0 and ξ2,st > 0. That is, the one-to-two hand transition occurs at αc,2 (see left panel in Fig. 9). In contrast, in the descending condition the order parameter ξ2,st of the two-handed grasping mode is finite, and the order parameter ξ1,st of the one-handed grasping mode vanishes for α > αc,1. For α < αc,1 we have ξ1,st > 0 and ξ2,st = 0. That is, the two-to-one hand transition occurs at αc,1 (see right panel in Fig. 9).
Fig. 9.

Stationary order parameters ξ1 (thin line) and ξ2 (thick line) versus α obtained by solving numerically the system (1) for the linearized attention parameter functions (12). In the ascending case (panel a) α was increased in steps Δα. The order parameter ξ1 of the one-handed mode vanished and the order parameter ξ2 of the two-handed mode emerged at the theoretically predicted value αc,2. In the descending case (panel b) α was decreased in steps Δα and the two-to-one hand transition was observed at αc,1. Parameters are defined as in Fig. 8 and Δα = 0.01. Equation 1 was solved using an forward Euler method with Δt = 0.1. Total number of iterations for every α: 10000
Participant populations and mean performance of individuals
Participant populations
Individual participants show transitions between grasping modes at different critical control parameters (see, for example, [5]). In order to account for this kind of individuality, we assume that in general the parameters B, C, β, λ1,0, and λ2,0 of Eqs. 1 and 12 differ slightly among participants. Consequently, in a population of participants the parameters αc,1 and αc,2 defined by Eq. 13 will be distributed according to a certain law. For sake of simplicity, we assume that αc,1 and αc,2 are Gaussian distributed around some mean values m1 and m2, respectively. That is, we consider probability densities P1(α1,c) and P2(α2,c) of the critical control parameters αc,1 and αc,2 defined by
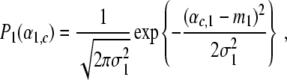 |
14 |
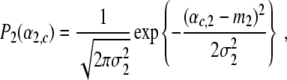 |
15 |
where 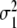 and
and 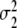 denote the variances of the parameters αc,1 and αc,2, as shown in Fig. 10. In order to compute the percentage of participants that perform two-handed grasps given a certain control parameter α in the ascending condition, we need to compute the probability p (αc,2 < α) to find critical control parameters αc,2 smaller than α. The probability p (αc,2 < α) can be computed from
denote the variances of the parameters αc,1 and αc,2, as shown in Fig. 10. In order to compute the percentage of participants that perform two-handed grasps given a certain control parameter α in the ascending condition, we need to compute the probability p (αc,2 < α) to find critical control parameters αc,2 smaller than α. The probability p (αc,2 < α) can be computed from
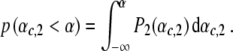 |
16 |
Fig. 10.

Probability densities of critical control parameters αc for the ascending (solid line) and descending condition (dashed line) as computed from Eqs. 14 and 15. Parameters: m1 = 1/3, m2 = 2/3, 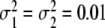
Using 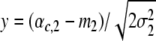 , we obtain
, we obtain
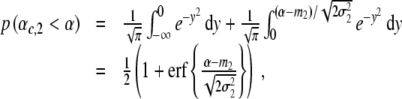 |
17 |
where the error-function 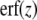 is defined by
is defined by 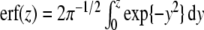 . Likewise, we can compute the probability p (αc,1 < α) to find participants that perform two-handed grasp in the descending condition given a certain control parameter α:
. Likewise, we can compute the probability p (αc,1 < α) to find participants that perform two-handed grasp in the descending condition given a certain control parameter α:
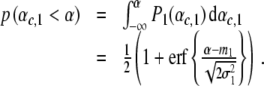 |
18 |
