

Abstract
Activating transcription factor 6 (ATF6) is located within the region of linkage to type 2 diabetes on chromosome 1q21-q23 and is a key activator of the endoplasmic reticulum stress response. We evaluated 78 single nucleotide polymorphisms (SNPs) spanning >213 kb in 95 people, from which we selected 64 SNPs for evaluation in 191 Caucasian case subjects from Utah and between 165 and 188 control subjects. Six SNPs showed nominal associations with type 2 diabetes (P = 0.001-0.04), including the nonsynonymous SNP rs1058405 (M67V) in exon 3 and rs11579627 in the 3′ flanking region. Only rs1159627 remained significant on permutation testing. The associations were not replicated in 353 African-American case subjects and 182 control subjects, nor were ATF6 SNPs associated with altered insulin secretion or insulin sensitivity in nondiabetic Caucasian individuals. No association with type 2 diabetes was found in a subset of 44 SNPs in Caucasian (n = 2,099), Pima Indian (n = 293), and Chinese (n = 287) samples. Allelic expression imbalance was found in transformed lymphocyte cDNA for 3′ untranslated region variants, thus suggesting cis-acting regulatory variants. ATF6 does not appear to play a major role in type 2 diabetes, but further work is required to identify the cause of the allelic expression imbalance.
The endoplasmic reticulum is a membranous labyrinthine network in eukaryotic cells that is the site for the synthesis, folding, assembly, and posttranslational modification of proteins. The endoplasmic reticulum includes a highly conserved system of proteins that facilitates protein folding and processing, protects cells from the toxic effects of accumulating unfolded proteins (endoplasmic reticulum stress), and, when these measures fail, initiates apoptosis. This system, known as the unfolded protein response, is activated by accumulating unfolded proteins (1-3). The endoplasmic reticulum stress and unfolded protein response pathways are particularly important in secretory cells, including pancreatic β-cells, hepatocytes, and adipocytes (2). In the presence of endoplasmic reticulum stress, the transcription factor activating factor 6 (ATF6) serves as a key proximal sensor and is transported to the Golgi complex, where it is cleaved by proteases to yield an active cytoplasmic domain that functions to upregulate chaperone proteins.
Considerable data have implicated endoplasmic reticulum stress in diabetes pathogenesis. In Akita mice, aberrant proinsulin processing induces endoplasmic reticulum stress and results in β-cell apoptosis (4), and mutations in other proximal sensors impair the endoplasmic reticulum stress response and lead to β-cell apoptosis and diabetes in experimental models and humans. Endoplasmic reticulum stress involvement in liver and adipose are activated in obese mice (5), and improved protein folding alleviated insulin resistance and improved glucose tolerance (6).
ATF6 includes 16 exons and spans 193 kb on chromosome 1q21-23, a region of well-replicated linkage to type 2 diabetes in eight different populations (7,8). Thus, ATF6 is a strong positional and functional candidate gene for type 2 diabetes. Furthermore, Thameem et al. (9) recently reported that the coding variant PRO145ALA (P145A) was nominally significantly associated with type 2 diabetes (P = 0.05). Additionally, we recently described an association of variants in the gene DUSP12, just upstream of ATF6, with type 2 diabetes in Caucasian and African-American subjects (10). To test the hypothesis that sequence variants in or near ATF6 contribute to type 2 diabetes susceptibility and to defects in insulin secretion and insulin resistance, we typed 64 single nucleotide polymorphisms (SNPs) spanning the ATF6 gene in a Northern European Caucasian population. We expanded the study by examining a subset of 44 SNPs in seven additional populations (U.K., French, and Amish Caucasians; Hong Kong and Shanghai Chinese; Pima Indians; and Arkansas African Americans) included in the International Type 2 Diabetes 1q Consortium. Nonsynonymous SNPs and SNPs showing the best evidence for an association with type 2 diabetes were tested for an association with insulin sensitivity (Si) and insulin secretion and for an association with type 2 diabetes in an Arkansas African-American population. Finally, we sought evidence that variation in or near ATF6 acting in a cis fashion might alter the expression ratio between alleles.
RESEARCH DESIGN AND METHODS
The primary study populations are summarized in Table 1. The primary study population comprised 191 unrelated Caucasian case subjects and 188 unrelated Caucasian control subjects ascertained from Utah and Arkansas for Northern European ancestry, as described previously (11,12). Case subjects were diabetic on treatment for type 2 diabetes and had a diabetic first-degree relative; control subjects had normal glucose tolerance tests and no first-degree relative with type 2 diabetes. A subset of 95 members of the control population was used to determine linkage disequilibrium and to select tag SNPs (see below). The same case subjects and a subset of the control subjects (165 control individuals) were genotyped as part of the International Type 2 Diabetes 1q Consortium (13).
TABLE 1.
Summary of primary study populations
| Population | Study description | Sex (M/F) | BMI (kg/m2) | Age (years) |
|---|---|---|---|---|
| Caucasian case/control | Case/control | Control 73/115, case 134/57 | 29.3 (19.8-43.2) | 56.9 ± 14.1 |
| African-American case/control | Case/control | Control 90/92, case 189/164 | 31.2 (20.0-48.8) | 50.0 ± 13.9 |
| Utah Caucasian metabolic | Family-based metabolic study | 50/72 | 27.5 (18.3-41.3) | 39.3 ± 10.5 |
| Arkansas Caucasian metabolic | Population-based metabolic study | 73/135 | 30.8 (18.2-41.9) | 36.4 ± 9.0 |
Data are arithmetic means for normal variables and geometric means for skewed variables (BMI), means (95% CI) transferred to the linear scale from the ln-transformation, or means ± SD.
