

Abstract
Cytoadherance of Plasmodium falciparum-infected erythrocytes in the brain, organs and peripheral microvasculature is linked to morbidity and mortality associated with severe malaria. Parasite-derived P. falciparum Erythrocyte Membrane Protein 1 (PfEMP1) molecules displayed on the erythrocyte surface are responsible for cytoadherance and undergo antigenic variation in the course of an infection. Antigenic variation of PfEMP1 is achieved by in situ switching and mutually exclusive transcription of the var gene family, a process that is controlled by epigenetic mechanisms. Here we report characterisation of the P. falciparum silent information regulator's A and B (PfSir2A and PfSir2B) and their involvement in mutual exclusion and silencing of the var gene repertoire. Analysis of P. falciparum parasites lacking either PfSir2A or PfSir2B shows that these NAD+-dependent histone deacetylases are required for silencing of different var gene subsets classified by their conserved promoter type. We also demonstrate that in the absence of either of these molecules mutually exclusive expression of var genes breaks down. We show that var gene silencing originates within the promoter and PfSir2 paralogues are involved in cis spreading of silenced chromatin into adjacent regions. Furthermore, parasites lacking PfSir2A but not PfSir2B have considerably longer telomeric repeats, demonstrating a role for this molecule in telomeric end protection. This work highlights the pivotal but distinct role for both PfSir2 paralogues in epigenetic silencing of P. falciparum virulence genes and the control of pathogenicity of malaria infection.
Author Summary
The unicellular parasite Plasmodium falciparum is the cause of the most severe form of malaria and is responsible for 300 million infections and ∼2 million deaths a year. Infected erythrocytes clump and block capillaries in the peripheral circulation, the brain, and placenta and are a major contributor to the pathology of malaria. A parasite-derived protein displayed on the surface of the infected erythrocyte is responsible for erythrocyte clumping in capillaries. Although 60 subtelomeric var genes can encode different versions of this “sticky” capillary-binding protein, only one protein is expressed at a time, and switches in expression between these genes causes variation of this pathogenic molecule enabling the parasite to evade the immune system. Here we identify two chromatin-modifying proteins that cooperate to mediate silencing and mutual exclusive expression of var genes. These proteins are thus important virulence factors of the malaria-causing parasite.
Investigation into two Sir2 histone deacetylases in the malaria-causing parasite revealstrans-acting epigenetic factors control mutually exclusive expression of a major subtelomeric virulence gene family.
Introduction
Phenotypic variation is essential for survival in a changing and competitive environment. In the case of pathogenic organisms, antigenic variation of cell surface proteins is a common mechanism to avoid clearance by the host immune system (for review see [1]). Although many different strategies exist for the regulation of antigenic variation, most rely on the mutually exclusive expression of gene families, that is, the expression of one variant and the silencing of all others [1,2]. By switching antigenic variants pathogens are able to avoid adaptive immune responses, thus maintaining a persistent infection and increasing the chances of transmission to the next host.
The protozoan parasite P. falciparum, the causative agent of the most severe form of malaria in humans, undergoes antigenic variation in its asexual blood stage, an important property of the parasite both for virulence and evasion of host immune responses (for review see, [3]) [4]. P. falciparum infects up to 300 million people a year resulting in the deaths of over 2 million annually [5]. Infection can manifest in a variety of ways and in severe cases cerebral abnormalities and organ failure can occur, which often leads to death. Such complications of infection are multifactorial but cytoadherance of infected red blood cells to the microvasculature and to uninfected erythrocytes plays an important role [3]. Parasite-derived P. falciparum Erythrocyte Membrane Protein 1 (PfEMP1) is exposed on the surface of the infected red blood cell and responsible for cytoadherance to receptors on the microvasculature, brain, and placenta. Capillary blockages and local inflammatory responses ensue and disease progresses. Thus, PfEMP1 is a key P. falciparum virulence factor and an important target of the host immune system [4].
To maintain cytoadherance in the presence of a mounting immune response PfEMP1 undergoes antigenic variation by switching expression between the ∼60-member var gene family that encodes these proteins [6–10]. Var gene expression is controlled at the level of transcription [11,12] and switching to other genes involves no DNA rearrangements [13]. Like many other antigenically variant gene families in pathogenic organisms, most var genes occupy subtelomeric regions of P. falciparum chromosomes where gene silencing and recombination occur at high frequency [14–16]. There are also clusters of var gene at more central regions on five of the 14 chromosomes [10,14].
Var genes have a highly conserved promoter region that fall into three distinct families on the basis of their physical position and orientation along the chromosome [10,17]. The UpsB promoters control var genes that are most telomere proximal and are transcribed towards the centromeres. UpsA promoters and the related but distinct UpsE promoter of the highly conserved placental PfEMP1 variant var2CSA drive expression of var genes that are also within subtelomeric regions but are transcribed towards the telomere. The UpsC promoter-type controls the expression of internal chromosome var genes [17,18]. Prediction of binding phenotypes based on ligand domain structure suggests that PfEMP1 molecules encoded by UpsB and UpsC var genes most commonly bind the endothelial CD36 and ICAM-1 receptors, whereas UpsA and the related UpsE var genes encode PfEMP1 molecules that are thought to bind to other receptors such as complement receptor on red blood cells (causing red blood cell rosetting) and chondroitin sulphate A (CSA) in the case of placental malaria [19,20].
Chromosome ends in P. falciparum are tethered to the nuclear periphery and are clustered into 3–7 distinct foci [15,16]. The subtelomeric DNA repeat rep20 participates in chromosome tethering [21,22] and an unknown proteinaceous molecule(s) binds telomere ends together [16]. Most of the nuclear periphery contains electron dense material consistent with heterochromatin, suggesting that this region is transcriptionally silent [11,23]. Indeed, var gene silencing involves heterochromatin formation, and activation of var loci correlates with a “relaxation” of nucleosomal architecture and intranuclear repositioning of the activated gene [11,24,25].
Recently, it has been shown that the highly conserved var gene promoters are key to mutual exclusive expression of var genes [26,27]. There is an indication that the single intron in the conserved genetic structure of var genes plays a role in silencing [28], although this is under debate [29]. Certain DNA elements within the highly conserved var gene promoters have been shown to bind specific, but yet unidentified proteins that could be involved in var activation and silencing [30]. One of these DNA elements has a role in nucleosomal organization although much work remains to decipher the role of cis-elements in control of var gene expression [27].
Reversible histone-tail modifications and ATP-dependent nucleosome remodelling are important for epigenetic control in eukaryotic gene regulation. Indeed, activation and silencing of var genes correlates with specific histone tail marks, suggesting epigenetic memory is involved in maintenance and switching of var gene repertoire. Histone H3 lysine 9 acetylation and methylation of lysine 4 have been shown to be associated with var gene activation [31], whereas tri-methylation of lysine 9 of histone H3 is associated with var gene silencing [31,32].
Given the association of histone modifications with var gene expression, molecules that catalyse their addition and removal are likely to be important factors controlling the expression of PfEMP1. The only molecule known to be involved in the regulation of var gene transcription is a homologue of the yeast silent information regulator 2 (Sir2) family [23,24]. Sir2 was originally isolated as a suppressor of subtelomeric gene expression in yeast [33] and has subsequently been shown to be a NAD+-dependent histone deacetylase [34]. Sir2 molecules, or sirtuins as they are commonly known, are found throughout the tree of life and catalyse the removal of acetyl groups from a variety of substrates in many cellular processes [35]. Enzymatically, PfSir2 represents a classic NAD+-dependent histone deacetylase, catalysing the removal of acetyl-groups from histone tails [36]. Some versions also harbour the ability to ribosylate [37], including the previously characterised PfSir2 (here called PfSir2A) [38]. PfSir2 has also been shown to associate with silenced var genes and localise to chromosome-end clusters all along the noncoding telomere-associated repeat elements (TARE 1–6) and telomere repeats within the nucleus [23,39]. Furthermore, the genetic ablation of PfSir2A in P. falciparum results in the derepression of a subset of var genes, thus further implicating histone hyper-acetylation in var gene activation and demonstrating PfSir2A as an important virulence factor of P. falciparum infection [23,24].
Very few trans-acting factors have been identified that control antigenic variation in pathogenic protozoa, and in this work we characterise the first two that are involved in regulation of var gene expression in P. falciparum. We show that two paralogues of the Sir2 NAD+-dependent histone deacetylase family cooperate to regulate the expression of the var gene repertoire. We show that var transcription is regulated by Sir2 paralogues interacting from within conserved promoter regions. Furthermore, we show that Sir2 paralogues also play a part in the regulation of other antigenically variant genes as well as telomere repeat length. Our work highlights these epigenetic factors as key regulators of antigenic variation and thus pathogenicity of the malaria parasite and further provides impetus that other pathogenic protozoa that undergo antigenic variation may also have a similar reliance on such factors for control of their virulence genes.
Results
Identification of Sir2 Homologues in P. falciparum
Previously, it has been shown that a P. falciparum homologue of the histone deacetylase Sir2 (here after referred to as PfSir2A) is involved in the regulation of antigenic variation [23,24]. We set out to identify other factors that could be involved in epigenetic regulation of P. falciparum var genes. BLAST searching on PlasmoDB (www.plasmodb.org) identified a protein with similarity to PfSir2A and other sirtuins (Figure 1A). This Sir2 homologue, here after referred to as PfSir2B (PlasmoDB accession number PF14_0489), is much larger than PfSir2A and is found on Chromosome 14 (Figure 1A). We compared PfSir2B with the ∼250-amino acid (AA) catalytic domains of sirtuin family members and with PfSir2A. PfSir2B shares 29%/43% identity/similarity with yeast Sir2, a “group I” sirtuin and 26%/51% identity/similarity with PfSir2A, which has previously been defined as a “type III” sirtuin (Figure 1A) [40]. PfSir2B shows the greatest similarity with group IV sirtuins with 38% and 53% identity and similarity, respectively (Figure 1A). Interestingly, the predicted catalytic domain of PfSir2B is split in two, separated by an insertion of approximately 180 amino acids (Figures 1A and Figure S1).
Figure 1. Identification of PfSir2B and Phylogenetic Relationships between PfSir2A and PfSir2B within Apicomplexan Parasites.

(A) PfSir2B is a large gene and encodes a ∼150-kD protein. PfSir2B, like all sirtuins, contains similarity to other Sir2 molecules within the catalytic regions (black). PfSir2B has 29%/43% identity/similarity with Schizosaccharomyces pombe Sir2 (SpSir2) and 26%/51% identity/similarity with the previously identified PfSir2A. PfSir2B most closely resembles a “type IV” sirtuin, which groups together other sirtuins with diverse roles in cellular signalling such as human SIRT6, involved in telomeric chromatin maintenance [82,83] as well as SIRT7, an activator of RNA polymerase I [84]. PfSir2B has with 38%/53% identity/similarity to type IV sirtuins.
