

Abstract
This paper is devoted to the quantization of the degree of nonlinearity of the relationship between two biological variables when one of the variables is a complex nonstationary oscillatory signal. An example of the situation is the indicial responses of pulmonary blood pressure (P) to step changes of oxygen tension (ΔpO2) in the breathing gas. For a step change of ΔpO2 beginning at time t1, the pulmonary blood pressure is a nonlinear function of time and ΔpO2, which can be written as P(t-t1 | ΔpO2). An effective method does not exist to examine the nonlinear function P(t-t1 | ΔpO2). A systematic approach is proposed here. The definitions of mean trends and oscillations about the means are the keys. With these keys a practical method of calculation is devised. We fit the mean trends of blood pressure with analytic functions of time, whose nonlinearity with respect to the oxygen level is clarified here. The associated oscillations about the mean can be transformed into Hilbert spectrum. An integration of the square of the Hilbert spectrum over frequency yields a measure of oscillatory energy, which is also a function of time, whose mean trends can be expressed by analytic functions. The degree of nonlinearity of the oscillatory energy with respect to the oxygen level also is clarified here. Theoretical extension of the experimental nonlinear indicial functions to arbitrary history of hypoxia is proposed. Application of the results to tissue remodeling and tissue engineering of blood vessels is discussed.
Keywords: degree of nonlinearity, Fourier spectrum, Hilbert spectrum, nonlinear relationships
In biomedical science, we often have to deal with variables that are stochastic, oscillatory, and nonstationary and the relationship of these variables to other chemical, mechanical, physical, and pharmacological variables. In the cardiovascular system blood pressure is such a variable. This paper illustrates the mathematical approach to deal with the question of linearity or nonlinearity of the dependence of blood pressure on other variables. As a specific illustration, we consider the changes that occur in the lung when a sea-level dwelling animal is flown to a ski resort at a higher altitude where the partial pressure of oxygen in the gas that the animal breathes is lower. What happens is that the pulmonary arterial blood pressure becomes higher (1–3), the arterial blood vessel wall becomes thicker (3–5), the different layers of the arteries thicken with different rates and different courses of time (2–6), the mechanical properties of the blood vessel wall change with specific historical courses (7–9), cells in the wall modify, grow, proliferate, or move (5, 6, 10–13), intercellular matrix and interstitial space change (14, 15), the stress and strain distribution in the vessel wall change with time in a specific way (16), and because of cellular and extracellular changes the zero-stress state of the blood vessel wall changes with time (7–9). The crucial fact is the blood pressure change, because the blood pressure imposes load on the blood vessel wall, causing stress and strain, and the subsequent tissue and mechanical properties remodeling are believed to be the results of these stress and strain changes. Therefore, the exact behavior of blood pressure when the oxygen tension changes is of paramount importance. But blood pressure is stochastic (see Fig. 1). To get a meaningful and precise description of the blood pressure history is the first step.
Figure 1.

Four typical sets of traces and IMFs of the pulmonary arterial pressure of rats subjected to a step lowering of pO2 at 6 h and a step rise of pO2 at 30 h. Lowest pO2: Column 1 20.9%; column 2 17.2%; column 3 13.6%; and column 4 10.0%.
We used a catheter that was implanted in the pulmonary arterial trunk of a rat to get the instantaneous reading of blood pressure continuously over many days in a simulated laboratory chamber. The oxygen tension in the breathing gas was controlled as a constant or varied as a step decrease in time or a step increase in time. Typical blood pressure records are shown in Fig. 1. Previous reports (17) have demonstrated various features of pulmonary hypertension caused by hypoxia, but no mathematical analysis of the blood pressure-oxygen relationship was shown. The present paper shows a systematic use of the method presented in refs. 18 and 19 to the study of the nonlinearity of the pulmonary blood pressure-oxygen tension relationship.
