

Abstract
Human skin is repeatedly exposed to ultraviolet radiation (UVR) that influences the function and survival of many cell types and is regarded as the main causative factor in the induction of skin cancer. It has been traditionally believed that skin pigmentation is the most important photoprotective factor, since melanin, besides functioning as a broadband UV absorbent, has antioxidant and radical scavenging properties. Besides, many epidemiological studies have shown a lower incidence for skin cancer in individuals with darker skin compared to those with fair skin. Skin pigmentation is of great cultural and cosmetic importance, yet the role of melanin in photoprotection is still controversial. This article outlines the major acute and chronic effects of UV radiation on human skin, the properties of melanin, the regulation of pigmentation and its effect on skin cancer prevention.
Introduction
Human skin is repeatedly exposed to various DNA-damaging environmental influences and therefore requires numerous endogenous mechanisms to protect against, reduce and/or repair such damage. These mechanisms include increasing epidermal thickness, DNA repair mechanisms and apoptosis, antioxidant enzymes and, last but not least, skin pigmentation. Ultraviolet radiation (UVR) is the major environmental factor that influences the function and survival of many cell types and is regarded as the main causative factor in the induction of skin tumors like basal cell carcinoma (BCC), squamous cell carcinoma (SCC) and malignant melanoma. It has been traditionally believed that skin pigmentation is the most important photoprotective factor, since melanin, besides functioning as a broadband UV absorbent, has antioxidant and radical scavenging properties. Besides, many epidemiological studies have shown a lower incidence for skin cancer in individuals with darker skin compared to those with fair skin. However, there is growing evidence that the relationship between pigmentation and photoprotection is far more complex than assumed. It has been suggested that UVR-induced photodamage and its repair are signals that induce melanogenesis. To put it another way, epidermal pigmentation may represent the mammalian equivalent of a bacterial SOS repair mechanism.
Skin pigmentation is of great cultural and cosmetic importance, yet the role of melanin is still controversial. In light of the increasing incidence for UV-induced skin cancer and the progressive depletion of the ozone layer, which contrasts to public perception of a tan as being healthy, a better understanding of the role of melanin in preventing UV-induced DNA damage and malignant transformation of skin cells would be more than desirable.
Solar Ultraviolet Spectra and Effects on the Skin
The UVR that reaches the earth's surface consists mainly of long wavelength ultraviolet A (UVA) (320 – 400 nm) radiation but only a minority (estimated at 5%) of short wavelength ultraviolet B (UVB) (280 - 320 nm) (1). UVC (200 – 280 nm) does not reach the surface of the earth, as it is screened out by atmospheric oxygen as well absorbed by the ozone layer. However, the penetration of UVB is enhanced by the increasing depletion of the ozone layer leading to a higher risk for UV-induced carcinogenesis (2). It has been estimated that a 1% decrease in ozone increases melanoma mortality by 1%-2% (3). Solar simulated radiation (SSR) refers to radiation emitted by a device that comprises a spectrum and intensity similar to the sun's radiation (global radiation) consisting of ∼50% visible light, ∼40% infrared light and ∼9% UV light (∼0.4% UVB). It is important to note that terrestrial radiation varies with the solar altitude which depends on geographic location, season and time of day (4).
UVR is responsible for a wide variety of different acute and chronic effects on the skin. Acute responses of human skin to UVR include photodamage, erythema, mutation, immunosuppression, synthesis of vitamin D and tanning. Chronic UVR effects include photoaging and photocarcinogenesis, which is considered to be induced by mutation and immunosuppression (5, 6).
DNA photodamage
One of the most important acute effects of UVR is DNA photodamage. UVA and UVB show different properties regarding their biological effects on the skin. UVB radiation is more cytotoxic and mutagenic than UVA and, according to wavelength dependent studies, is 3-4 orders of magnitude more effective per unit physical dose (J/cm2) than UVA for DNA photodamage (7), erythema (7, 8), tanning (8) and skin cancer in mice (9). However, UVA, which in contrast to UVB is not filtered by window glass, is able to penetrate deeper into the skin and reach the dermis. It has been estimated that 50% of exposure to UVA occurs in the shade (10). While UVB is absorbed directly by DNA, and induces base structural DNA damage, UVA is mainly responsible for indirect DNA damage by the generation of reactive oxygen species (ROS) that include superoxide anion, hydrogen peroxide, and singlet oxygen (11) and result in single-strand breaks in DNA and in DNA-protein crosslinks (12, 13). UVR from 245 to 290 nm is absorbed maximally by DNA (14), thus implicating UVB as a primary mutagen (15). The most important photolesions that are induced by UVB with potentially mutagenic properties are cyclobutane pyrimidine dimers (CPD) and pyrimidine (6-4) pyrimidone photoproducts (64PP) that occur at a ratio that varies from 4:1 to 10:1 (16, 17). Animal studies confirm that CPD, and to a lesser extent 64PP, have roles in photocarcinogenesis (18). CPD and 64PP can lead to highly specific mutations, namely CC → TT double base substitutions and C→T substitutions at dipyrimidine sites that are known as UVR fingerprint mutations (19). It was reported that over 50% of BCC and SCC contain UVB fingerprint mutations (19, 20). Even suberythemal doses of UVB and SSR can immediately induce CPD and 64PP in human skin (7, 21, 22). However, it seems that pyrimidine dimerization is not limited to UVB radiation. It was reported that UVA, that is very weakly absorbed by DNA is able to induce CPDs (22, 23) and that UVA accounts for a much higher induction of T=T CPDs than does UVB (24). There is strong evidence that the underlying mechanism is based on a photosensitizing process that involves a still unidentified photoexcited chromophore (24, 25). The removal of 64PP after the application of a single dose of SSR is relatively fast in contrast to the repair of CPD, where many lesions can still be found after 24 hr (26, 27). There is also evidence that cytosine-containing lesions such as C=C and C=T, are repaired more rapidly than are those that contain thymidine only (T=T) (26, 28).
Photodamage to DNA can further result in highly characteristic gene mutations in various critical genes (29, 30), e.g. in p53 which is thought to be the first step in the induction of non-melanoma skin cancer (31).
Sunburn
The effectiveness of UV to induce erythema declines rapidly with longer wavelengths. To produce the same erythemal response, approximately 1000 times more UVA dose is needed compared with UVB (8). UVB-induced erythema occurs approximately 4 hr after exposure, peaks around 8 to 24 hr, and fades over a day or so; in fair-skinned and older individuals, UVB erythema may be persistent, sometimes lasting for weeks. The time courses for UVA-induced erythema and tanning are biphasic. Erythema is often evidenced immediately at the end of the irradiation period; it fades in several hr, followed by a delayed erythema starting at 6 hr and reaching its peak at 24 hr. Erythema is associated with a wide variety of changes at the cell and molecular levels, but especially with the appearance of apoptotic keratinocytes (sunburn cells) (32, 33). The action spectrum for UV-induced tanning and erythema are almost identical, but UVA is more efficient in inducing tanning whereas UVB is more efficient in inducing erythema. The observation that the action spectrum for erythema is very similar to that for CPD induction suggests that DNA damage is an important trigger for erythema (7).
UV and immunosuppression
Besides its deleterious photodamaging and erythema-inducing effects, UVR also has local and systemic immunosuppressive properties (34) and is capable of reducing the capability for surveillance against tumor or viral antigens (35). After UV exposure, the antigen-presenting Langerhans cells undergo numerous, functional, and morphologic changes, resulting in their depletion from the skin. UV-induced immunosuppression may have arisen to prevent autoimmune responses to inflammatory products resulting from UV-mediated damage (e.g. UV-damaged DNA). Most of the immunosuppressive effects of UV exposure have been ascribed to UVB. However, more recent studies indicated that UVA radiation is even more immunosuppressive than UVB (36).
There is considerable evidence from mouse studies that UV-induced immunosuppression might be a risk factor for the development of skin tumors (5). In an animal mouse model, skin tumors that developed in adult mice after long-term high dose UVB irradiation were rejected when transplanted to syngeneic healthy mice, but continued to grow when transplanted on mice that were pre-irradiated with UVB (37).
In humans, UVR is also able to suppress development of the contact hypersensitivity response, which is regarded as a model for photoimmunological events that are important in skin carcinogenesis (38, 39). Interestingly, the contact hypersensitivity response is more easily suppressed in fair skin types (types I/II) than in darker skin types (types III/IV) (40). Furthermore, there is evidence for skin type dependent differences in response to oxidative stress (41), which is thought to play a role in carcinogenesis (13).
UV, Vitamin D and folic acid
The epidermis is the site for the synthesis of vitamin D. During exposure to sunlight, 7-dehydrocholesterol in the epidermis is photolyzed by UVB to previtamin D3, which is then converted to vitamin D3 (cholecalciferol) by thermal isomerization. In the liver, vitamin D3 is hydroxylated to form 25-hydroxy-vitamin-D3, which in turn is transported to the kidney where it is converted into the biologically active form 1α,25-dihydroxyvitamin-D3 (calcitriol) (42). Skin pigmentation influences the effectiveness of vitamin D3 synthesis in the skin as melanin absorbs UVB photons and competes for them with 7-dehydrocholesterol (43). However, available data about the relationship between solar UVR, skin pigmentation and vitamin D status show contradictory results. On the one hand, it was reported that skin pigmentation greatly reduces the UVR-mediated synthesis of vitamin D3 as those with Black skin require at least a 6-fold greater UVR dose to increase circulating levels of vitamin D3 than do those with White skin (44). It was also reported that many African Americans who live in northern parts of the US suffer severe vitamin D deficiencies in spite of supplementing foods with vitamin D (45). In contrast, Matsuoka (46) found no difference in 1,25-(OH)2-vitamin D3 levels in the sera of different ethnic groups, although there was a significant association between skin color and vitamin D3 synthesis. This view was supported by Nelson et al. (47) who reported that there is no difference in the disposition for calcium deficiency in dark-skinned compared to fair-skinned individuals and in that report they further emphasized that African women have the same bone mass as Caucasian women. It is possible that a higher incidence of vitamin D deficiency results from other factors such as diet. Further, melanin absorbs only 50-75% of the UVR. Generally, it can be stated that in geographical areas of high UV-exposure there is selection for dark skin to prevent sunburn, skin cancer and photolysis of folic acid (48, 49) and to protect the sweat glands to prevent abnormal thermoregulation (50). The gradient between UVR level and constitutive pigmentation seems to represent a compromise between the deleterious effect of UVR and its beneficial effects regarding vitamin D synthesis (49).