As argued earlier, p (αc,2 < α) describes the probability of finding participants that perform two-handed grasping movements in the ascending condition. Consequently, p 2H,ascend(α) = 100 p (αc,2 < α) is the percentage of participants that perform two-handed grasps in the ascending condition. Likewise, p 2H,descend(α) = 100 p (αc,1 < α) is the percentage of participants that perform two-handed grasps in the descending condition. We have
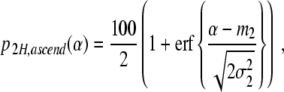 |
19 |
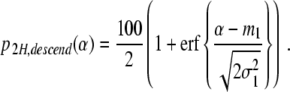 |
20 |
Let p 1H,ascend(α) and p 1H,descend(α) denote the percentages of participants performing unimanual grasps in the ascending and descending condition, respectively. From p 1H,ascend(α) + p 2H,ascend(α) = 100 and p 1H,descend(α) + p 2H,descend(α) = 100 it follows that
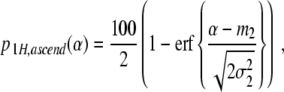 |
21 |
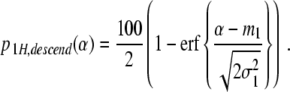 |
22 |
Figure 11 depicts the functions (19,...,22) for particular values of 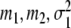 and
and 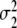 . The parameters m1 and m2 determine 50 percent levels under ascending and descending conditions. These 50 percent levels have been interpreted as critical transition parameters for participant populations [7]. The variances
. The parameters m1 and m2 determine 50 percent levels under ascending and descending conditions. These 50 percent levels have been interpreted as critical transition parameters for participant populations [7]. The variances 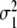 and
and 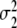 determine the slope of the functions p 1H/2H,ascend(α) and p 1H/2H,descend(α) at α = m1 and α = m2, respectively. For example, for large values the functions p 2H,ascend(α) and p 2H,descend(α) increase slowly, whereas for very small values the functions p 2H,ascend(α) and p 2H,descend(α) jump from 0 to 100 at α = m1 and α = m2. In Fig. 11 the case
determine the slope of the functions p 1H/2H,ascend(α) and p 1H/2H,descend(α) at α = m1 and α = m2, respectively. For example, for large values the functions p 2H,ascend(α) and p 2H,descend(α) increase slowly, whereas for very small values the functions p 2H,ascend(α) and p 2H,descend(α) jump from 0 to 100 at α = m1 and α = m2. In Fig. 11 the case 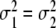 and m2 > m1 is shown. Population results p 1H/2H,ascend(α) and p 1H/2H,descend(α) have been reported in several empirical studies [4, 5, 7]. The empirical graphs presented in these studies correspond qualitatively to the functions p 1H/2H,ascend(α) and p 1H/2H,descend(α) defined by Eqs. 19, ..., 22.
and m2 > m1 is shown. Population results p 1H/2H,ascend(α) and p 1H/2H,descend(α) have been reported in several empirical studies [4, 5, 7]. The empirical graphs presented in these studies correspond qualitatively to the functions p 1H/2H,ascend(α) and p 1H/2H,descend(α) defined by Eqs. 19, ..., 22.
Fig. 11.

Percentages of one-handed grasps (panel a) and two-handed grasps (panel b) as functions of the control parameter α for the ascending (solid lines) and descending condition (dashed lines) as computed from Eqs. 19, ..., 22. Parameters as used in Fig. 10
Mean performance of individuals
In Section 2.2.1, we derived the functions p 1H/2H,ascend(α) and p 1H/2H,descend(α) under the assumption that the parameters of our system (1) and (12) vary between participants. The technical arguments made in Section 2.2.1 remain valid if we assume alternatively that the system parameters vary between trials performed by a single individual. In this case, we can re-interpret the function p 1H/2H,ascend(α) and p 1H/2H,descend(α) as describing the mean grasping behavior of individuals. In fact, in [1] sigmoidal functions for individual participants have been reported that correspond qualitatively to the functions p 1H/2H,ascend(α) and p 1H/2H,descend(α) defined by Eqs. 19, ..., 22.