Confirmatory association studies were conducted in seven additional populations representing four ethnic groups included in the International Type 2 Diabetes 1q Consortium (13): 424 African-American individuals from Arkansas (245 case subjects and 179 control subjects), 662 samples from the Amish Family Diabetes Study (308 case subjects with diabetes or impaired glucose tolerance and 354 control subjects), 547 samples from the French diabetes study (259 case subjects and 288 control subjects), 890 samples from the U.K. (443 and 447, respectively), 293 samples from the Pima Indian study (147 and 146), 128 samples from the Hong Kong Chinese study (64 and 64), and 159 samples from the Shanghai Chinese study (79 and 80). This resource is described in detail elsewhere (13). Additional African-American samples were tested in confirmatory genotyping studies conducted in Arkansas for a total population of 182 control subjects with normal fasting and/or postchallenge glucose levels and no family history of type 2 diabetes and 353 type 2 diabetic subjects who also had a diabetic first-degree relative and diabetes diagnosis before age 65 years. The Consortium Pima Indian samples overlap with those published previously for analysis of ATF6 (9).
Physiological studies were conducted in 121 members of Utah families ascertained for two diabetic siblings; each individual had undergone a tolbutamide-modified frequently sampled intravenous glucose tolerance test (FSIGT) (14). A second population of 209 unrelated Caucasian individuals from Arkansas underwent either a tolbutamide-modified (n = 100) or an insulin-modified (0.04 units/kg, n = 109) FSIGT (10). Subjects ascertained in Utah provided written informed consent under a protocol approved by the University of Utah institutional review board. Subjects studied in Arkansas provided written informed consent under protocols approved by the University of Arkansas for Medical Sciences institutional review board. Other Consortium samples were collected under approved procedures for local institutional review boards.
SNP genotyping
We evaluated 78 SNPs selected from public NCBI (National Center for Biotechnology Information) databases (15) and spanning from 8 kb upstream to 205 kb downstream of the ATF6 ATG start site (158,459,659-158,672,877 bp; NCBI Build 35) (supplemental Table 3S, which can be found in an online appendix [available at http://dx.doi.org/10.2337/db06-1305]). SNPs were selected from three sources: 1) SNPs typed as part of an initial 10-kb map across the region in the Utah sample; 2) known exonic variants (L35L, M67V, P90P, S103S, P145A, and S157P) in the ATF6 gene; and 3) 44 SNPs that fell within this region that were typed as part of a dense linkage disequilibrium map by the International Type 2 Diabetes 1q Consortium (13). We selected additional SNPs from HapMap (16) to fill in gaps identified based on initial HapMap linkage disequilibrium measures. These 78 SNPs were then typed in at least 95 Utah Caucasians to select a subset of SNPs with a minor allele frequency >5% and that fully captured the HapMap Phase 2 SNPs (r2 >0.95). Coding SNPs and SNPs already typed in the full Utah case-control set were forced in. Additional SNPs from the 44 Consortium SNPs that were required to capture the common variants were typed in the remaining control samples. The 44 Consortium SNPs were typed using 1536-plex Illumina Golden Gate assays (Illumina, San Diego, CA) (13). All other SNPs in the Utah and Arkansas samples were typed using either pyrosequencing on a PSQ-96 (Biotage, Uppsala, Sweden) or oligonucleotide ligation assays. For SNPs initially typed by the International Type 2 Diabetes 1q Consortium but expanded to additional control subjects, typing was accomplished by pyrosequencing but with at least 20 samples overlapping with the Illumina Golden Gate assay. Additionally, full typing of seven SNPs and partial typing of two additional SNPs showed excellent correspondence between pyrosequencing and Illumina Golden Gate assays.
SNPs were selected for full typing using the Tagger program in Haploview version 3 (17). We typed 39 SNPs in 191 case and 188 control subjects; an additional 25 SNPs typed by the International Type 2 Diabetes 1q Consortium but not required to capture the common Caucasian variants were typed in 191 case and 165 control subjects and included in all analyses. Nondiabetic Caucasian subjects who had undergone detailed metabolic studies were typed for nonsynonymous exonic SNPs or SNPs shown to be strongly associated in the Caucasian case-control cohort. African Americans were typed for the 44 Consortium SNPs, as well as exonic SNPs and SNPs showing an association in Caucasians, but no attempt was made to fully capture the common African-American variation based on HapMap Yoruban samples.
Allelic expression imbalance
Total RNA was isolated from Epstein-Barr virus-transformed lymphocytes, and allele-specific expression of two 3′ untranslated region SNPs was quantified from heterozygous individuals as described elsewhere (11). Briefly, total RNA was converted to cDNA using random primers. Both cDNA and genomic DNA were typed using the same assay by pyrosequencing, using a PSQ-96 machine (Biotage) with a universal biotinylated primer. Peak heights for the two alleles were quantified in both cDNA and genomic DNA using Allele Quantification software (Biotage), which was designed for quantification of pooled genotypes and which provides a ratio (percentage of total peak summing to 100%) for each allele. Allelic expression imbalance was considered to be present if the cDNA ratio fell outside the 95% CI determined from the DNA ratio in heterozygous individuals, in which the expected ratio is 1 (50% for each allele).