UV and Pigmentation
The regulatory mechanisms that lead to pigmentation are complex and at present not completely understood. However, extensive data suggest that UV-induced DNA damage and/or its repair produce initiating signals that induce an increase in melanogenesis after UV irradiation (51, 52), although the chromophore(s) for melanogenesis have not been established with certainty.
Studies have shown that the action spectrum that leads to tanning in human skin is identical to the spectrum that induces erythema (8), which in turn is similar to that for the typical DNA photoproduct CPD (7). Treatment of UV-exposed melanocytes with the excision enzyme T4 endonuclease V, that is responsible for the repair of CPD, increased DNA repair as well as doubling the melanin content compared with cells treated with diluent alone or with heat-inactivated enzyme (53). Several studies have examined the effect of small DNA fragments such as thymidine dinucleotide (pTpT) which serve as a model for thymidine dimers. In vitro, a 7-fold increase in melanin content and a 2-fold increase in tyrosinase mRNA (the rate limiting enzyme for melanin synthesis) was observed after treatment with pTpT, while topical treatment of shaved guinea pig skin with pTpT induced a visible tan (54). Further, agents that induce single strand DNA breaks were also able to stimulate melanogenesis in vitro (55).
Melanin Properties
Skin color is mainly determined by the mix of carotenoids, oxy-/deoxy-hemoglobin and, most importantly, different types of melanin and also the way that melanin is packaged and distributed in melanosomes (56). The production of melanin takes place in specific ovoid organelles known as melanosomes which are produced in dendritic melanocytes that account for only 1% of epidermal cells. Each basal layer melanocyte is associated with about 36 keratinocytes and one Langerhans cell (epidermal melanin unit) (57). Melanin synthesized within melanosomes is transported via dendrites to adjacent keratinocytes, and accumulate within keratinocytes and melanocytes in the perinuclear area as supranuclear “caps” that are thought to shield DNA from UV rays (58). The composition of melanin (discussed in more detail in Chapters XX and XX (note to Editor – please replace ‘XX’ with relevant Chapter numbers in this Symposium-in-Print) is a complex of lighter red/yellow, alkali soluble sulfur-containing pheomelanin and darker brown/black insoluble eumelanin (59, 60). Differences in skin pigmentation do not result from differences in the number of melanocytes in the skin, as one might assume, but from differences in the melanogenic activity, the type of melanin produced in melanosomes and the size, number and packaging of melanosomes, with melanin content of melanosomes ranging from 17.9% to 72.3% (61, 62). The number of melanocytes in the skin is race-independent, but can vary at different body sites with densities between 2000 mm-2 in head or forearm skin to 1000 mm-2 elsewhere. Melanocyte density in the skin of the palms and soles is about 10-20% that in skin on the trunk (63). Red hair can contain high levels of pheomelanin, but generally, all types of skin contain a higher proportion of eu- than pheo-melanin. While total melanin content in the epidermis differs only by ∼2-fold in Asian and White skin, Black skin contains higher levels of melanin (∼3 to 6-fold higher, respectively), more eumelanin (64) and has larger and more melanosomes (65) than fair skin (Figure 1). However, no similar correlation has been observed for pheomelanin (59). Eumelanin is synthesized and deposited in elliptical melanosomes, whereas pheomelanin is synthesized in smaller round melanosomes. Melanosomes in Black skin have much longer axes than do melanosomes in White skin (800 vs. 400 nm). They exist as single entities and it has been hypothesized that they are transferred to the keratinocytes individually (thus absorbing light more efficiently), while melanosomes in light skin tend to form clusters and are packaged and transferred as complexes (66).
Figure 1. Melanin Content in Skin of Different Ethnicity.
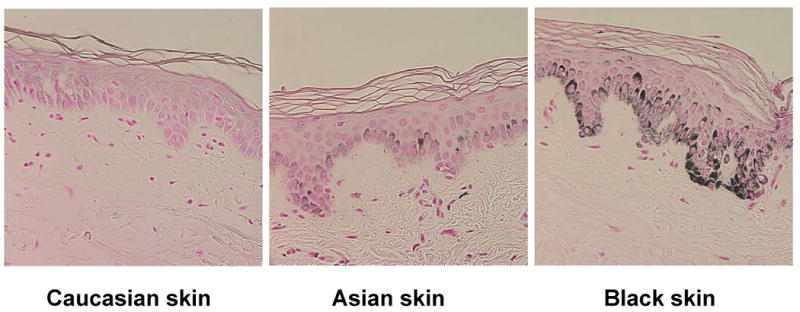
Histological detection of melanin content using Fontana-Masson staining. Melanin content in the basal layers of the epidermis is substantially higher in Black skin compared to Asian or White skin, although the number of melanocytes is virtually identical in skins of different ethnicity.
Photoprotective Role of Melanin
Epidemiological data strongly support the photoprotective role of melanin as there exists an inverse correlation between skin pigmentation and the incidence of sun-induced skin cancers (1) and subjects with White skin are approximately 70 times more likely to develop skin cancer than subjects with Black skin (67). The shielding effect of melanin, especially eumelanin, is achieved by its ability to serve as a physical barrier that scatters UVR, and as an absorbent filter that reduces the penetration of UV through the epidermis (68). The efficacy of melanin as a sunscreen was assumed to be about 1.5-2.0 sun protective factors (SPF); possibly as high as 4 SFP, implying that melanin absorbs 50% to 75% of UVR. An SPF of 2 means the doubling of protection of the skin against sunburn. Dark skin, which contains more eumelanin than fair skin is better protected against UV-induced damage, and eumelanin is thought to be superior to pheomelanin in its photoprotective properties. As discussed by Gloster and Neal (69) melanin in Black skin is twice as effective compared to White skin (70) in inhibiting UVB radiation from penetrating. While Black epidermis allows only 7.4% of UVB and 17.5% of UVA to penetrate, 24% UVB and 55% UVA passes through White skin (70). Further, melanosomes in dark skin are resistant to degradation by lysosomal enzymes, remain intact throughout the epidermal layers (65) and form supranuclear caps in keratinocytes and melanocytes which contribute considerably to photoprotection against UV-induced damage (71). In contrast, in lightly pigmented skin, melanosomes are degraded and only persist as “melanin dust” in the suprabasal layers. This reduction of melanosomes in the upper epidermis is considered to be an important factor in carcinogenesis, as it compromises the photoprotection of the skin. Other important properties of eumelanin are its functions as a free radical scavenger and superoxide dismutase that reduce ROS (72).
Deleterious Effects of Melanin
Melanin is generally considered to be the perfect protection against UV-induced photodamage. However, melanin can also have toxic properties, especially after exposure to UVR (73-75). In in vitro studies, melanin has been shown able to react with DNA and is known to act as a photosensitizer that produces ROS after UVA radiation (76) that can produce single strand DNA breaks in skin cells in vitro (77). In contrast to eumelanin, pheomelanin is especially prone to photodegradation (78, 79) and is thought to contribute to the damaging effects of UVR because it can generate hydrogen peroxide and superoxide anions (80-82) and might cause mutations in melanocytes or other cells (83). In addition, pheomelanin has been associated with higher rates of apoptotic cells after UVR (84). Pheomelanin also increases the release of histamine, which contributes to the sun-induced erythema and edema in fair-skinned individuals (85). In vivo studies of mice also suggest that melanin, especially pheomelanin, is a UVA and UVB sensitizer that causes cell death (86). At present, the clinical significance of deleterious effects of melanogenic intermediates in carcinogenesis is not known, but it is noteworthy that albinos with tyrosinase-deficient melanocytes that cannot produce melanin are more prone to nonmelanoma skin cancers, but have a lower incidence of malignant melanoma (87). Thus, it is possible that pheomelanin has a weak carcinogenic effect that can contribute to melanoma formation.
Constitutive Versus Facultative Pigmentation
Human skin has developed two main defense mechanisms to guard against the damaging effects of UV: epidermal thickening and the stimulation of melanin synthesis. With regard to photoprotection, the increased melanogenesis is more important. The ability to adjust melanin content of epidermal cells after UV exposure is based on the individual's melanogenic potential, which led to the concept of facultative and constitutive skin color (88). Constitutive skin color describes the genetically determined level of melanin in the skin that is not influenced by exogenous or endogenous factors. Recent studies indicate that the distribution of melanosomes within keratinocytes (89) as well as melanosome size (90) play roles in determining skin color. On the other hand, facultative pigmentation designates an induced level of increased epidermal melanin content as a result of environmental factors such as solar radiation or hormones.