Discussion
We studied body-scaled transitions in human grasping from a theoretical perspective using a dynamical system defined by Eq. 1 that was originally proposed in [37]. Equation 1 describes the evolution of the amplitudes (or order parameters) ξ1 and ξ2 of unimanual and bimanual grasping modes. If an order parameter becomes finite, then the corresponding grasping mode emerges. If an order parameter vanishes, then the corresponding grasping mode vanishes as well. Our analysis showed that the system (1) accounts for several experimental findings: the hysteresis in grasping transitions, the exclusivity of grasping modes, the interaction between grasping modes, and the existence of biomechanical constraints that make the performance of certain grasping modes (such as the unimanual mode) impossible.
Our analysis suggests that hysteresis of grasping transitions results from the bistability of the underlying order parameter dynamics. That is, although the dynamics described in Eq. 1 should be regarded as an approximation of the actual order parameter dynamics of grasping, our analysis suggests that the actual order parameter dynamics will also involve a bistable parameter domain that gives rise to the hysteresis patterns observed in experimental studies.
By means of Eq. 1 we demonstrated that a winner-takes-all dynamics is tailored to describe transitions between mutually exclusive grasping modes. Again, we may generalize this result by stating that human-environmental systems that feature mutually exclusive behavioral patterns are likely to feature an underlying winner-takes-all dynamics. The winner-takes-all dynamics may be approximately described by Eq. 1 or by another winner-takes-all model.
We exploited the attention parameters λ1 and λ2 of Eq. 1 in order to distinguish between biomechanical limitations and the mode-mode interactions that may result in grasping transitions. If an attention parameter of a grasping mode becomes negative, then a stationary finite order parameter of that mode no longer exists because the fixed point of the mode entirely vanishes. In this sense, a mode with a negative attention parameter is no longer part of the repertoire of available behavioral patterns. For this reason we interpreted in Section 2.1.8 negative attention parameters as counterparts to biomechanical constraints that make the performance of behavioral patterns impossible. For example, if we ask a participant to grasp a cube with index finger and thumb, then the participant simply can not perform this task with one hand if the size of the cube exceeds the span between index finger and thumb. This experimental condition would correspond to a negative attention parameter λ1 for the unimanual grasping mode. In contrast, if an attention parameter is positive but critically smaller than the other attention parameter, then the corresponding grasping mode is available but unstable. In this case due to mode-mode interactions a mode cannot be performed although it is part of the repertoire of available behavioral patterns. For example, in the descending condition a transition from bimanual to unimanual grasping occurs as illustrated schematically in Fig. 3. However, if we would instruct participants in the descending condition to grasp objects always with two hands even if the objects become relatively small, the transition would not occur or would be shifted to very small object sizes. In view of Eq. 1 the instruction to use bimanual grasps would imply that the order parameter ξ1 is fixed to zero. Under the condition ξ1 = 0, Eq. 1 reduces to 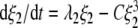 and the two-to-one hand transition for λ2 > 0 shown in Fig. 9 (panel b) would disappear. This illustrates that if ξ1 is not fixed to zero, the two-to-one hand transition at λ2 = λ1/g > 0 shown in Fig. 9 (panel b) is due to a mode-mode interaction. Since human behavior is frequently subjected to biomechanical constraints, dynamical systems approaches to human behavior in general should account for the aforementioned two types of transitions. Equation 1 helps us to illustrate explicitly in the context of grasping transitions how this can be done.
and the two-to-one hand transition for λ2 > 0 shown in Fig. 9 (panel b) would disappear. This illustrates that if ξ1 is not fixed to zero, the two-to-one hand transition at λ2 = λ1/g > 0 shown in Fig. 9 (panel b) is due to a mode-mode interaction. Since human behavior is frequently subjected to biomechanical constraints, dynamical systems approaches to human behavior in general should account for the aforementioned two types of transitions. Equation 1 helps us to illustrate explicitly in the context of grasping transitions how this can be done.