Statistical methods
The primary analyses compared genotypes using the Armitage Trend Test, which tests the null hypothesis: p2 + 1/2(p1) = q2 + 1/2(q1), where p1 and q1 are heterozygous case and control subjects, respectively, and p2 and q2 are rare homozygous case and control subjects, respectively. The test, which is distributed as χ2 with 1 degree of freedom, retains the power of an allelic association but remains valid in the presence of deviations from Hardy-Weinberg equilibrium. We considered an uncorrected two-sided P < 0.05 to be nominally significant. Secondary, exploratory analyses of genotypic association were conducted under additive, dominant, and recessive models using χ2 or Fisher’s exact tests. Tests of association and Hardy-Weinberg equilibrium were conducted using the DeFinetti program (18). Pairwise linkage disequilibrium measures were estimated from the genotype data using the EM (expectation maximization) algorithm implemented in HaploView version 3.2 or 3.32 (17). Haplotype case-control association was tested within each haplotype block using HaploView version 3.2, which tests the frequency of each haplotype in case versus control subjects; corrected P values are subsequently estimated by permutation tests (10,000 replicates). Haplotype blocks were established using HaploView version 3.32 with the solid spline of linkage disequilibrium definition.
Si and acute insulin response to glucose (AIRg) were calculated from FSIGT data using the MinMod (19) or MinMod Millennium programs (20). Genotype effects on insulin secretion and Si were tested using general linear models with sex and genotype as factors and BMI and age as covariates. For the Utah study, glucose tolerance status and family membership were included as additional factors, whereas for unrelated individuals from Arkansas, protocol type (tolbutamide or insulin) was included as an additional factor.
For analysis of Consortium data, between-group differences in allele frequency were evaluated on a population-specific basis using standard contingency table methods and exact P values calculated using Stata SE version 8 (Stata, College Station, TX). Single-point data from the case-control samples were combined using the Mantel-Haenszel fixed-effects method (Stata SE version 8), and combined odds ratios (ORs) were generated under dominant models for each allele. Studies in the Amish population accounted for family structure.
RESULTS
From 78 SNPs spanning 213.2 kb (-8,150 to 205,069 bp relative to the ATG start) (Fig. 1), we selected 64 SNPs for full evaluation in the Utah Northern European Caucasian case-control sample (denoted Utah Caucasian in the tables); nonsynonymous variants were forced into inclusion. The 64 typed SNPs fell into five linkage disequilibrium blocks of sizes 1, 61, 90, 40, and 4 kb (Fig. 2 and supplemental Figs. 3S and 4S). We found a nominal association with 6 of 64 SNPs (uncorrected P values from allelic association of 0.001-0.03) (Table 2 and Fig. 1), including a nonsynonymous SNP in exon 3 (rs1058405, M67V, P = 0.011) and a noncoding SNP in the 3′ flanking region (rs11579627, P = 0.001). Two other nonsynonymous variants in exon 5, rs2070150 (P145A) and rs1135983 (S157P), were more common in control subjects, but differences were not significant (P > 0.1). Eight SNPs were modestly out of Hardy-Weinberg equilibrium, even after assay redesign, in either case or control subjects (P = 0.001 for rs1135983 to P = 0.043 in rs1058405) (supplemental Table 4S), including several SNPs with a nominal association with diabetes (rs1058405, P = 0.043 in case subjects for deviation from expected Hardy-Weinberg proportions; rs1135983, P = 0.001 for deviation from expected proportions in control subjects). For both rs1058405 and rs11579627, the minor allele was overrepresented in case compared with control subjects (Table 2). Other nominally associated variants included intronic SNPs rs1027700 (P = 0.021) and rs11576878 (P = 0.029) and 3′ flanking SNPs rs2499849 (P = 0.025) and rs2490433 (P = 0.054) (Table 2). Linkage disequilibrium was similar in case and control subjects, and no haplotype was more strongly associated than individual SNPs (supplemental Table 5S).
FIG. 1.

ATF6 region map in Caucasians. A total of 16 exons are shown as darkened blocks. Bars above the figure denote -log of P values for tests of allelic association in the Utah Caucasian sample without correction for multiple hypothesis testing. Arrows denote SNPs typed in the full Consortium sample.
FIG. 2.

Linkage disequilibrium structure of 64 typed markers in the Utah Caucasian sample. Image is taken from HaploView 3.32 for the 64 SNPs typed by the International Type 2 Diabetes 1q Consortium and/or in house. Blocks are defined using the solid spline of linkage disequilibrium. Shades of gray refer to the strength of pairwise linkage disequilibrium based on r2, and numbers refer to pairwise D′ values. Black squares indicate r2 ~1, white squares are very low values of r2. Squares without numbers indicate D′ values of 1.00, and all lower values are indicated.
TABLE 2.