Increased skin pigmentation in response to UV exposure proceeds in several distinct steps: the first step is a transient phenomenon, termed immediate pigment darkening (IPD), which occurs within min of UV exposure. It appears as a grayish coloration that gradually fades to a brown color over a period of min to days, depending on the UV dose and the individual's skin color. IPD is not based on the synthesis of new melanin, but rather results from the photooxidation of preexisting melanin and the redistribution of existing melanosomes from a perinuclear to a peripheral dendritic location (91). IPD shows a broad peak in the UVA region (92) and is only marginally photoprotective (93), hence its biological function remains unknown. IPD is followed by a prolonged second phase of the tanning reaction which varies among individuals, called persistent pigment darkening (PPD). PPD, which is tan to brown in color, is thought to result from the oxidation of melanin (similar to IPD), occurs within hr after UV exposure and persists at least 3-5 days (94, 95). PPD is elicited more strongly by UVA than by UVB and has been successfully used for testing sunscreen formulations with UVA protection factors (95) The last phase of skin tanning, the delayed tanning (DT) response, can be induced by UVB or UVA, is clearly distinct from PPD and becomes apparent 2-3 days after UV exposure (62). Its action spectrum is completely different from the spectrum of IPD, which suggests that they are completely distinct processes (8). DT results from the stimulation of melanin synthesis and involves increases in the number and activity of functional melanocytes, increased dendricity, increased synthesis and transfer as well as altered packaging of melanosomes. Corresponding to the tanning reaction, there is an increase in the activity of tyrosinase, the rate-limiting enzyme in the melanogenic pathway (96). UVB-induced DT is photoprotective (it is estimated to have a SPF of 3)- while DT induced by UVA is not considered to be photoprotective. DT is maximal from 10 days to 3-4 weeks, depending on the UV dose and the individual's skin color. It may take several weeks or months for the skin to return to its base constitutive color. UVA-induced DT is 2-3 orders of magnitude less efficient per unit dose than UVB and has an earlier onset, often directly after IPD. Furthermore, it has a different oxygen-dependent pathophysiology (52, 97) unlike that of UVB.
The pigmentary response of the skin to UV is determined to a large extent by constitutive pigmentation and is more pronounced with darker skin color. The classification of skin phototypes (type I to VI) has been based on the ability of individuals with different constitutive pigmentation to burn or tan in response to sun exposure (62). The concept of skin phototype was originally created to optimize UVR doses in phototherapy (98). Skin phototype I represents individuals with very fair skin who always burn and never tan when exposed to the sun. Skin phototype II individuals tan minimally with difficulty and burn easily, skin phototype III individuals tan moderately and uniformly and burn moderately while type IV individuals burn minimally and tan moderately and easily. Finally, skin phototype V represent individuals with very dark skin who rarely burn and tan profusely, while type VI individuals never burn and tan profusely upon sun exposure. Sensitivity to sunburn is routinely evaluated by minimal erythema dose (MED) determination. MED describes the lowest dose (J/cm2) of UVR that will cause erythema assessed at 24 hr after exposure. In general, tanning capacity correlates with MED. but – in contrast to MED measurement - the determination of skin phototype is based on personal experience and is therefore not a reliable index for UVB sensitivity of the skin.
Hormonal Regulation of Pigmentation
Tanning is determined directly by the response of melanocytes to UVR, but is also affected indirectly by a complex system of paracrine and autocrine factors such as hormones, cytokines and growth factors, whose synthesis in epidermal cells is influenced by UVR. In vitro experiments have examined the expression of a wide variety of factors synthesized in skin cells after UVR. In keratinocytes, UVR increased synthesis of the cytokines interleukin-1 (IL1) (99) and tumor necrosis factor α (TNFα) (100), while these factors decreased the proliferation and tyrosinase activity of human melanocytes (101), suggesting that they might be part of a negative feedback loop that down-regulates the pigmentary effect of UVR. The levels of prostaglandins D2, E2 and F2α have been found to increase in the skin after UVR (102). It has been shown that prostaglandins E2 and F2α increase the dendricity of melanocytes, an effect that is associated with increased skin pigmentation, as it facilitates the transfer of melanosomes to keratinocytes (103).
Endothelin-1 (ET1) is produced by keratinocytes, especially after UVR, and has been reported to stimulate human melanocyte proliferation synergistically with basic fibroblast growth factor [bFGF) and α-melanocyte stimulating hormone [αMSH] (104). ET1 reduces UV-induced apoptosis as well as promotes melanocyte survival (105, 106).
More than 120 genes have been shown to regulate pigmentation in mammals. The major determinant of pigment phenotype is the melanocortin 1 receptor (MC1R) (107). Its product, the MC1R protein, is a G-protein coupled receptor that is expressed on the surface of melanocytes, and which regulates melanin synthesis qualitatively and quantitatively. MC1R function is regulated by the physiological agonists αMSH and adrenocorticotropic hormone (ACTH) and by one antagonist agouti-signaling protein (ASP). Activation of the MC1R by binding an agonist stimulates the synthesis of eumelanin. Epidermal keratinocytes and melanocytes respond to UVR by increasing their expression of αMSH and ACTH, which up-regulates the expression of MC1R and consequently enhances the response of melanocytes to αMSH and ACTH. Those melanocortins, as well as ET1, are survival factors that rescue human melanocytes from apoptosis induced by UVR (105). Those 3 factors activate survival, DNA repair and antioxidant pathways in human melanocytes. Their pigmentary effects protect against the damaging effects of subsequent UVR exposures. Variants of MC1R that function weakly show a relative inability to tan (for example red-heads cannot tan). In mice with mutant MC1R there is no detectable tanning response; however, a rescue strategy of topically applied forskolin was able to bypass the mutant MC1R and induced a profound skin darkening in genetically fair-skinned mice (108).
Vitamin D3 (cholecalciferol) is synthesized in the skin upon sun exposure. There have been studies to examine whether vitamin D3 metabolites might mediate the melanogenic effects of UVR. It was shown that human melanocytes express specific receptors for vitamin D3 (109). Experiments addressing the correlation between vitamin D and pigmentation also showed contradictory results. In an in vitro study, melanocytes responded to 1,25 (OH)2 vitamin D3 with a decrease in tyrosinase activity (109). But contrary to those findings, Mansur et al. did not detect an effect of different vitamin D derivatives on human melanocytes in vitro (110). In contrast, topically applied 1,25 (OH)2 vitamin D3 increased the number of DOPA-positive melanocytes in mouse skin and augmented the melanogenic effect of UVR on murine skin (109).
Sex steroid hormones have long been recognized for their role in cutaneous pigmentation. Although no specific receptors for estrogens could be detected in melanocytes, dose-dependent increases in tyrosinase activity to β-estradiol have been reported and the response did not correlate with constitutive pigmentation (111, 112).
Melanin and UV Exposure
Epidemiological as well as experimental data suggest that melanin in the skin plays an important role in photoprotection, and not merely as a sunscreen. The relationship between melanogenesis and DNA photodamage was summarized in an earlier review by Agar and Young (113). In an early study, Kaidbey et al. (68) examined Black and White skin after irradiation with UVA and UVB with regard to phototoxicity and erythema. They found that five times less UV reaches the upper dermis of Black skin compared to White skin and assumed that was due to the increased melanin content, its more efficient distribution and the thickness of the stratum corneum. However, another study that used white-and-black guinea pigs to examine the relationship between the protective role of melanin and UV-induced DNA repair by measuring unscheduled DNA synthesis (UDS) in guinea pig skin after treatment with UV radiation (at different wavelengths) in vivo found no difference in UDS in pigmented and unpigmented skin at any UV dose. They concluded that epidermal melanin does not significantly protect DNA of basal cells against UV radiation (114). In a recent study, using sunburn cells as markers, a close correlation between constitutive skin pigmentation and UV sensitivity was observed. In brown or dark skin, DNA damage was restricted to upper layers, but occurred in all layers of lightly pigmented skin (115). Other studies concentrated on the relation of facultative pigmentation and photoprotection. Young et al. (116) exposed subjects with skin type II who were treated with or without sunscreen to SSR (0.7 MED) 10 times over a 2-week period followed by a 2 MED SSR challenge one week later. The results of measurements of unscheduled DNA synthesis, melanin content, distribution and skin thickness in biopsies taken at the end of the experiment led to the conclusion that increased melanin content alone without sunscreen protection was not sufficient to protect completely against DNA damage. In a follow-up study (117) that extended the original study, Young et al. found that pigmentation and skin thickening were induced in all skin types examined (I-IV). Combined with the use of sunscreen, the increased pigmentation and skin thickening improved protection against subsequent UV challenge. In another clinical study Sheehan et al. (118) reported that tanning offers a ‘modest’ photoprotection against erythema and concluded that induced tanning provides a higher protection against UV than does the thickening of the stratum corneum. De Winter and colleagues (119) examined whether repeated UV exposures (SSR 3 times a week for 3 weeks; the first dose was 0.5 MED, the second 1.0 MED and the following doses with an increment of 20% MED) lead to significant protection against subsequent UV (3 MED SSR) challenge. Their study concluded that repetitive UV exposure increased pigmentation and skin thickness, and decreased its sensitivity to erythema by 75%. They found less DNA damage (average reduction in CPD formation by about 60%) and concluded that the pigmentation induced was photoprotective to some extent. Photoprotection by melanin not only depends on its amount, but is also determined by its distribution in the skin. We completed a clinical study with more than 90 subjects in 7 distinct racial/ethnic backgrounds that were exposed to a single 1 MED dose of UVA/UVB. The analysis of biopsies taken immediately, at 1 d and 7 d after the irradiation showed that DNA damage was significantly greater in lighter, more UV-sensitive skin types and significantly lower in darker, more UV-resistant skin, as shown in Figure 2, although subjects with darker skin typically received at least 3 – 4 times the physical UV dose (17, 120, 121). In all skin types, DNA damage occurs to a greater extent in the upper layers of the epidermis, while the lower layers of the skin are protected as the melanin content of the skin increases (122). Interestingly, the results showed that UVR not only induces less DNA damage in darker skin than in lighter skin, but also the rate of apoptotic cell formation - even at the same physical UV dose - is dramatically higher in darker skin (122). This indicates that besides decreased UVR-induced DNA damage, darker skin is also more efficient at removing damaged cells. This combination results in a greatly reduced risk of carcinogenesis.