We linked the attention parameters λ1, λ2 to the experimentally accessible object-body scale ratio α. To this end, in Section 2.1.9 we wrote the attention parameters λ1 and λ2 as functions of α: λ1(α) and λ2(α). Taking a dynamical systems perspective, α corresponds to a control parameter. In Section 2 we showed that α captures the unimanual and bimanual grasping mode in a dynamical framework. Equations 1 and 12 predict, for any time point t, for any given pair ξ1(t) and ξ2(t), and for any given parameter α, which grasping mode will be selected. By solving Eqs. 1 and 12 numerically (see simulations in Fig. 9) and analytically (see the fixed point analysis in Sections 2.1.1,...,2.1.5) from Eqs. 1 and 12 a mapping
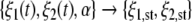 |
23 |
can be derived. This mapping involves a dynamical aspect because the time point t on the left hand side of Eq. 23 does not equal the time point that occurs on the right hand side of Eq. 23. Dynamic perception as opposed to static or passive perception accounts for the fact that human-environmental systems evolve in time and that the emergence of behavioral patterns is also a process that involves time. Thus, Eqs. 1 and 12 illustrate how the issues of dynamic perception and action can be addressed explicitly by recourse to dynamical systems theory.
In general, order parameters of a system can be obtained by first identifying the entire set of evolution equations that describe the system and second by diagonalizing the linearized parts of these evolution equations [30]. In the context of body-scaled transitions, we are dealing with humans acting and perceiving in environments. Consequently, in order to derive Eq. 1 from a structural (bottom-up) approach we would need to take microscopic evolution equations into account that involve both environmental components and components of human motor control systems. Therefore, in our context, the order parameter dynamics (1), and the potential function V(ξ1,ξ2), as well as the attractors and repellers that can be associated with the dynamics (1), reflect both environmental components and structural components of human actors. In particular, we argued that the parameters λ1, λ2, B, and C depend on a control parameter α that represents an object-size to body-scale ratio. However, other properties of human-environmental systems such as the age and gender of actors, the mass of objects, and the availability of tools that assist grasping of larger objects may affect these parameters as well. In fact, such human-environment properties have been addressed in experimental studies [1–7]. Future efforts may be devoted to explore the impacts of such properties on body-scaled transitions from a dynamical systems perspective centered on Eq. 1.
In our study we did not focus on the magnitudes of the order parameters ξ1 and ξ2. The reason for this is that according to our approach the deterministic grasping behavior is completely determined by two types of fixed points. Fixed point I has coordinates ξ1 > 0, ξ2 = 0 and reflects unimanual grasps. Fixed point II has coordinates ξ1 = 0, ξ2 > 0 and reflects bimanual grasping patterns. That is, all states with ξ1 > 0, ξ2 = 0 are mapped to one-handed grasping and all states with ξ1 = 0, ξ2 > 0 are considered as states that represent two-handed grasping. However, when stochastic aspects of grasping transitions are studied, the magnitudes of the order parameters have to be examined carefully. Close to critical points in the parameter space of Eq. 1 at which grasping mode attractors of the potential V(ξ1,ξ2) vanish, fluctuations may induce transitions between attractors of unimanual and bimanual grasping. Such transitions depend on the magnitudes of the order parameters ξ1 and ξ2 and particularly on the height of the potential barriers that separate minima of V(ξ1,ξ2). A stochastic account in this direction, however, is beyond the scope of the present study.
The order parameter dynamics (1) was proposed by Haken (see [37]) in the context of pattern recognition processes. Haken not only addressed two order parameters but an arbitrarily large set of order parameters. Accordingly, the recognition of a particular face or scene implies that the order parameter of that face or scene becomes finite. In Section 2 we interpreted the order parameters ξ1 and ξ2 of Eq. 1 as amplitudes of two behavioral modes in order to make contact with experimental studies on transitions between unimanual and bimanual grasping [1–7]. In line with Haken’s original model, we may interpret ξ1 and ξ2 as amplitudes of different perceptual judgments or cognitive modes. In grasping experiments there are two fundamental perceptual judgments. Participants judge an object as something that they would grasp with one hand (1H cognitive mode) or participants judge an object as something that they would grasp with two hands (2H cognitive mode). In fact, in [7] grasping was studied this way. Participants were asked to judge whether or not they would grasp an object with one or two hands. In these judgment trials grasping acts themselves were not performed. Transitions were observed between the 1H and 2H cognitive or perceptual judgment modes. Thus, when interpreting ξ1 and ξ2 in this latter way, Eq. 1 in principle also applies to the judgment findings reported in [7].