METHODS
Sixteen male Sprague–Dawley rats (Harlan, San Diego, CA), 358.9 ± 4.9 g body weight, were used in the study. The protocol and experimental methods are presented in refs. 19 and 20. Briefly, each rat was implanted with an indwelling catheter in the pulmonary arterial trunk when breathing a gaseous anesthetic of isoflurane. After the implantation of the catheter, the rat could move freely in a standard-sized cage in a quiet room that was illuminated from 0600 to 1800 h. The catheter tubing had 0.305 mm i.d. and 0.635 mm o.d. and floated in the pulmonary arterial trunk facing downstream. The other end of the tubing was connected to a Statham pressure transducer (P23ID, Hato Rey, PR). The pressure recorded therefore was the stagnation or total pressure. The pressure was recorded continuously by a computer at a sampling rate of 100 points/sec over a 36-h period with a time lag of 2 sec in every 60 sec for computer processing. The analog-to-digital conversion was accomplished by a data translation board (DT31-EZ, Data Translation, Marlboro, MA). After 6 h in normal sea-level air with 20.9% oxygen, the rats were exposed to a hypoxic gas containing nitrogen and 17.2% O2, 13.6% O2, or 10% O2 for 24 h followed by returning to breathing normal sea-level air for 6 h. The changeover from one level of oxygen tension to the next was accomplished in 1.5 ± 0.5 min each time. Hence, in the perspective of 1 day we say approximately that the oxygen level was changed as a step function of time. Four rats were used at each hypoxic level; four served as control without hypoxia.
For mathematical analysis of the pressure signal, we begin by considering the time between successive extrema as the local time scale and define an intrinsic mode function (IMF) as a function that satisfies two conditions: (i) in the whole data set, the number of extrema and the number of zero crossings must either equal or differ at most by one; and (ii) at any point, the mean value of the envelope defined by the local maxima and the envelope defined by the local minima is zero. Let X(t) be an oscillatory signal shown in Fig. 1. A practical computational way to obtain the first IMF is as follows. Let the successive maxima of X(t) be connected by an envelope with the method of cubic spline. Similarly the successive minima are connected by an envelope. The mean of these envelopes is computed and plotted and is designated m1(t). The difference,
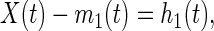 |
1 |
represents the oscillations of X(t) about m1(t). The function h1(t) is treated as a new signal whose upper and lower envelopes are constructed, and the new mean of the envelopes, m11(t), is computed. The process is continued until the mean of the envelopes becomes zero or insignificantly small. Then the converged oscillatory function is the first intrinsic mode and is designated as C1(t). The local mean of C1(t) is zero.
Next, the difference X(t) and C1(t) is computed and is called the first residue, R1(t),
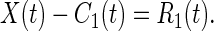 |
2 |
R1(t) is treated as a new signal and is analyzed as before to yield the second intrinsic mode, C2(t), whose local mean is zero. The local time scale (the time interval between successive extrema) of C2(t) is in general longer than that of C1. Iteration continues until the signal X(t) is resolved into a series of IMFs in the form
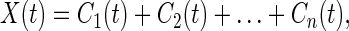 |
3 |
in which each term Ci(t) is an oscillatory mode with zero local mean, and the last mode, Cn(t), is nonoscillatory. The local time scale of the last mode is the total length of the data set.
The last mode, Cn(t), in Eq. 3 represents a mean trend. The sum Cn(t)+Cn−1(t) also represents a mean trend that is somewhat oscillatory. Generalizing, the set of functions
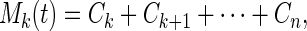 |
4 |
where 2 ≤ k ≤ n, represents a sequence of mean trends with various degree of oscillations. The lower the k, the more oscillation Mk(t) contains.
When the signal X(t) is influenced by another variable, S(t), we are interested in the correlation of X(t) and S(t). If S(t) represents a simple step function
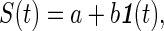 |
5 |
where 1(t) is a unit-step function, then both the IMFs C1(t), C2(t), … Cn(t) and the mean trends Mn(t), Mn−1(t), … Mk(t) are functions of a and b. Traditionally, the functions Mk(t) in response to a step S(t) as specified by Eq. 5 are called the indicial response functions.