Figure 2. CPD in Skin of Different Ethnicity.
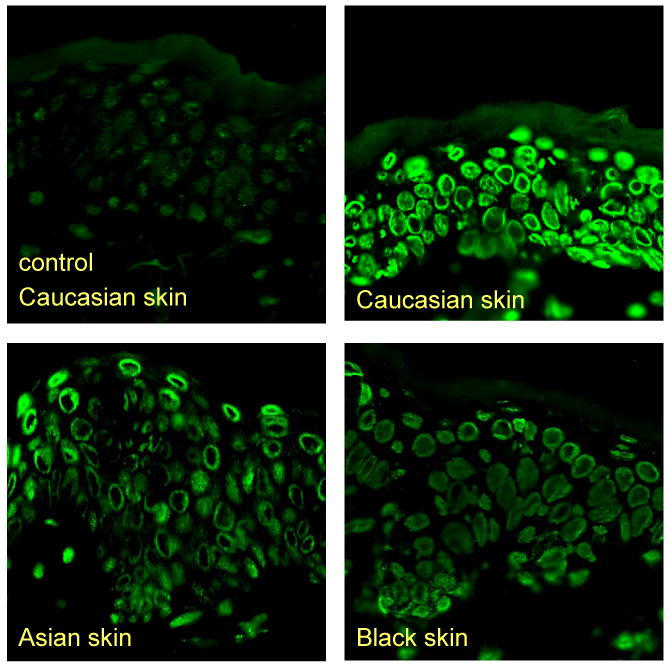
Immunohistochemical detection of CPDs in skin of different ethnicities immediately after UVR. The fluorescence detection technique uses the binding of FITC-labeled antibodies that are reactive with DNA photoproducts. A considerably higher amount of CPDs is detected in White skin compared to Asian or Black skin.
Our irradiation studies in skin of different color and racial/ethnic origin demonstrated that, although a visible tan developed after a single 1 MED UV irradiation, melanin content did not significantly change within 1 week (17, 120). Even after a series of UV exposures over a period of 3 weeks and despite a significant increase in visible pigmentation, there are less than 2-fold increases in melanin content in White and Asian skin (123, 124). A possible explanation for this observation might be the fact that after UVR melanin migrates towards the surface of the skin and is thereby more visible and also more effective in shielding the lower layers of the skin from UV irradiation. The observations made in these studies allow the conclusions that DNA damage is greatest immediately after UV exposure and is gradually repaired thereafter and that subjects with darker skin type incur less DNA damage than subjects with lighter skin. However, even the darkest skin type incurred significant DNA damage at levels less than or equal to 1 MED, underscoring the fact that there is no such thing as totally UV-resistant human skin.
In another irradiation study we examined the effect of 3 different schedules of repeated UV exposure (cumulative dose of schedule A = 1900 J/m2, of schedule B = 2900 J/m2 and of schedule C = 4200 J/m2). Exposures to all 3 schedules resulted in effective increases in skin pigmentation and significantly increased melanocyte density. Interestingly, the level of CPD measured was significantly lower after schedule C compared to schedules A and B, implying that pigmentation induced by repeated UV exposure protects against subsequent CPD formation. However, increased cumulative UV doses raised the number of p53-positive cells, potentially associated with a higher risk of UV induced malignant transformation (124).
Melanin and Skin Cancer
Since the 1960s, incidences in BCC, SCC and melanoma among predominantly white populations have increased at a rate of 5% to 8% annually (125-127). In contrast, the incidence of skin cancer has remained relatively constant in Blacks.
Non-melanoma skin cancers such as SCC in subjects with White skin most commonly occur on areas of skin exposed to the sun, and occurs in sun-protected sites in subjects with Black skin (67, 128). When the protective effect of melanin is calculated using MED measurements, protection for even the darkest-skinned individuals is no more than 10- to 15-fold that seen in the absence of melanin. But in terms of skin cancer risk, the protection is 500-1000 (66, 68) indicating that highly pigmented skin is profoundly protected from carcinogenesis. Even less deeply pigmented Asians have a very low skin cancer rate (129). One possible explanation for the high cancer protection afforded by darker skin pigmentation might involve mechanisms of increased risk associated with blonde/red pigments. MC1R variants have been shown to confer an increased risk of melanoma and non-melanoma skin cancers, independently of skin pigment (including red hair phenotype) (130, 131).
Recently, it was shown that the transcription factor and tumor-suppressor protein p53 promotes cutaneous pigmentation after UVR by activation of proopiomelanocortin (POMC) (a precursor of αMSH) expression, while absence of p53 inhibits the tanning response (132). POMC/αMSH is of considerable importance for skin carcinogenesis, as mutations in the POMC gene lead to a red-hair phenotype (comparable to that of MC1R alleles) (133).
UVA induces 40 times more DNA single strand breaks in human melanocytes from dark skin than in melanocytes from light skin (134). Given that UVA damage is largely thought to occur via oxidative mechanisms this may account for a positive role for UVA in melanoma induction (84).
Conclusions
To minimize the deleterious effects of UVR, public education on photoprotective measures should be continued. Although there are a wide variety of agents with photoprotective properties, which range from antioxidants to plant extracts to DNA repair enzymes, a better understanding of melanin, its photoprotective properties and contributions of melanocytes to cancer would be desirable. This should permit new approaches to safely modulate pigmentation in the absence of sun to increase pigmentation for cosmetic reasons as well as to prevent skin cancer.
Acknowledgments
This research was supported by the Intramural Research Program of the NIH, National Cancer Institute.
Abbreviations used
- 64PP
6,4-photoproducts
- ACTH
adrenocorticotropic hormone
- ASP
agouti signaling protein
- BCC
basal cell carcinoma
- bFGF
basic fibroblast growth factor
- CPD
cyclobutane pyrimidine dimer
- DT
delayed tanning
- ET1
endothelin 1
- IL1
interleukin 1
- IPD
immediate pigment darkening
- MC1R
melanocortin 1 receptor
- MED
minimal erythema dose
- αMSH
α-melanocyte stimulating hormone
- PPD
persistent pigment darkening
- POMC
proopiomelanocortin
- ROS
reactive oxygen species
- SCC
squamous cell carcinoma
- SPF
sun protective factor
- SSR
solar simulated radiation
- TNFα
tumor necrosis factor α
- UVR
ultraviolet radiation
References
- 1.Gilchrest BA, Eller MS, Geller AC, Yaar M. The pathogenesis of melanoma induced by ultraviolet radiation. N Engl J Med. 1999;340:1341–1348. doi: 10.1056/NEJM199904293401707. [DOI] [PubMed] [Google Scholar]
- 2.De Fabo EC. Arctic stratospheric ozone depletion and increased UVB radiation: potential impacts to human health. Int J Circumpolar Health. 2005;64:509–522. doi: 10.3402/ijch.v64i5.18032. [DOI] [PubMed] [Google Scholar]
- 3.Kripke ML. Impact of ozone depletion on skin cancers. J Dermatol Surg Oncol. 1988;14:853–857. doi: 10.1111/j.1524-4725.1988.tb03589.x. [DOI] [PubMed] [Google Scholar]
- 4.Holzle E, Honigsmann H. UV-radiation--sources, wavelength, environment. J Dtsch Dermatol Ges. 2005;3 2:S3–10. doi: 10.1111/j.1610-0387.2005.04392.x. [DOI] [PubMed] [Google Scholar]
- 5.Ullrich SE. Photoimmune suppression and photocarcinogenesis. Front Biosci. 2002;7:d684–703. doi: 10.2741/A804. [DOI] [PubMed] [Google Scholar]
- 6.Ziegler A, Jonason AS, Leffell DJ, Simon JA, Sharma HW, Kimmelman J, Remington L, Jacks T, Brash DE. Sunburn and p53 in the onset of skin cancer. Nature. 1994;372:773–776. doi: 10.1038/372773a0. [DOI] [PubMed] [Google Scholar]
- 7.Young AR, Chadwick CA, Harrison GI, Nikaido O, Ramsden J, Potten CS. The similarity of action spectra for thymine dimers in human epidermis and erythema suggests that DNA is the chromophore for erythema. J Invest Dermatol. 1998;111:982–988. doi: 10.1046/j.1523-1747.1998.00436.x. [DOI] [PubMed] [Google Scholar]
- 8.Parrish JA, Jaenicke KF, Anderson RR. Erythema and melanogenesis action spectra of normal human skin. Photochem Photobiol. 1982;36:187–191. doi: 10.1111/j.1751-1097.1982.tb04362.x. [DOI] [PubMed] [Google Scholar]
- 9.de Gruijl FR. Action spectrum for photocarcinogenesis. Recent Results Cancer Res. 1995;139:21–30. doi: 10.1007/978-3-642-78771-3_2. [DOI] [PubMed] [Google Scholar]
- 10.Schaefer H, Moyal D, Fourtanier A. Recent advances in sun protection. Semin Cutan Med Surg. 1998;17:266–275. doi: 10.1016/s1085-5629(98)80023-0. [DOI] [PubMed] [Google Scholar]