(B) Phylogenetic analysis of apicomplexan Sir2 paralogues. Sir2 homologues were identified from the sequencing projects of four Plasmodium species; P. vivax [85], P. chabaudi [86], P. yoelli [86], P. berghei [86], together with Sir2 homologues from other apicomplexan parasites: T. gondii (http://toxodb.org), T. annulata [87], T. parva [87], B. bovis (www.tigr.org), and Cryptosporidium hominis [88]. Within the analysis we also included Sir2 from yeast, other related species, and unicellular protozoan parasites. Bootstraps values are indicated for important nodes, where in all cases the upper number is the bootstrap using distance, the lower is the bootstrap using maximum likelihood. “-“ sign indicates a value below 50. Two Sir2 paralogues are a common feature of apicomplexan parasites. Sir2B paralogues (red branches) show strong support for the Sir2B clade, especially within Plasmodium species. Sir2B paralogues also groups together type IV sirtuins (SIRT6 and 7) with good support. Sir2A clade (blue branches) groups together Sir2A paralogues of apicomplexan species with excellent support. Sir2A's moderate grouping with SIRT5 and Archaeoglobus fulgidus Sir2-Af1 also shows a possible functional relationship with type III sirtuins. For more in depth analysis see Text S1. P. ber, P. berghei; P. yoe, P. yoellii; P. cha, P. chabaudi; P. viv, P. vivax; P. fal, P. falciparum; T. ann, T. annulata; T. parv, T. parva; T. gondi, T. gondii; B. bov, B. bovis; A. ful, A. fulgidus; H. sap, Homo sapiens; C. hom, C. hominis; T. pse, Thalassiosira pseudonana (diatom); G. lam, Giardia lamblia; K. lac, Kluyveromyces lactis; S. cer, S. cerevisiae; L. maj, Leishmania major.
Phylogenetic analysis was performed on Sir2 homologues identified from a range of apicomplexan parasites as well as a diverse selection of eukaryotes. Unrooted trees suggest there are two ancient and distinct apicomplexan Sir2 types. One of these groups strongly with other Sir2 proteins previously described as type IV sirtuins (here Sir2B), while the other does weakly with type III sirtuins (here Sir2A) (Figure 1B). Every Plasmodium species analysed, plus Toxoplasma gondi (a coccidian apicomplexan parasite), possesses one of each of these types, while the piroplasm apicomplexans Theileria and Babesia appear to have secondarily lost their Sir2A paralogue.
What is evident from our analysis is that Sir2 paralogues have been co-opted for the regulation of var genes. T. gondii also contains Sir2A and Sir2B orthologues although they are unlikely to be involved in antigenic variation given that no subtelomeric gene family has been identified, even with the genome project near completion (www.toxodb.org). Other Plasmodium species analysed here lack var genes, but genome projects have revealed other subtelomeric gene families in these parasites that appear to be antigenically variant. We suggest in this work that Sir2 paralogues in P. falciparum directly regulate var gene promotors, thus it seems likely that specific factors have co-opted Sir2 paralogues to regulate var genes.
To unravel potential similarities and differences in function between Sir2 paralogues an alignment of the catalytic domain was created (Figure S1). We noted that PfSir2B largely has good conservation of residues required for catalytic activity and zinc binding. Residues that represent the acetylated substrate-binding site show some diversity suggesting a change in substrate (Figure S1). For further analysis of the phylogenetic tree and similarities and differences within the catalytic domain between other sirtuins see Text S1. Our analysis clearly demonstrates two Sir2 paralogues are a common feature of malaria parasites and points to a possible role of these molecules in subtelomeric silencing and antigenic variation.
Disruption of the PfSir2B Gene in P. falciparum
To determine the function of PfSir2B in P. falciparum we constructed a parasite line, from the parent strain 3D7, in which the gene encoding the corresponding protein had been disrupted using the plasmid vector pHHΔpfsir2B (Figure 2A) [41]. Southern blot analysis confirmed that the locus had been disrupted by insertion of the transfected plasmid by single crossover homologous recombination (Figure 2B). Furthermore, pulse-field gel electrophoresis showed the plasmid was integrated into a high molecular weight chromosome consistent with Chromosome 14, which encodes pfsir2B (Figure S2) [10]. To confirm that the gene had been disrupted we probed RNA from ΔPfSir2B with pfsir2B and no specific transcript could be detected, whereas ΔPfSir2A parasites, which encoded an intact pfsir2A gene, transcribed similar levels when compared to the parental line 3D7, suggesting there is no compensatory mechanism of Sir2 paralogues (Figure 2C). An antibody derived from the C-terminal portion of PfSir2B recognised a protein of the predicted size of 150 kDa in 3D7 parasites that was absent in ΔPfSir2B parasites (Figure 2D). We wished to compare the expression of var and other subtelomeric genes between wildtype and Sir2-disrupted lines, and given the high variation in these areas between isolates we confirmed that all of the derived parasite lines were isogenic by fingerprint analysis using the rep20 repetitive probe (Figure S2) [42].
Figure 2. PfSir2B Gene Disruption in P. falciparum .

Single crossover recombination was used to disrupt PfSir2B gene in P. falciparum. Bacterially derived plasmids were introduced containing an ∼1-kb fragment of the 5′ end of the PfSir2B gene and an hDHFR expression cassette for stable selection of transgenic parasites. Cycles of “on” and “off” drug were used to select for parasites that had integrated the plasmid by homologous integration into the PfSir2B locus.
(A) Theoretical restriction mapping of the Sir2B locus, pre- and postepisome integration. Hatched line denotes bacterially derived plasmid sequence, while AmpR signifies beta lactamase expression cassette for selection for resistance on Ampicillin. The line under the 5′ end of the PfSir2B gene shows the probe (P) used for Southern blot mapping. B, BglII site.
(B) Southern blot of wildtype (wt) and ΔPfSir2B disrupted parasites. Banding pattern is consistent with disruption of the Sir2B gene by multiple plasmid copies (as mapped in [A]). DNA ladder sizes are indicated on the left of the blot.
(C) Northern blot showing a loss of PfSir2B transcript in ΔPfSir2B-disrupted parasites but not in ΔPfSir2A-disrupted parasites. Same probe (P) in Southern blot was used for northern blot.
(D) Western blot of antisera raised against a C-terminal portion of PfSir2B. Antisera reacting with a protein in wildtype parasites contain the predicted size for PFSir2B, while this protein is absent as expected in ΔPfSir2B parasites. Cross-reactive bands indicate equal loading of lanes.
PfSir2A and PfSir2B Regulate Silencing of Distinct Groups of var Genes in P. falciparum
To determine if PfSir2B, a putative histone deacetylase, has a role in gene regulation within the intraerythrocytic life cycle of P. falciparum we performed global transcription analysis with ΔPfSir2A, ΔPfSir2B, and the parental 3D7 strain using a custom-designed, 2.5-million feature Affymetrix chip based on the fully sequenced P. falciparum genome (3D7 strain version January 2005). Parasites were synchronised and RNA prepared from three time points within the ∼48-h intraerythrocytic lifecycle corresponding to: ring stage, 8–12 h post-invasion (pi); trophozoite stage, 24–28 h pi; and schizont stage, 38–42 h pi. Ring-stage parasites represent a time of host cell remodelling and expression of antigenically variant proteins that are exported to the P. falciparum-infected erythrocyte surface [43,44]. Trophozoites represent the period of growth and haemoglobin digestion, whereas schizonts define a period of daughter cell formation for subsequent egress and invasion into new erythrocytes. Three biological replicates for each stage and for all parasite lines were used for quality control. Similar variability between all samples demonstrated that no bias was introduced during the processing of parasites handling and sample preparation (unpublished data).
The Sir2 protein in yeast affects the expression of subtelomeric genes, and the P. falciparum homologue PfSir2A has previously been implicated in the silencing of a subset of var genes in early ring stages [24]. Consequently we determined the transcriptional profile of subtelomeric genes within the P. falciparum genome in both ΔPfSir2A and ΔPfSir2B parasites. In ΔPfSir2A and ΔPfSir2B we physically mapped the pattern of gene expression within subtelomeric regions of all 14 P. falciparum chromosomes, and the five internal var gene clusters (Figure 3). Strikingly, we found that between the ΔPfSir2A and ΔPfSir2B parasite lines, essentially all var genes were transcribed. Also, it was evident that there was a correlation between the particular pfsir2 gene disrupted and the physical position and direction of transcription of the activated gene (Figure 3). That is, primarily the most telomere proximal var genes (i.e., controlled by an UpsB promoter) were highly activated in ΔPfSir2B, whereas the subtelomeric var genes transcribed towards the telomere and those in internal chromosomal clusters (i.e., controlled by UpsA, UpsE, and UpsC) were highly transcribed in ΔPfSir2A (Figure 3, rainbow pattern of expression blue to green and yellow to purple, respectively, see also Figure S3). This suggests that PfSir2A plays an important role in silencing of UpsA, UpsC, and UpsE controlled var genes whilst PfSir2B silences the nonoverlapping group controlled by UpsB promoters.
Figure 3. PfSir2A and PfSir2B Control the Expression of Subtelomeric Genes in P. falciparum .

The expression profile of subtelomeric genes and internal var gene clusters were analysed in Sir2-disrupted lines and applied to a physical map of all 28 chromosome ends. Subtelomeric ends contain most var genes as well as the rifin gene family, which are also known to be antigenically variant. Other gene families occupy subtelomeric regions of P. falciparum chromosomes, but at this stage their biological function or whether they are antigenically variant remains elusive. As seen by the key the colour of each gene relates to the level of expression in each PfSir2-disrupted line. Black genes indicate that expression of that gene is not observed in any condition (expression signal <75th percentile). In the unusual case where PfSir2A or PfSir2B both affected the expression of a single var gene only the most dramatic change is shown. This physical map of gene expression accompanies Figure S3, which graphs the expression values of all subtelomeric genes in 3D7 and the two PfSir2-disrupted lines.
Analysis of var Gene Transcription in P. falciparum Parasites Lacking PfSir2A or PfSir2B Function
The expression of var genes was further analysed in wildtype and the two Sir2-disrupted lines. A Gaussian “self”-clustering analysis was performed on the transcription pattern of the full repertoire of var genes in the 3D7 genome given nine different conditions set out in our array experiments (i.e., three parasite lines; ΔPfSir2A, ΔPfSir2B, and 3D7 at three different time points; rings, trophozoites, and schizonts) (Figure 4A). Var genes were also labelled by their promoter type, UpsA, B, C, E, D, var-like, and hybrid promoters BC and BA [18]. Gaussian self-clustering grouped transcription profiles into three distinct clades with cluster 4 containing outliers (Figure 4A). Cluster 1 included var genes that are expressed in parental 3D7 during standard culture conditions and they were also expressed in ΔPfSir2A and ΔPfSir2B parasites (Figure 4A). There was no obvious bias towards the promoter type driving the expression of these var genes, nor could we find any other feature that defined this cluster except that no UpsA-driven var gene was found. This most likely represents the “stable” var gene transcription observed in the parental population of 3D7 in standard culture conditions that has not been selected for any cytoadherence phenotype.