When the oxygen tension changes as a step decrease beginning at time t0, the mean trend of the change in pulmonary arterial blood pressure may be represented by the equation
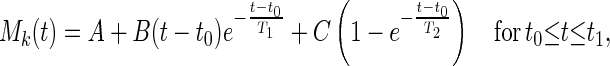 |
6 |
where t0 is the instant of time when O2 concentration drops suddenly, and t1 is the instant when O2 concentration increases suddenly. A is the mean value of Mk(t) before time t0; B, C, T1, and T2 are constants. On the other hand, for a step increase of oxygen tension beginning at time t1, the following equation with new constants D and T3 appears good enough:
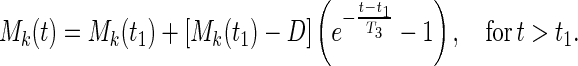 |
7 |
Conjugate to the mean trends defined by Eq. 4, the oscillations about the mean trend Mk(t) is defined as:
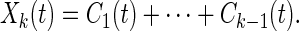 |
8 |
For oscillations about the mean, Hilbert transform allows us to define instantaneous frequency and amplitude of the signal (21). The Hilbert transform of X(t) is Y(t) defined by the reciprocal relationship:
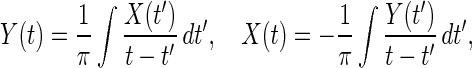 |
9 |
in which the integral is defined in the sense of Cauchy principal value (21). Titchmarsh (21) attributes the idea to consider the complex variable Z(t) = X(t) + iY(t) to Hilbert. Z(t) can be written in polar coordinates as:
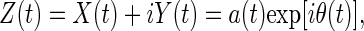 |
10 |
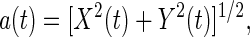 |
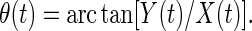 |
11 |
Huang et al. (18) defined the instantaneous frequency ω(t) as the derivative of θ(t) with respect to time t:
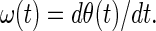 |
12 |
Then the amplitude a(t) of Eq. 10 and the frequency ω(t) of Eq. 12 are functions of time t and can be plotted in contour maps as shown in figures 2 and 3 of ref. 19. The frequency-time distribution of the amplitude is designated as the Hilbert amplitude spectrum, H(ω,t).
The vanishing of the local means of the functions C1, … Cn-1 is very important, because the Hilbert spectral amplitude a(t) and the phase angle θ(t) have unique physical meaning only if the local means are zero. The last mode, Cn, is the nonoscillatory residue and could contain no more than one maximum and one minimum. In most cases, this component is a monotonic function representing the trend of the data. This component is not involved in the spectral concept and should not be included in the instantaneous frequency computation unless justified independently based on physical considerations.
The Hilbert amplitude spectrum of Xk(t) may be designated as Hk(ω,t). The square of Hk(ω,t) represents an oscillatory energy density. We can define the oscillatory energy about the mean, Ek(t), by an integration over all frequencies:
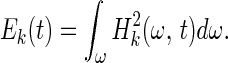 |
13 |
In practice, at any given time within a time interval of one heartbeat, there are only two or three frequencies that contribute to the energy Ek(t). Clearly, Ek(t) is another nonstationary stochastic variable that can be treated in the same manner as outlined above. Each Ek(t) has its IMFs, mean trends, and oscillations about the mean.
The degree of nonlinearity of the hypertension-hypoxia relationship is revealed by the dependence of the material constants A, B, …, T1, T2 … in Eqs. 6 and 7 as functions of the constants a and b in the step function given in Eq. 5.
RESULTS
Fig. 1 shows the records of the pulmonary blood pressure history (24- or 36-h traces) and their IMFs of four typical rats, one at each level of hypoxia. The records were plotted from the digital recording. The blood pressure was measured in units of cm H2O (≈ 98.063 N/m2 at 4°C). The concentration of oxygen was expressed in terms of % partial pressure in the breathing gas. In normal atmosphere the concentration of oxygen was 20.9%. We tested rats at pO2 = 20.9, 17.2, 13.6, and 10% atm (n = 4). The pO2 value is shown by straight line segments in Fig. 1 (Upper). The level of hypoxia is designated as −ΔpO2 and is equal to 20.9 minus the test concentration. The IMFs were computed as in refs. 18–20.