Summary of SNPs typed in Caucasian individual samples for ATF6 region
| Location | Name | Gene location | Allele (+ strand) | MAF |
OR (95% CI) | P | |
|---|---|---|---|---|---|---|---|
| Case | Control | ||||||
| -8150 | rs12745240 | 5′flanking | G/A | 0.439 | 0.362 | 1.375 (1.025-1.846) | 0.03 |
| -6402 | rs1417580 | 5′flanking | C/A | 0.482 | 0.414 | 1.313 (0.982-1.755) | 0.066 |
| -6250 | rs1417581 | 5′flanking | G/C | 0.476 | 0.420 | 1.257 (0.943-1.676) | 0.119 |
| -116 | rs2298019 | 5′flanking | G/A | 0.130 | 0.149 | 0.853 (0.563-1.292) | 0.44 |
| 323 | rs2271010* | Intron 1 | T/C | 0.094 | 0.092 | 1.023 (0.616-1.699) | 0.925 |
| 2088 | rs4657101 | Intron 1 | A/C | 0.195 | 0.249 | 0.731 (0.517-1.032) | 0.082 |
| 5124 | rs905594* | Intron 1 | C/T | 0.336 | 0.376 | 0.841 (0.621-1.138) | 0.258 |
| 10528 | rs2340823 | Intron 1 | T/C | 0.042 | 0.072 | 0.568 (0.301-1.073) | 0.069 |
| 10622 | rs2340824* | Intron 1 | T/C | 0.072 | 0.089 | 0.791 (0.46-1.362) | 0.376 |
| 11906 | rs2070151* | Exon 2 (L35L) | C/T | 0.042 | 0.070 | 0.585 (0.308-1.109) | 0.087 |
| 15591 | rs1058405 | Exon 3 (M67V) | A/G | 0.316 | 0.228 | 1.568 (1.13-2.175) | 0.011 |
| 17652 | rs2271013* | Exon 4 (P90P) | T/C | 0.069 | 0.074 | 0.927 (0.53-1.622) | 0.783 |
| 20886 | rs1027700* | Intron 4 | A/T | 0.321 | 0.241 | 1.493 (1.083-2.059) | 0.021 |
| 21477 | rs6427630* | Intron 4 | A/G | 0.047 | 0.070 | 0.653 (0.346-1.23) | 0.171 |
| 25126 | rs2070150* | Exon 5 (P145A) | C/G | 0.047 | 0.069 | 0.67 (0.359-1.251) | 0.192 |
| 25162 | rs1135983 | Exon 5 (S157P) | C/T | 0.045 | 0.075 | 0.579 (0.311-1.076) | 0.114 |
| 27783 | rs2134690 | Intron 6 | G/A | 0.122 | 0.142 | 0.845 (0.551-1.295) | 0.425 |
| 29845 | rs10917955* | Intron 6 | T/C | 0.047 | 0.069 | 0.655 (0.351-1.261) | 0.195 |
| 36092 | rs3738326* | Intron 7 | G/A | 0.050 | 0.075 | 0.648 (0.348-1.204) | 0.153 |
| 39427 | rs11576878* | Intron 7 | A/G | 0.320 | 0.241 | 1.482 (1.065-2.062) | 0.029 |
| 42544 | rs4656334* | Intron 7 | G/A | 0.047 | 0.072 | 0.636 (0.337-1.2) | 0.146 |
| 44829 | rs2134697 | Intron 7 | A/G | 0.271 | 0.274 | 0.986 (0.712-1.366) | 0.936 |
| 45050 | rs2134696 | Intron 7 | T/A | 0.128 | 0.138 | 0.922 (0.597-1.423) | 0.706 |
| 48255 | rs4657109* | Intron 7 | G/A | 0.069 | 0.090 | 0.758 (0.438-1.313) | 0.3 |
| 53475 | rs3817085* | Intron 8 | G/C | 0.071 | 0.090 | 0.774 (0.457-1.31) | 0.317 |
| 61198 | rs12403862* | Intron 9 | A/G | 0.047 | 0.069 | 0.665 (0.351-1261) | 0.195 |
| 62089 | rs4657110* | Intron 9 | C/T | 0.069 | 0.090 | 0.758 (0.438-1.313) | 0.3 |
| 69738 | rs2340715* | Intron 9 | G/T | 0.066 | 0.092 | 0.7 (0.409-1.198) | 0.172 |
| 75405 | rs1006310* | Intron 9 | C/T | 0.336 | 0.367 | 0.873 (0.641-1.189) | 0.385 |
| 77365 | rs1553444* | Intron 9 | T/C | 0.135 | 0.149 | 0.891 (0.585-1.358) | 0.582 |
| 84754 | rs3767635* | Intron 10 | A/G | 0.285 | 0.313 | 0.875 (0.639-1.198) | 0.408 |
| 90465 | rs3767633* | Intron 12 | A/T | 0.151 | 0.151 | 1.002 (0.672-1.495) | 0.992 |
| 94778 | rs1394066* | Intron 13 | A/G | 0.155 | 0.131 | 1.211 (0.797-1.840) | 0.368 |
| 97083 | rs3767632 | Intron 14 | G/A | 0.343 | 0.391 | 0.813 (0.605-1.093) | 0.175 |
| 98631 | rs6427632* | Intron 14 | C/T | 0.138 | 0.132 | 1.052 (0.684-1.618) | 0.818 |
| 101978 | rs1503815* | Intron 14 | C/T | 0.140 | 0.132 | 1.074 (0.705-1.634) | 0.744 |
| 111220 | rs12024729* | Intron 14 | T/A | 0.069 | 0.089 | 0.759 (0.438-1.313) | 0.3 |
| 113235 | rs2340721* | Intron 14 | C/A | 0.330 | 0.362 | 0.865 (0.636-1.177) | 0.344 |
| 120274 | rs2340722* | Intron 14 | A/G | 0.089 | 0.090 | 0.998 (0.604-1.648) | 0.992 |
| 121930 | rs4147177 | Intron 14 | C/T | 0.073 | 0.093 | 0.762 (0.452-1.287) | 0.286 |
| 122710 | rs1806594* | Intron 14 | C/T | 0.128 | 0.145 | 0.867 (0.564-1.334) | 0.509 |
| 127773 | rs6427635* | Intron 14 | T/C | 0.092 | 0.090 | 1.019 (0.609-1.704) | 0.939 |
| 135657 | rs12033967* | Intron 14 | G/A | 0.069 | 0.089 | 0.759 (0.438-1.313) | 0.3 |
| 136621 | rs2341473* | Intron 14 | G/A | 0.082 | 0.093 | 0.867 (0.521-1.443) | 0.564 |
| 144332 | rs12079579* | Intron 14 | G/A | 0.092 | 0.082 | 1.143 (0.686-1.904) | 0.602 |