- 11.Fisher GJ, Kang S, Varani J, Bata-Csorgo Z, Wan Y, Datta S, Voorhees JJ. Mechanisms of photoaging and chronological skin aging. Arch Dermatol. 2002;138:1462–1470. doi: 10.1001/archderm.138.11.1462. [DOI] [PubMed] [Google Scholar]
- 12.Kvam E, Tyrrell RM. Induction of oxidative DNA base damage in human skin cells by UV and near visible radiation. Carcinogenesis. 1997;18:2379–2384. doi: 10.1093/carcin/18.12.2379. [DOI] [PubMed] [Google Scholar]
- 13.Sander CS, Chang H, Hamm F, Elsner P, Thiele JJ. Role of oxidative stress and the antioxidant network in cutaneous carcinogenesis. Int J Dermatol. 2004;43:326–335. doi: 10.1111/j.1365-4632.2004.02222.x. [DOI] [PubMed] [Google Scholar]
- 14.Tornaletti S, Pfeifer GP. UV damage and repair mechanisms in mammalian cells. Bioessays. 1996;18:221–228. doi: 10.1002/bies.950180309. [DOI] [PubMed] [Google Scholar]
- 15.Linge C. Relevance of in vitro melanocytic cell studies to the understanding of melanoma. Cancer Surv. 1996;26:71–87. [PubMed] [Google Scholar]
- 16.Sage E. Distribution and repair of photolesions in DNA: genetic consequences and the role of sequence context. Photochem Photobiol. 1993;57:163–174. doi: 10.1111/j.1751-1097.1993.tb02273.x. [DOI] [PubMed] [Google Scholar]
- 17.Tadokoro T, Kobayashi N, Zmudzka BZ, Ito S, Wakamatsu K, Yamaguchi Y, Korossy KS, Miller SA, Beer JZ, Hearing VJ. UV-induced DNA damage and melanin content in human skin differing in racial/ethnic origin. Faseb J. 2003;17:1177–1179. doi: 10.1096/fj.02-0865fje. [DOI] [PubMed] [Google Scholar]
- 18.Vink AA, Roza L. Biological consequences of cyclobutane pyrimidine dimers. J Photochem Photobiol B. 2001;65:101–104. doi: 10.1016/s1011-1344(01)00245-7. [DOI] [PubMed] [Google Scholar]
- 19.Brash DE, Rudolph JA, Simon JA, Lin A, McKenna GJ, Baden HP, Halperin AJ, Ponten J. A role for sunlight in skin cancer: UV-induced p53 mutations in squamous cell carcinoma. Proc Natl Acad Sci U S A. 1991;88:10124–10128. doi: 10.1073/pnas.88.22.10124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ziegler A, Leffell DJ, Kunala S, Sharma HW, Gailani M, Simon JA, Halperin AJ, Baden HP, Shapiro PE, Bale AE, et al. Mutation hotspots due to sunlight in the p53 gene of nonmelanoma skin cancers. Proc Natl Acad Sci U S A. 1993;90:4216–4220. doi: 10.1073/pnas.90.9.4216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chadwick CA, Potten CS, Nikaido O, Matsunaga T, Proby C, Young AR. The detection of cyclobutane thymine dimers, (6-4) photolesions and the Dewar photoisomers in sections of UV-irradiated human skin using specific antibodies, and the demonstration of depth penetration effects. J Photochem Photobiol B. 1995;28:163–170. doi: 10.1016/1011-1344(94)07096-7. [DOI] [PubMed] [Google Scholar]
- 22.Young AR, Potten CS, Nikaido O, Parsons PG, Boenders J, Ramsden JM, Chadwick CA. Human melanocytes and keratinocytes exposed to UVB or UVA in vivo show comparable levels of thymine dimers. J Invest Dermatol. 1998;111:936–940. doi: 10.1046/j.1523-1747.1998.00435.x. [DOI] [PubMed] [Google Scholar]
- 23.Applegate LA, Scaletta C, Panizzon R, Niggli H, Frenk E. In vivo induction of pyrimidine dimers in human skin by UVA radiation: initiation of cell damage and/or intercellular communication? Int J Mol Med. 1999;3:467–472. doi: 10.3892/ijmm.3.5.467. [DOI] [PubMed] [Google Scholar]
- 24.Douki T, Reynaud-Angelin A, Cadet J, Sage E. Bipyrimidine photoproducts rather than oxidative lesions are the main type of DNA damage involved in the genotoxic effect of solar UVA radiation. Biochemistry. 2003;42:9221–9226. doi: 10.1021/bi034593c. [DOI] [PubMed] [Google Scholar]
- 25.Cadet J, Sage E, Douki T. Ultraviolet radiation-mediated damage to cellular DNA. Mutat Res. 2005;571:3–17. doi: 10.1016/j.mrfmmm.2004.09.012. [DOI] [PubMed] [Google Scholar]
- 26.Bykov VJ, Sheehan JM, Hemminki K, Young AR. In situ repair of cyclobutane pyrimidine dimers and 6-4 photoproducts in human skin exposed to solar simulating radiation. J Invest Dermatol. 1999;112:326–331. doi: 10.1046/j.1523-1747.1999.00523.x. [DOI] [PubMed] [Google Scholar]
- 27.Young AR, Chadwick CA, Harrison GI, Hawk JL, Nikaido O, Potten CS. The in situ repair kinetics of epidermal thymine dimers and 6-4 photoproducts in human skin types I and II. J Invest Dermatol. 1996;106:1307–1313. doi: 10.1111/1523-1747.ep12349031. [DOI] [PubMed] [Google Scholar]
- 28.Xu G, Snellman E, Bykov VJ, Jansen CT, Hemminki K. Effect of age on the formation and repair of UV photoproducts in human skin in situ. Mutat Res. 2000;459:195–202. doi: 10.1016/s0921-8777(99)00069-5. [DOI] [PubMed] [Google Scholar]
- 29.Mitchell DL, Jen J, Cleaver JE. Relative induction of cyclobutane dimers and cytosine photohydrates in DNA irradiated in vitro and in vivo with ultraviolet-C and ultraviolet-B light. Photochem Photobiol. 1991;54:741–746. doi: 10.1111/j.1751-1097.1991.tb02084.x. [DOI] [PubMed] [Google Scholar]
- 30.Mitchell DL, Nairn RS. The biology of the (6-4) photoproduct. Photochem Photobiol. 1989;49:805–819. doi: 10.1111/j.1751-1097.1989.tb05578.x. [DOI] [PubMed] [Google Scholar]
- 31.Brash DE, Ziegler A, Jonason AS, Simon JA, Kunala S, Leffell DJ. Sunlight and sunburn in human skin cancer: p53, apoptosis, and tumor promotion. J Investig Dermatol Symp Proc. 1996;1:136–142. [PubMed] [Google Scholar]
- 32.Murphy G, Young AR, Wulf HC, Kulms D, Schwarz T. The molecular determinants of sunburn cell formation. Exp Dermatol. 2001;10:155–160. doi: 10.1034/j.1600-0625.2001.010003155.x. [DOI] [PubMed] [Google Scholar]
- 33.Sheehan JM, Cragg N, Chadwick CA, Potten CS, Young AR. Repeated ultraviolet exposure affords the same protection against DNA photodamage and erythema in human skin types II and IV but is associated with faster DNA repair in skin type IV. J Invest Dermatol. 2002;118:825–829. doi: 10.1046/j.1523-1747.2002.01681.x. [DOI] [PubMed] [Google Scholar]
- 34.Aubin F. Mechanisms involved in ultraviolet light-induced immunosuppression. Eur J Dermatol. 2003;13:515–523. [PubMed] [Google Scholar]
- 35.Kripke ML. Immunology and photocarcinogenesis. New light on an old problem. J Am Acad Dermatol. 1986;14:149–155. doi: 10.1016/s0190-9622(86)70017-0. [DOI] [PubMed] [Google Scholar]
- 36.Nghiem DX, Kazimi N, Clydesdale G, Ananthaswamy HN, Kripke ML, Ullrich SE. Ultraviolet a radiation suppresses an established immune response: implications for sunscreen design. J Invest Dermatol. 2001;117:1193–1199. doi: 10.1046/j.0022-202x.2001.01503.x. [DOI] [PubMed] [Google Scholar]
- 37.Fisher MS, Kripke ML. Systemic alteration induced in mice by ultraviolet light irradiation and its relationship to ultraviolet carcinogenesis. Proc Natl Acad Sci U S A. 1977;74:1688–1692. doi: 10.1073/pnas.74.4.1688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Elmets CA, Bergstresser PR, Tigelaar RE, Wood PJ, Streilein JW. Analysis of the mechanism of unresponsiveness produced by haptens painted on skin exposed to low dose ultraviolet radiation. J Exp Med. 1983;158:781–794. doi: 10.1084/jem.158.3.781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yoshikawa T, Rae V, Bruins-Slot W, Van den Berg JW, Taylor JR, Streilein JW. Susceptibility to effects of UVB radiation on induction of contact hypersensitivity as a risk factor for skin cancer in humans. J Invest Dermatol. 1990;95:530–536. doi: 10.1111/1523-1747.ep12504877. [DOI] [PubMed] [Google Scholar]