In [7] grasping was also studied from a social perspective. Participants were grouped into pairs. These pairs of participants were asked to transport planks of different sizes from one position to another. The participants could carry these planks alone or as couples. In [7] one-person grasping as well as two-person grasping was observed. Transitions between the one-person and two-person grasping mode were observed as well as hysteresis. The results found in the study on social interaction can also be addressed from the dynamical systems perspective developed in Section 2 by re-interpreting the order parameters ξ1 and ξ2 as order parameters of modes of social behavior.
The winner-takes-all model (1) allows for the study of individual behavior (see Section 2.1) and the behavior of a population (see Section 2.2). In fact, experimental studies have revealed qualitative differences in individual performance [1, 7]. Future research may be directed at studying individual performance differences from the perspective of Eq. 1. In particular, questions regarding the extent to which performance differs qualitatively between individuals (or pairs of individuals) can be related to the parameters λ1, λ2, B, and C of Eq. 1.
Finally, in Section 2.1.9 the attention parameters λ1 and λ2 were assumed to depend linearly on the control parameters. For the sake of simplicity we assumed that λ1 and λ2 increase and decrease by the same amount for a varying control parameter α. That is, in Eq. 12 we used the same β-factor for the functions λ1(α) and λ2(α). A natural generalization of Eq. 12 involves different β-factors such that we have
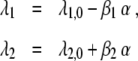 |
24 |
with β1, β2 > 0. In general, the functions λ1(α) and λ2(α) may depend in a nonlinear fashion on the control parameter α. Generalized relationships such as given by Eq. 24 may be useful in order to relate experimental data to the order parameter dynamics (1). As mentioned in Section 2.1.9, in the most general case, the nonlinearity parameters B and C will depend on α as well. This would imply that the factor g changes with α. In this case the transition between one-handed and two-handed grasps in the ascending condition would occur at the critical control parameters αc satisfying
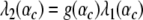 |
25 |
whereas the transition between two-handed and one-handed grasps in the descending condition would occur at
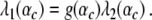 |
26 |
A challenge for future experimental and theoretical work, therefore, is to design experiments and to develop data analysis methods such that the functions λ1(α), λ2(α), and g(α) can be extracted from experimental data. Identifying the information that specifies the different behavioral modes as well as manipulating different biomechanical, cognitive, and social variables will be essential to such experimentation.