Fig. 2 shows the mean trends of the mean blood pressure (MBP) in response to a step decrease of oxygen tension (−ΔpO2) at 6 h and step increase of oxygen tension at 30 h. In each column, Fig. 2, Upper shows the Mk(t) as defined by Eq. 4, for k = 9, 10, ⋅ ⋅ ⋅ 18. Fig. 2, Lower shows M10 fitted by an analytic function in the form of Eqs. 6 and 7. Because the oxygen level was changed as step functions, the fitted analytic functions are the indicial response functions of the MBP in response to step change of oxygen level. The coefficients in the analytic functions of Eq. 6 are listed in Table 1 and are used to study the nonlinearity of the indicial functions with regard to the level of hypoxia (see Fig. 5A).
Figure 2.

Mean trends of the MBP and their analytical expressions. Mk(t)MBP is the mean trend of order k, a function representing the MBP history. The highest order, Mn(t), has no oscillations. The number n is data specific. Mk(t)MBP with k < n contains oscillations of various degree (see Eq. 4). (Upper) Mk(t) for various values of k. (Lower) One example at a larger scale and the curve fitting by Eqs. 6 and 7 is illustrated. The coefficients of the fitted curves are listed in Table 1 except for the case of rat no. 1212, which was not subjected to hypoxia. For the normal rat 1212, the mean trends of order 16 and the analytic approximation of M16(t) are shown in the lowest left panel.
Table 1.
The coefficients of the analytic functions that fitted the kth-order trend Mk(t)MBP for the MBP history
| ΔpO2, % atm | Rat code | Order no. k of Mk(t)MBP | A | B | BT1 | T1 | C | T2 | D | A–D | T3 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| −3.7 | 0607 | 10 | 29.3 | 7 | 16.1 | 2.3 | 2.5 | 6 | 25.7 | 3.6 | 0.3 |
| 12 | 29.7 | 7 | 16.1 | 2.3 | 2.5 | 3 | 25.8 | 3.9 | 0.3 | ||
| 14 | 29.5 | 6 | 15.6 | 2.6 | 2.0 | 3 | 25.8 | 3.7 | 0.3 | ||
| 16 | 29.5 | 4 | 12.0 | 3.0 | 2.0 | 3 | 25.3 | 4.2 | 2.0 | ||
| −3.7 | 0601 | 10 | 27.6 | 2 | 7.0 | 3.5 | 1 | 6 | 26.9 | 0.7 | 0.5 |
| −3.7 | 0609 | 10 | 32.0 | 5 | 20.0 | 4.0 | 1 | 2 | 29.7 | 2.3 | 0.3 |
| −3.7 | 0621 | 10 | 27.5 | 3 | 10.5 | 3.5 | 1 | 5 | 26.1 | 1.4 | 0.5 |
| −7.3 | 0501 | 10 | 31.2 | 13 | 45.5 | 3.5 | 2.0 | 6.5 | 27.2 | 4.0 | 0.4 |
| 12 | 31.1 | 13 | 45.5 | 3.5 | 2.0 | 6.5 | 27.3 | 3.8 | 0.4 | ||
| 14 | 31.4 | 13 | 45.5 | 3.5 | 1.5 | 6.5 | 27.7 | 3.7 | 0.5 | ||
| 16 | 30.8 | 10 | 40.0 | 4.0 | 0.5 | 6.5 | 27.6 | 3.2 | 2.0 | ||
| −7.3 | 0427 | 10 | 29.2 | 8 | 32.0 | 4.0 | 1 | 5 | 25.6 | 3.6 | 0.3 |
| −7.3 | 0429 | 10 | 27.9 | 24 | 55.2 | 2.3 | 4 | 4 | 23.7 | 4.2 | 0.5 |
| −7.3 | 0504 | 10 | 30.3 | 13 | 52.0 | 4.0 | 3 | 2 | 29.1 | 1.2 | 0.5 |
| −10.9 | 0718 | 10 | 29.7 | 23 | 46 | 2.0 | 13 | 2 | 26.7 | 3.0 | 0.5 |
| 12 | 29.8 | 23 | 46 | 2.0 | 13 | 2 | 26.5 | 3.3 | 0.5 | ||
| 14 | 29.9 | 20 | 46 | 2.3 | 12 | 2 | 26.4 | 3.5 | 0.5 | ||
| 16 | 34.1 | 4.5 | 36 | 8.0 | 1 | 1 | 24.8 | 9.3 | 2.0 | ||
| −10.9 | 0417 | 10 | 28.0 | 15 | 58.5 | 3.9 | 16 | 8 | 27.9 | 0.1 | 0.5 |
| −10.9 | 0422 | 10 | 27.1 | 9 | 49.5 | 5.5 | 8 | 6 | 23.8 | 3.3 | 0.5 |
| −10.9 | 0508 | 10 | 33.4 | 40 | 40.0 | 1.0 | 13 | 2 | 27.2 | 6.2 | 0.5 |
The units of A, BT1, C, D, and A–D are cm H2O. The unit of B is cm H2O per h. The units of T1, T2, and T3 are h.
