

Abstract
Barx1 modulates cellular adhesion molecule expression and participates in specification of tooth-types, but little is understood of its role in patterning the pharyngeal arches. We examined barx1 expression during zebrafish craniofacial development and performed a functional analysis using morpholino oligonucleotides. Barx1 is expressed in the rhombencephalic neural crest, the pharyngeal arches, the pectoral fin buds and the gut in contrast to its paralogue barx2, which is most prominently expressed in the arch epithelium. Additionally, barx1 transient expression was observed in the posterior lateral line ganglia and developing trunk/tail. We show that Barx1 is necessary for proliferation of the arch osteochondrogenic progenitors, and that morphants exhibit diminished and dysmorphic arch cartilage elements due to reductions in chondrocyte differentiation and condensation. Attenuation of Barx1 results in lost arch expression of osteochondrogenic markers col2a1, runx2a and chondromodulin, as well as odontogenic marker dlx2b. Further, loss of barx1 positively influenced gdf5 and chordin, markers of jaw joint patterning. FGF signaling is required for maintaining barx1 expression, and that ectopic BMP4 induces expression of barx1 in the intermediate region of the second pharyngeal arch. Together, these results indicate an essential role for barx1 at early stages of chondrogenesis within the developing zebrafish viscerocranium.
Keywords: Barx1, pharyngogenesis, viscerocranium, zebrafish, pharyngeal arches, neural crest, chondrogenesis, cartilage, BMP, FGF
Introduction
The viscerocranium consists of highly adapted skeletal elements derived from the embryonic pharyngeal arch ectomesenchyme that function in concert but allow for diverse pharyngolaryngeal activities. The viscerocranium is composed of membranous and endochondral bones. Endochondral bone formation within the pharyngeal arches is a multi-step process that requires the migration of the cranial neural crest into the facial prominences and their subsequent epitheliomesenchymal interactions; these interactions are necessary for fate determination, aggregation of the cells into discrete condensations, and their terminal differentiation (Hall and Miyake, 2000; Helms and Schneider, 2003; Goldring et al., 2006). Signaling factors that include Bone Morphogenic Proteins (BMPs) and Fibroblast Growth Factors (FGFs), through reciprocal epitheliomesenchymal interactions, influence the expression of downstream factors that pattern the chondrogenic elements (de Crombrugghe et al., 2000; Hall and Miyake, 2000; Tuan, 2004; Goldring et al., 2006). The precise mechanism of patterning osteochondrogenic progenitors is not well understood.
The Barx family of homeodomain-containing transcription factors participates in the formation of mesenchymal condensations through the modulation of cellular adhesion molecules (CAMs), and regulation of fibril-forming type II collagen (Col2a1) (Jones et al., 1997; Edelman et al., 2000; Meech et al., 2005). The regulation of CAMs and Col2a1, a major component of the cartilage extracellular matrix, by Barx1 indicates a direct link between expression of a tissue-specific transcription factor and changes in cartilage morphology. In the mouse, Barx1 expression is located at sites of mesenchymal condensation that include the pharyngeal arches, the limb buds, developing joints, molar tooth papillae and the stomach wall (Tissier-Seta et al., 1995; Jones et al., 1997; Barlow et al., 1999; Kim et al., 2005). Ectopic Barx1 gene expression in mouse mandible cultures results in the alteration of incisor teeth to an unusual molariform shape (Tucker et al., 1998; Miletich et al., 2005), while attenuation of Barx1 by RNA interference results in arrest of molar mesenchyme at the bud stage (Song et al., 2006). Manipulation of the closely related Barx2 gene expression in limb bud cultures affects cellular aggregation and chondrocyte differentiation, indicating a familial role in patterning chondrocytes (Meech et al., 2005). The function of Barx1 in craniofacial development has yet to be fully examined. In humans, rare duplications and deletions of the BARX1 locus result in craniofacial and joint anomalies, but no disease-causing mutations have been associated directly with this gene (Stalker et al., 1993; Gould and Walter, 2000; Gould and Walter, 2004; Redon et al., 2006). To explore the relationship of this gene with craniofacial development we characterized zebrafish barx1 and the phenotype on the barx1morphant.
Here we show that the zebrafish barx1 gene is expressed in the migrating cranial neural crest and subsequently in the developing cartilage elements of the pharyngeal arches. Transient expression is observed in the posterior lateral line ganglia and in the trunk and tail. In contrast to barx1, the barx2 paralogue is expressed predominantly in the proximal aspects of the fin buds and in oral and aboral epithelium of the first and second pharyngeal arches. Attenuation of barx1 expression using antisense morpholinos results in a loss of arch cartilage tissue and micrognathia without an increase in apoptosis, but with a reduction in Phosphohistone-H3 stained nuclei, indicating a role for barx1 in cellular proliferation and chondrocyte differentiation. As FGF and BMP signaling is known to pattern arch development, we examined the influence of these factors on barx1 expression. Examination of mutant embryos deficient in fgf3 (lia) and fgf8 (ace), as well as embryos treated with an FGF receptor antagonist (SU5402), show that FGF signaling is necessary for maintaining and patterning barx1 expression in the arches. In contrast, ectopic BMP4 signaling induces misspecification of barx1 patterning within the intermediate aspect of the second pharyngeal arch. This work provides insight into barx1 function and patterning during prechondrogenic condensation events in the developing zebrafish viscerocranium.
Materials and Methods
Animal maintenance and transgenic zebrafish
Zebrafish embryo lines, EK wild-type, fli1:GFP transgenics (Motoike et al., 2000), and the fgf8/ace mutant, were maintained according to Westerfield (1995). Fixed wild-type and sibling fgf3/lia embryos were a gift from the Hammerschmidt laboratory (Germany). Embryos and larvae were fixed in 4% paraformaldehyde/phosphate buffered saline and dehydrated in methanol for storage at −20°C. FGF inhibition was performed by soaking 24 hours post-fertilization (hpf) embryos in 10 μm SU5402 (Pfizer) in 5% DMSO.
In vitro transcription of synthetic RNA
The zebrafish barx1 (NM001024949) and the barx2 predicted open reading frame (XM001342008) were amplified from cDNA produced by reverse transcription from 96 hpf RNA and cloned into pCRII-TOPO (Invitrogen). For capped sense mRNA, the barx1 open reading frame was cloned into pCS2+ plasmid, digested with Not I, and transcribed with SP6 Polymerase. Capping was performed with 7′ methylguanosine GTP analogue cap (NEB).
Attenuation of barx1 expression
Morpholinos (MO) complementary to the translational start site of barx1 (Bx MO) and the splice acceptor (Sa MO) of the second exon (Fig. 1A) were microinjected into one to two cell-stage embryos. Lissamine conjugated MOs (Gene-Tools) used are as follows: Bx MO, [5′CCCCAATCTCCAAAGGATGTTGCAT3′], Sa MO [5′GCCTTCAGAACTGGAATG AAATAAG3′], and a standard control [5′CCTCTTACCTCAGTTACAATTTATA3′]. MOs were diluted in Danieau buffer (58 mM NaCl, 0.7 mM KCl, 0.4 mM MgSO4, 0.6 mM Ca(NO3)2, 5 mM Hepes, pH 7.6) and 0.01% Phenol Red. Embryos were injected with approximately 1 nl of MO at a concentration of 4 ng/nl. For rescue, the MOs were coinjected with 25 ng/μl of barx1 mRNA containing five base pair mismatches within the ATG MO target site. To test inhibition of translation, a barx1:eGFP fusion construct was injected together with the MO, and proteins extracted from 24 hpf embryos were blotted with anti-gfp (1/1000; Santa Cruz) and anti-α-tubulin (1/1000; CalBiochem) antibodies, and detected by a Dura Chemiluminescence Kit (Pierce). To test the effectiveness of the Sa MO, total RNA was extracted from 24 hpf morphant and control embryos, DNase I treated and column purified (Qiagen). RT-PCR, using random hexamer primers was performed on 1 μg of RNA for each sample (Superscript III, Invitrogen). Forward (5′-AATGCAACATCCTTTGGAGATT-3′) and reverse (5′-ATCCCGTTTATTCCTCTTGGTT-3′) primers were used to test for properly spliced barx1 mRNA. β-actin amplification was used as a control.
Figure 1.

Molecular analysis of zebrafish barx1. (A) The coding region of barx1 consists of four exons (boxes, numbered with Roman numerals) containing the homeobox (HB, blue) and the Bar Basic Region (BBR, green); UTR’s are indicated by outlined boxes. Target sites of the barx1 ATG morpholino (Bx MO) and the Splice acceptor site morpholino (Sa MO) are indicated. (B) Phylogenetic tree comparing zebrafish (z) Barx1 with the human (h), mouse (m), and chicken (c) Barx amino acid sequences, as examined using Clustal W. GenBank Accession Nos.: (h1) NM021570, (m1) NM007526, (c1) NM204193, (z1) NM001024949, (h2) NM003658, (m2) NM013800, (c2) NM204896, (z2) XM001342008 respectively. (C) Syntenic relationship between zebrafish chromosome 11 fragment contig: CR548622.8 (Ensembl release 49, March 2008) and the human chromosome 9q22.32 (loci distances indicated in Mb). The barx1 and phf2, PHD finger protein 2, are linked in zebrafish and humans.