- 40.Kelly DA, Young AR, McGregor JM, Seed PT, Potten CS, Walker SL. Sensitivity to sunburn is associated with susceptibility to ultraviolet radiation-induced suppression of cutaneous cell-mediated immunity. J Exp Med. 2000;191:561–566. doi: 10.1084/jem.191.3.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kerb R, Brockmoller J, Reum T, Roots I. Deficiency of glutathione S-transferases T1 and M1 as heritable factors of increased cutaneous UV sensitivity. J Invest Dermatol. 1997;108:229–232. doi: 10.1111/1523-1747.ep12335337. [DOI] [PubMed] [Google Scholar]
- 42.Holick MF, MacLaughlin JA, Clark MB, Holick SA, Potts JT, Jr, Anderson RR, Blank IH, Parrish JA, Elias P. Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science. 1980;210:203–205. doi: 10.1126/science.6251551. [DOI] [PubMed] [Google Scholar]
- 43.Chen TC, Chimeh F, Lu Z, Mathieu J, Person KS, Zhang A, Kohn N, Martinello S, Berkowitz R, Holick MF. Factors that influence the cutaneous synthesis and dietary sources of vitamin D. Arch Biochem Biophys. 2007;460:213–217. doi: 10.1016/j.abb.2006.12.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Clemens TL, Adams JS, Henderson SL, Holick MF. Increased skin pigment reduces the capacity of skin to synthesise vitamin D3. Lancet. 1982;1:74–76. doi: 10.1016/s0140-6736(82)90214-8. [DOI] [PubMed] [Google Scholar]
- 45.Holick MF, Jenkins M. The UV advantage. ibooks; New York, NY: 2003. [Google Scholar]
- 46.Matsuoka LY, Wortsman J, Haddad JG, Kolm P, Hollis BW. Racial pigmentation and the cutaneous synthesis of vitamin D. Arch Dermatol. 1991;127:536–538. [PubMed] [Google Scholar]
- 47.Nelson DA, Kleerekoper M, Peterson E, Parfitt AM. Skin color and body size as risk factors for osteoporosis. Osteoporos Int. 1993;3:18–23. doi: 10.1007/BF01623172. [DOI] [PubMed] [Google Scholar]
- 48.Aoki K. Sexual selection as a cause of human skin colour variation: Darwin's hypothesis revisited. Ann Hum Biol. 2002;29:589–608. doi: 10.1080/0301446021000019144. [DOI] [PubMed] [Google Scholar]
- 49.Jablonski NG, Chaplin G. The evolution of human skin coloration. J Hum Evol. 2000;39:57–106. doi: 10.1006/jhev.2000.0403. [DOI] [PubMed] [Google Scholar]
- 50.Pandolf KB, Gange RW, Latzka WA, Blank IH, Young AJ, Sawka MN. Human thermoregulatory responses during cold water immersion after artificially induced sunburn. Am J Physiol. 1992;262:R617–623. doi: 10.1152/ajpregu.1992.262.4.R617. [DOI] [PubMed] [Google Scholar]
- 51.Gilchrest BA, Eller MS. DNA photodamage stimulates melanogenesis and other photoprotective responses. J Investig Dermatol Symp Proc. 1999;4:35–40. doi: 10.1038/sj.jidsp.5640178. [DOI] [PubMed] [Google Scholar]
- 52.Eller MS, Gilchrest BA. Tanning as part of the eukaryotic SOS response. Pigment Cell Res. 2000;13 8:94–97. doi: 10.1034/j.1600-0749.13.s8.17.x. [DOI] [PubMed] [Google Scholar]
- 53.Gilchrest BA, Zhai S, Eller MS, Yarosh DB, Yaar M. Treatment of human melanocytes and S91 melanoma cells with the DNA repair enzyme T4 endonuclease V enhances melanogenesis after ultraviolet irradiation. J Invest Dermatol. 1993;101:666–672. doi: 10.1111/1523-1747.ep12371673. [DOI] [PubMed] [Google Scholar]
- 54.Eller MS, Yaar M, Gilchrest BA. DNA damage and melanogenesis. Nature. 1994;372:413–414. doi: 10.1038/372413a0. [DOI] [PubMed] [Google Scholar]
- 55.Eller MS, Ostrom K, Gilchrest BA. DNA damage enhances melanogenesis. Proc Natl Acad Sci U S A. 1996;93:1087–1092. doi: 10.1073/pnas.93.3.1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Stamatas GN, Zmudzka BZ, Kollias N, Beer JZ. Non-invasive measurements of skin pigmentation in situ. Pigment Cell Res. 2004;17:618–626. doi: 10.1111/j.1600-0749.2004.00204.x. [DOI] [PubMed] [Google Scholar]
- 57.Fitzpatrick TB, Breathnach AS. the Epidermal Melanin Unit System. Dermatol Wochenschr. 1963;147:481–489. [PubMed] [Google Scholar]
- 58.Montagna W, Carlisle K. The architecture of black and white facial skin. J Am Acad Dermatol. 1991;24:929–937. doi: 10.1016/0190-9622(91)70148-u. [DOI] [PubMed] [Google Scholar]
- 59.Thody AJ, Higgins EM, Wakamatsu K, Ito S, Burchill SA, Marks JM. Pheomelanin as well as eumelanin is present in human epidermis. J Invest Dermatol. 1991;97:340–344. doi: 10.1111/1523-1747.ep12480680. [DOI] [PubMed] [Google Scholar]
- 60.Zanetti R, Prota G, Napolitano A, Martinez C, Sancho-Garnier H, Osterlind A, Sacerdote C, Rosso S. Development of an integrated method of skin phenotype measurement using the melanins. Melanoma Res. 2001;11:551–557. doi: 10.1097/00008390-200112000-00001. [DOI] [PubMed] [Google Scholar]
- 61.Duchon J. Chemical composition of melanosomes. Dermatol Monatsschr. 1970;156:371. [PubMed] [Google Scholar]
- 62.Pathak MA, Jimbow K, Fitzpatrick T. Photobiology of pigment cells. In: Seiji M, editor. Phenotypic expression in pigment cells. University of Tokyo Press; Tokyo: 1980. pp. 655–670. [Google Scholar]
- 63.Yamaguchi Y, Itami S, Watabe H, Yasumoto K, Abdel-Malek ZA, Kubo T, Rouzaud F, Tanemura A, Yoshikawa K, Hearing VJ. Mesenchymal-epithelial interactions in the skin: increased expression of dickkopf1 by palmoplantar fibroblasts inhibits melanocyte growth and differentiation. J Cell Biol. 2004;165:275–285. doi: 10.1083/jcb.200311122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ito S, Wakamatsu K. Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: a comparative review. Pigment Cell Res. 2003;16:523–531. doi: 10.1034/j.1600-0749.2003.00072.x. [DOI] [PubMed] [Google Scholar]
- 65.Szabo G, Gerald AB, Pathak MA, Fitzpatrick TB. Racial differences in the fate of melanosomes in human epidermis. Nature. 1969;222:1081–1082. doi: 10.1038/2221081a0. [DOI] [PubMed] [Google Scholar]
- 66.Kollias N, Sayre RM, Zeise L, Chedekel MR. Photoprotection by melanin. J Photochem Photobiol B. 1991;9:135–160. doi: 10.1016/1011-1344(91)80147-a. [DOI] [PubMed] [Google Scholar]
- 67.Halder RM, Bang KM. Skin cancer in blacks in the United States. Dermatol Clin. 1988;6:397–405. [PubMed] [Google Scholar]
- 68.Kaidbey KH, Agin PP, Sayre RM, Kligman AM. Photoprotection by melanin--a comparison of black and Caucasian skin. J Am Acad Dermatol. 1979;1:249–260. doi: 10.1016/s0190-9622(79)70018-1. [DOI] [PubMed] [Google Scholar]
- 69.Gloster HM, Jr, Neal K. Skin cancer in skin of color. J Am Acad Dermatol. 2006;55:741–760. 761–744. doi: 10.1016/j.jaad.2005.08.063. quiz. [DOI] [PubMed] [Google Scholar]
- 70.Halder RM, Bridgeman-Shah S. Skin cancer in African Americans. Cancer. 1995;75:667–673. doi: 10.1002/1097-0142(19950115)75:2+<667::aid-cncr2820751409>3.0.co;2-i. [DOI] [PubMed] [Google Scholar]
- 71.Kobayashi N, Nakagawa A, Muramatsu T, Yamashina Y, Shirai T, Hashimoto MW, Ishigaki Y, Ohnishi T, Mori T. Supranuclear melanin caps reduce ultraviolet induced DNA photoproducts in human epidermis. J Invest Dermatol. 1998;110:806–810. doi: 10.1046/j.1523-1747.1998.00178.x. [DOI] [PubMed] [Google Scholar]
- 72.Bustamante J, Bredeston L, Malanga G, Mordoh J. Role of melanin as a scavenger of active oxygen species. Pigment Cell Res. 1993;6:348–353. doi: 10.1111/j.1600-0749.1993.tb00612.x. [DOI] [PubMed] [Google Scholar]
- 73.Kipp C, Young AR. The soluble eumelanin precursor 5,6-dihydroxyindole-2-carboxylic acid enhances oxidative damage in human keratinocyte DNA after UVA irradiation. Photochem Photobiol. 1999;70:191–198. [PubMed] [Google Scholar]
- 74.Kvam E, Dahle J. Melanin synthesis may sensitize melanocytes to oxidative DNA damage by ultraviolet A radiation and protect melanocytes from direct DNA damage by ultraviolet B radiation. Pigment Cell Res. 2004;17:549–550. doi: 10.1111/j.1600-0749.2004.00168.x. [DOI] [PubMed] [Google Scholar]