Figure 5.

(A) The degree of nonlinearity of the effects of the oxygen tension in breathing gas on the parameters that characterize the indicial responses of the MBP given in Eqs. 6 and 7 and Table 1 is shown. The physical meaning of the parameters are: A, the MBP (unit: cm H2O) at time t = t0; B, the amplitude of the second term (unit: cm H2O per h); BT1, the peak height of the MBP indicial function above A for step ΔpO2 (unit: cm H2O); C, the asymptotic hypertensive MBP above A as t→0 for step −ΔpO2 (unit: cm H2O); D, the asymptotic MBP as t→0 for step increase of ΔpO2 (unit: cm H2O); t0, time of step lowering of pO2 (unit: h); t1, time of step rising of pO2 (unit: h); T1, time constant for MBP rise (unit: h); T2, time constant for MBP asymptote as t→∞ (unit: h). (B) Similar to A except related to the energy of oscillations of the blood pressure about the mean. Mk(t) is the kth-order mean of the E6(t) about the sixth order of MBP. Data are from Table 2.
Fig. 3 shows the oscillations about the mean trends, Xk(t), as defined by Eq. 8, and the “energy” of the oscillations, Ek(t), as defined by Eq. 13. In each column, Xk(t) (k = 6, 8, 9, … 12), Ek(t) (k = 6, 8, 9), a large scale plot of X6(t), and a large scale plot of E6(t) are shown.
Figure 3.

Oscillations about the mean trends and their energy history. Xk(t) is the kth-order oscillations about the kth-order trend of the MBP, Mk(t)MBP (see Eqs. 4 and 8). The Hilbert spectrum of Xk(t) was computed. An integration of the square of the Hilbert amplitude spectrum over all frequencies yields the energy of oscillations (EO). EO(t) corresponding to Xk(t) is denoted by Ek(t). (Upper) Xk(t) (k = 6, 8, 9, … 12) and Ek(t) (k = 6, 8, 9). (Lower) X6(t) and E6(t) at a larger scale.
Fig. 4 shows the analysis of the energy of oscillation E6(t) shown in the lowest panel of Fig. 3. The results of IMFs, Mks, and Xks of E6(t) are shown. The Mk(t)s of E6(t) are fitted by analytic functions in the same manner as by Eqs. 6 and 7. The coefficients in the analytic functions are listed in Table 2. The correlation with hypoxia is shown in Fig. 5B.
Figure 4.

Analysis of the energy of oscillations E6(t), which is a stochastic signal with its own IMFs (shown in A), Mk(t)s (shown in B), and Xk(t)s. In C and D, Mk(t) of E6(t) and Xk(t) of E6(t) are shown for k = 12 and 9, respectively. They are fitted with analytic functions as in Eqs. 6 and 7. The coefficients of the fitted curves are given in Table 2, except for the normal rat 1221, whose results are shown in C.
Table 2.