Figure 4. PfSir2 Paralogues Silence Discrete Subsets of var Genes Associated with Conserved Promoter Types That Drive Their Expression.

(A) Unsupervised nonhierarchical Gaussian self-clustering analysis was performed in order to understand the relationship between var gene subtypes and the role of PfSir2 paralogues in their regulation. Genes were clustered by similarity using Pearson coefficients. Heatmap colour-scale values were assigned by ArrayMiner after further transformation of expression data (see Material and Methods for details) with +3 (yellow) being the highest expression and −3 (blue) being the lowest expression. Expression profiles of the var gene repertoire were grouped given the nine conditions examined by Affymetrix arrays (i.e., rings, trophozoites, and schizonts in 3D7 [wildtype], ΔPfSir2A, and ΔPfSir2B parasite lines). The heatmap clearly identifies three major clusters that are biologically relevant. Cluster 1 represents var genes that are expressed in standard culture conditions in 3D7 parasite isolate and are somewhat expressed in PfSir2-disrupted parasites. No correlation in promoter type is seen in var genes found is this cluster with the exception that no UpsA-var genes were found there. Var genes controlled by UpsA conserved promoter type are found within cluster 2 as are most of the internally located UpsC var genes. This finding suggests that PfSir2A plays a suppressive role in expression of var genes with an UpsA or UpsC promoter type. Cluster 3 contains almost exclusively UpsB var genes and these are de-repressed in the absence of PfSir2B. This suggests that PfSir2B is involved in the suppression of the most telomere proximal, UpsB driven var genes. Cluster 4 contains several var genes that seem to be outliers, not conforming or reaching the threshold required to fall into one of the three main clusters. Scale bar of expression goes from −3 to 3 in transformed log2 expression.
(B) Raw normalised log2 expression of all var genes in ring-stage parasites aligned with their corresponding heatmap in the Gaussian self-clustering analysis. Raw normalised expression values for all var genes grouped by their promotor type as assayed by array analysis can be viewed in Figure S4. The colour of each bar refers to its strain identity: green, 3D7; red, ΔPfSir2A; blue, ΔPfSir2B.
Cluster 2 represents var genes that are poorly expressed in parental 3D7 but that are highly upregulated in ΔPfSir2A at ring stage suggesting a role for this molecule in silencing this subset of genes (Figure 4A). Interestingly, all var genes controlled by an UpsA promoter (subtelomeric and transcribed towards the telomere) are contained within this cluster (Figure 4A). This cluster also contains a significant proportion of UpsC promoter controlled var genes. UpsC promoters drive the expression of var genes that are found in internal chromosome locations, showing that PfSir2-mediated silencing is not restricted to subtelomeric genes (Figure 4A) and that PfSir2A is required for their silencing. Several var genes controlled by hybrid promoters, in which the 3′ end represents an UpsB promoter and the 5′ end has similarity to UpsC or UpsA promoters (i.e., UpsBC and UpsBA), are also found within this cluster, which suggests that a common feature of these hybrid var promotors allows their control by PfSir2A.
Cluster 3 includes var genes that are derepressed in the absence of PfSir2B and therefore suggests a role for this histone deacetylase in the silencing of this var gene subset (Figure 4A). Of the 15 var genes within this cluster, UpsB promoters control 13, the other two are hybrid promoters (UpsBA and UpsBC). Several UpsB var genes also show a high degree of statistical significance in transcription between parental and knockout (Figure S4) and these were confirmed by quantitative PCR (qPCR) (see below and Table S1). The remaining seven var genes do not fall into a distinct cluster (although here called cluster 4 in Figure 4A) owing to atypical expression patterns (MAL7P1.186, MAL7P1.212, MAL8P1.220, PFF1640w), but also perhaps because of the derepression phenotype not meeting the threshold for the Gaussian clustering (MAL13P1.1, PFL0005w) (Figure 4A). Figure 4B displays the normalised log2 raw expression data of all var genes, in rings, in a bar graph format as an alternative way to view the data. Individual var genes are aligned with the var genes as they appear in the Gaussian clustering analysis (Figure 4B). This data can also be viewed in an alternate way in supporting information as raw normalised expression of each var gene, grouped according to var promoter type in bar graph format (Figure S4). qPCR was used to confirm the Affymetrix array results (see below and Table S1). In conclusion, P. falciparum Sir2 paralogues play a role in silencing the var gene repertoire and there is a strong correlation between the var promoter type and the PfSir2 molecule that regulates its activity.
PfSir2 Paralogues Control Mutual Exclusive Transcription of var Genes
Antigenic variation of PfEMP1 occurs by monoallelic activation of a single var gene and silencing of all others [13]. Switching between PfEMP1 variants occurs at approximately 1% per generation and this involves, through unknown mechanisms, silencing of one var gene and activation of another [45]. Given that we are analysing a population of parasites expressing a range of var genes it was not possible to conclude from our initial analysis whether the absence of PfSir2A and B function results in a greater frequency of switching or a breakdown in the mutually exclusive program by the derepression of multiple var genes in a single parasite. To discern between these possibilities parasites were selected for adherence to CSA, which is mediated by a PfEMP1 protein encoded by the var2csa gene (3D7 genome: PFL0030c) [10,46], and determined the transcriptional levels of the var gene repertoire. Selection of ΔPfSir2A and ΔPfSir2B parasites on CSA allowed us to determine the integrity of var gene mutual exclusion in a ΔPfSir2A and ΔPfSir2B genetic background (Figure 5).
Figure 5. Disruption of PfSir2 Paralogues Destroys Mutual Exclusive Expression of var Genes.

In wildtype parasites selection on CSA enriches for parasites exclusively expressing var2CSA. We used this mono-allelic binding phenotype to assay if PfSir2-disrupted parasites still maintain mutual exclusive expression of var genes.
(A) 2-D FACS plots showing binding of R1945 rabbit serum specifically recognises infected erythrocytes (ethidium bromide [EtBr] positive) containing var2CSA-expressing parasites. R1945 percentage positivity and negativity are separated on the basis of fluorescence levels of uninfected erythrocytes. Selection on CSA of 3D7 (3D7_CSA) greatly enhances the reactivity to R1945 serum and is comparable to the CS2 parasite line that stably expresses var2CSA. Given that ΔPfSir2A already expresses var2CSA in the absence of selection it is not surprising that CSA selection of this parasite line (ΔPfSir2A_CSA) does nothing (if anything a negative effect) to the binding of R1945 serum. ΔPfSir2A parasites show an intermediate binding of R1945 reactivity suggesting that they have lower levels of VAR2CSA on their surface. Selection of ΔPfSir2B parasites on CSA (ΔPfSir2B_CSA) also greatly enhanced R1945 binding to infected erythrocytes suggesting the ΔPfSir2B parasites are a heterogenous pool and are not confined to expressing only UpsB-driven var genes.
(B) As expected qPCR of var2CSA (PFL0030c) transcript levels shows a dramatic increase upon CSA selection on 3D7 and ΔPfSir2B parasites, but not ΔPfSir2A.
(C) Mutual exclusive expression of var genes is broken down in the absence of both PfSir2A and PfSir2B. As expected, 3D7_CSA mainly expresses var2CSA and while other var genes are only at a very low levels (see also Figure S5). However, when compared to 3D7_CSA, PfSir2-disrupted parasites, not only express var2CSA but also express many other var genes at much higher level than 3D7_CSA does (silenced state). This finding strongly suggests that mutual exclusive expression has broken down in these parasites. Interestingly, expression of var genes in the Sir2-disrupted lines selected on CSA also shows a bias towards var genes with certain promoter types thus confirming by another method our earlier findings.
3D7, ΔPfSir2A, and ΔPfSir2B were selected for high levels of CSA adherence to produce the lines 3D7_CSA, ΔPfSir2A_CSA, and ΔPfSir2B_CSA. These lines were firstly analysed by flow cytometry (FACS) for expression of VAR2CSA, the PfEMP1 protein responsible for CSA adherence [46], using antibodies that could specifically detect this protein (Figure 5A) [47]. 3D7_CSA parasite-infected erythrocytes expressed increased levels of VAR2CSA compared to 3D7 consistent with selection of parasites expressing this PfEMP1 protein (Figure 5A) [48]. The level of VAR2CSA expression for 3D7_CSA was comparable to CS2, a parasite line that more stably expresses this PfEMP1 variant (Figure 5A). The ΔPfSir2A parasites transcribe elevated levels of var2csa along with other var genes controlled by UpsA and E promoters [24], consequently it was not surprising that ΔPfSir2A_CSA parasites expressed similar levels of VAR2CSA on the surface of infected erythrocytes compared to ΔPfSir2A_CSA (Figure 5A). It should be noted that by scatter plot PfSir2A-disrupted parasites do not bind as much anti-VAR2CSA antibody suggesting that they have less VAR2CSA on their surface (Figure 5A). In contrast, ΔPfSir2B parasites do not express VAR2CSA as anti-VAR2CSA binding to the parasite-infected erythrocyte was low. However, upon CSA selection ΔPfSir2B_CSA-infected erythrocytes had high levels of anti-VAR2CSA (R1945) reactivity, suggesting these parasites retain the capacity to activate expression of other PfEMP1 antigenic types (Figure 5A).
To address the role of PfSir2A and B on var gene switching and monoallelic exclusion at the transcriptional level qPCR was performed with cDNA from 3D7, ΔPfSir2A, and ΔPfSir2B parasites before and after CSA selection to determine alterations in transcription of this gene family (Figure 5B and 5C; Table S1). As expected the var2csa gene in 3D7 was dramatically increased in transcription after selection in 3D7_CSA, which correlated with increased reactivity to VAR2CSA antibodies (Figure 5B) [49]. All other var genes were down regulated to apparently basal levels (Figures 5C and S5) consistent with an intact mutually exclusive var expression program. Given that ΔPfSir2A already expresses var2csa at high levels [24], selection on CSA did not substantially affect its level of gene transcription (Figure 5B). Interestingly, and in concordance with the anti-VAR2CSA antibody binding, selection of ΔPfSir2B parasites on CSA dramatically increases var2csa transcription in these parasites 5–10-fold compared to 3D7_CSA (Figure 5B). These results suggest that loss of PfSir2B function has not disrupted the ability of these parasites to activate specific silenced var genes.