References
- 1.Cesari, P., Newell, K.M.: Body-scaled transitions in human grip configurations. J. Exp. Psychol. Hum. Percept. Perform. 26, 1657–1668 (1999) [DOI] [PubMed]
- 2.Cesari, P., Newell, K.M.: Scaling of human grip configurations. J. Exp. Psychol. Hum. Percept. Perform. 25, 927–935 (1999) [DOI] [PubMed]
- 3.Newell, K.M., Scully, D.M., McDonald, P.V., Baillargeon, R.: Task constraints and infant grip configuration. Dev. Psychobiol. 22, 817–832 (1989) [DOI] [PubMed]
- 4.Newell, K.M., Scully, D.M., Tenenbaum, F., Hardiman, S.: Body scale and the development of prehension. Dev. Psychobiol. 22, 1–13 (1989) [DOI] [PubMed]
- 5.van der Kamp, J., Savelsbergh, G.J.P., Davis, W.E.: Body-scaled ratio as a control parameter for prehension in 5- to 9-year old children. Dev. Psychobiol. 33, 351–361 (1998) [DOI] [PubMed]
- 6.van Hof, P., van der Kamp, J., Savelsbergh, G.J.P.: The relation of unimanual and bimanual reaching to crossing the midline. Child Dev. 73, 1353–1362 (2002) [DOI] [PubMed]
- 7.Richardson, M.J., Marsh, K.L., Baron, R.M.: Judging and actualizing intrapersonal and interpresonal affordances. J. Exp. Psychol. Hum. Percept. Perform. 33, 845–859 (2007) [DOI] [PubMed]
- 8.Marsh, K.L., Richardson, M.J., Baron, R.M.: Contrasting approaches to perceiving and acting with others. Ecol. Psychol. 18, 1–38 (2006) [DOI]
- 9.Gibson, J.J.: The Ecological Approach to Visual Perception. Houghton-Mifflin, Boston (1979)
- 10.Michaels, C.F., Carello, C.: Direct Perception. Prentice-Hall, Inc., Englewood Cliffs, N.J. (1981)
- 11.Turvey, M.T.: Affordances and prospective control: an outline of the ontology. Ecol. Psychol. 4(3), 173–187 (1992) [DOI]
- 12.Warren, W.H.: Perceiving affordances: visual guidance of stair climbing. J. Exp. Psychol. Hum. Percept. Perform. 10, 683–703 (1984) [DOI] [PubMed]
- 13.Beek, P.J., Peper, C.E., Stegeman, D.F.: Dynamical models of movement coordination. Hum. Mov. Sci. 14, 573–608 (1995) [DOI]
- 14.Frank, T.D.: On a nonlinear master equation and the Haken-Kelso-Bunz model. J. Biol. Phys. 30, 139–159 (2004) [DOI] [PMC free article] [PubMed]
- 15.Frank, T.D.: Nonlinear Fokker-Planck Equations: Fundamentals and Applications. Springer, Berlin (2005)
- 16.Frank, T.D., Daffertshofer, A., Beek, P.J., Haken, H.: Impacts of noise on a field theoretical model of the human brain. Physica D 127, 233–249 (1999) [DOI]
- 17.Frank, T.D., Daffertshofer, A., Peper, C.E., Beek, P.J., Haken, H.: Towards a comprehensive theory of brain activity: coupled oscillator systems under external forces. Physica D 144, 62–86 (2000) [DOI]
- 18.Frank, T.D., Michelbrink, M., Beckmann, H., Schöllhorn, W.I.: On a quantitative dynamical systems approach to differential learning: self-organization principle and order parameter equations. Biol. Cybern. 98, 19–31 (2008) [DOI] [PubMed]
- 19.Haken, H.: Principles of Brain Functioning. Springer, Berlin (1996)
- 20.Jirsa, V.K., Kelso, J.A.S.: Coordination Dynamics: Issues and Trends. Springer, Berlin (2004)
- 21.Kelso, J.A.S.: Phase transitions and critical behavior in human bimanual coordination. Am. J. Physiol. 15, R1000–R1004 (1984) [DOI] [PubMed]
- 22.Kelso, J.A.S.: Dynamic Patterns - The Self-organization of Brain and Behavior. MIT Press, Cambridge (1995)
- 23.Mechsner, F., Kerzel, D., Knoblich, G., Prinz, W.: Perceptual basis of bimanual coordination. Nature 414, 69–73 (2001) [DOI] [PubMed]
- 24.Peper, C.E., Beek, P.J., van Wieringen, P.C.W.: Frequency-induced transitions in bimanual tapping. Biol. Cybern. 73, 301–309 (1995) [DOI] [PubMed]