The curve-fitted coefficients of the kth trend, Mk(t) of E6(t), of the energy of the oscillations of the signal Xk(t)MBP oscillating about the Mk(t)MBP
| ΔpO2, % atm | Rat code | Order no. k of Mk(t)E6 | A | B | BT1 | T1 | C | T2 | D | A–D | T3 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| −3.7 | 0607 | 9 | 10 | 1.5 | 5.25 | 3.5 | 4.0 | 6 | 12.5 | −2.5 | 1.5 |
| 11 | 10 | 1.5 | 5.25 | 3.5 | 4.0 | 6 | 12.5 | −2.5 | 1.5 | ||
| 13 | 10 | 1.5 | 5.25 | 3.5 | 4.0 | 6 | 13.5 | −3.5 | 1.5 | ||
| −3.7 | 0601 | 9 | 11 | 2.2 | 7.7 | 3.5 | 4.5 | 6 | 13.0 | −2.0 | 1.5 |
| −3.7 | 0609 | 9 | 12 | 1.0 | 3.5 | 3.5 | 6.0 | 6 | 16.5 | −3.5 | 1.5 |
| −3.7 | 0621 | 9 | 7 | 2.3 | 11.5 | 5.0 | 9.0 | 6 | 14.0 | −7.0 | 1.5 |
| −7.3 | 0427 | 9 | 7 | 1.3 | 5.2 | 4.0 | 1.7 | 6 | 4.5 | 2.5 | 1.5 |
| 11 | 7 | 1.3 | 5.2 | 4.0 | 1.7 | 6 | 4.5 | 2.5 | 1.5 | ||
| 13 | 7 | 1.3 | 5.2 | 4.0 | 1.7 | 6 | 5.3 | 1.7 | 1.5 | ||
| −7.3 | 0429 | 9 | 12 | 50.0 | 100.0 | 2.0 | 7.0 | 8 | 17.0 | −5.0 | 1.5 |
| −7.3 | 0501 | 9 | 13 | 22.0 | 66.0 | 3.0 | 8.0 | 6 | 18.0 | −5.0 | 1.5 |
| −7.3 | 0504 | 9 | 14 | 19.0 | 76.0 | 4.0 | 6.0 | 8 | 16.0 | −2.0 | 1.5 |
| −10.9 | 0417 | 9 | 14 | 35.0 | 122.5 | 3.5 | 15.0 | 4 | 20.0 | −6.0 | 1.5 |
| 11 | 14 | 35.0 | 122.5 | 3.5 | 15.0 | 4 | 20.0 | −6.0 | 1.5 | ||
| 13 | 14 | 30.0 | 105.0 | 3.5 | 15.0 | 4 | 23.0 | −9.0 | 1.5 | ||
| −10.9 | 0422 | 9 | 22 | 40.0 | 200.0 | 5.0 | 30.0 | 8 | 40.0 | −18.0 | 1.5 |
| −10.9 | 0508 | 9 | 17 | 100.0 | 100.0 | 1.0 | 10.0 | 3 | 20.0 | −3.0 | 1.5 |
| −10.9 | 0718 | 9 | 25 | 60.0 | 120.0 | 2.0 | 20.0 | 6 | 32.0 | −7.0 | 1.5 |
The units of A, BT1, C, D, and A–D are (cm H2O)2. The unit of B is (cm H2O)2 per h. The units of T1, T2, and T3 are h. The physical significance of the parameters A, B, … T3 is given in the legend of Fig. 3.
Table 1 shows the coefficients of the analytic functions that fit the mean trend of order k, [Mk(t)], of the MBP [Mk(t)MBP]. Table 2 shows the coefficients of the kth order mean of the energy of oscillations of blood pressure about the M6MBP, i.e., E6(t).
Fig. 5A shows the correlation of the coefficients of the analytical indicial functions of the MBP listed in Table 1 with the level of hypoxia −ΔpO2. It is seen that A, B, D, T1, and T2 are remarkably linear functions of −ΔpO2. T3 is mildly nonlinear. But the product BT1, which represents physically the magnitude of the peak of the indicial function above the base line for the case of decreased oxygen tension, is nonlinear. BT1 is the product of two linear factors; hence, it is a quadratic function of hypoxia. The coefficient C, which represents physically the asymptotic level of the blood pressure hypertension, is also nonlinear, and the nonlinearity is of the simple exponential type.