- 75.Kvam E, Tyrrell RM. The role of melanin in the induction of oxidative DNA base damage by ultraviolet A irradiation of DNA or melanoma cells. J Invest Dermatol. 1999;113:209–213. doi: 10.1046/j.1523-1747.1999.00653.x. [DOI] [PubMed] [Google Scholar]
- 76.Korytowski W, Pilas B, Sarna T, Kalyanaraman B. Photoinduced generation of hydrogen peroxide and hydroxyl radicals in melanins. Photochem Photobiol. 1987;45:185–190. doi: 10.1111/j.1751-1097.1987.tb05362.x. [DOI] [PubMed] [Google Scholar]
- 77.Marrot L, Belaidi JP, Meunier JR, Perez P, Agapakis-Causse C. The human melanocyte as a particular target for UVA radiation and an endpoint for photoprotection assessment. Photochem Photobiol. 1999;69:686–693. [PubMed] [Google Scholar]
- 78.Chedekel MR. Photochemistry and photobiology of epidermal melanins. Photochem Photobiol. 1982;35:881–885. doi: 10.1111/j.1751-1097.1982.tb02663.x. [DOI] [PubMed] [Google Scholar]
- 79.Chedekel MR, Post PW, Deibel RM, Kalus M. Photodestruction of phaeomelanin. Photochem Photobiol. 1977;26:651–653. doi: 10.1111/j.1751-1097.1977.tb07546.x. [DOI] [PubMed] [Google Scholar]
- 80.Chedekel MR, Smith SK, Post PW, Pokora A, Vessell DL. Photodestruction of pheomelanin: role of oxygen. Proc Natl Acad Sci U S A. 1978;75:5395–5399. doi: 10.1073/pnas.75.11.5395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Felix CC, Hyde JS, Sarna T, Sealy RC. Melanin photoreactions in aerated media: electron spin resonance evidence for production of superoxide and hydrogen peroxide. Biochem Biophys Res Commun. 1978;84:335–341. doi: 10.1016/0006-291x(78)90175-4. [DOI] [PubMed] [Google Scholar]
- 82.Prota G. The chemistry of melanins and melanogenesis. Fortschr Chem Org Naturst. 1995;64:93–148. doi: 10.1007/978-3-7091-9337-2_2. [DOI] [PubMed] [Google Scholar]
- 83.Harsanyi ZP, Post PW, Brinkmann JP, Chedekel MR, Deibel RM. Mutagenicity of melanin from human red hair. Experientia. 1980;36:291–292. doi: 10.1007/BF01952282. [DOI] [PubMed] [Google Scholar]
- 84.Hill HZ, Hill GJ. UVA, pheomelanin and the carcinogenesis of melanoma. Pigment Cell Res. 2000;13 8:140–144. doi: 10.1034/j.1600-0749.13.s8.25.x. [DOI] [PubMed] [Google Scholar]
- 85.Cesarini JP. Photo-induced events in the human melanocytic system: photoaggression and photoprotection. Pigment Cell Res. 1988;1:223–233. doi: 10.1111/j.1600-0749.1988.tb00420.x. [DOI] [PubMed] [Google Scholar]
- 86.Takeuchi S, Zhang W, Wakamatsu K, Ito S, Hearing VJ, Kraemer KH, Brash DE. Melanin acts as a potent UVB photosensitizer to cause an atypical mode of cell death in murine skin. Proc Natl Acad Sci U S A. 2004;101:15076–15081. doi: 10.1073/pnas.0403994101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Streutker CJ, McCready D, Jimbow K, From L. Malignant melanoma in a patient with oculocutaneous albinism. J Cutan Med Surg. 2000;4:149–152. doi: 10.1177/120347540000400307. [DOI] [PubMed] [Google Scholar]
- 88.Pathak MA. Activation of the melanocyte system by ultraviolet radiation and cell transformation. Ann N Y Acad Sci. 1985;453:328–339. doi: 10.1111/j.1749-6632.1985.tb11821.x. [DOI] [PubMed] [Google Scholar]
- 89.Thong HY, Jee SH, Sun CC, Boissy RE. The patterns of melanosome distribution in keratinocytes of human skin as one determining factor of skin colour. Br J Dermatol. 2003;149:498–505. doi: 10.1046/j.1365-2133.2003.05473.x. [DOI] [PubMed] [Google Scholar]
- 90.Alaluf S, Barrett K, Blount M, Carter N. Ethnic variation in tyrosinase and TYRP1 expression in photoexposed and photoprotected human skin. Pigment Cell Res. 2003;16:35–42. doi: 10.1034/j.1600-0749.2003.00005.x. [DOI] [PubMed] [Google Scholar]
- 91.Routaboul C, Denis A, Vinche A. Immediate pigment darkening: description, kinetic and biological function. Eur J Dermatol. 1999;9:95–99. [PubMed] [Google Scholar]
- 92.Irwin C, Barnes A, Veres D, Kaidbey K. An ultraviolet radiation action spectrum for immediate pigment darkening. Photochem Photobiol. 1993;57:504–507. doi: 10.1111/j.1751-1097.1993.tb02326.x. [DOI] [PubMed] [Google Scholar]
- 93.McGregor JM, Hawk JLM. Acute effects of ultraviolet radiation on the skin. In: Freedberg IM, Eisen AZ, Wolff K, Austen KF, Goldsmith LA, Katz SI, Fitzpatrick TB, editors. Dermatology in general medicine. McGraw-Hill; New York: 1999. pp. 1555–1561. [Google Scholar]
- 94.Moyal D, Chardon A, Kollias N. Determination of UVA protection factors using the persistent pigment darkening (PPD) as the end point. (Part 1). Calibration of the method. Photodermatol Photoimmunol Photomed. 2000;16:245–249. doi: 10.1034/j.1600-0781.2000.160602.x. [DOI] [PubMed] [Google Scholar]
- 95.Moyal D, Wichrowski K, Tricaud C. In vivo persistent pigment darkening method: a demonstration of the reproducibility of the UVA protection factors results at several testing laboratories. Photodermatol Photoimmunol Photomed. 2006;22:124–128. doi: 10.1111/j.1600-0781.2006.00223.x. [DOI] [PubMed] [Google Scholar]
- 96.Friedmann PS, Gilchrest BA. Ultraviolet radiation directly induces pigment production by cultured human melanocytes. J Cell Physiol. 1987;133:88–94. doi: 10.1002/jcp.1041330111. [DOI] [PubMed] [Google Scholar]
- 97.Lavker RM, Kaidbey KH. Redistribution of melanosomal complexes within keratinocytes following UV-A irradiation: a possible mechanism for cutaneous darkening in man. Arch Dermatol Res. 1982;272:215–228. doi: 10.1007/BF00509049. [DOI] [PubMed] [Google Scholar]
- 98.Fitzpatrick TB. The validity and practicality of sun-reactive skin types I through VI. Arch Dermatol. 1988;124:869–871. doi: 10.1001/archderm.124.6.869. [DOI] [PubMed] [Google Scholar]
- 99.Oxholm A, Oxholm P, Staberg B, Bendtzen K. Immunohistological detection of interleukin I-like molecules and tumour necrosis factor in human epidermis before and after UVB-irradiation in vivo. Br J Dermatol. 1988;118:369–376. doi: 10.1111/j.1365-2133.1988.tb02430.x. [DOI] [PubMed] [Google Scholar]
- 100.Kock A, Schwarz T, Kirnbauer R, Urbanski A, Perry P, Ansel JC, Luger TA. Human keratinocytes are a source for tumor necrosis factor alpha: evidence for synthesis and release upon stimulation with endotoxin or ultraviolet light. J Exp Med. 1990;172:1609–1614. doi: 10.1084/jem.172.6.1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Abdel-Malek Z, Swope V, Collins C, Boissy R, Zhao H, Nordlund J. Contribution of melanogenic proteins to the heterogeneous pigmentation of human melanocytes. J Cell Sci. 1993;106(Pt 4):1323–1331. doi: 10.1242/jcs.106.4.1323. [DOI] [PubMed] [Google Scholar]
- 102.Black AK, Fincham N, Greaves MW, Hensby CN. Time course changes in levels of arachidonic acid and prostaglandins D2, E2, F2 alpha in human skin following ultraviolet B irradiation. Br J Clin Pharmacol. 1980;10:453–457. doi: 10.1111/j.1365-2125.1980.tb01788.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Scott G, Leopardi S, Printup S, Malhi N, Seiberg M, Lapoint R. Proteinase-activated receptor-2 stimulates prostaglandin production in keratinocytes: analysis of prostaglandin receptors on human melanocytes and effects of PGE2 and PGF2alpha on melanocyte dendricity. J Invest Dermatol. 2004;122:1214–1224. doi: 10.1111/j.0022-202X.2004.22516.x. [DOI] [PubMed] [Google Scholar]
- 104.Swope VB, Medrano EE, Smalara D, Abdel-Malek ZA. Long-term proliferation of human melanocytes is supported by the physiologic mitogens alpha-melanotropin, endothelin-1, and basic fibroblast growth factor. Exp Cell Res. 1995;217:453–459. doi: 10.1006/excr.1995.1109. [DOI] [PubMed] [Google Scholar]