- 25.Schöner, G.S., Kelso, J.A.S.: A synergetic theory of environmentally-specified and learned patterns of movement coordination. I. Relative phase dynamics. Biol. Cybern. 58, 71–80 (1988) [DOI] [PubMed]
- 26.Schöner, G.S., Kelso, J.A.S.: A synergetic theory of environmentally-specified and learned patterns of movement coordination. II. Component oscillator dynamics. Biol. Cybern. 58, 81–89 (1988) [DOI] [PubMed]
- 27.Sternad, D., Turvey, M.T., Saltzman, E.L.: Dynamics of 1:2 coordination: generalizing relative phase to n:m rhythms, sources of symmetry breaking, temporal scaling, latent 1:1, and bistability. J. Mot. Behav. 31, 207–247 (1999) [DOI] [PubMed]
- 28.Tass, P., Wunderlin, A., Schanz, M.: A theoretical model of sinusoidal forearm tracking with delayed visual feedback. J. Biol. Phys. 21, 83–112 (1995) [DOI]
- 29.Turvey, M.T.: Coordination. Am. Psychol. 45, 938–953 (1990) [DOI] [PubMed]
- 30.Haken, H.: Synergetics: Introduction and Advanced Topics. Springer, Berlin (2004)
- 31.Lee, D.N., Reddish, D.E.: Plummeting gannets: a paradigm of ecological optics. Nature 293, 293–294 (1981) [DOI]
- 32.Turvey, M.T.: Dynamic touch. Am. Psychol. 51, 1134–1152 (1996) [DOI] [PubMed]
- 33.Withagen, R.: The pick-up of non-specifying variables does not entail indirect perception. Ecol. Psychol. 16, 237–253 (2004) [DOI]
- 34.Lee, D.N., Young, D.S., Reddish, D.E., Lough, S., Clyton, T.M.H.: Visual timing in hitting an accelerating ball. Q. J. Exp. Psychol. 35, 333–346 (1983) [DOI] [PubMed]
- 35.Savelsbergh, G.J.P., Whiting, H.T.A., Bootsma, R.J.: Grasping tau. J. Exp. Psychol. Hum. Percept. Perform. 17, 315–322 (1991) [DOI] [PubMed]
- 36.Fitzpatrick, P., Carello, C., Schmidt, R.C., Corey, D.: Haptic and visual perception of an affordance for upright posture. Ecol. Psychol. 6(4), 265–287 (1994) [DOI]
- 37.Haken, H.: Synergetic Computers and Cognition. Springer, Berlin (1991)
- 38.Fuchs, A., Haken, H.: Pattern recognition and associative memory as dynamical processes in a synergetic system. I. Translational invariance, selective attention and decomposition of scene. Biol. Cybern. 60, 17–22 (1988) [DOI] [PubMed]
- 39.Fuchs, A., Haken, H.: Pattern recognition and associative memory as dynamical processes in a synergetic system. II. Decomposition of complex scenes, simultaneous invariance with respect to translation, rotation, and scaling. Biol. Cybern. 60, 107–109 (1988) [DOI] [PubMed]
- 40.Haken, H.: Nonequilibrium phase transitions in pattern recognition and associative memory. Z. Phys. B 70, 121–123 (1988) [DOI]
- 41.Ditzinger, T., Haken, H.: Oscillations in the perception of ambigious patterns: a model based on synergetics. Biol. Cybern. 61, 279–287 (1989) [DOI]
- 42.Bressloff, P.C.: Neural networks, lattice instantons, and the anti-integrable limit. Phys. Rev. Lett. 75, 962–965 (1995) [DOI] [PubMed]
- 43.Bressloff, P.C.: A self-organizing neural network in the weak-coupling limit. Physica D 110, 195–208 (1997) [DOI]
- 44.Bressloff, P.C., Roper, P.: Stochastic dynamics of the diffusive Haken model with subthreshold periodic forcing. Phys. Rev. E 58, 2282–2287 (1998) [DOI]
- 45.Turvey, M.T.: Impredicativity, dynamics, and the perception-action divide. In: Jirsa, V.K., Kelso, J.A.S. (eds.) Coordination Dynamics: Issues and Trends, vol. I, pp. 1–20. Springer, New York (2004)
- 46.Warren, W.H.: The dynamics of perception and action. Psychol. Rev. 113, 358–389 (2006) [DOI] [PubMed]