Whole-mount in situ hybridization and histological characterization
The following antisense riboprobes were used: chondromodulin (AF322374) (Sachdev et al., 2001); col2a1 (U23822) (Yan et al., 1995); crestin (AF195881) (Luo et al., 2001); dlx2a (NM131311) (Akimenko et al., 1994); dlx2b (NM131297) (Jackman et al., 2004); crestin (Rubinstein et al., 2000; Luo et al., 2001); goosecoid (gsc) (NM131017) (Stachel et al., 1993); hand2 (dhand; NM131626) (Angelo et al., 2000); inca (Luo et al., 2007); neurod, (NM130978) (Korzh et al., 1998); runx2b (NM212862) (Flores et al., 2004); sox9a (NM161343) (Chiang et al., 2001). Whole-mount in situ hybridizations were performed as described by Akimenko et al. (1994), using anti-digoxigenin, and anti-fluorescein antibodies (Roche).
Phosphohistone-H3 antibody (Upstate) was used at 1/1000 dilution, followed by anti-mouse biotinylated secondary antibody (Vectastain). Cartilage staining was performed as in Ellies et al., (1997). Histological analysis was performed on embryos embedded in JB-4 plastic (Polysciences) and serially sectioned. Sections were stained with 13% Methylene blue and 0.13% Basic Fuchsin. Apoptosis was determined by TUNEL labeling, using Terminal Transferase (Roche) and dUTP-DIG (Roche) as described in Ellies et al. (1997).
Bead implantation
Recombinant BMP4 (1 μg/ml) (R&D Systems) was used with Affi-Gel Blue beads (Bio-Rad). One percent BSA coated Heparin beads (Sigma) were used as control. Beads were soaked at 4°C overnight. A sharp needle (Fine Instruments) was used to embed beads anterior to the otic vesicle on the left side of the embryo.
Photography
Samples were digitally photographed with a Leica MZ APO dissecting microscope connected to a computer running Northern Eclipse 6.0 image capture software (Empix Imaging Inc.). Live animals were immobilized with tricaine and mounted in egg water or methylcellulose. For higher magnification, fixed embryos were cleared in glycerol and digitally photographed using a Zeiss Axioplan 2.
Results
Zebrafish barx1 is the orthologue of the mouse and human Barx1 genes
The sequence encoding zebrafish barx1 has recently been deposited in GenBank (Accession No. NM001024949). Vertebrates have at least two barx genes, and BLAST-searches of the most recent version of the zebrafish genome (Zv7), yielded two barx paralogues, barx1 and a predicted barx2 (XM001342008) located on chromosome 17. The barx1 open reading frame is encoded in four exons and includes an engrailed homology (EH) domain, the homeobox (HB) and a conserved Bar Basic Region (BBR, Fig. 1A, S1). The Bar Basic Region is thought to assist in both positive and negative regulation of downstream genes in a tissue-specific manner (Edelman et al., 2000). Similarly, the barx2 sequence indicates a conservation of the EH, HB and BBR domains. ClustalW alignment and phylogenetic analysis of the zebrafish, human, mouse and chick Barx proteins indicate that the zebrafish barx1 gene is closely related to the Barx1 clade (Fig. 1B; Supplementary Fig. S1). The zebrafish barx1 gene, located on chromosome 11 (DNA contig: CR548622.8; Zv7 genome assembly; Ensembl release 49, March 2008), exhibits conserved synteny with human BARX1 located on chromosome 9q22.32 (Ensembl release 49), but not with human BARX2 located on chromosome 11q24.3 (Hjalt and Murray, 1999), confirming orthology between the two genes (Fig. 1C).
barx1 expression in the developing zebrafish embryo
In zebrafish, barx1 expression has been previously described and used as a marker of pattering within the developing pharyngeal arches (Walker et al., 2006; Walker et al., 2007); however, its expression pattern has not been fully characterized. To further understand its role in development we performed a series of in situ hybridizations in embryos from the one-cell stage to 120 hours post fertilization (hpf) larvae. Transcripts of barx1 were not detected by RT-PCR or whole-mount in situ hybridization prior to somitogenesis (data not shown). We observed expression at 9 somites (13 hpf) during the initiation of rhombencephalic neural crest migration in bilateral domains on either side of the neural tube (arrows; Fig. 2A, B). Cells expressing barx1 appear to be a subset of the rhombencephalic neural crest, as these cells are located in the most ventral aspect of the streams as compared to the pan-neural crest marker crestin (ctn, red) (Fig. D). At 24 hpf, expression was observed in the three streams of the cranial neural crest, in the gut primordia, fin bud primordia and in the trunk and tail (Fig. 2E, F; Supplementary Fig. S2A and B). Trunk and tail expression was progressively lost by 48 hpf (Supplementary Fig. S2A, B and C). Transcripts in the differentiating ectomesenchyme of the pharyngeal arches at 48 hpf were concentrated in the developing prechondrogenic condensations (Fig. 2G). Barx1 expression within the dorsal aspect of the second arch appeared to be rostrally enriched, potentially correlating with sites of increased proliferation within the hyomandibular condensation that subsequently fuses with the symplectic element (Kimmel et al., 1998). Expression was also maintained in the pectoral fin bud (Fig. 2G, inset). In mammals, Barx1 participates in tooth development (Tucker et al., 1998; Song et al., 2006; Mitsiadis and Drouin, 2008).. We examined whether barx1 is expressed in the developing tooth buds at 56 hpf, a time when tooth primordia are readily observable (Jackman et al., 2004) (asterisk, Supplementary Fig. S2D). Expression of barx1 appears to be adjacent to the condensing tooth bud primordia. In 72 hpf embryos, barx1 continued to be expressed for the most part in the stacked arrays of chondrogenic cells, but diminishes as the elements mature (Fig. 2H and data not shown). At this time, we also identified barx1 expression in the posterior lateral line ganglia (pllg, Supplementary Fig. S2E, F, H, and H inset). Transcripts of barx1 continued to be detected in the intestinal wall of the gut (Supplementary Fig. S2H and I). By 96 hpf, serial sections of larvae showed a progressive loss of barx1 expression in the stacked cartilage cells, with expression remaining in the surrounding mesenchyme (Fig. 2I). In summary, expression in the head skeleton was restricted to the ectomesenchyme of the developing pharyngeal arches up to 120 hpf, and was not observed in the neurocranium.
Figure 2.

Barx1 expression. Embryos at (A, B) 13 hpf, (C)18 hpf, (D) 19 hpf, (E, F) 24 hpf, (G) 48 hpf, (H) 72 hpf, and (I) 96 hpf, were examined by in situ hybridization. (A) barx1 expression in the migrating neural crest in bilateral domains adjacent to the hindbrain (arrows). Anterior is at the top. (B) coronal section through the hindbrain. (C) lateral view of barx1 expression (arrow). (D) double in situ of barx1 (dark blue, arrowhead) and crestin (red) co-expression in the neural crest streams indicated by Roman numerals. (E, F) expression in the cranial neural crest streams. (G, G inset) barx1 expression in the pharyngeal arches and fin bud; a1, first arch; a2, second arch; d, dorsal expression domain; ga, gill arches; v, ventral expression domain; asterisk indicates the a2 intermediate region. (H) lateral oblique view of barx1 expression in the chondrocytes. (I) parasagittal section, open arrowhead indicates jaw joint. ch, ceratohyal; ceratobranchials are numbered; e, eye; fb, fin bud; g, gut primordium; hmc, hyomandibular condensation; M, Meckel’s cartilage; ov, otic vesicle; pq, palatoquadrate; tr, trabecula. Scale bar: (B and I) 50 μm; (C, E, F, and G) 100 μm.