To determine if the PfSir2 paralogues play a role in control of mutually exclusive expression, the transcription of var genes was analysed before and after selection for adherence to CSA (Figure 5C). Given that all var genes except var2csa are silenced in 3D7 parasites selected on CSA, this analysis was used as a benchmark for the var gene silenced state (Figures 5C and S5). Comparing the expression of var genes in ΔPfSir2-disrupted parasites selected on CSA with 3D7_CSA parasites it can be seen that in both ΔPfSir2A_CSA and ΔPfSir2B_CSA express multiple var genes at higher levels than 3D7_CSA. Indeed, some var genes in ΔPfSir2A and ΔPfSir2B are expressed higher than in 3D7 (Table S1). These data suggest that mutually exclusive expression of var genes has broken down in the absence of PfSir2 paralogues and thus both these molecules play a critical role in the control of mutually exclusive expression of var genes (Figure 5C). It is also worth noting that ΔPfSir2A_CSA and ΔPfSir2B_CSA infected erythrocytes can bind to CSA and CD36 and this together with anti-VAR2CSA antibody reactivity is consistent with expression of multiple PfEMP1 variants on individual infected erythrocytes and the breakdown of mutual exclusion (unpublished data). Both before and after selection of ΔPfSir2A and ΔPfSir2B for CSA adherence a similar pattern of var gene transcription was observed consistent with an important role for PfSir2A and PfSir2B in silencing of specific var gene subsets (Figure 5C; Table S1).
Using this type of analysis it also looks as though PfSir2A and PfSir2B can have an effect on the transcription of the same var gene suggesting that they are acting cooperatively rather than exclusively (Figure 5C). For example, UpsB-driven var genes are largely regulated by PfSir2B, however, using qPCR and CSA selection PfSir2A clearly has an effect on UpsB controlled gene transcription (greater log2 value) (Figure 5C). The reciprocal is true for UpsA and UpsC var genes where parasites lacking PfSir2B function transcribe greater levels of UpsA var genes compared to the same genes silenced in 3D7 parasites. This result suggests that PfSir2 paralogues act cooperatively to control the transcription of individual var genes and warranted further investigation.
PfSir2 Paralogues Have a Direct Effect on the Silencing of var Gene Promoters
Previously, we showed that the default state of a var promoter is silenced and stochastic activation occurs at low frequency [26,27]. Additionally, a drug resistance marker can be inserted into the var gene antigenic program by placing it under the control of a var promoter so that its activation by drug selection enforces silencing of all var genes [26,27]. To investigate whether PfSir2 paralogues directly regulate the silencing of var genes by acting on the promoter we introduced plasmids with an UpsB (PFL0005w; subtelomeric) or UpsC (PFL1960w; internal) var promoter driving hdhfr in 3D7, ΔPfSir2A, and ΔPfSir2B parasites and assayed promoter activity by qPCR. Plasmids also contained the blasticidin-S deaminase (bsd) gene driven by the constitutive hsp86 promoter for selection of transfected parasites (pUpsB/hDHFR and pUpsC/hDHFR) (Figure 6A).
Figure 6. PfSir2 Paralogues Directly Regulate var Gene Promoters and Contribute to Spreading of Transcriptional Silencing into Neighbouring Regions.

(A) Plasmids bearing two expression cassettes were introduced into 3D7, ΔPfSir2A, and ΔPfSir2B parasites. The bsd cassette is driven by the housekeeping hsp86 promoter, which provides a selection mechanism for parasites stably maintaining episomes. UpsB and UpsC var promoters were placed in front of the selectable marker hdhfr and therefore var promoter activity can be monitored. All values plotted were normalised against the var promoter in its activated state.
(B) PfSir2A and B paralogues differentially affect the silencing of isolated var promoters. hdhfr transcript level was measured in 3D7 parasites with silenced and activated episomally located UpsB and UpsC promoters as well as the level of hdhfr transcription in the absence of PfSir2 paralogues. Both PfSir2 paralogues can directly affect the activity of a var promoter strongly suggesting that PfSir2 paralogues do not function independently of one another on different var promoters but rather cooperate to regulate expression. In both subtelomeric and internally derived var promoters isolated from their natural context PfSir2B has a larger effect on the suppression of var promoter activity.
(C) PfSir2A and PfSir2B control the spreading of silencing from var promoters. Upon activation of the downstream var promoter activity of bsd expression cassette increases suggesting that factors that normally silence var gene promoters can spread in cis to modulate expression of neighbouring regions. In the absence of both PfSir2A and PfSir2B spreading of silencing is reduced suggesting that both paralogues mediate the spreading of silencing from both internal and subtelomerically derived var gene promoters. In both cases PfSir2B has a greater influence on spreading of silencing to within the bsd cassette. qPCR was performed on three biological replicates harvested from ring-stage parasites. 95% confidence error bars derived from a calculated standard error are also displayed.
In 3D7-pUpsB/hDHFR and 3D7-pUpsC/hDHFR, selection with the antifolate WR99210 provided a parasite population expressing activated versions of the UpsB and UpsC promoters respectively [26,27]. As expected hdhfr gene transcript levels (after normalising for plasmids copy number) are dramatically increased compared to WR99210 unselected lines (Figure 6B). ΔPfSir2A and ΔPfSir2B parasites containing either pUpsB/hDHFR or pUpsC/hDHFR transfected parasites could not be selected on WR99210 because hDHFR (and WR99210 selection) was used to make the PfSir2 gene disruptions. Consequently var promoter activity was determined in the default state using qPCR.
Analysis of the UpsB and UpsC var promoter activity in ΔPfSir2A-pUps(B/C)/hDHFR and ΔPfSir2B-pUps(B/C)/hDHFR (after subtracting the transcript signal of hDHFR from ΔPfSir2A and B lines) showed that absence of PfSir2A and B function results in a partial derepression of the hdhfr gene (Figure 6B). These results demonstrate that PfSir2-mediated silencing can act directly on var promoters and does not require cis-spreading of silenced chromatin from neighbouring telomeres as is observed in Sir2-mediated subtelomeric silencing in yeast [50]. Interestingly, this analysis shows that both Sir2 paralogues play a role in the silencing of individual var promoters, as first observed earlier in qPCR of var genes in PfSir2-disrupted lines selected on CSA (Figure 5C). Interestingly, loss of either PfSir2A or B function causes only partial derepression compared to a fully activated var gene promoter, strengthening the idea that both molecules cooperate in silencing these var promoters. The absence of PfSir2B shows greater activation of the isolated UpsB var promoter (PFL0005w), strongly supporting the evidence that PfSir2B plays a greater role in the regulation of UpsB var genes than does PfSir2A. In the case of the UpsC var promoter (PFL1960w) there is a greater derepression of this promoter in the absence of PfSir2B, which is somewhat contradictory to the var qPCR and microarray analysis of the endogenous PFL1960w gene in all the cell lines (Figure 6B). One possible explanation for this is slight differences in the complex regulatory network of var genes when they are located in different chromosomal contexts (i.e., internal chromosome location versus extrachromosomal episome). Var gene promotors are key to the control of mutual exclusive expression of var genes, and our results strongly implicate PfSir2 paralogues as major trans-acting factors influencing the control of these highly conserved elements. This clearly highlights the importance of PfSir2 paralogues in the control of var gene silencing and mutual exclusion.
Spreading of Silencing Is Mediated by PfSir2 Paralogues
The spreading of heterochromatin from its nucleation point is important to achieve gene and regional silencing [51]. Previously we have shown that silencing can spread from a var promotor [26,27]. Sir2 in yeast is known to be involved in spreading of silencing chromatin [50], and we reasoned that PfSir2A and B could also be involved in this process in P. falciparum. We investigated whether PfSir2 paralogues had a role in spreading of silencing by monitoring the expression level of a bsd expression cassette upstream of a var promotor (Figure 6A). We assayed the transcript level of bsd (driven by the housekeeping hsp86 promoter) by qPCR in 3D7, ΔPfSir2A, and ΔPfSir2B parasites containing pUps(B/C)hDHFR plasmids (Figure 6C). Parasites with an active, plasmid-based UpsB or UpsC promoter driving the hDHFR gene in 3D7 had significantly increased levels of the bsd transcript, suggesting spreading of var gene silencing factors to the hsp86 promoter, as has been observed previously [26]. Interestingly, upon introduction of pUpsB/hDHFR into ΔPfSir2A and B parasites the activation state of the bsd cassette differed from that of parental 3D7 parasites with a silenced, plasmid-located var promoter (Figure 6C). As observed with the activation of var promoter in Sir2-disrupted lines, the effect on the upstream bsd cassette differed depending on which PfSir2 paralogue was not functional. In the case of UpsB-based plasmid we found an almost complete abrogation of silencing in the PfSir2B-disrupted line and a more mild alleviation of silencing in parasites lacking PfSir2A (Figure 6C). A similar phenomenon was observed with UpsC-based plasmids, suggesting that spreading of silencing is less reliant on the function of PfSir2B (Figure 6C). These data strongly suggest that both PfSir2 molecules, to differing degrees, play a role in the spreading of a transcriptionally repressed state from var promoters.
Temporal and Transcriptional Regulation of the Variant rif Gene Family Is Altered by Loss of PfSir2B or PfSir2A Function
Repetitive interdispersed family (rif) genes constitute the largest gene family in P. falciparum consisting of upwards of 150 members, most of which are within subtelomeric regions [10,52]. Other than var, rif genes are the only other gene family to date that has been shown to be antigenically variant [52] and are normally expressed in trophozoite-stage parasites [52]. The encoded Rifin proteins are trafficked to the infected erythrocyte, and some data suggest that they are located on the red blood cell surface [52]. By analysing microarrays we found that a proportion of the rif gene family was activated in ΔPfSir2A and ΔPfSir2B parasites and these often have a close physical association with an activated var gene (Figure 3) (see below for further analysis). Gaussian self-clustering was performed on the rif genes with respect to the nine conditions under which the microarray analysis was performed (Figure 7). Ninety rif genes sorted into ten different clusters, whilst the remainder were not transcribed under any of the conditions and were therefore excluded from the analysis.
Figure 7. rif Genes Change Expression Timing upon Disruption of PfSir2A, but not PfSir2B.

Gaussian self-clustering was used to further analyse the role of PfSir2 paralogues in rif gene expression. Expression throughout rings, trophozoite, and schizont-stage parasites show that PfSir2A can modulate the temporal expression of rif genes. Normally most rif are expressed in trophozoite stage but upon disruption of PfSir2A expression of some of this antigenically variant gene family changes to either ring- or schizont stage, or both. Interestingly the lack of PfSir2B shows a suppressive effect on rif gene expression. Unfortunately no other similarities either in promoter sequence or physical position relative to var genes could be found in any of the clusters.