- 105.Kadekaro AL, Kavanagh R, Kanto H, Terzieva S, Hauser J, Kobayashi N, Schwemberger S, Cornelius J, Babcock G, Shertzer HG, Scott G, Abdel-Malek ZA. alpha-Melanocortin and endothelin-1 activate antiapoptotic pathways and reduce DNA damage in human melanocytes. Cancer Res. 2005;65:4292–4299. doi: 10.1158/0008-5472.CAN-04-4535. [DOI] [PubMed] [Google Scholar]
- 106.Kadekaro AL, Kavanagh RJ, Wakamatsu K, Ito S, Pipitone MA, Abdel-Malek ZA. Cutaneous photobiology. The melanocyte vs. the sun: who will win the final round? Pigment Cell Res. 2003;16:434–447. doi: 10.1034/j.1600-0749.2003.00088.x. [DOI] [PubMed] [Google Scholar]
- 107.Rees JL. Genetics of hair and skin color. Annu Rev Genet. 2003;37:67–90. doi: 10.1146/annurev.genet.37.110801.143233. [DOI] [PubMed] [Google Scholar]
- 108.D'Orazio JA, Nobuhisa T, Cui R, Arya M, Spry M, Wakamatsu K, Igras V, Kunisada T, Granter SR, Nishimura EK, Ito S, Fisher DE. Topical drug rescue strategy and skin protection based on the role of Mc1r in UV-induced tanning. Nature. 2006;443:340–344. doi: 10.1038/nature05098. [DOI] [PubMed] [Google Scholar]
- 109.Abdel-Malek ZA, Ross R, Trinkle L, Swope V, Pike JW, Nordlund JJ. Hormonal effects of vitamin D3 on epidermal melanocytes. J Cell Physiol. 1988;136:273–280. doi: 10.1002/jcp.1041360209. [DOI] [PubMed] [Google Scholar]
- 110.Mansur CP, Gordon PR, Ray S, Holick MF, Gilchrest BA. Vitamin D, its precursors, and metabolites do not affect melanization of cultured human melanocytes. J Invest Dermatol. 1988;91:16–21. doi: 10.1111/1523-1747.ep12463282. [DOI] [PubMed] [Google Scholar]
- 111.Ranson M, Posen S, Mason RS. Human melanocytes as a target tissue for hormones: in vitro studies with 1 alpha-25, dihydroxyvitamin D3, alpha-melanocyte stimulating hormone, and beta-estradiol. J Invest Dermatol. 1988;91:593–598. doi: 10.1111/1523-1747.ep12477126. [DOI] [PubMed] [Google Scholar]
- 112.McLeod SD, Ranson M, Mason RS. Effects of estrogens on human melanocytes in vitro. J Steroid Biochem Mol Biol. 1994;49:9–14. doi: 10.1016/0960-0760(94)90295-x. [DOI] [PubMed] [Google Scholar]
- 113.Agar N, Young AR. Melanogenesis: a photoprotective response to DNA damage? Mutatt Res. 2005;571:121–132. doi: 10.1016/j.mrfmmm.2004.11.016. [DOI] [PubMed] [Google Scholar]
- 114.Ishikawa T, Kodama K, Matsumoto J, Takayama S. Photoprotective role of epidermal melanin granules against ultraviolet damage and DNA repair in guinea pig skin. Cancer Res. 1984;44:5195–5199. [PubMed] [Google Scholar]
- 115.Del Bino S, Sok J, Bessac E, Bernerd F. Relationship between skin response to ultraviolet exposure and skin color type. Pigment Cell Res. 2006;19:606–614. doi: 10.1111/j.1600-0749.2006.00338.x. [DOI] [PubMed] [Google Scholar]
- 116.Young AR, Potten CS, Chadwick CA, Murphy GM, Cohen AJ. Inhibition of UV radiation-induced DNA damage by a 5-methoxypsoralen tan in human skin. Pigment Cell Res. 1988;1:350–354. doi: 10.1111/j.1600-0749.1988.tb00131.x. [DOI] [PubMed] [Google Scholar]
- 117.Young AR, Potten CS, Chadwick CA, Murphy GM, Hawk JL, Cohen AJ. Photoprotection and 5-MOP photochemoprotection from UVR-induced DNA damage in humans: the role of skin type. J Invest Dermatol. 1991;97:942–948. doi: 10.1111/1523-1747.ep12491807. [DOI] [PubMed] [Google Scholar]
- 118.Sheehan JM, Potten CS, Young AR. Tanning in human skin types II and III offers modest photoprotection against erythema. Photochem Photobiol. 1998;68:588–592. [PubMed] [Google Scholar]
- 119.de Winter S, Vink AA, Roza L, Pavel S. Solar-simulated skin adaptation and its effect on subsequent UV-induced epidermal DNA damage. J Invest Dermatol. 2001;117:678–682. doi: 10.1046/j.0022-202x.2001.01478.x. [DOI] [PubMed] [Google Scholar]
- 120.Tadokoro T, Yamaguchi Y, Batzer J, Coelho SG, Zmudzka BZ, Miller SA, Wolber R, Beer JZ, Hearing VJ. Mechanisms of skin tanning in different racial/ethnic groups in response to ultraviolet radiation. J Invest Dermatol. 2005;124:1326–1332. doi: 10.1111/j.0022-202X.2005.23760.x. [DOI] [PubMed] [Google Scholar]
- 121.Zmudzka BZ, Hearing VJ, Beer JZ. Photobiologic role of melanin distribution in the epidermis. J Photochem Photobiol B. 2006;84:231. doi: 10.1016/j.jphotobiol.2006.05.008. [DOI] [PubMed] [Google Scholar]
- 122.Yamaguchi Y, Takahashi K, Zmudzka BZ, Kornhauser A, Miller SA, Tadokoro T, Berens W, Beer JZ, Hearing VJ. Human skin responses to UV radiation: pigment in the upper epidermis protects against DNA damage in the lower epidermis and facilitates apoptosis. Faseb J. 2006;20:1486–1488. doi: 10.1096/fj.06-5725fje. [DOI] [PubMed] [Google Scholar]
- 123.Hennessy A, Oh C, Diffey B, Wakamatsu K, Ito S, Rees J. Eumelanin and pheomelanin concentrations in human epidermis before and after UVB irradiation. Pigment Cell Res. 2005;18:220–223. doi: 10.1111/j.1600-0749.2005.00233.x. [DOI] [PubMed] [Google Scholar]
- 124.Yamaguchi Y, Coelho SG, Zmudzka BZ, Takahashi K, Beer JZ, Hearing VJ, Miller SA. Cyclobutane pyrimidine dimer formation and p53 production in human skin after repeated UV exposures. J Invest Dermatol (revised and resubmitted) 2007 doi: 10.1111/j.1600-0625.2008.00722.x. [DOI] [PubMed] [Google Scholar]
- 125.Armstrong BK, Kricker A. Skin cancer. Dermatol Clin. 1995;13:583–594. [PubMed] [Google Scholar]
- 126.Gloster HM, Jr, Brodland DG. The epidemiology of skin cancer. Dermatol Surg. 1996;22:217–226. doi: 10.1111/j.1524-4725.1996.tb00312.x. [DOI] [PubMed] [Google Scholar]
- 127.Hall HI, Miller DR, Rogers JD, Bewerse B. Update on the incidence and mortality from melanoma in the United States. J Am Acad Dermatol. 1999;40:35–42. doi: 10.1016/s0190-9622(99)70562-1. [DOI] [PubMed] [Google Scholar]
- 128.Gray DT, Suman VJ, Su WP, Clay RP, Harmsen WS, Roenigk RK. Trends in the population-based incidence of squamous cell carcinoma of the skin first diagnosed between 1984 and 1992. Arch Dermatol. 1997;133:735–740. [PubMed] [Google Scholar]
- 129.Armstrong BK, Kricker A. The epidemiology of UV induced skin cancer. J Photochem Photobiol B. 2001;63:8–18. doi: 10.1016/s1011-1344(01)00198-1. [DOI] [PubMed] [Google Scholar]
- 130.Kennedy C, Huurne Jter, Berkhout M, Gruis N, Bastiaens M, Bergman W, Willemze R, Bavinck JN. Melanocortin 1 receptor (MC1R) gene variants are associated with an increased risk for cutaneous melanoma which is largely independent of skin type and hair color. J Invest Dermatol. 2001;117:294–300. doi: 10.1046/j.0022-202x.2001.01421.x. [DOI] [PubMed] [Google Scholar]
- 131.Scott MC, Wakamatsu K, Ito S, Kadekaro AL, Kobayashi N, Groden J, Kavanagh R, Takakuwa T, Virador V, Hearing VJ, Abdel-Malek ZA. Human melanocortin 1 receptor variants, receptor function and melanocyte response to UV radiation. J Cell Sci. 2002;115:2349–2355. doi: 10.1242/jcs.115.11.2349. [DOI] [PubMed] [Google Scholar]
- 132.Cui R, Widlund HR, Feige E, Lin JY, Wilensky DL, Igras VE, D'Orazio J, Fung CY, Schanbacher CF, Granter SR, Fisher DE. Central role of p53 in the suntan response and pathologic hyperpigmentation. Cell. 2007;128:853–864. doi: 10.1016/j.cell.2006.12.045. [DOI] [PubMed] [Google Scholar]
- 133.Krude H, Biebermann H, Luck W, Horn R, Brabant G, Gruters A. Severe early-onset obesity, adrenal insufficiency and red hair pigmentation caused by POMC mutations in humans. Nat Genet. 1998;19:155–157. doi: 10.1038/509. [DOI] [PubMed] [Google Scholar]
- 134.Wenczl E, Van der Schans GP, Roza L, Kolb RM, Timmerman AJ, Smit NP, Pavel S, Schothorst AA. (Pheo)melanin photosensitizes UVA-induced DNA damage in cultured human melanocytes. J Invest Dermatol. 1998;111:678–682. doi: 10.1046/j.1523-1747.1998.00357.x. [DOI] [PubMed] [Google Scholar]