Fig. 5B shows the corresponding relations of the coefficients of the indicial functions of the kth order mean trend of E6(t), the energy of oscillations of the signal X6(t) (Eq. 8), oscillating about the MBP trend M6(t). The energy (Eq. 13) is proportional to the mean square of the systolic-diastolic pressure differences. It is seen that the indicial functions of the oscillations about the mean behave quite similarly to those of the MBP itself.
Now, we have completed what we set out to do. The clarity of the results offered by the IMF method was unique. We have found that when the oxygen tension in the breathing gas is decreased or increased as a step function of time, both the MBP and the Hilbert energy spectrum of the oscillations about the mean change with time and with the level of change of the partial pressure of O2. If the MBP or the mean energy of oscillation is denoted by y(t), and the hypoxia (the negative change of the oxygen partial pressure) is denoted by x(t), then our results may be expressed by the following equation:
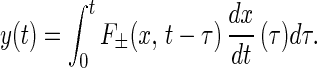 |
14 |
When x is a positive step function, dx/dτ is a positive delta function, then F+ applies and is called the positive indicial response function. When x is a negative step function, dx/dτ is a negative delta function, the F− applies and is called the negative indicial response function. We have determined the functions F+(x, t-τ) and F−(x, t-τ). As a theoretical extension of our experimental results, we would like to propose that Eq. 14 is valid for an arbitrary function x(t), with F+(x, t-τ) associated with an arbitrary history of increasing positive hypoxia, whenever dx/dτ is positive; and with F−(x, t-τ) associated with increasing oxygen tension, when dx/dτ is negative. When dx/dτ = 0, either F+ or F− persists, until it becomes nonvanishing.
If x = 0 all the time, i.e., if the oxygen tension in the breathing gas was maintained constant, we found the typical trend of the MBP was not a periodic function with a 24 h. period, as shown in Figs. 1–4 (Left). The fluctuations about the mean are fractal, e.g., the IMFs of 10-sec segments shown in figures 1C and 3 A and B of ref. 19 are fairly similar to those shown in Fig. 1 of this paper. The total number of modes are fewer for the short segments and more for the long segments (eight for the 10-sec segment but 16 for the 24-h segment).
CONCLUSION AND DISCUSSION
We conclude that the IMF method leads to clear and reasonable definition of the mean trends and oscillations about the mean trends of blood pressure. The energy of oscillations can be similarly resolved to mean trends and oscillations about the mean. These trends, when fitted with analytic functions, tell us that under step hypoxia or step recovery, the outcomes of rise and decay of the MBP and the energy of oscillations are quite similar. Most parameters of the indicial functions vary linearly with the degree of hypoxia, whereas a few parameters vary as quadratic function of hypoxia.
The successive IMFs are like successive subharmonic oscillations. The last IMF is nonoscillatory and is the basic mean trend. The subharmonic feature differs fundamentally from the Fourier analysis, which emphasizes higher harmonics.
In our laboratory, the study of blood pressure fluctuation caused by hypoxic gas breathing was motivated by research on the tissue remodeling of pulmonary blood vessels. We have collected tissue specimens of rat lungs subjected to periods of step changes of oxygen tension for various lengths of time, measured the changed morphology, diameter, opening angle, thicknesses of the intima-media, adventitia, and mechanical properties, made measurements of blood flow, and calculated the stresses (22). Like blood pressure, most of these variables are nonstationary oscillatory functions of time, especially the flow and the shear stress acting on the endothelium, and can be benefited by the IMF method.
Acknowledgments
This work was supported by National Heart, Lung, and Blood Institute Grant HL 43026; American Heart Association, California Affiliate, Postdoctoral Fellowship 96–95 (to W.H.); National Science Foundation Grant CM-9615897 and National Aeronautics and Space Administration Grant NAG 5–5149 (to Z.S.); and National Aeronautics and Space Administration Grant RTOP 622–47-11–20 (to N.E.H.).