Analysis of rif transcription across the nine conditions demonstrated differences between both ΔPfSir2A and B parasites (Figure 7). It was apparent that ΔPfSir2A parasites contain changes in raw transcriptional levels of rif genes compared to parental, as was observed with var genes, however, a shift in transcriptional timing was dramatically evident accounting for most of the variation observed compared with 3D7 parasites (Figure 7). Lack of PfSir2A function results in alteration of the temporal regulation for rif gene transcription from trophozoites to ring stage (clusters 1, 9, and 10) and in some cases also to schizont stage (clusters 4 and 7). Comparison between 3D7 and ΔPfSir2A parasites showed that rif genes in clusters 1, 2, 3, and 8 in ΔPfSir2A were expressed at ring stages, whereas clusters 4 and 7 were expressed mainly at schizont stages, even though some transcription can still be accounted for at ring-stage parasites (Figure 7). Clusters 5, 6, 9, and 10 showed slightly different variations. While the rif genes in clusters 5 and 6 remain expressed at the trophozoites stage, their level of transcription varied in ΔPfSir2A trophozoites and was higher in cluster 5 but lower in cluster 6 (Figure 7). For the rif genes classified in clusters 9 and 10, whilst they showed no detectable transcription in parental 3D7, they were activated in ΔPfSir2A and also showing a shift in their temporal pattern (Figure 7). Interestingly, ΔPfSir2B parasites do not transcribe any rif genes at the schizonts stages suggesting that loss of PfSir2B function results in repression of this gene family (Figure 7). Transcription of some rif genes showed a shift towards ring stages as was found for ΔPfSir2A parasites but had a decreased level of transcription when compared to the 3D7 parent (Figure 7, clusters 1, 8, and 9).
We attempted to find features common to the rif genes found in each individual cluster, similar to the var gene analysis showing that the conserved promoter types also fall into distinct clusters. Rif promoters are also conserved [53], and the encoded rifin genes have been phylogenetically classified into two groups [54]. We performed phylogenetic analysis on the 1.5-kb promoter regions of the rif genes in Figure 7, and the cluster number was mapped to each rif gene within the phylogenetic tree. We found no correlation of relatedness between promoter sequence and the likelihood of being expressed in either PfSir2-disrupted line, suggesting that unlike var genes the rif promoter does not have a prominent role with respect to regulation of this gene set by PfSir2A or B. Additionally, there was no strong correlation between the physical positioning of rif genes compared to var genes other than the strongest expressed rif transcripts are found head-to-head with expressed var genes in ΔPfSir2A parasites (unpublished data). These results show that loss of PfSir2A or B function has a strong effect on a proportion of the rif genes suggesting they are important in the regulation of this variant gene family.
The Role of PfSir2A and PfSir2B in Regulation of Other Genes in P. falciparum
Transcriptional analysis of the ΔPfSir2A and ΔPfSir2B parasites showed that PfSir2A and B proteins played an important role in silencing of var genes and regulation of the rif gene family, but surprisingly had virtually no effect on other genes within the P. falciparum genome with some notable exceptions. A small number of genes other than var and rif located within the subtelomeric region of chromosomes showed increased transcription in ΔPfSir2A, which included the pfmc-2tm gene family [55]. This family consists of 12 members and ten are transcribed in the 3D7 parent (unpublished data). Removal of PfSir2A function in ΔPfSir2A resulted in increased transcription of the ten genes that were already transcribed to some extent (unpublished data). Figure S3 displays the physical map of subtelomeric genes and internal var gene loci with accompanying expression graphs for each gene throughout the intraerythrocytic time points applied to the array.
After removing all the var and rif genes from the analysis we scanned the remaining differentially expressed genes for common features (Table S2). Interestingly, we found that most differentially expressed genes found in either ΔPfSir2A or B was transcribed in ring-stage parasites. We also noticed that PfSir2A disruption affected expression of more genes compared to loss of PfSir2B function. Members of several other gene families changed expression profile in ΔPfSir2A and B lines, including several members of the large 72-member Plasmodium helical interdispersed subtelomeric (PHIST) family [43,56], a stevor gene, and an etramp (Table S2) [57]. The differentially expressed genes were compared with respect to their features and physical position along the chromosome and no obvious shared properties were identified (Table S2). We also specifically looked for other genes known to confer parasite phenotypic variation, and the expression of rDNA genes (which are affected by Sir2 in yeast) and we found no differences. For more in depth analysis please consult Text S1.
PfSir2A Plays a Role in Maintenance of Telomere Length
Telomeres recruit proteins that facilitate spreading of heterochromatin into subtelomeric noncoding and coding regions [58]. In P. falciparum, PfSir2 and PfORC1 associate with telomere and subtelomeric heterochromatin [39]. It has been shown in other systems that telomere length is influenced by factors that affect the chromatin fibre, including Sir3 and Sir4 from yeast [59]. Thus, it has been speculated that these factors are involved in telomere end protection. We reasoned that PfSir2 paralogues might be involved in this process. To assay the effect of PfSir2 paralogues on maintenance of telomere length we performed telomere restriction fragment (TRF) analysis as described previously [22]. This technique takes advantage of the lack of restriction sites within telomeric repeat DNA as a means to liberate intact telomeres. Southern blot on genomic DNA digested from 3D7, ΔPfSir2A, and ΔPfSir2B with four restriction enzymes that cut frequently within the P. falciparum genome but do not cut in telomeric repeats were probed with telomeric repeats (Figure 8). In a population of wildtype (3D7) parasites telomere repeat length varies from ∼1 kb to ∼2 kb over all chromosomes with a mean length of approximately 1.5 kb. In parasites lacking PfSir2A, but not PfSir2B an increase in telomere length is evident with an increase in mean length of 3 kb in length, approximately double the length of telomeric repeats in wildtype (3D7) and ΔPfSir2B parasites (Figure 8). It is also interesting to note that four sharp bands are evident in parasites lacking ΔPfSir2A in the range of 1 to 1.5 kb (Figure 8). At this stage it is uncertain what these bands represent in the chromosomal context. Given the fuzzy nature of chromosome ends due to variable number of telomere repeats, a likely explanation is that significant blocks of repeats have been moved to internal chromosome regions, leading to defined bands after TRF analysis. Thus, PfSir2A appears to increase chromosome end instability. This is supported by the Rep20 RFLP analysis (arrow, Figure S1, which indicates possible changes in the RFLP profile compared to 3D7 and ΔPfSir2A.) Interestingly, we noticed that there are four chromosome ends that do not contain a functional var most proximal to the telomeric repeats: left arm of Chromosome 6 (psuedo-var), the right arm of Chromosome 8 (hypothetical genes), and the two arms of Chromosome 14 (pseudo-vars) (Figure 3). This finding suggests a link between var gene regulation and telomere length by PfSir2 paralogues.
Figure 8. Sir2A, but Not PfSir2B Has a Role in Telomere Length Regulation.
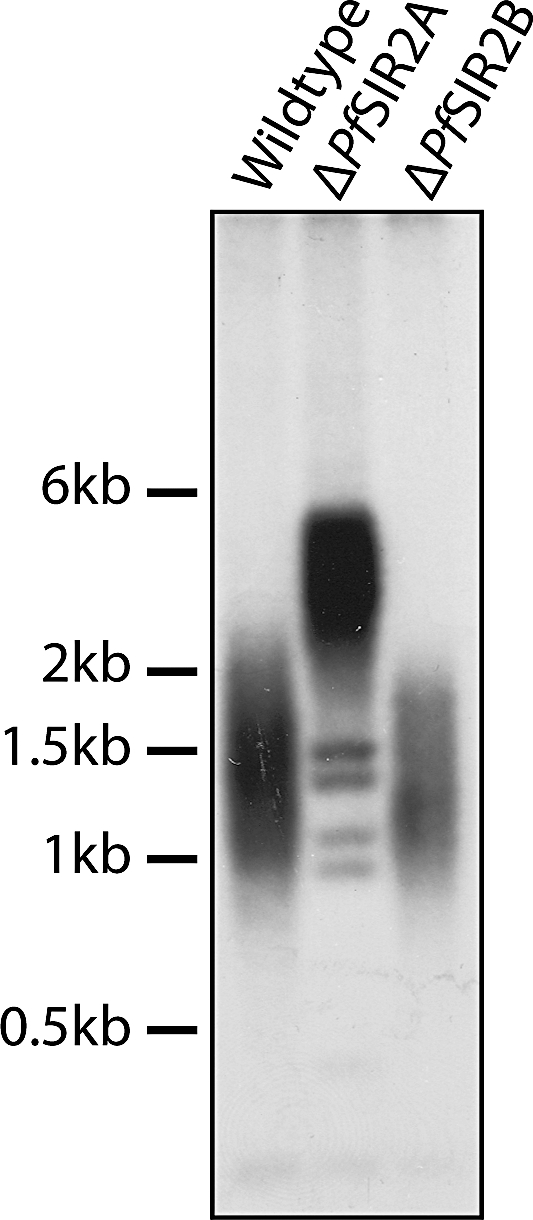
Genomic DNA from wildtype (3D7), ΔPfSir2A, and ΔPfSir2B was digested with four frequently cutting restriction enzymes that do not cut telomeric repeats. Samples were run on 1% agarose gel and probed with telomeric repeat-labelled with 32P. Telomere length is seen as a smear as telomere lengths vary between chromosomes and within a population telomere. 3D7 parasites have telomere lengths of between 1–2 kB. Whereas upon disruption of PfSir2A but not PfSir2B, telomere length approximately doubles in size. PfSir2B-disrupted parasites have several distinct bands of a length that correspond to nonlengthened telomeres. It is also possible that they represent telomeric repeats that have recombined into chromosome internal locations and thus are not able to lengthen.
Discussion
Antigenically variant PfEMP1 molecules are encoded by the var gene family and transcribed in a mutually exclusive fashion and controlled by epigenetic mechanisms [23,24,60]. We have shown here that two paralogues of the Sir2 NAD+-dependent histone deacetylase family cooperate to achieve silencing of the full var gene repertoire. This is, to our knowledge, the first demonstration of paralogous epigenetic modifiers cooperating to control the expression of a gene family and the first evidence of cooperation by Sir2 paralogous in the control of mutual exclusion of pathogen virulence factors.
The PfSir2 paralogues have separate but somewhat overlapping functions, and although both proteins have an effect on transcriptional repression of individual var genes PfSir2A has a greater role in silencing the subtelomeric UpsA promoter-driven genes (which are transcribed towards the telomeres) and the chromosomal internally located UpsC driven var genes [10]. PfSir2B, on the other hand, has a more prominent role in the silencing of var genes controlled by the subtelomerically located UpsB promoter type (which are transcribed towards the centromere). Both PfSir2A and B regulate var gene transcription by acting directly on the conserved promoters, which agrees with previous results analysing var promoters [26,30]. It appears likely that DNA elements within the conserved promoter types are a point that initiates var gene promoter silencing and spreading by recruitment of protein complexes containing either PfSir2A or PfSir2B. DNA elements that bind nuclear proteins have been experimentally identified in the different var promoter subsets [30]. The DNA element SPE2 was defined in UpsB promoters and mediates repression throughout ring and trophozoite-stage parasites when inserted into the housekeeping calmodulin promoter [27]. This DNA element is a candidate for initiating PfSir2B-mediated suppression in UpsB promoters by recruitment of proteins for heterochromatin formation and spreading of silencing.