To determine whether there might be genetic redundancy between barx1 and barx2, we examined the expression pattern of the barx2 paralogue (Supplementary Fig. S3). In contrast to barx1, barx2 is expressed in the proximal aspect of the pectoral fin buds from 24 to 32 hpf (Fig. S3A–C). From 36 hpf to 72 hpf, expression is observed in the in the oral and aboral epithelium of the mandibular and hyoid arches (Fig. S3D–H), and not in the condensing mesenchyme of the chondrogenic elements (arrowheads, Fig. S3G). Expression is also observed in the olfactory epithelium (Fig. S3F inset). The expression patterns of barx1 and barx2 do not overlap in the condensing mesenchyme of the prechondrogenic elements, and suggests that limited or no genetic redundancy exists between the paralogues.
barx1 is necessary for pharyngeal arch chondrogenesis
To determine the role of barx1 in the developing pharyngeal arches and viscerocranium, we used antisense morpholinos (MO) targeted to the ATG site (Bx MO) and a splice-acceptor site (Sa MO) (Fig. 1A). MOs were injected into 1–2 cell embryos, and development was followed to 120 hpf. Compared to control MO-injected larvae at 120 hpf, morphants exhibited reduction in head size ranging from mild to severe hypoplastic pharyngeal arch morphology exemplified by micro- or agnathia (Fig. 3A–C, arrow), due to poor pharyngeal arch outgrowth (open arrowhead, Fig. 3A–C). Alcian Blue staining showed reductions or a lack of pharyngeal arch-derived chondrogenic elements (Fig. 3D–F). Generally, all the elements were dysmorphic, while the ceratobranchial elements were the most likely to be lost, which included the fifth arch elements containing the pharyngeal teeth. The most severely affected larvae exhibited either small cartilage nodules in place of elements or lacked any cartilage in the viscerocranium (open arrowhead, Fig. 3F, and data not shown). In more mildly affected larvae, reductions in chondrocyte condensation and incomplete fusions were observed between the symplectic and the hyomandibular elements (asterisk, Fig. 3G, H), as well as abnormal joint fusions between Meckel’s cartilage and the palatoquadrate element (black arrowhead, Fig. 3G, H). Histological examination of larvae at 72 hpf during cartilage condensation in the pharyngeal arches showed a lack of cellular organization within the arches, as shown by a deficiency of prechondrocyte aggregation and absence of characteristic extracellular matrix surrounding the cells (arrow, Fig. 3I, J; see also boxed region in Supplementary Fig. S4A, B).
Figure 3.

Attenuation of barx1 expression perturbs viscerocranial morphology and cartilage patterning. Larvae injected with (A) control MO (4 ng), and (B and C) Bx MO (4 ng). Alcian Blue staining of control MO (D) and Bx MO injected (E and F) 120 hpf larvae. Dissection of the first and second arches of control (G) and mildly affected larvae (H). (I and J) parasagittal sections of 72 hpf embryos. (J) Bx MO (4 ng) injected compared with (I) control MO injected (4 ng). Anterior is to the left; (A–C, G–J) lateral views, (D–F) ventral view. (A–C) Arrows indicate mouth, and arrowheads the pharyngeal arches; (D–F) open arrowheads indicate the ceratohyal cartilage; (G, H) asterisk, perturbed fusion; black arrowhead, joint fusion between Meckel’s cartilage (M) and the palatoquadrate (pq); cb, ceratobranchials; ch, ceratohyal; ep, ethmoid plate; hm, hyomandibular; ov, otic vesicle; sy, symplectic element. Scale bars: (A–C) 250 μm; (D–H) 100 μm; (J) 50 μm. (K, L) Effectiveness and specificity of the barx1 (Bx) and splice acceptor (Sa) morpholinos. (K) Western analysis of in vivo attenuation of barx1:GFP translation in 24 hpf zebrafish embryos. (L) total RNA extracts (18S and 28S bands shown) used for RT-PCR of barx1. β-actin used as a control. (M) Percentage of affected embryos comparing control MO, Bx MO and Sa MO, and rescue by co-injection of barx1 mRNA with a 5-bp mismatch to the ATG target site. Affected embryos are defined as those exhibiting micrognathia as a result of a reduction or loss of arch cartilage elements, as seen by Alcian Blue staining at 120 hpf. (1) Control MO 4 ng; (2) Bx MO 4 ng; (3) Sa MO 4 ng; (4) Bx MO 2 ng + Sa MO 2 ng. Rescue: (5) gfp mRNA 25 pg; (6) barx1 mRNA 25 pg; (7) Bx MO + Sa MO 2 ng each + barx1 mRNA 25 pg. Number of embryos indicated on top of each bar.
To test the effectiveness of the Bx MO, a barx1:GFP fusion mRNA containing the ATG MO target site was co-injected with the morpholino into 1–2 cell embryos. Western blot analysis at 24 hpf showed that the morpholino effectively inhibited barx1:GFP translation indicating that injected embryos are strong hypomorphs (Fig. 3K). Additionally, to verify that the phenotype was due to the attenuation of barx1 translation, a second MO targeting the second exon splice-acceptor site was tested (Sa MO; Fig. 1A). RT-PCR of Sa MO-injected embryo RNA extracts showed a reduced amount of product of spliced mRNAs when compared to control embryos (Fig. 2L). Embryos injected with 4 ng of Bx MO or the Sa MO exhibited identical phenotypes of dysmorphic viscerocranial patterning at similar frequencies (Fig. 3M, bars 2 and 3). Co-injection of both Bx MO and Sa MO (2 ng each) produced the same phenotype with a higher effectiveness (Fig. 3M, bar 4). Finally, rescue of the phenotype was achieved by co-injecting both morpholinos with barx1 mRNA containing a 5-base pair mismatch in the ATG target sequence (Fig. 3M, bar 7). Trials showed that low doses of barx1 mRNA (25 pg) co-injected with Bx+Sa MO (2 ng each) were effective at rescuing the phenotype in most larvae (89/111, 80%; 3M, bar7) as determined by apparently normal development at 120 hpf, and Alcian Blue staining of arch cartilage elements. However, higher doses (50 pg) of barx1 mRNA alone or with Bx+Sa MO caused most embryos to exhibit aberrant gastrulation resulting in highly dysmorphic development.
barx1 is not required for early neural crest specification
To test whether barx1 participates in specification of the prechondrocyte lineage, we examined cranial neural crest markers expressed in the migrating cells in barx1 morphants. Molecular markers of the cranial neural crest (see Materials and Methods) that include inca (Fig. 4A, B), dlx2a (Fig. 4C, D), as well as neurod, a marker of nascent ganglion cells (Fig. 4E, F), and hand2, an endodermal marker, were tested (Fig. 4G, H). In all cases, expression patterns of these markers in barx1 morpholino-injected embryos appeared normal at 24 hpf, although development of the injected embryos appeared mildly delayed. This suggests that a deficiency in barx1 expression does not strongly affect specification and migration of the hindbrain-derived neural crest. Likewise, examination of fli1:GFP transgenic embryos that express GFP in the pharyngeal arches (Lawson and Weinstein, 2002; Crump et al., 2004), suggested that neural crest migration into the arches is normal (Supplementary Fig. S4C–F).
Figure 4.

Initial patterning of the arches is maintained in barx1 morphants. (A, C, E and G) control MO injected embryos, (B, D, F and H) embryos co-injected with Bx MO + Sa MO, 2 ng each. Neural crest expression of inca (A, B) and dlx2a (C and D) at 24 hpf. (E, F) neurod expression at 24 hpf in neural ganglia precursors; (G and H) hand2 expression in ventral endoderm at 32 hpf. Neural crest streams marked in Roman numerals, gill arches indicated by numbers; e, eye; m, mandibular arch; h, hyoid arch; ov, otic vesicle; ad/av/f, anterodorsal/anteroventral lateral line/facial placode/ganglia; o, octavel/statoacoustic ganglia precursors; p, posterior lateral line placode/ganglion. Scale bar: 100 μm.
Pharyngeal arch chondrocyte differentiation requires barx1
To determine the role of barx1 in cartilage patterning, we examined markers of differentiation of osteochondroprogenitor cells in MO-injected embryos. The expression patterns examined included goosecoid (gsc), a marker of cellular condensation in the hyoid arch (Schulte-Merker et al., 1994); sox9a, a transcription factor necessary for chondrogenesis (Yan et al., 2002; Yan et al., 2005); runx2b, a marker of mesenchymal condensations and chondrocyte differentiation (Flores et al., 2006); fibril-forming type II collagen (col2a1), the major component of the cartilage extracellular matrix (Yan et al., 1995), chondromodulin (chm), a secreted glycoprotein that stimulates maturation of chondrocytes and inhibits vascularization (Sachdev et al., 2001); and dlx2b, a marker of tooth differentiation on the fifth ceratobranchial arch (Jackman et al., 2004) (Fig. 5). While the heads of barx1 morphants were reduced in size compared to controls, gsc and sox9a expression was maintained in the hypoplastic arches, suggesting that gsc and sox9a expression may be regulated independently of barx1 (Fig. 5A–D). However, molecular markers of the chondrogenic and subsequent osteogenic program, col2a1, chm, and runx2b, were reduced or lost (Fig. 5E–J). In addition to the perturbation of the osteochondrogenic program, morphants showed a loss of dlx2b expression suggesting that tooth development was also affected (Fig. 5K, L). These data point to the importance of Barx1 function during the earliest stages of chondrogenesis.
Figure 5.

Barx1 is necessary for expression of chondrogenic markers within the pharyngeal arches. (A, C, E, G, I and K) control embryos; (B, D, F, H, J and L) barx1 morphant embryos (see Fig. 4). Whole-mount in situ hybridization for (A, B) goosecoid (gsc), (C, D) sox9a, (E, F) runx2b, (G, H) type II collagen (col2a1), (I, J) chondromodulin (chm), and (K, L) dlx2b. Anterior is to the left; open arrowheads indicate sites of altered marker expression in the morphants; bcc, basicranial commissures; cb, ceratobranchials; ch, ceratohyal; cl, cleithrum; d, diencephalon; ep, ethmoid plate; fb, fin bud; hsc, hyosymplectic condensation; ov, otic vesicle; pq, palatoquadrate; t, telencephalon. Scale bar: 100 μm.