| 149202 | rs2341479* | Intron 15 | A/C | 0.167 | 0.162 | 1.036 (0.695-1.543) | 0.859 |
| 152805 | rs4417044* | Intron 15 | C/T | 0.195 | 0.232 | 0.799 (0.563-1.136) | 0.193 |
| 159788 | rs2341480* | Intron 15 | T/G | 0.234 | 0.263 | 0.858 (0.609-1.210) | 0.361 |
| 163581 | rs10918215* | Intron 15 | A/G | 0.246 | 0.282 | 0.832 (0.595-1.162) | 0.263 |
| 166885 | rs7514053* | Intron 15 | G/A | 0.244 | 0.280 | 0.831 (0.594-1.162) | 0.263 |
| 168348 | rs2341481 | Intron 15 | A/G | 0.237 | 0.281 | 0.795 (0.573-1.102) | 0.159 |
| 172188 | rs10737517* | Intron 15 | A/G | 0.237 | 0.263 | 0.872 (0.620-1.227) | 0.411 |
| 178133 | rs10800151* | Intron 15 | A/G | 0.371 | 0.404 | 0.869 (0.647-1.167) | 0.348 |
| 181842 | rs10918279* | Intron 15 | C/A | 0.220 | 0.264 | 0.785 (0.561-1.1) | 0.152 |
| 188599 | rs2341471* | Intron 15 | G/A | 0.372 | 0.396 | 0.905 (0.668-1.225) | 0.514 |
| 192418 | rs13401 | 3′UTR | A/G | 0.241 | 0.296 | 0.767 (0.555-1.06) | 0.107 |
| 192621 | rs1136046 | 3′UTR | G/A | 0.090 | 0.116 | 0.752 (0.468-1.208) | 0.224 |
| 193611 | rs2499855 | 3′flanking | T/C | 0.124 | 0.149 | 0.803 (0.528-1.221) | 0.299 |
| 194804 | rs11579627 | 3′flanking | G/A | 0.334 | 0.223 | 1.751 (1.265-2.423) | 0.001 |
| 195478 | rs2499849* | 3′flanking | C/G | 0.223 | 0.293 | 0.695 (0.499-0.968) | 0.025 |
| 199870 | rs9783050 | 3′flanking | A/G | 0.376 | 0.418 | 0.84 (0.626-1.127) | 0.239 |
| 200638 | rs2490438* | 3′flanking | G/A | 0.379 | 0.315 | 1.328 (0.979-1.802) | 0.076 |
| 202745 | rs2490433 | 3′flanking | C/A | 0.411 | 0.340 | 1.354 (1.008-1.821) | 0.054 |
| 205069 | rs2490431* | 3′flanking | C/T | 0.308 | 0.337 | 0.876 (0.53-1.237) | 0.376 |
This table summarizes the association data in Utah and Arkansas Caucasians. SNPs that were in complete LD (r2 = 1) were typed only in 95 control samples and are not included in this table (see supplemental Table 3S). Likewise, SNPs with low minor allele frequencies (MAFs) (<5%) were generally not tested unless nonsynonymous. Raw counts are available in supplemental Table 3S. Alleles are listed as the major/minor allele. ORs were taken from the allelic association test considering the minor allele as the risk allele. P values are shown for genotypic association by Armitage’s Trend Test.
SNPs also genotyped by the International Type 2 Diabetes 1q Consortium in eight populations but shown here only for the Utah sample. Locations are given with respect to the A of ATG start as 1 and first base upstream as -1, based on NCBI Build 35 (University of California Santa Cruz, May 2004).
We sought to replicate the modest associations in seven additional samples, including Caucasian (1,010 case and 1,089 control subjects, excluding the Utah sample), Chinese (143 case and 144 control subjects), African-American (245 and 179, respectively), and Pima Indian (147 and 146) populations using Consortium SNPs. No SNP was associated in individual samples (data not shown), by meta-analysis of all samples or meta-analysis of Caucasian samples without the Utah contribution (supplemental Table 6S), nor did we find a trend to an association for the three nonsynonymous SNPs, rs1135983, or other SNPs associated in Utah Caucasians in 369 African-American diabetic and 186 control subjects (supplemental Table 7S). Similarly, the three nonsynonymous SNPs showed no trend to association with type 2 diabetes in the Amish Family Diabetes study (data not shown).
Defects in endoplasmic reticulum stress response could contribute to impaired insulin secretion and/or insulin action. We tested the nonsynonymous variants M67V (rs1058405, exon 3), P145A (rs2070150, exon 5), and S157P (rs1135983, exon 5) and the most strongly associated 3′ flanking SNP (rs11579627) in two Caucasian populations: a family-based study of 121 subjects from Utah and a population-based study of 209 individuals from Arkansas. No SNP altered Si or insulin secretion, measured as either AIRg, the ability of the β-cell to compensate for insulin resistance (disposition index: Si × AIRg), or AIRmax (the maximum insulin secretory capacity) (supplemental Tables 8S and 9S).