ABBREVIATIONS
- IMF
intrinsic mode function
- MBP
mean blood pressure
References
- 1.Atkinson J E, Olson J W, Altiere R J, Gillespie M N. J Appl Physiol. 1987;62:1562–1568. doi: 10.1152/jappl.1987.62.4.1562. [DOI] [PubMed] [Google Scholar]
- 2.Meyrick B, Reid L. Lab Invest. 1980;42:603–615. [PubMed] [Google Scholar]
- 3.Meyrick B, Reid L. Am J Pathol. 1980;100:151–178. [PMC free article] [PubMed] [Google Scholar]
- 4.Reid L. Chest. 1986;89:279–288. doi: 10.1378/chest.89.2.279. [DOI] [PubMed] [Google Scholar]
- 5.Sobin S S, Tremer H M, Hardy J D, Chiodi H P. J Appl Physiol. 1983;55:1445–1455. doi: 10.1152/jappl.1983.55.5.1445. [DOI] [PubMed] [Google Scholar]
- 6.Davies P, Reid L. J Appl Physiol. 1991;71:1886–1891. doi: 10.1152/jappl.1991.71.5.1886. [DOI] [PubMed] [Google Scholar]
- 7.Fung Y C, Liu S Q. J Appl Physiol. 1991;70:2455–2470. doi: 10.1152/jappl.1991.70.6.2455. [DOI] [PubMed] [Google Scholar]
- 8.Fung Y C, Liu S Q. Am J Physiol. 1992;262:H544–H552. doi: 10.1152/ajpheart.1992.262.2.H544. [DOI] [PubMed] [Google Scholar]
- 9.Fung Y C, Liu S Q, Zhou J B. J Biomech Eng. 1993;115:453–459. doi: 10.1115/1.2895523. [DOI] [PubMed] [Google Scholar]
- 10.Dicarlo V S, Chen S J, Meng Q C, Durand J, Yano M, Chen Y F, Oparil S. Am J Physiol. 1995;269:L690–L697. doi: 10.1152/ajplung.1995.269.5.L690. [DOI] [PubMed] [Google Scholar]
- 11.Hung K S, McKenzie J C, Mattioli L, Klein R M, Meno C D, Poulose A K. Acta Anat. 1986;126:13–20. doi: 10.1159/000146180. [DOI] [PubMed] [Google Scholar]
- 12.Stenmark K R, Aldashev A A, Orton E C, Durmowicz A G, Badesch D B, Parks W C, Mecham R P, Voelkel N F, Reeves J T. Am J Physiol Suppl. 1991;261:97–104. doi: 10.1152/ajplung.1991.261.4.L97. [DOI] [PubMed] [Google Scholar]
- 13.Wohrley J D, Frid M G, Moiseeva E P, Orton E C, Belknap J K, Stenmark K R. J Clin Invest. 1995;96:273–281. doi: 10.1172/JCI118031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liu S Q. J Appl Physiol. 1996;81:2147–2155. doi: 10.1152/jappl.1996.81.5.2147. [DOI] [PubMed] [Google Scholar]
- 15.Liu S Q. J Appl Physiol. 1997;82:1677–1684. doi: 10.1152/jappl.1997.82.5.1677. [DOI] [PubMed] [Google Scholar]
- 16.Fung Y C. Ann Biomed Eng. 1991;19:237–249. doi: 10.1007/BF02584301. [DOI] [PubMed] [Google Scholar]
- 17.Wagner W W, Jr, Weir E K. The Pulmonary Circulation and Gas Exchange. Armonk, NY: Futura; 1994. [Google Scholar]
- 18.Huang N E, Shen Z, Long S R, Wu M L, Shih H H, Zheng Q, Yen N C, Tung C C, Liu H H. Proc R Soc London Ser A. 1998;454:903–995. [Google Scholar]
- 19.Huang W, Shen Z, Huang N E, Fung Y C. Proc Natl Acad Sci USA. 1998;95:4816–4821. doi: 10.1073/pnas.95.9.4816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Huang W, Shen Z, Huang N E, Fung Y C. Proc Natl Acad Sci USA. 1998;95:12766–12771. doi: 10.1073/pnas.95.22.12766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Titchmarsh E C. Introduction to the Theory of Fourier Integrals. Oxford: Oxford Univ. Press; 1948. [Google Scholar]
- 22.Fung Y C, Liu S Q. Proc Natl Acad Sci USA. 1995;92:2169–2173. doi: 10.1073/pnas.92.6.2169. [DOI] [PMC free article] [PubMed] [Google Scholar]