Attenuation of barx1 enhances markers of jaw joint patterning
Morphants stained with Alcian Blue exhibited a range of altered cartilage morphology that included mispatterning of the joints (Fig. 3D–H). Barx1 is expressed in the mouse and chick joints of the limbs and digits and has been suggested to regulate joint patterning by inhibiting chondrogenesis in limb micromass cultures (Tissier-Seta et al., 1995; Church et al., 2005). To determine the influence of barx1 on joint patterning within the viscerocranium, we examined between 48 and 80 hpf molecular markers, bapx1, chordin, and gdf5, which are expressed in the developing jaw joint (Miller et al., 2003). Expression of bapx1 at 48 hpf (Fig. 6A, B) and 56 hpf (data not shown), showed no to perhaps a mild enhancement in expression in morphant embryos. However, expression patterns of chordin as well as gdf5 were noticeably enhanced within the jaw joint by the loss of barx1 (open arrowheads, Fig. 6C–F). These results suggest that barx1 influences the regulation of these two genes necessary for the proper patterning of the jaw joint in zebrafish.
Figure 6.

Loss of barx1 enhances chordin and gdf5 expression in the jaw joint Whole-mount in situ hybridization for (A, B) bapx1, (C, D) chordin (chd), and (E, F) gdf5. (A, C, E) control uninjected embryos; (B, D, F) barx1 morphants (see Fig. 4). Embryonic ages as indicated. (A, B) lateral view, anterior to the left. (C–F) ventral view, anterior at the top; asterisk indicates Meckel’s cartilage, open arrowheads indicate jaw joint expression domains; e, eye; ch, ceratohyal; o orifice; pq, palatoquadrate. Scale bar: (A–F) 50 μm.
barx1 is necessary for cellular proliferation of chondrogenic progenitors
We examined the effect of loss of barx1 function on cell death and proliferation within the migrating neural crest streams and the pharyngeal arches. Control and barx1 morphant embryos during migration and pharyngogenesis (13, 24 and 48 hpf) showed no apparent increase in apoptosis as determined by TUNEL labeling (data not shown). The hypoplastic arches suggested that barx1 might participate in cellular proliferation. The pharyngeal arch mesenchyme was examined in serially sectioned control and barx1 morphants stained with Phosphohistone-H3 (PH3) antibody at 34, 48, 56, and 72 hpf, a period of prominent arch outgrowth. Stained nuclei were counted within the arch ectomesenchyme, as indicated by the light green overlay marking the region of interest in Figure 7 (A, B), in approximately 50 sections per embryo. The results show that barx1 morphants exhibited 1.5 to 2.5-fold fewer dividing cells in the mesenchymal regions than control embryos at 48 and 56 hpf, but similar numbers at different time points (Fig. 7C). Since the decrease in PH3 labeled nuclei within the pharyngeal arch mesenchyme is at a critical period of arch outgrowth, our data suggest that barx1 is involved in proliferation of the arch osteochondrogenic progenitors.
Figure 7.
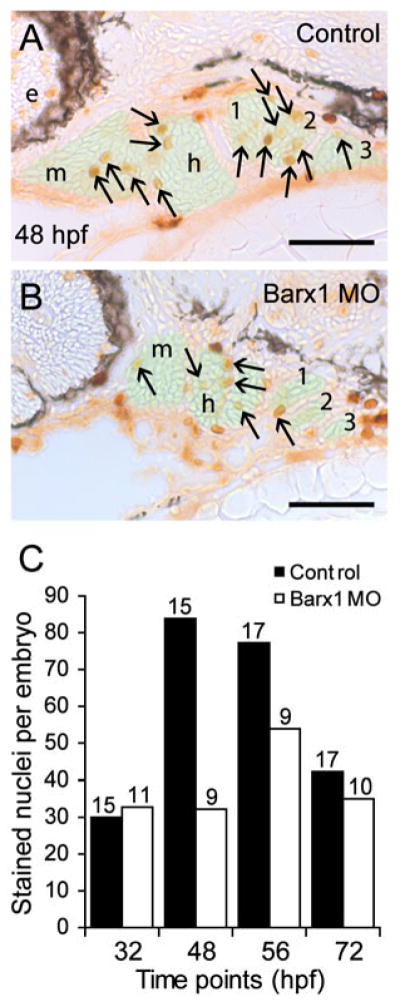
Barx1 positively influences cellular proliferation within the pharyngeal arches. (A, B) Sagittal serial sections (10 μm) of the pharyngeal arch region stained with Phosphohistone-H3 antibody. (A) Control embryo at 48 hpf. (B) barx1 morphant. The neural crest derived mesenchyme is overlaid in green; arrows indicate stained nuclei; e, eye; m, mandibular arch; h, hyoid arch; gill arches are numbered. Anterior is to the left. Scale bar: 50 μm. (C) Averaged cell proliferation in control uninjected embryos (black bars) and barx1 morphants (see Fig. 4; white bars) was determined within the mesenchyme as shown in (A,B) at the indicated four time points. Approximately fifty serial sections per embryo were examined. Number of embryos examined is indicated above each bar. Scale bar: (A, B) 50 μm.
FGF signaling is necessary for barx1 expression in the pharyngeal arches
FGF signaling is required for proper pharyngeal arch patterning (Abu-Issa et al., 2002; David et al., 2002; Frank et al., 2002; Trokovic et al., 2003; Walshe and Mason, 2003; Crump et al., 2004; Trokovic et al., 2005). In chick and mouse, FGFs emanating from the oral ectoderm positively influence the underlying Barx1 mesenchymal expression of the first arch (Tucker et al., 1998; Barlow et al., 1999). In zebrafish, fgf8 and fgf3 are expressed in the pharyngeal arch endoderm (Walshe and Mason, 2003). To determine the influence of FGF signaling on barx1 expression in zebrafish, we examined the fgf3 (lia) and the fgf8 (ace) mutants. In fgf3 mutants, larvae exhibit malformed mandibular and hyoid derived cartilages, and lack more posterior cartilages in the gill arches (Crump et al., 2004; Herzog et al., 2004). Expression of barx1 was similarly affected in that transcripts were detected in the anterior arches but were reduced or lacking in the gill arches (arrows, Fig. 8A, B).
Figure 8.

Barx1 expression in fgf mutant embryos. (A, B) Wild-type sibling and lia/fgf3 mutant. Arrows indicate sites of barx1 expression in the gill arches. (C, D and E) Wild-type sibling and left and right views of an ace/fgf8 mutant embryo. Left/right asymmetric expression indicated in the mutant (arrowhead); asterisk indicates reduced expression in the third gill arch. (F, G) Overstained control (DMSO) and embryo treated with the FGF receptor inhibitor SU5402 from 24 hpf to 48 hpf. Lateral views with anterior to the left, panel (D) was flipped for comparison; d, dorsal and v, ventral expression domains of the second arch; e, eye; gill arches are numbered; g, gut primordium; h, hyoid arch; m, mandibular arch; ov, otic vesicle. Scale bar: (A–G) 50 μm.
In fgf8 mutants, left/right asymmetries were previously observed in pharyngeal arch cartilage patterning (Albertson and Yelick, 2005). Therefore, using barx1 as a marker of chondrocyte condensation, we examined ace mutants for differences between control embryos and the left- and the right-hand sides of mutant embryos. Subtle differences were observed in the cellular condensation within the stacked arrays on the two sides of the same embryo in ace mutants (Fig. 8C–E); the differences included reduced (black arrowheads) or absence of barx1 expression (asterisk). As multiple FGF ligands are expressed in the embryo, we blocked all FGF receptor signaling with the pharmacological inhibitor SU5402 (Mohammadi et al., 1997). Embryos soaked in the inhibitor beginning at 24 hpf and compared to control siblings at 48 hpf lose barx1 expression (Fig. 8F, G). These data indicate that FGF signaling is necessary for maintaining barx1 expression in the arches and suggests that several FGF ligands are involved in this process.
barx1 is influenced by exogenous BMP4 signaling within the hyoid arch intermediate zone
Signaling factors that include BMPs emanating from the encapsulating tissues are necessary for patterning the pharyngeal arches and influencing subsequent chondrogenesis (Hall and Miyake, 2000; Helms and Schneider, 2003; Goldring et al., 2006; Nie et al., 2006b). We examined the effects of exogenous BMP4 signaling on barx1 expression. The effectiveness of using beads coated in BMPs have previously been demonstrated in chick and mouse organ cultures as well as in zebrafish embryos (Barlow and Francis-West, 1997; Tucker et al., 1998; Holzschuh et al., 2005). Beads were implanted on the left side at 24 hpf and the embryos were allowed to develop until 48 hpf. Control embryos were implanted with beads coated in 1% BSA (Fig. 9A, A′), and exhibited identical barx1 expression on their left and right-hand sides (n=17/17). Embryos implanted with BMP4 coated beads expressed barx1 within the intermediate domain of the second (hyoid) arch, where expression is not normally observed (open arrowhead in Fig. 9B, B′; n=4/7). Embryos implanted with BMP4 (n=11) beads at 36 hpf and examined at 60 hpf did not show any difference in barx1 expression compared to the control side (data not shown). These observations indicate that BMP4 can lead to ectopic barx1 expression within the hyoid intermediate zone, indicating a regional and time-dependent specificity in the response.