We tested for cis-acting variants by testing the allelic expression balance in cDNA prepared from Epstein-Barr virus-transformed lymphocytes from Caucasian individuals who were heterozygous for the 3′ untranslated SNPs rs1136046 and rs13401. The allelic ratio in cDNA for SNP rs1136046 fell outside the 95% CIs established from DNA from 73 heterozygotes (ratio 1.16, 95% CI 1.15-1.17) in 13 of 16 heterozygous individuals (<1 expected), with a range of 0.63-1.60 (P < 10-25) (Fig. 3), and in 3 of 10 heterozygotes for rs13401 (95% CI in genomic DNA from 148 individuals of 0.74-0.95, P = 0.0003) (Fig. 3).
FIG. 3.
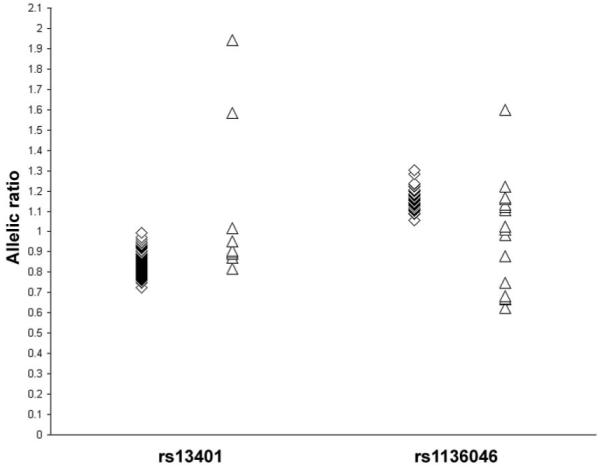
Allelic expression imbalance in ATF6 gene. Scatter plot showing allele-specific expression in DNA ( ) and cDNA (Δ) for 10 cDNA samples at rs13401 and 16 samples for rs1136046. Both cDNA ratios are significantly different from the genomic DNA ratios using the same assay.
) and cDNA (Δ) for 10 cDNA samples at rs13401 and 16 samples for rs1136046. Both cDNA ratios are significantly different from the genomic DNA ratios using the same assay.
DISCUSSION
As a key transcriptional activator at the head of the unfolded protein response, ATF6 is a strong positional and functional candidate for type 2 diabetes, particularly given recent animal data supporting a role for endoplasmic reticulum stress response in peripheral and hepatic insulin resistance (5) and insulin secretion (21). The three common nonsynonymous SNPs M67V (rs1058405), P145A (rs2070150), and S157P (rs1135983) are potentially functional. Both exon 5 variants are nonconservative substitutions, and the three nonsynonymous SNPs are located in the NH2-terminal cytoplasmic-localized transcription activation domain of ATF6, which is essential for the activation of unfolded protein response. A recent study in Pima Indians reported borderline associations of P145A (rs2070150) and S157P (rs1135983) with type 2 diabetes (P < 0.05) and reduced insulin response to oral glucose (9). We did not find an association with type 2 diabetes for SNPs rs2070150 (P145A) and rs1135983 (S157P), which are much less common in Caucasians, but we did find a nominal association with type 2 diabetes for M67V (rs1058405). However, no coding variant remained significant when P values were estimated by permutation testing. Furthermore, no SNP altered glucose homeostasis traits in our population or was associated with type 2 diabetes in our African-American population. SNP rs2070150 (P145A) was directly typed by the International Type 2 Diabetes 1q Consortium, and other Consortium SNPs were excellent proxies for M67V (rs1058405) and S157P (1135983); hence, an association with type 2 diabetes was excluded in >1,000 Caucasian case and 1,000 Caucasian control subjects. If ATF6 coding variants have an effect on diabetes susceptibility, the effect size is likely small.
The power of this study is limited for markers of effect comparable to the potassium channel KCNJ11 E23K (22) or PPARγ P12A variants (23). In the Utah sample, over the range of minor allele frequencies observed in this study, we have >70% power at P < 0.05 to detect a difference in allele frequency of 7-10% between Caucasian case and control subjects, corresponding to an OR of 1.5-1.8, or approximately the range observed for the well-replicated TCF7L2 diabetes gene (24). Considering the Consortium Caucasian sample of 1,000 case and 1,000 control subjects, we had nearly 100% power to replicate the Utah associations at SNPs rs12745240, rs11576878, rs1058405, and rs2070151. Power to detect changes in Si ranged from 66% to detect a 30% drop in log-transformed Si for common alleles (rs1058405) to 74% to detect a 50% fall in Si for an uncommon SNP, such as rs2070150. Hence, small changes would not have been detected without a considerably larger sample. Clearly, our power to detect ethnic-specific variants in Chinese or African-American populations is low, in the former case because of the small sample, and in the latter because of the low likelihood that we captured all common variants with the selected SNPs.
Based on HapMap phase 2 SNPs, we selected our markers for the detailed Utah Caucasian study to capture 95% of the variation; hence, we are unlikely to have missed common variants. Based on the 64 markers typed in the Utah Caucasian set, the 44 markers typed by the International Type 2 Diabetes 1q Consortium captured all but five SNPs (rs1275240, rs1417580, rs1417581, rs4657101, and rs1136046) at r2 > 0.8. SNPs rs1275240 and rs1417580 showed nominal associations in the Utah population and thus might become significant in a larger population. We have not resequenced the ATF6 gene, given the low likelihood that additional common variants exist that were not captured by the 64 SNPs that we chose to type. However, we have likely not captured rare variants in the ATF6 gene, if these exist. Theoretically, rare exon variants may be present and may contribute to type 2 diabetes.