Var promoters were analysed in an attempt to define a region that may be involved in initiating binding of PfSir2-containing complexes by using the hybrid UpsBA and UpsBC promoters that are regulated differentially by PfSir2A and B. The 3′ end of these promoters represents an UpsB-type promoter, whilst the 5′ resembles either an UpsA or C promoter generated by recombination between the different var gene subsets [18]. By utilizing the breakpoint in homology between these hybrid promoters we attempted to identify candidate regions of the promoter that could bind and initiate PfSir2-mediated silencing. However, we did not find any obvious conserved DNA elements in the promoter regions and it is likely the sequences involved are not well conserved. It will require further analyses using chromatin immunoprecipitation (ChIP) with PfSir2-specific antibodies and functional assay of the var promoter regions to identify these elements. Unfortunately the antibodies described here were not specific enough to address this question.
Both PfSir2 paralogues are required for mutually exclusive expression of var genes because ΔPfSir2A and ΔPfSir2B parasites express multiple genes and PfEMP1 proteins. Previous work has suggested that an active var gene is located in a specific subnuclear expression site at the nuclear periphery [11,24,25], and a candidate region is located at the nuclear periphery that lacks the electron dense appearance of heterochromatin [11]. The telomeric regions of chromosomes form clusters of 3–7 chromosome ends attached to the nuclear periphery, and in order for a var gene to be active it moves either to an “active” cluster [24] or outside a cluster to a transcriptionally permissive domain [11]. The UpsC var genes, that are more centrally located, also appear to be located within clusters at the nuclear periphery consistent with this area being important for both silencing and activation of this gene family [11]. Telomeric clusters are formed in the absence of PfSir2A [16] and PfSir2B (A. Marty, B.S. Crabb, and A.F. Cowman, unpublished data) function, and thus these proteins are not important for formation and maintenance of these chromosomal structures. It is not clear how parasites lacking PfSir2A or B function are able to accommodate transcription of multiple var genes once monoallelic exclusion has broken down, as these active genes must be located at the nuclear periphery. It is possible that loss of PfSir2A or B function results in breakdown of the subnuclear architecture spreading the transcriptionally permissive area or alternatively that multiple var genes can be transcribed within the existing permissive domain.
Both PfSir2A and B have a role in var regulation by acting directly on specific var promoter subsets and mediating spreading of the silenced state, which is in agreement with previous data [26,27]. This result is in contrast to studies in yeast where subtelomeric gene silencing is reliant on the spreading of silenced chromatin from the telomeric repeats and not direct binding to promoter regions (see for review [51]). Mechanistically, yeast subtelomeric transcriptional silencing is reliant on the spreading of silenced chromatin from telomeres and is mediated by the histone deacetylase activity of Sir2 [50]. At a molecular level this is thought to occur by the binding of RAP1 to the telomeric repeats, binding of the SIR complex to RAP1, followed by deacetylation of adjacent N-terminal Histone H3 and H4 tails. Subsequent binding of the SIR complex to the deacetylated histone tails then allows for deacetylation of adjacent histone tails and it is this iterative process that propagates the spreading of a silenced chromatin state inward into subtelomeric regions [50]. The apparent lack of RAP1 and SIR homologues in any of the sequenced apicomplexan genomes (www.plasmodb.org) [10] together with results presented here, suggest that this model is unlikely in P. falciparum. Rather our data support a model in which complexes containing either PfSir2A or PfSir2B can directly bind var promoter regions, regulating their expression and furthermore, spread silent chromatin in cis.
Subtelomeric regions of chromosomes in a variety of organisms are associated with epigenetic gene regulation [61]. Acetylation of H3 and H4 histone tails are associated with euchromatin and active gene transcription while the removal of these marks by histone deacetylases is linked to gene silencing and conversion of nucleosomes into a heterochromatic state [62]. In yeast Sir2, a specialised NAD+-dependent lysine histone deacetylase is found that suppresses the activity of subtelomeric genes [36]. Recently in P. falciparum it has been shown that histone H3 lysine 9 (H3K9ac) and acetylated histone H4 marks an active UpsE (related to UpsA) var promoter, while the PfSir2A homologue we also describe here binds to these promoter regions in P. falciparum [23,31]. Furthermore, we have shown here and elsewhere that PfSir2A is involved in its suppression [24].
PfSir2B also has a role in subtelomeric var gene silencing but mechanistically and biochemically it is not clear how this takes place. Although there is a dramatic size difference in the P. falciparum Sir2 paralogues, they share common features of the catalytic domains found in sirtuin homologues that have been studied structurally [63]. Interestingly, PfSir2B shows differences in its substrate-binding pocket suggesting these molecules bind different cellular targets. PfSir2A has previously been shown to localise to the nuclear periphery and associate with silenced, but not active var genes [23]. Furthermore, PfSir2A can deacetylate H3 and H4 histone tails and also ribosylate [38]. Given that PfSir2A and PfSir2B have different functions and contain differences in their substrate-binding pocket, it is likely that their cellular substrates differ.
It was surprising that there were so few genes affected by loss of PfSir2A or B function with the biggest effect being contained within the var and rif variant families suggesting epigenetic regulation may not be widely relevant to other genes in the asexual stages. PfSir2A, and to a lesser extent PfSir2B, has an effect on the level and temporal expression of the rif gene family. Rif genes sometimes have close physical proximity to var genes and have highly conserved promoters [10]. However, neither of these features correlated with derepression patterns in the absence of the PfSir2 paralogues. It appears that spreading of silenced chromatin from var genes is not a major factor in activation or alteration of temporal transcription of this gene family. The small number of genes affected by PfSir2A or B, other than var and rif, have no common features except that some are located within the subtelomeric region of the chromosomes. It will be interesting to determine if they have any common functional region within their respective promoters that render them sensitive to the PfSir2 proteins.
Telomeres are special nucleoprotein structures that protect chromosome ends from recombination and degradation. In most eukaryotes telomeres are assembled into tightly packed chromatin fibre [64]. Telomeres of P. falciparum form nucleosome and non-nucleosomal chromatin at the chromosome [65]. The biology of P. falciparum telomeres remains elusive apart from the characterisation of an unusually large telomerase subunit (TERT) [66,67]. No other trans-acting factors have been characterised that are required for telomere stability or length regulation. It has been shown that telomere length changes occur at truncated chromosomes in P. falciparum and this DNA sequence effect occurs only in cis [22]. Here we have identified PfSir2A, a heterochromatin component, that not only contributes to gene repression but affects telomere length at chromosome ends.
In the ΔPfSir2A parasites four DNA bands of sizes corresponding to unlengthened telomeres were present either corresponding to a subset of telomeres not lengthening in the absence of PfSir2A function or telomeric repeats that have recombined to internal locations. It was also evident using the rep20 RFLP analysis (a measure of subtelomeric integrity) that there were some differences in the PfSir2A-disrupted line when compared to wildtype parasites suggesting a PfSir2A role in chromosome stability. It is also interesting to note that there are four chromosome arm ends that do not have a functional var gene proximal [10] to the subtelomeric repeats drawing a possible link between var gene and telomere length regulation. It is also worth noting that in other systems telomere length is linked to genomic instability [68], and our data suggest also that this might also be the case for P. falciparum.
In summary, we have characterised the role of two Sir2 NAD+-dependent histone deacetylases on var gene regulation and show an important role for both paralogues in regulating antigenic variation through mutually exclusive transcription of the full var gene repertoire. The PfSir2 paralogues interact directly with the conserved var promoter subsets and have overlapping but distinct roles. It is not clear why P. falciparum parasites require the interplay of two PfSir2 paralogues for silencing of this important gene family. UpsA promoters control var genes that contain unusual cytoadherance-binding phenotypes, and studies suggest that their expression is associated with disease, whereas other var genes are associated with asymptomatic infections [69,70]. If these findings are confirmed, then perhaps PfSir2A has a role in suppressing the exposure of this limited repertoire of unusual cytoadherance phenotypes, and PfSir2B acts more as a general mediator of silencing and protection of the mutually exclusive program. Whatever the reason, given the clear importance of PfEMP1 in antigenic variation and cytoadherance, PfSir2 paralogues and other unidentified epigenetic factors are key determinants of pathogenicity in malaria infection.
Materials and Methods
Phylogenetic analysis.
Sequences were aligned using ClustalX v1.83 and alignments were manually adjusted using BioEdit Alignment editor. Phylogenetic trees were inferred from 253 positions of the alignments using both maximum likelihood and distance methods. Distance trees were made using both Neighbor joining and Kitsch methods using the PHYLIP 3.67 package. Distance bootstrapping was carried out for 1,000 replicates using the Seqboot bootstrapping method in PHYLIP 3.67 [71]. Maximum likelihood trees were made using PhyML version 2.5 [72] at http://www.phylogeny.fr/phylo_cgi/phyml.cgi, with the Whelan and Goldman (WAG) substitution model. Bootstrapping was performed for 100 replicates. In addition to the trees depicted, phylogenies were inferred from several alignment variants, including or excluding taxa or using fewer or greater alignment characters. Most nodes were insensitive to these variants, but the positioning and order of group III sirtuins varied with different datasets, indicating that this may not be a robust grouping.
Cloning, transfection, and culturing.
Parasites lacking PfSir2B were created by the electroporation and selection for homologous integration of a bacterially derived plasmid harbouring a 5′ flank of the genomic sequence of the PfSir2B gene. The first 1,023 bp (including ATG) of the PfSir2B gene was amplified from 3D7 gDNA and ligated into the BglII/XhoI sites of pHHM (primer sequences available on request) [21]. Transfection and selection proceeded as previously described [73,74]. Cycling on and off drug selected for a population of parasites in which the plasmid has integrated via single crossover homologous recombination into the endogenous PfSir2B locus. Parasites were then cloned out by limiting dilution.
For investigations into the direct role that PfSir2 paralogues play on var promoter silencing and spreading, pHBUpsB [27] and pHBUpsC were introduced into wildtype (3D7), ΔPfSir2A, and ΔPfSir2B parasites and selected for stable maintenance with blasticidin-S. Wildtype parasites harbouring either plasmid were then also treated on WR99210 for selection of parasites expressing the ectopic UpsB promoter.
Northern blotting.
Trophozoite-stage parasites were harvested using ice-cold saponin and resuspended in TRIzol (Invitrogen). Northern blots were performed as previously described [75]. Blots were probed with [32P]-dATP radiolabelled DNA and washed at 60 °C with 0.5 × SSC and 0.1% SDS prior to autoradiography. The ∼500-bp PfSir2B probe was amplified from within the flank used for the knockout construct (same as for southern blot).
Southern blotting.