Figure 9.

Exogenous BMP signaling influences barx1 expression within the zebrafish pharyngeal arches. (A, B) Whole-mount in situ hybridization of barx1 in the left (bead implanted) and right (control) side of single embryos. Right-side images were flipped for easier comparison. (A, A′) Control BSA white bead, (B, B′) BMP4 blue bead. Anterior is to the left; d and v, dorsal and ventral expression domains in the second arch; e, eye. Arrowhead indicates sites of barx1 misexpression; asterisk indicates intermediate region, normally devoid of barx1 expression at this stage. Number of affected embryos over the total embryos with successful bead implantation is indicated in the panels (A, B). Black dashed line outlines the bead. Scale bar: 50 μm.
Discussion
Endochondral bone formation is preceded by chondrogenesis that forms the anlage for shaping of the bones (Hall and Miyake, 2000; Karsenty and Wagner, 2002; Goldring et al., 2006). The viscerocranium consists of highly adapted skeletal elements necessary for mastication, auditory amplification, and vocalization (Sperber and Sperber, 1996). How these elements are patterned is not well understood. Here we report analysis of the zebrafish barx1 transcription factor expression, its functional role in patterning chondrocyte condensation within the pharyngeal arches and the influence of morphogenic cues on its expression. Our experiments indicate that zebrafish barx1 is essential for proper patterning of the osteochondrogenic progenitors within the pharyngeal arches.
Zebrafish barx1
We have characterized the zebrafish barx1 gene and compared it to its paralogue, barx2. The expression of barx1 in the zebrafish embryo shows both conserved and divergent patterns compared to the mouse and chick. In the mouse, Barx1 is expressed in the mesenchyme of the maxillary facial prominences restricted in the first and second arches to the lateral aspects abutting the stomodeum and the pharyngeal cavity, and is prominently found in the molar field, yet lacking in Meckel’s cartilage condensation (Tissier-Seta et al., 1995). Expression is also observed in the mouse olfactory epithelium. In chick, Barx1 is expressed in the mesenchyme and the epithelium of the facial primordia and is detected after neural crest migration into the arches (Barlow et al., 1999). In contrast, zebrafish barx1 expression is detected early in the migrating hindbrain-derived neural crest and subsequently throughout the pharyngeal arch mesenchyme. During pharyngogenesis, barx1 expression is apparently enriched in putative proliferative regions located for the most part in rostral and ventral aspects of the arches (Walker et al., 2006; Walker et al., 2007). As chondrogenesis progresses, barx1 expression is down regulated in arrayed cells (Fig. 2I) while being maintained in the surrounding mesenchyme, suggesting that loss of barx1 expression may be a prerequisite for osteogenic differentiation. Barx1 expression was not detected in the ethmoid plate or trabeculae of the neurocranium, even though midbrain-hindbrain neural crest contributes to these structures, which undergo endochondral bone formation (Eberhart et al., 2006). These observations illustrate genetic differences in the chondrogenic program of neural crest cells derived from a similar location undergoing condensation within different elements of the chondrocranium. In comparison, barx2 is expressed most prominently in the oral and aboral epithelium of the first and second arches as well as in the olfactory epithelium, but not in the gut and therefore divergent of the observed pattern in higher vertebrates (Tissier-Seta et al., 1995; Barlow et al., 1999; Sander and Powell, 2004). Comparison of these patterns suggests that an alternate evolutionary subfunctionalization of the barx genes has occurred in teleosts in contrast to higher vertebrate model systems. Barx1 expression was also conserved in the developing gut wall in zebrafish as in the mouse and chick, as well as in the pectoral fin/limb buds. Transient expression was detected in the posterior lateral line ganglia at 72 hpf. The primary lateral line is established between 55 and 72 hpf (Ledent, 2002), and its expression suggests that barx1 may have a role in the differentiation or specification of this structure. Additional transient expression from 24 to 32 hpf was observed in the somite chevrons of the trunk and tail.
Initial arch patterning is maintained in barx1 morphants
The loss of barx1 function does not affect the specification of the cranial neural crest or the initial patterning of the pharyngeal arches as expression of neural crest markers dlx2a and inca are maintained during migration in barx1 morphants. Initial arch patterning appears normal as the pouches form when visualized in the fli1:GFP transgenic line. The behavior of other derivatives of the neural crest, such as aspects of the facial ganglia which express neuroD, and melanophores responsible for pigment in the morphants, indicate that their respective differentiation is maintained. As development progresses, however, we observed dysmorphogenesis within the pouches, presumably because of reduced cellular proliferation and compromised cellular condensation.
Chondrocyte differentiation requires barx1 in the pharyngeal arches
Two phases have been recognized in cartilage morphogenesis of the zebrafish pharyngeal arches, deposition of matrix by condensed chondrocytes, and a period of rapid growth (Kimmel et al., 1998). Here we show that attenuation of barx1 resulted in poor facial outgrowth due to fewer chondrocytes and poor deposition of matrix, causing dysmorphic cartilage elements as shown by poor fusion between the symplectic and hyomandibular elements (Fig. 3H). In other systems Barx genes are expressed in articular cartilage and the epiphyseal growth plate (Tissier-Seta et al., 1995; Barlow et al., 1999; Church et al., 2005; Meech et al., 2005), and that attenuation of Barx2 in mouse limb bud cultures disrupts primary mesenchymal condensations (Meech et al., 2005). Barx genes regulate the cell adhesion molecule NCAM (Hirsch et al., 1991; Edelman et al., 2000), which is expressed in aggregating prechondrocytes (Widelitz et al., 1993; DeLise et al., 2000; Hall and Miyake, 2000), and the fibril-forming type II collagen (Col2a1), the main component of cartilage extracellular matrix (Meech et al., 2005). We observed that barx1 morphants cause the perturbation of osteogenic promoting factors that include runx2b and chondromodulin (Fig. 5). Additionally, factors co-expressed in the arch neural crest, dlx2b and sox9a, which are required for chondrogenesis maintain their expression in the barx1 morphants. These observations suggest that barx1 functions at the earliest stages of prechondrocyte aggregation in an apparently independent regulatory mechanism compared to other cartilage and bone associated transcription factors. We further observed an enhancement in chordin and gdf5 expression within the jaw joint of morphants. Meech et al. (2005) showed that Gdf5 is able to induce Barx2 expression suggesting a functional connection. Here we show that the loss of the barx1 induces gdf5 expression suggesting a potential negative feedback regulatory mechanism that may function to balance their respective roles in chondrogenesis and joint patterning.
Ectopic BMP influences barx1 hyoid patterning
Reciprocal epithelial-mesenchymal interactions mediated by BMPs are essential for pharyngeal arch patterning and chondrogenesis (Tucker et al., 1998; Barlow et al., 1999; Tucker et al., 1999; Mina et al., 2002; Nie et al., 2006b). During chondrogenesis, FGF and BMP signaling is thought to act antagonistically, balancing proliferation and terminal differentiation in a complex regulatory network (Minina et al., 2002; Ornitz, 2005; Nie et al., 2006a; Nie et al., 2006b). In higher vertebrates, FGF8 and BMP2/4 induce and negatively influence Barx1 expression, respectively, in the maxillary and mandibular prominences (Tucker et al., 1998; Barlow et al., 1999). Conversely, in limb cultures, BMP4 positively influences Barx2 expression (Meech et al., 2005). Fgf8 and Fgf3 emanating from the endoderm, participate in the segmentation of the arches in zebrafish by promoting neural crest survival and cellular proliferation (Walshe and Mason, 2003; Crump et al., 2004). In our report, fgf3 and fgf8 deficient fish perturb but do not abolish barx1 expression. However, incubation with SU5402, a FGF receptor antagonist completely abolishes barx1 expression. This data suggests that multiple FGFs are necessary to regulate and maintain barx1 expression in an apparent coordinate manner.
BMP signaling is involved at different stages of the chondrogenic program (Yoon and Lyons, 2004), and BMPs emanate from the pouch endoderm of the zebrafish arches (Holzschuh et al., 2005). BMP2/4 beads implanted in the chick mandible show the loss of surrounding Barx1 expression (Tucker et al., 1998; Barlow et al., 1999). However, we observed that BMP4 bead implantation induced ectopic barx1 expression within the receptive cells of the second arch intermediate region between dorsally and ventrally located expression domains. Taken together, these reports and our data suggest that barx1 expression influenced by BMP cues reflects the cellular proliferation and extracellular matrix production necessary for shaping of cartilage elements within the viscerocranium.