Several facts support a role for ATF6 variation in disease. Several SNPs, particularly those in strong linkage disequilibrium with M67V (rs1058405), had genotypes that departed from Hardy-Weinberg equilibrium expectations, despite assay redesign, and may suggest genetic selection. SNP rs11579627, which showed the strongest association, is predicted to alter the binding of transcription factor CdxA, and the minor allele of SNP rs1136046 in the 3′ untranslated region, which showed allelic imbalance, is predicted to create binding sites for transcription factors PBX-1 and c/EBPα. Most significantly, we found evidence for cis-acting sequence variants in the gene based on the highly significant allelic expression imbalance. Because the expression ratios were both above and below the 95% CIs in different individuals, the causative cis-acting variant is likely in incomplete linkage disequilibrium with rs1136046 and rs13401. Additional studies, including allelic imbalance in other tissues, are needed to identify the likely causative variant and to determine whether the expression levels alter diabetes risk. The true functional variant, which may be considerably distant, may not have been tested for association with type 2 diabetes in this study.
In conclusion, we have thoroughly evaluated a strong chromosome 1 candidate gene. The strongest association in Utah Caucasians is in the 3′ flanking region (P = 0.02 based on 100,000 permutations), but it could not be confirmed in other populations. Both the 3′ untranslated SNP and the most strongly associated 3′ flanking region SNP alter transcription factor binding, and thus may be functional, but an undiscovered cis-acting regulatory variant appears more likely to explain the allelic expression imbalance. Hence, the possibility remains that a regulatory variant that alters ATF6 message levels also contributes to defective insulin action or insulin secretion.
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by grants from National Institutes of Health (NIH)/National Institutes of Diabetes and Digestive and Kidney Diseases (NIDDK) (DK39311) and by the Research Service of the U.S. Department of Veterans Affairs. Subject ascertainment was supported in part by grants from the American Diabetes Association and NIH/NIDDK (DK59311). Subject ascertainment and metabolic studies were supported by General Clinical Research Center Grant M01RR14288 from the National Center for Research Resources (NIH) to the University of Arkansas for Medical Sciences. Studies of the International Type 2 Diabetes 1q Consortium were supported primarily as a supplement to NIDDK Award U01-DK58026 and by NIH/NIDDK Grant DK073490 (to M.I.M.). E.Z. is a Wellcome Trust Career Development Fellow. Amish studies were supported by Grant R01 DK54361 from the NIH/NIDDK (to A.R.S.).
Glossary
- AIRg
acute insulin response to glucose
- FSIGT
frequently sampled intravenous glucose tolerance test
- SNP
single nucleotide polymorphism
Footnotes
Publisher's Disclaimer: The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
W.S.C. and S.K.D. contributed equally to this work.
Additional information for this article can be found in an online appendix at http://dx.doi.org/10.2337/db06-1305.
REFERENCES
- 1.Kaufman RJ. Orchestrating the unfolded protein response in health and disease. J Clin Invest. 2002;110:1389–1398. doi: 10.1172/JCI16886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rutkowski DT, Kaufman RJ. A trip to the ER: coping with stress. Trends Cell Biol. 2004;14:20–28. doi: 10.1016/j.tcb.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 3.Schroder M, Kaufman RJ. The mammalian unfolded protein response. Annu Rev Biochem. 2005;74:739–789. doi: 10.1146/annurev.biochem.73.011303.074134. [DOI] [PubMed] [Google Scholar]
- 4.Ron D. Proteotoxicity in the endoplasmic reticulum: lessons from the Akita diabetic mouse. J Clin Invest. 2002;109:443–445. doi: 10.1172/JCI15020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ozcan U, Cao Q, Yilmaz E, Lee AH, Iwakoshi NN, Ozdelen E, Tuncman G, Gorgun C, Glimcher LH, Hotamisligil GS. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science. 2004;306:457–461. doi: 10.1126/science.1103160. [DOI] [PubMed] [Google Scholar]
- 6.Ozcan U, Yilmaz E, Ozcan L, Furuhashi M, Vaillancourt E, Smith RO, Gorgun CZ, Hotamisligil GS. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science. 2006;313:1137–1140. doi: 10.1126/science.1128294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Elbein SC, Hoffman MD, Teng K, Leppert MF, Hasstedt SJ. A genome-wide search for type 2 diabetes susceptibility genes in Utah Caucasians. Diabetes. 1999;48:1175–1182. doi: 10.2337/diabetes.48.5.1175. [DOI] [PubMed] [Google Scholar]
- 8.McCarthy MI. Growing evidence for diabetes susceptibility genes from genome scan data. Curr Diab Rep. 2003;3:159–167. doi: 10.1007/s11892-003-0040-y. [DOI] [PubMed] [Google Scholar]