Southern blot analysis on ΔPfSir2B parasites was performed using standard procedures. gDNA was cut using BglII and runout on a 0.8% agarose gel. After transfer onto nitrocellulose a ∼500-bp fragment from the homologous flank containing incorporated dioxygenin-dATP was used as a probe (Roche). Primers for the amplification of the probe are available on request. Probe was detected on the membrane using anti-DIG-HRP antibodies and chemoluminescence detection (Roche).
Telomere length blot was performed as previously described [22]. Briefly, gDNA from 3D7, ΔSir2A(3D7), and ΔSir2B(3D7) was harvested from mixed-stage parasites. 2 μg of gDNA was cut with the four enzymes AluI, DdeI, MboII, and RsaI, which frequently cut within the Plasmodium genome, but not within the telomeric repeats. gDNA was run out on a 1% gel and transferred to Hybond N+ nitrocellulose membrane (Amersham). Membranes were probed with two complementary 99-mer oligonucleotides corresponding to nucleotides 11–110 of the left arm of Chromosome 13 (oligonucleotide sequences available on request).
Antibody production.
DNA corresponding to amino acids 811–953 of PfSir2B was amplified from 3D7 gDNA and ligated into pGEX-4T1. Purified, bacterially expressed GST-PfSir2B 3′ was used to immunize rabbits. Before use, rabbit antiserum was purified over Protein-G sepharose (Amersham).
Global expression analysis.
To analyse global gene expression profiles within intraerythrocytic cycles RNA was collected from highly synchronous cultures of ring-, trophozoite-, and schizont-stage parasites. Briefly, 3D7, ΔPfSir2A, and ΔPfSir2B parasites were tightly synchronised with 5% sorbitol twice 16 h apart. Parasites were harvested at 8–12 h, 24–28 h, and 38–42 h post invasion. Infected red blood cell pellets were immediately resuspended in TRIzol (Invitrogen) and frozen at −80 °C. RNA was extracted as per manufacturers protocol (Invitrogen) and further digested with RNase free DNase (Ambion).
P. falciparum (Pf) samples were hybridised on the custom PFSANGER Affymetrix array. PFSANGER Affymetrix arrays are high-density 8-μm custom 25-mer oligonucleotide arrays, whose tiling-like design was based on the P. falciparum (3D7 strain) genomic sequence released in January 2005 (www.genedb.org) [76]. 10 μg of total RNA was reverse transcribed and biotin-labelled as cRNA, using the GeneChip IVT Labelling kit as recommended by Affymetrix. Hybridisations were carried out at 45 °C for 16 h with constant rotation at 60 rpm. Following hybridisation, the solutions were removed and the arrays washed and stained on a fluidics station (Affymetrix FS450). Gene arrays were then scanned at an emission wavelength of 570 nm at 1.56 μm pixel-resolution using a confocal scanner (Affymetrix GeneChip Scanner 3000 7G). After scanning, the hybridisation intensity for each 25-mer feature was computed using Affymetrix GCOS version 1.3 software.
The raw data was then transferred into R/Bioconductor [77] and preprocessed using the Affy package [78], principally the RMA function, which relies on a robust multi-array averaging algorithm (RMA) for background adjustment, normalization, and summarization of the probe sets [79]. All probes mapping a CDS on P. falciparum genome (www.genedb.org, June 2007 update) were assigned in a probe set, giving a total of 5356 Pf CDS.
For a specific gene-analysis, we undertook a nonhierarchical clustering based on expression signals for each of the nine conditions (three cell lines: ΔPfSir2A, ΔPfSir2B, and wildtype (3D7) at three different time points: rings, trophozoites, and schizonts) for all the probe sets representing the genes of interest. The genes were clustered by measuring similarity of shapes using Pearson coefficients. The unsupervised clustering program enabled us to discriminate biological clusters (ArrayMiner, OptimalDesign). ArrayMiner software searches for solutions of minimal total variance in a given number of clusters. However, unlike most other currently available methods, ArrayMiner does not rely on k-means in its search for high quality clusters as it uses a GGA optimisation process (the total variance of the clusters being minimized). To draw heatmaps, the expression values for each gene were slid vertically so that their average was zero (zero mean normalisation) then multiplied so that the length of the associated vector (square root of the sum of squares of all values) be equal to 1 (unit-norm normalisation of length 1). The resulting contrasts were plotted on a scale from −3 (blue, low expression) to +3 (yellow, high expression).
CSA selection of P. falciparum-infected erythrocytes.
Parasites were cultured to trophozoite stage and enriched away from uninfected erythrocytes by gelatin selection [80] and bound to CSA as described previously [81]. Parasite lines were selected in parallel five times, which was enough to change R1945 binding profile of 3D7 to close to 100% positive suggesting that selection had gone to completion.
FACS analysis.
Expression of Var2CSA-specific PfEMP1 was assessed by FACS using established methods [47]. In short, cells were sequentially incubated with R1945 (rabbit serum), and Alexa-Fluor 488 conjugated donkey anti-rabbit Ig (Molecular Probes)/ethidium bromide (BioRad). All incubations were performed in PBS/0.1% casein for 30 min at room temperature. Samples were analysed on a Becton-Dickinson FACSCalibur flow cytometer. Using FlowJo (TriStar) IgG binding for each sample was expressed as the geometric mean fluorescence intensity (MFI) for infected erythrocytes (ethidium bromide positive), after subtracting the MFI for uninfected erythrocytes.
qPCR.
For the expression analysis RNA was harvested from ring-stage parasites synchronised in their previous erythrocytic cycle by treatment with sorbitol. Cooled saponin lysed parasites were resuspended in TRIzol and RNA harvested as per manufacturers recommendations (Invitrogen). RNA was further purified using RNeasy columns (Qiagen) and treated with Turbo DNase to remove any residual DNA (Ambion). RNA samples were tested for gDNA contamination by PCR. First strand cDNA synthesis took place using a Retroscript kit (Ambion) or Superscript III Reverse Transcriptase III (Invitrogen). Extraction of gDNA was performed at trophozoite stage. qPCR primers used for analysing expression of the 3D7 var repertoire was repeated as previously described [46]. cDNA quantification was performed on a Roche LC480 light cycler. Transcript quantification was performed by comparing to standard dilution series of gDNA for all var genes and then normalising for cDNA concentration with the expression level of actin (PFL2215w). In experiments describing the expression of pUpsB/C plasmids hDHFR was normalised against Msp8 (another ring-stage expressed gene) and hsp86 was used to normalise bsd expression (bsd is driven by the hsp86 promoter).
Plasmid copy number was quantified using the level of bsd versus the signal from the single copy gene hsp86. hdhfr transcript levels were normalised against the expression of the ring-expressed msp8 gene and then divided by the plasmid copy number calculated previously. ΔPfSir2 parasites also contain hDHFR as a selectable marker for gene disruption therefore the hdhfr transcript level from these parasites without pUpsB/C was subtracted from parasites harbouring these plasmids as to normalise for the hdhfr signal from the UpsB-driven hdhfr alone. bsd transcript levels were normalised against the expression of hsp86 and further by taking into account plasmid copy number. Standard error was calculated across the three biological replicates and used to derive confidence intervals. All expression values were normalised against wildtype parasites with an active UpsB/C-hdhfr gene as a measure of the suppression status of all other samples.
Supporting Information
Alignment of the Catalytic Domain of Sir2A and B Paralogues with Other Sirtuins
Shading level denotes level of conservation within aligned sequences. Boxes 1, 2, and 3 form the “Rossman fold” and are the binding pocket for NAD+ . Residues that are considered critical in Hst2 for the catalytic activity of the NAD+ binding pocket are marked with “*” and these are completely conserved in PfSir2B. “#” marks the cystine residues that form a triad for binding zinc ions and these are completely conserved in PfSir2B. Boxes 4 and 5 denote the region that binds the acetylated substrate and this shows diversity between PfSir2A and PfSir2B and other sirtuin members. “*” residues highlighted in boxes 4 and 5 are those that make contact with the acetylated substrate in Hst2 and these also show differences. Hs, Homo sapiens; Pf, P. falciparum; Pv, P. vivax; Sc, Saccharomyces cerevisiae.
(1.11 MB TIF)
Genomic Characterisation of PfSir2 Mutants
(A) Pulse field gel and southern blot using plasmid backbone as a probe. Membrane shows integration of plasmid into two high molecule weight chromosomes consistent with Chromosomes 13 and 14 and proves the identity of these parasites for further analysis.
(B) Rep20 fingerprinting of parasite lines signifying that they are all 3D7. Var genes and subtelomeric ends are highly polymorphic between strains and thus Rep20 fingerprinting provides evidence that comparisons of var (and other genes) within subtelomeric regions is comparable between all these cell lines. Arrow points to an RFLP between ΔPfSir2A and 3D7.
(905 KB TIF)
Physical Map of Subtelomeric Gene Expression in Sir2-Disrupted Lines
Physical map of subtelomeric and colour-coded expression of each gene denotes expression level and Sir2-disrupted line it is expressed in. Subtelomeric ends of each of the 14 P. falciparum chromosomes are accompanied by graphs of the expression of subtelomeric genes. Expression profile of every biological replicate for each individual gene across the three intraerythrocytic time points is shown as a dot while the average expression across the replicates is shown as a line: red, ΔPfSir2A; blue, ΔPfSir2B; and grey, wildtype 3D7 parasites.
(3.73 MB PDF)
Raw Expression Values of var Genes in 3D7 and Sir2-Disrupted Parasites
Var genes were grouped according to the conserved promoter type that drives their expression and plotted on a graph as a log2 values. Expression values of var genes in 3D7 are in green, ΔPfSir2A in red, and ΔPfSir2B in blue. Highly statistically different values from 3D7 are marked with a “*.” As seen with Gaussian self-clustering disruption of PfSir2A has a greater effect on the derepression of UpsA and UpsC-driven var genes suggesting a role for this molecule in their suppression, whereas UpsB var gene suppression is controlled by PfSir2B. This Affymetrix expression data is corroborated by qPCR expression values outlined in Table S2.
(576 KB TIF)
Analysis of var Genes by qPCR on CSA Selected Parasite Lines
In standard culture conditions 3D7 parasites express a standard set of var genes and largely do not express var2CSA. Upon selection of infected red blood cells to bind to CSA parasites expressing var2CSA are highly enriched and as a consequence of mutual exclusion the expression of all other var genes in these parasites are suppressed as they drop to near basal levels.
(501 KB TIF)
Log2 Ratio of All Genes Differentially Expressed at Any Stages in ΔSir2A and ΔSir2B Parasite Lines
(54 KB XLS)
Expression of 3D7 var Repertoire in PfSir2A, PfSir2B, and Wildtype Parasites before and after Selection on CSA
(68 KB XLS)
(52 KB DOC)
Acknowledgments
We would like to thank Anu Sakthianandeswaren and Dania Mueller for help with qPCR analysis and Jenny Thompson for excellent technical support. We thank donors and Red Cross Blood Service (Melbourne, Australia) for supply of erythrocytes and serum.