Supplementary Material
Figure S1. Alignment of amino acid sequences of human (h) BARX1, -2, mouse (m) Barx1 and -2, chick (c) Barx1, -2 and zebrafish (z) Barx1, -2. Identical residues are highlighted in yellow, conserved residues are highlighted in green. The red dotted underline indicates the engrailed homology domain; the blue underline indicates the homeodomain; the green double underline indicates the Bar Basic Region (BBR).
Figure S2. Whole-mount in situ hybridization of barx1 expression. (A–B) 24 hpf, (C) 32 hpf, (D) 56 hpf, and (E–I) 72 hpf. (A) tail expression. (B) cross-section of the trunk, open arrowheads indicates barx1 expression. (D) Coronal section of the fifth branchial arch adjacent to the tooth bud primordium (asterisk). (E) lateral view of pharyngeal arch expression, posterior lateral line ganglia (pllg), and fin (f) expression domains. (F) dorsal view indicating coronal serial sections in panels (G, H and I). (G) expression in the arch mesenchyme of the jaw. (H) cross-section through the pllg. (H inset) higher magnification of expression in the posterior lateral line ganglia; dotted line outlines ganglion. (I) cross-section through the gut wall (g). f, fin; FB, forebrain; M, Meckel’s cartilage; nc, notochord; nt, neural tube; ov, otic vesicle; tb, tooth bud. Scale bar: (A) 100; μm, (B, D, G, H inset and I) 50 μm.
Figure S3. Whole-mount in situ hybridization of barx2 expression. (A) dorsal view, anterior is to the left. (B, D, H) lateral views, anterior to the left, (C) dorsal view; (E) ventral view; (C and E) anterior is to the top. (F) lateral oblique view. (F inset) sagittal section. (G) coronal section. (A, B) black arrowheads indicate bilateral sites of expression in the proximal aspect of the fin bud primordia. ae, aboral epithelium; bc, buccal cavity; e, eye; f, fin; fb, fin bud; h, hyoid arch; oe, oral epithelium; ole, olfactory bulb epithelium; ov, otic vesicle; m, mandibular arch. Scale bar: (A–F and H) 100 μm; (F inset and G) 50 μm.
Figure S4. Perturbation of pharyngeal arch outgrowth and chondrocyte condensation in barx1 morphants. (A, C and E) Control MO injected embryos, (B, D and F) barx1 morphants (see Fig. 4). (A, B) Pharyngeal arch sagittal sections at 72 hpf (See Fig. 3I and J). Arrow indicates the extracellular matrix surrounding the stacked chondrocytes. (C–F) fli1:GFP transgenic line at (C, D) 32 hpf and (E, F) 48 hpf. ch, ceratohyal cartilage; M, Meckel’s cartilage; m, mandibular arch; gill arches are numbered; h, hyoid arch; ov, otic vesicle. Anterior is to the left in all panels. Scale bars: (A–F) 100 μm.
Acknowledgments
We thank Pfizer Inc. for the generous gift of the SU5402, Dr. Chisa Shukunami for chondromodulin, Dr. Matthias Hammerschmidt for the lia embryos; Drs. Tom Sargent and Ting Luo for the inca probe, Dr. Marc Ekker for the dlx2b probe, our laboratory colleagues for helpful discussions, Dr. G.H. Sperber and the NIH Fellows Editorial Board for their comments on the manuscript, and Mark Rath and John Gonzales for their assistance with zebrafish husbandry.
References
- Abu-Issa R, Smyth G, Smoak I, Yamamura K-i, Meyers EN. Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development. 2002;129:4613–4625. doi: 10.1242/dev.129.19.4613. [DOI] [PubMed] [Google Scholar]
- Akimenko MA, Ekker M, Wegner J, Lin W, Westerfield M. Combinatorial expression of three zebrafish genes related to distal-less: part of a homeobox gene code for the head. J Neurosci. 1994;14:3475–86. doi: 10.1523/JNEUROSCI.14-06-03475.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albertson RC, Yelick PC. Roles for fgf8 signaling in left-right patterning of the visceral organs and craniofacial skeleton. Dev Biol. 2005;283:310–21. doi: 10.1016/j.ydbio.2005.04.025. [DOI] [PubMed] [Google Scholar]
- Angelo S, Lohr J, Lee KH, Ticho BS, Breitbart RE, Hill S, Yost HJ, Srivastava D. Conservation of sequence and expression of Xenopus and zebrafish dHAND during cardiac, branchial arch and lateral mesoderm development. Mech Dev. 2000;95:231–7. doi: 10.1016/s0925-4773(00)00334-8. [DOI] [PubMed] [Google Scholar]
- Barlow AJ, Bogardi JP, Ladher R, Francis-West PH. Expression of chick Barx-1 and its differential regulation by FGF-8 and BMP signaling in the maxillary primordia. Dev Dyn. 1999;214:291–302. doi: 10.1002/(SICI)1097-0177(199904)214:4<291::AID-AJA2>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Barlow AJ, Francis-West PH. Ectopic application of recombinant BMP-2 and BMP-4 can change patterning of developing chick facial primordia. Development. 1997;124:391–8. doi: 10.1242/dev.124.2.391. [DOI] [PubMed] [Google Scholar]
- Chiang EF, Pai CI, Wyatt M, Yan YL, Postlethwait J, Chung B. Two sox9 genes on duplicated zebrafish chromosomes: expression of similar transcription activators in distinct sites. Dev Biol. 2001;231:149–63. doi: 10.1006/dbio.2000.0129. [DOI] [PubMed] [Google Scholar]
- Church V, Yamaguchi K, Tsang P, Akita K, Logan C, Francis-West P. Expression and function of Bapx1 during chick limb development. Anat Embryol (Berl) 2005;209:461–9. doi: 10.1007/s00429-005-0464-z. [DOI] [PubMed] [Google Scholar]
- Crump JG, Maves L, Lawson ND, Weinstein BM, Kimmel CB. An essential role for Fgfs in endodermal pouch formation influences later craniofacial skeletal patterning. Development. 2004;131:5703–16. doi: 10.1242/dev.01444. [DOI] [PubMed] [Google Scholar]
- David NB, Saint-Etienne L, Tsang M, Schilling TF, Rosa FM. Requirement for endoderm and FGF3 in ventral head skeleton formation. Development. 2002;129:4457–4468. doi: 10.1242/dev.129.19.4457. [DOI] [PubMed] [Google Scholar]
- de Crombrugghe B, Lefebvre V, Behringer RR, Bi W, Murakami S, Huang W. Transcriptional mechanisms of chondrocyte differentiation. Matrix Biol. 2000;19:389–94. doi: 10.1016/s0945-053x(00)00094-9. [DOI] [PubMed] [Google Scholar]
- DeLise AM, Fischer L, Tuan RS. Cellular interactions and signaling in cartilage development. Osteoarthritis and Cartilage. 2000;8:309–334. doi: 10.1053/joca.1999.0306. [DOI] [PubMed] [Google Scholar]
- Eberhart JK, Swartz ME, Crump JG, Kimmel CB. Early Hedgehog signaling from neural to oral epithelium organizes anterior craniofacial development. Development. 2006;133:1069–1077. doi: 10.1242/dev.02281. [DOI] [PubMed] [Google Scholar]
- Edelman DB, Meech R, Jones FS. The homeodomain protein Barx2 contains activator and repressor domains and interacts with members of the CREB family. J Biol Chem. 2000;275:21737–45. doi: 10.1074/jbc.M909998199. [DOI] [PubMed] [Google Scholar]
- Ellies DL, Langille RM, Martin CC, Akimenko MA, Ekker M. Specific craniofacial cartilage dysmorphogenesis coincides with a loss of dlx gene expression in retinoic acid-treated zebrafish embryos. Mech Dev. 1997;61:23–36. doi: 10.1016/s0925-4773(96)00616-8. [DOI] [PubMed] [Google Scholar]
- Flores MV, Lam EY, Crosier P, Crosier K. A hierarchy of Runx transcription factors modulate the onset of chondrogenesis in craniofacial endochondral bones in zebrafish. Dev Dyn. 2006;235:3166–76. doi: 10.1002/dvdy.20957. [DOI] [PubMed] [Google Scholar]
- Flores MV, Tsang VW, Hu W, Kalev-Zylinska M, Postlethwait J, Crosier P, Crosier K, Fisher S. Duplicate zebrafish runx2 orthologues are expressed in developing skeletal elements. Gene Expr Patterns. 2004;4:573–81. doi: 10.1016/j.modgep.2004.01.016. [DOI] [PubMed] [Google Scholar]
- Frank DU, Fotheringham LK, Brewer JA, Muglia LJ, Tristani-Firouzi M, Capecchi MR, Moon AM. An Fgf8 mouse mutant phenocopies human 22q11 deletion syndrome. Development. 2002;129:4591–603. doi: 10.1242/dev.129.19.4591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldring MB, Tsuchimochi K, Ijiri K. The control of chondrogenesis. J Cell Biochem. 2006;97:33–44. doi: 10.1002/jcb.20652. [DOI] [PubMed] [Google Scholar]