- 9.Thameem F, Farook VS, Bogardus C, Prochazka M. Association of amino acid variants in the activating transcription factor 6 gene (ATF6) on 1q21-q23 with type 2 diabetes in Pima Indians. Diabetes. 2006;55:839–842. doi: 10.2337/diabetes.55.03.06.db05-1002. [DOI] [PubMed] [Google Scholar]
- 10.Das SK, Chu WS, Hale TC, Wang X, Craig RL, Wang H, Shuldiner AR, Froguel P, Deloukas P, McCarthy MI, Zeggini E, Hasstedt SJ, Elbein SC. Polymorphisms in the glucokinase-associated, dual-specificity phosphatase 12 (DUSP12) gene under chromosome 1q21 linkage peak are associated with type 2 diabetes. Diabetes. 2006;55:2631–2639. doi: 10.2337/db05-1369. [DOI] [PubMed] [Google Scholar]
- 11.Wang H, Zhang Z, Chu W, Hale T, Cooper JJ, Elbein SC. Molecular screening and association analyses of the interleukin 6 receptor gene variants with type 2 diabetes, diabetic nephropathy, and insulin sensitivity. J Clin Endocrinol Metab. 2005;90:1123–1129. doi: 10.1210/jc.2004-1606. [DOI] [PubMed] [Google Scholar]
- 12.Das SK, Chu W, Zhang Z, Hasstedt SJ, Elbein SC. Calsquestrin 1 (CASQ1) gene polymorphisms under chromosome 1q21 linkage peak are associated with type 2 diabetes in Northern European Caucasians. Diabetes. 2004;53:3300–3306. doi: 10.2337/diabetes.53.12.3300. [DOI] [PubMed] [Google Scholar]
- 13.Zeggini E, Damcott CM, Hanson RL, Karim MA, Rayner NW, Groves CJ, Baier LJ, Hale TC, Hattersley AT, Hitman GA, Hunt SE, Knowler WC, Mitchell BD, Ng MC, O’Connell JR, Pollin TI, Vaxillaire M, Walker M, Wang X, Whittaker P, Kunsun X, Jia W, Chan JC, Froguel P, Deloukas P, Shuldiner AR, Elbein SC, McCarthy MI. Variation within the gene encoding the upstream stimulatory factor 1 does not influence susceptibility to type 2 diabetes in samples from populations with replicated evidence of linkage to chromosome 1q. Diabetes. 2006;55:2541–2548. doi: 10.2337/db06-0088. [DOI] [PubMed] [Google Scholar]
- 14.Elbein SC, Hasstedt SJ, Wegner K, Kahn SE. Heritability of pancreatic beta-cell function among nondiabetic members of Caucasian familial type 2 diabetic kindreds. J Clin Endocrinol Metab. 1999;84:1398–1403. doi: 10.1210/jcem.84.4.5604. [DOI] [PubMed] [Google Scholar]
- 15.Sherry ST, Ward MH, Kholodov M, Baker J, Phan L, Smigielski EM, Sirotkin K. dbSNP: the NCBI database of genetic variation. Nucleic Acids Res. 2001;29:308–311. doi: 10.1093/nar/29.1.308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Altshuler D, Brooks LD, Chakravarti A, Collins FS, Daly MJ, Donnelly P. A haplotype map of the human genome. Nature. 2005;437:1299–1320. doi: 10.1038/nature04226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005;21:263–265. doi: 10.1093/bioinformatics/bth457. [DOI] [PubMed] [Google Scholar]
- 18.Weinker S. DeFinetti Program. 2005. [Accessed 20 January 2007]. Available from http://ihg.gsf.de/cgi-bin/hw/hwal.pl.
- 19.Pacini G, Bergman RN. MINMOD: a computer program to calculate insulin sensitivity and pancreatic responsivity from the frequently sampled intravenous glucose tolerance test. Comput Methods Programs Biomed. 1986;23:113–122. doi: 10.1016/0169-2607(86)90106-9. [DOI] [PubMed] [Google Scholar]
- 20.Boston RC, Stefanovski D, Moate PJ, Sumner AE, Watanabe RM, Bergman RN. MINMOD Millennium: a computer program to calculate glucose effectiveness and insulin sensitivity from the frequently sampled intravenous glucose tolerance test. Diabetes Technol Ther. 2003;5:1003–1015. doi: 10.1089/152091503322641060. [DOI] [PubMed] [Google Scholar]
- 21.Harding HP, Ron D. Endoplasmic reticulum stress and the development of diabetes: a review. Diabetes. 2002;51(Suppl. 3):S455–S461. doi: 10.2337/diabetes.51.2007.s455. [DOI] [PubMed] [Google Scholar]
- 22.Florez JC, Burtt N, de Bakker PI, Almgren P, Tuomi T, Holmkvist J, Gaudet D, Hudson TJ, Schaffner SF, Daly MJ, Hirschhorn JN, Groop L, Altshuler D. Haplotype structure and genotype-phenotype correlations of the sulfonyl-urea receptor and the islet ATP-sensitive potassium channel gene region. Diabetes. 2004;53:1360–1368. doi: 10.2337/diabetes.53.5.1360. [DOI] [PubMed] [Google Scholar]
- 23.Altshuler D, Hirschhorn JN, Klannemark M, Lindgren CM, Vohl MC, Nemesh J, Lane CR, Schaffner SF, Bolk S, Brewer C, Tuomi T, Gaudet D, Hudson TJ, Daly M, Groop L, Lander ES. The common PPARgamma Pro12Ala polymorphism is associated with decreased risk of type 2 diabetes. Nat Genet. 2000;26:76–80. doi: 10.1038/79216. [DOI] [PubMed] [Google Scholar]
- 24.Grant SF, Thorleifsson G, Reynisdottir I, Benediktsson R, Manolescu A, Sainz J, Helgason A, Stefansson H, Emilsson V, Helgadottir A, Styrkarsdottir U, Magnusson KP, Walters GB, Palsdottir E, Jonsdottir T, Gudmundsdottir T, Gylfason A, Saemundsdottir J, Wilensky RL, Reilly MP, Rader DJ, Bagger Y, Christiansen C, Gudnason V, Sigurdsson G, Thorsteinsdottir U, Gulcher JR, Kong A, Stefansson K. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat Genet. 2006;38:320–323. doi: 10.1038/ng1732. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.