- Gould DB, Walter MA. Cloning, characterization, localization, and mutational screening of the human BARX1 gene. Genomics. 2000;68:336–42. doi: 10.1006/geno.2000.6307. [DOI] [PubMed] [Google Scholar]
- Gould DB, Walter MA. Mutational analysis of BARHL1 and BARX1 in three new patients with Joubert syndrome. Am J Med Genet A. 2004;131:205–8. doi: 10.1002/ajmg.a.30227. [DOI] [PubMed] [Google Scholar]
- Hall BK, Miyake T. All for one and one for all: condensations and the initiation of skeletal development. BioEssays. 2000;22:138–147. doi: 10.1002/(SICI)1521-1878(200002)22:2<138::AID-BIES5>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- Helms JA, Schneider RA. Cranial skeletal biology. Nature. 2003;423:326–31. doi: 10.1038/nature01656. [DOI] [PubMed] [Google Scholar]
- Herzog W, Sonntag C, von der Hardt S, Roehl HH, Varga ZM, Hammerschmidt M. Fgf3 signaling from the ventral diencephalon is required for early specification and subsequent survival of the zebrafish adenohypophysis. Development. 2004;131:3681–92. doi: 10.1242/dev.01235. [DOI] [PubMed] [Google Scholar]
- Hirsch M, Valarche I, Deagostini-Bazin H, Pernelle C, Joliot A, Goridis C. An upstream regulatory element of the NCAM promoter contains a binding site for homeodomains. FEBS Lett. 1991;287:197–202. doi: 10.1016/0014-5793(91)80050-d. [DOI] [PubMed] [Google Scholar]
- Hjalt TA, Murray JC. The human BARX2 gene: genomic structure, chromosomal localization, and single nucleotide polymorphisms. Genomics. 1999;62:456–9. doi: 10.1006/geno.1999.6037. [DOI] [PubMed] [Google Scholar]
- Holzschuh J, Wada N, Wada C, Schaffer A, Javidan Y, Tallafuss A, Bally-Cuif L, Schilling TF. Requirements for endoderm and BMP signaling in sensory neurogenesis in zebrafish. Development. 2005;132:3731–42. doi: 10.1242/dev.01936. [DOI] [PubMed] [Google Scholar]
- Jackman WR, Draper BW, Stock DW. Fgf signaling is required for zebrafish tooth development. Dev Biol. 2004;274:139–157. doi: 10.1016/j.ydbio.2004.07.003. [DOI] [PubMed] [Google Scholar]
- Jones FS, Kioussi C, Copertino DW, Kallunki P, Holst BD, Edelman GM. Barx2, a new homeobox gene of the Bar class, is expressed in neural and craniofacial structures during development. Proc Natl Acad Sci U S A. 1997;94:2632–7. doi: 10.1073/pnas.94.6.2632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karsenty G, Wagner EF. Reaching a genetic and molecular understanding of skeletal development. Dev Cell. 2002;2:389–406. doi: 10.1016/s1534-5807(02)00157-0. [DOI] [PubMed] [Google Scholar]
- Kim B-M, Buchner G, Miletch I, Sharpe PT, Shivdasani RA. The Stomach Mesenchymal Transcription Factor Barx1 Specifies Gastric Epithelial Identity through Inhibition of Transient Wnt Signaling. Dev Cell. 2005;8:611–622. doi: 10.1016/j.devcel.2005.01.015. [DOI] [PubMed] [Google Scholar]
- Kimmel CB, Miller CT, Kruze G, Ullmann B, BreMiller RA, Larison KD, Snyder HC. The shaping of pharyngeal cartilages during early development of the zebrafish. Dev Biol. 1998;203:245–63. doi: 10.1006/dbio.1998.9016. [DOI] [PubMed] [Google Scholar]
- Korzh V, Sleptsova I, Liao J, He J, Gong Z. Expression of zebrafish bHLH genes ngn1 and nrd defines distinct stages of neural differentiation. Dev Dyn. 1998;213:92–104. doi: 10.1002/(SICI)1097-0177(199809)213:1<92::AID-AJA9>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- Lawson ND, Weinstein BM. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev Biol. 2002;248:307–18. doi: 10.1006/dbio.2002.0711. [DOI] [PubMed] [Google Scholar]
- Ledent V. Postembryonic development of the posterior lateral line in zebrafish. Development. 2002;129:597–604. doi: 10.1242/dev.129.3.597. [DOI] [PubMed] [Google Scholar]
- Luo R, An M, Arduini BL, Henion PD. Specific pan-neural crest expression of zebrafish Crestin throughout embryonic development. Dev Dyn. 2001;220:169–74. doi: 10.1002/1097-0177(2000)9999:9999<::AID-DVDY1097>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- Luo T, Xu Y, Hoffman TL, Zhang T, Schilling T, Sargent TD. Inca: a novel p21-activated kinase-associated protein required for cranial neural crest development. Development. 2007;134:1279–1289. doi: 10.1242/dev.02813. [DOI] [PubMed] [Google Scholar]
- Meech R, Edelman DB, Jones FS, Makarenkova HP. The homeobox transcription factor Barx2 regulates chondrogenesis during limb development. Development. 2005;132:2135–46. doi: 10.1242/dev.01811. [DOI] [PubMed] [Google Scholar]
- Miletich I, Buchner G, Sharpe PT. Barx1 and evolutionary changes in feeding. Journal of Anatomy. 2005;207:619–622. doi: 10.1111/j.1469-7580.2005.00486.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller CT, Yelon D, Stainier DY, Kimmel CB. Two endothelin 1 effectors, hand2 and bapx1, pattern ventral pharyngeal cartilage and the jaw joint. Development. 2003;130:1353–65. doi: 10.1242/dev.00339. [DOI] [PubMed] [Google Scholar]
- Mina M, Wang YH, Ivanisevic AM, Upholt WB, Rodgers B. Region- and stage-specific effects of FGFs and BMPs in chick mandibular morphogenesis. Dev Dyn. 2002;223:333–52. doi: 10.1002/dvdy.10056. [DOI] [PubMed] [Google Scholar]
- Minina E, Kreschel C, Naski MC, Ornitz DM, Vortkamp A. Interaction of FGF, Ihh/Pthlh, and BMP signaling integrates chondrocyte proliferation and hypertrophic differentiation. Dev Cell. 2002;3:439–49. doi: 10.1016/s1534-5807(02)00261-7. [DOI] [PubMed] [Google Scholar]
- Mitsiadis TA, Drouin J. Deletion of the Pitx1 genomic locus affects mandibular tooth morphogenesis and expression of the Barx1 and Tbx1 genes. Dev Biol. 2008;313:887–96. doi: 10.1016/j.ydbio.2007.10.055. [DOI] [PubMed] [Google Scholar]
- Mohammadi M, McMahon G, Sun L, Tang C, Hirth P, Yeh BK, Hubbard SR, Schlessinger J. Structures of the tyrosine kinase domain of fibroblast growth factor receptor in complex with inhibitors. Science. 1997;276:955–60. doi: 10.1126/science.276.5314.955. [DOI] [PubMed] [Google Scholar]
- Motoike T, Loughna S, Perens E, Roman BL, Liao W, Chau TC, Richardson CD, Kawate T, Kuno J, Weinstein BM, Stainier DY, Sato TN. Universal GFP reporter for the study of vascular development. Genesis. 2000;28:75–81. doi: 10.1002/1526-968x(200010)28:2<75::aid-gene50>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- Nie X, Luukko K, Kettunen P. FGF signalling in craniofacial development and developmental disorders. Oral Dis. 2006a;12:102–11. doi: 10.1111/j.1601-0825.2005.01176.x. [DOI] [PubMed] [Google Scholar]
- Nie X, Luukko K, Kettunen P. BMP signalling in craniofacial development. Int J Dev Biol. 2006b;50:511–21. doi: 10.1387/ijdb.052101xn. [DOI] [PubMed] [Google Scholar]
- Ornitz DM. FGF signaling in the developing endochondral skeleton. Cytokine Growth Factor Rev. 2005;16:205–13. doi: 10.1016/j.cytogfr.2005.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redon R, Baujat G, Sanlaville D, Le Merrer M, Vekemans M, Munnich A, Carter NP, Cormier-Daire V, Colleaux L. Interstitial 9q22.3 microdeletion: clinical and molecular characterisation of a newly recognised overgrowth syndrome. Eur J Hum Genet. 2006;14:759–67. doi: 10.1038/sj.ejhg.5201613. [DOI] [PubMed] [Google Scholar]
- Rubinstein AL, Lee D, Luo R, Henion PD, Halpern ME. Genes dependent on zebrafish cyclops function identified by AFLP differential gene expression screen. Genesis. 2000;26:86–97. doi: 10.1002/(sici)1526-968x(200001)26:1<86::aid-gene11>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- Sachdev SW, Dietz UH, Oshima Y, Lang MR, Knapik EW, Hiraki Y, Shukunami C. Sequence analysis of zebrafish chondromodulin-1 and expression profile in the notochord and chondrogenic regions during cartilage morphogenesis. Mechanisms of Development. 2001;105:157–162. doi: 10.1016/s0925-4773(01)00417-8. [DOI] [PubMed] [Google Scholar]
- Sander GR, Powell BC. Expression of the homeobox gene barx2 in the gut. J Histochem Cytochem. 2004;52:541–4. doi: 10.1177/002215540405200412. [DOI] [PubMed] [Google Scholar]
- Schulte-Merker S, Hammerschmidt M, Beuchle D, Cho KW, De Robertis EM, Nusslein-Volhard C. Expression of zebrafish goosecoid and no tail gene products in wild-type and mutant no tail embryos. Development. 1994;120:843–852. doi: 10.1242/dev.120.4.843. [DOI] [PubMed] [Google Scholar]
- Song Y, Zhang Z, Yu X, Yan M, Zhang X, Gu S, Stuart T, Liu C, Reiser J, Zhang Y, Chen Y. Application of lentivirus-mediated RNAi in studying gene function in mammalian tooth development. Dev Dyn. 2006;235:1347–1357. doi: 10.1002/dvdy.20706. [DOI] [PubMed] [Google Scholar]
- Sperber GH, Sperber SM. Pharyngogenesis. J Dent Assoc S Afr. 1996;51:777–82. [PubMed] [Google Scholar]
- Stachel SE, Grunwald DJ, Myers PZ. Lithium perturbation and goosecoid expression identify a dorsal specification pathway in the pregastrula zebrafish. Development. 1993;117:1261–74. doi: 10.1242/dev.117.4.1261. [DOI] [PubMed] [Google Scholar]
- Stalker H, Ayme S, Delneste D, Scarpelli H, Vekemans M, Der Kaloustian VM. Duplication of 9q12-q33: a case report and implications for the dup(9q) syndrome. Am J Med Genet. 1993;45:456–459. doi: 10.1002/ajmg.1320450412. [DOI] [PubMed] [Google Scholar]
- Tissier-Seta JP, Mucchielli ML, Mark M, Mattei MG, Goridis C, Brunet JF. Barx1, a new mouse homeodomain transcription factor expressed in cranio-facial ectomesenchyme and the stomach. Mech Dev. 1995;51:3–15. doi: 10.1016/0925-4773(94)00343-l. [DOI] [PubMed] [Google Scholar]
- Trokovic N, Trokovic R, Mai P, Partanen J. Fgfr1 regulates patterning of the pharyngeal region. Genes Dev. 2003;17:141–153. doi: 10.1101/gad.250703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trokovic N, Trokovic R, Partanen J. Fibroblast growth factor signalling and regional specification of the pharyngeal ectoderm. Int J Dev Biol. 2005;49:797–805. doi: 10.1387/ijdb.051976nt. [DOI] [PubMed] [Google Scholar]
- Tuan RS. Biology of developmental and regenerative skeletogenesis. Clin Orthop Relat Res. 2004:S105–17. doi: 10.1097/01.blo.0000143560.41767.ee. [DOI] [PubMed] [Google Scholar]
- Tucker AS, Matthews KL, Sharpe PT. Transformation of tooth type induced by inhibition of BMP signaling. Science. 1998;282:1136–8. doi: 10.1126/science.282.5391.1136. [DOI] [PubMed] [Google Scholar]
- Tucker AS, Yamada G, Grigoriou M, Pachnis V, Sharpe PT. Fgf-8 determines rostral-caudal polarity in the first branchial arch. Development. 1999;126:51–61. doi: 10.1242/dev.126.1.51. [DOI] [PubMed] [Google Scholar]
- Walker MB, Miller CT, Coffin Talbot J, Stock DW, Kimmel CB. Zebrafish furin mutants reveal intricacies in regulating Endothelin1 signaling in craniofacial patterning. Dev Biol. 2006;295:194–205. doi: 10.1016/j.ydbio.2006.03.028. [DOI] [PubMed] [Google Scholar]
- Walker MB, Miller CT, Swartz ME, Eberhart JK, Kimmel CB. phospholipase C, beta 3 is required for Endothelin1 regulation of pharyngeal arch patterning in zebrafish. Dev Biol. 2007;304:194–207. doi: 10.1016/j.ydbio.2006.12.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walshe J, Mason I. Fgf signalling is required for formation of cartilage in the head. Dev Biol. 2003;264:522–36. doi: 10.1016/j.ydbio.2003.08.010. [DOI] [PubMed] [Google Scholar]
- Westerfield M. The zebrafish book. University of Oregon Press; Eugene: 1995. [Google Scholar]
- Widelitz R, Jiang T, Murray B, Chuong C. Adhesion molecules in skeletogenesis: II. Neural cell adhesion molecules mediate precartilaginous mesenchymal condensations and enhance chondrogenesis. J Cell Physiol. 1993;156:399–411. doi: 10.1002/jcp.1041560224. [DOI] [PubMed] [Google Scholar]
- Yan YL, Hatta K, Riggleman B, Postlethwait JH. Expression of a type II collagen gene in the zebrafish embryonic axis. Dev Dyn. 1995;203:363–76. doi: 10.1002/aja.1002030308. [DOI] [PubMed] [Google Scholar]
- Yan YL, Miller CT, Nissen RM, Singer A, Liu D, Kirn A, Draper B, Willoughby J, Morcos PA, Amsterdam A, Chung BC, Westerfield M, Haffter P, Hopkins N, Kimmel C, Postlethwait JH, Nissen R. A zebrafish sox9 gene required for cartilage morphogenesis. Development. 2002;129:5065–79. doi: 10.1242/dev.129.21.5065. [DOI] [PubMed] [Google Scholar]
- Yan YL, Willoughby J, Liu D, Crump JG, Wilson C, Miller CT, Singer A, Kimmel C, Westerfield M, Postlethwait JH. A pair of Sox: distinct and overlapping functions of zebrafish sox9 co-orthologs in craniofacial and pectoral fin development. Development. 2005;132:1069–83. doi: 10.1242/dev.01674. [DOI] [PubMed] [Google Scholar]
- Yoon BS, Lyons KM. Multiple functions of BMPs in chondrogenesis. J Cell Biochem. 2004;93:93–103. doi: 10.1002/jcb.20211. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Alignment of amino acid sequences of human (h) BARX1, -2, mouse (m) Barx1 and -2, chick (c) Barx1, -2 and zebrafish (z) Barx1, -2. Identical residues are highlighted in yellow, conserved residues are highlighted in green. The red dotted underline indicates the engrailed homology domain; the blue underline indicates the homeodomain; the green double underline indicates the Bar Basic Region (BBR).
Figure S2. Whole-mount in situ hybridization of barx1 expression. (A–B) 24 hpf, (C) 32 hpf, (D) 56 hpf, and (E–I) 72 hpf. (A) tail expression. (B) cross-section of the trunk, open arrowheads indicates barx1 expression. (D) Coronal section of the fifth branchial arch adjacent to the tooth bud primordium (asterisk). (E) lateral view of pharyngeal arch expression, posterior lateral line ganglia (pllg), and fin (f) expression domains. (F) dorsal view indicating coronal serial sections in panels (G, H and I). (G) expression in the arch mesenchyme of the jaw. (H) cross-section through the pllg. (H inset) higher magnification of expression in the posterior lateral line ganglia; dotted line outlines ganglion. (I) cross-section through the gut wall (g). f, fin; FB, forebrain; M, Meckel’s cartilage; nc, notochord; nt, neural tube; ov, otic vesicle; tb, tooth bud. Scale bar: (A) 100; μm, (B, D, G, H inset and I) 50 μm.
Figure S3. Whole-mount in situ hybridization of barx2 expression. (A) dorsal view, anterior is to the left. (B, D, H) lateral views, anterior to the left, (C) dorsal view; (E) ventral view; (C and E) anterior is to the top. (F) lateral oblique view. (F inset) sagittal section. (G) coronal section. (A, B) black arrowheads indicate bilateral sites of expression in the proximal aspect of the fin bud primordia. ae, aboral epithelium; bc, buccal cavity; e, eye; f, fin; fb, fin bud; h, hyoid arch; oe, oral epithelium; ole, olfactory bulb epithelium; ov, otic vesicle; m, mandibular arch. Scale bar: (A–F and H) 100 μm; (F inset and G) 50 μm.
Figure S4. Perturbation of pharyngeal arch outgrowth and chondrocyte condensation in barx1 morphants. (A, C and E) Control MO injected embryos, (B, D and F) barx1 morphants (see Fig. 4). (A, B) Pharyngeal arch sagittal sections at 72 hpf (See Fig. 3I and J). Arrow indicates the extracellular matrix surrounding the stacked chondrocytes. (C–F) fli1:GFP transgenic line at (C, D) 32 hpf and (E, F) 48 hpf. ch, ceratohyal cartilage; M, Meckel’s cartilage; m, mandibular arch; gill arches are numbered; h, hyoid arch; ov, otic vesicle. Anterior is to the left in all panels. Scale bars: (A–F) 100 μm.