Abbreviations
- bsd
blasticidin-S deaminase
- CSA
chondroitin sulphate A
- FACS
flow cytometry
- PfEMP1
Plasmodium falciparum erythrocyte membrane protein 1
- PfSir2A
P. falciparum silent information regulator A
- PfSir2B
P. falciparum silent information regulator B
- qPCR
quantitative PCR
- sir
silencing information regulator
- sir2
silent information regulator 2
Footnotes
¤a Current address: Instituto de Medicina Molecular, Lisboa, Portugal
¤b Current address: Harvard School of Public Health, Boston, Massachusetts, United States of America
Author contributions. CJT, CKC, MTD, TSV, SAR, MH, MFD, LMdS, AS, AI, TPS, JGB, and AFC worked closely together to design, perform, and interpret experiments. CJT and AFC wrote the paper.
Funding. CJT is supported by a National Health and Medical Research Council (NHMRC) Peter Doherty Postdoctoral Research Fellowship. MTD was funded by a Wellcome Trust Advanced Training Fellowship. TSV was supported by a Swiss National Science Foundation fellowship. AFC is an NHMRC Australia Fellow and a Howard Hughes Medical Institute International Fellow. This work was supported by the Wellcome Trust and the NHMRC of Australia.
Competing interests. The authors have declared that no competing interests exist.
References
- Merrick CJ, Duraisingh MT. Heterochromatin-mediated control of virulence gene expression. Mol Microbiol. 2006;62:612–620. doi: 10.1111/j.1365-2958.2006.05397.x. [DOI] [PubMed] [Google Scholar]
- Deitsch KW, Moxon ER, Wellems TE. Shared themes of antigenic variation and virulence in bacterial, protozoal, and fungal Infections. Micro Mol Biol Reviews. 1997;61:281–293. doi: 10.1128/mmbr.61.3.281-293.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller LH, Baruch DI, Marsh K, Doumbo OK. The pathogenic basis of malaria. Nature. 2002;415:673–679. doi: 10.1038/415673a. [DOI] [PubMed] [Google Scholar]
- Bull PC, Lowe BS, Kortok M, Molyneux CS, Newbold CI, et al. Parasite antigens on the infected red cell surface are targets for naturally acquired immunity to malaria. Nature Med. 1998;4:358–360. doi: 10.1038/nm0398-358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snow RW, Guerra CA, Noor AM, Myint HY, Hay SI. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature. 2005;434:214–217. doi: 10.1038/nature03342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su XZ, Heatwole VM, Wertheimer SP, Guinet F, Herrfeldt JA, et al. The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infected erythrocytes. Cell. 1995;82:89–100. doi: 10.1016/0092-8674(95)90055-1. [DOI] [PubMed] [Google Scholar]
- Smith JD, Chitnis CE, Craig AG, Roberts DJ, Hudson-Taylor DE, et al. Switches in expression of Plasmodium falciparum var genes correlate with changes in antigenic and cytoadherent phenotypes of infected erythrocytes. Cell. 1995;82:101–110. doi: 10.1016/0092-8674(95)90056-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baruch DI, Pasloske BL, Singh HB, Bi X, Ma XC, et al. Cloning the P.falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. doi: 10.1016/0092-8674(95)90054-3. [DOI] [PubMed] [Google Scholar]
- Kyes S, Horrocks P, Newbold C. Antigenic variation at the infected red cell surface in malaria. Annu Rev Microbiol. 2001;55:673–707. doi: 10.1146/annurev.micro.55.1.673. [DOI] [PubMed] [Google Scholar]
- Gardner MJ, Hall N, Fung E, White O, Berriman M, et al. Genome sequence of the human malaria parasite Plasmodium falciparum . Nature. 2002;419:498–511. doi: 10.1038/nature01097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ralph SA, Scheidig-Benatar C, Scherf A. Antigenic variation in Plasmodium falciparum is associated with movement of var loci between subnuclear locations. Proc Natl Acad Sci U S A. 2005;102:5414–5419. doi: 10.1073/pnas.0408883102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyes S, Christodoulou Z, Pinches R, Kriek N, Horrocks P, et al. Plasmodium falciparum var gene expression is developmentally controlled at the level of RNA polymerase II-mediated transcription initiation. Mol Microbiol. 2007;63:1237–1247. doi: 10.1111/j.1365-2958.2007.05587.x. [DOI] [PubMed] [Google Scholar]
- Scherf A, Hernandez-Rivas R, Buffet P, Bottius E, Benatar C, et al. Antigenic variation in malaria: in situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocytic development in Plasmodium falciparum . EMBO J. 1998;17:5418–5426. doi: 10.1093/emboj/17.18.5418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubio JP, Thompson JK, Cowman AF. The var genes of Plasmodium falciparum are located in the subtelomeric region of most chromosomes. EMBO J. 1996;15:4069–4077. [PMC free article] [PubMed] [Google Scholar]
- Freitas-Junior LH, Bottius E, Pirrit LA, Deitsch KW, Scheidig C, et al. Frequent ectopic recombination of virulence factor genes in telomeric chromosome clusters of P. falciparum . Nature. 2000;407:1018–1022. doi: 10.1038/35039531. [DOI] [PubMed] [Google Scholar]
- Marty AJ, Thompson JK, Duffy MF, Voss TS, Cowman AF, et al. Evidence that Plasmodium falciparum chromosome end clusters are cross-linked by protein and are the sites of both virulence gene silencing and activation. Mol Microbiol. 2006;62:72–83. doi: 10.1111/j.1365-2958.2006.05364.x. [DOI] [PubMed] [Google Scholar]
- Kraemer SM, Smith JD. Evidence for the importance of genetic structuring to the structural and functional specialization of the Plasmodium falciparum var gene family. Mol Microbiol. 2003;50:1527–1538. doi: 10.1046/j.1365-2958.2003.03814.x. [DOI] [PubMed] [Google Scholar]
- Lavstsen T, Salanti A, Jensen AT, Arnot DE, Theander TG. Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J. 2003;2:27. doi: 10.1186/1475-2875-2-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howell DP, Levin EA, Springer AL, Kraemer SM, Phippard DJ, et al. Mapping a common interaction site used by Plasmodium falciparum Duffy binding-like domains to bind diverse host receptors. Mol Microbiol. 2008;67:78–87. doi: 10.1111/j.1365-2958.2007.06019.x. [DOI] [PubMed] [Google Scholar]
- Kraemer SM, Kyes SA, Aggarwal G, Springer AL, Nelson SO, et al. Patterns of gene recombination shape var gene repertoires in Plasmodium falciparum: comparisons of geographically diverse isolates. BMC Genomics. 2007;8:45. doi: 10.1186/1471-2164-8-45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Donnell RA, Freitas-Junior LH, Preiser PR, Williamson DH, Duraisingh M, et al. A genetic screen for improved plasmid segregation reveals a role for Rep20 in the interaction of Plasmodium falciparum chromosomes. EMBO J. 2002;21:1231–1239. doi: 10.1093/emboj/21.5.1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Figueiredo LM, Freitas-Junior LH, Bottius E, Olivo-Marin JC, Scherf A. A central role for Plasmodium falciparum subtelomeric regions in spatial positioning and telomere length regulation. EMBO J. 2002;21:815–824. doi: 10.1093/emboj/21.4.815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freitas-Junior LH, Hernandez-Rivas R, Ralph SA, Montiel-Condado D, Ruvalcaba-Salazar OK, et al. Telomeric heterochromatin propagation and histone acetylation control mutually exclusive expression of antigenic variation genes in malaria parasites. Cell. 2005;121:25–36. doi: 10.1016/j.cell.2005.01.037. [DOI] [PubMed] [Google Scholar]
- Duraisingh MT, Voss TS, Marty AJ, Duffy MF, Good RT, et al. Heterochromatin silencing and locus repositioning linked to regulation of virulence genes in Plasmodium falciparum . Cell. 2005;121:13–24. doi: 10.1016/j.cell.2005.01.036. [DOI] [PubMed] [Google Scholar]
- Dzikowski R, Li F, Amulic B, Eisberg A, Frank M, et al. Mechanisms underlying mutually exclusive expression of virulence genes by malaria parasites. EMBO Rep. 2007;8:959–965. doi: 10.1038/sj.embor.7401063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voss TS, Healer J, Marty AJ, Duffy MF, Thompson JK, et al. A var gene promoter controls allelic exclusion of virulence genes in Plasmodium falciparum malaria. Nature. 2006;439:1004–1008. doi: 10.1038/nature04407. [DOI] [PubMed] [Google Scholar]
- Voss TS, Tonkin CJ, Marty AJ, Thompson JK, Healer J, et al. Alterations in local chromatin environment are involved in silencing and activation of subtelomeric var genes in Plasmodium falciparum . Mol Microbiol. 2007;66:139–150. doi: 10.1111/j.1365-2958.2007.05899.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank M, Dzikowski R, Costantini D, Amulic B, Berdougo E, et al. Strict pairing of var promoters and introns is required for var gene silencing in the malaria parasite Plasmodium falciparum . J Biol Chem. 2006;281:9942–9952. doi: 10.1074/jbc.M513067200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deitsch K, Duraisingh M, Dzikowski R, Gunasekera A, Khan S, et al. Mechanisms of gene regulation in Plasmodium . Am J Trop Med Hyg. 2007;77:201–208. [PubMed] [Google Scholar]
- Voss TS, Kaestli M, Vogel D, Bopp S, Beck HP. Identification of nuclear proteins that interact differentially with Plasmodium falciparum var gene promoters. Mol Microbiol. 2003;48:1593–15607. doi: 10.1046/j.1365-2958.2003.03528.x. [DOI] [PubMed] [Google Scholar]
- Lopez-Rubio JJ, Gontijo AM, Nunes MC, Issar N, Hernandez Rivas R, et al. 5′ flanking region of var genes nucleate histone modification patterns linked to phenotypic inheritance of virulence traits in malaria parasites. Mol Microbiol. 2007;66:1296–1305. doi: 10.1111/j.1365-2958.2007.06009.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chookajorn T, Dzikowski R, Frank M, Li F, Jiwani AZ, et al. Epigenetic memory at malaria virulence genes. Proc Natl Acad Sci U S A. 2007;104:899–902. doi: 10.1073/pnas.0609084103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shore D, Squire M, Nasmyth KA. Characterization of two genes required for the position-effect control of yeast mating-type genes. EMBO J. 1984;3:2817–2823. doi: 10.1002/j.1460-2075.1984.tb02214.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moazed D. Enzymatic activities of Sir2 and chromatin silencing. Curr Opin Cell Biol. 2001;13:232–238. doi: 10.1016/s0955-0674(00)00202-7. [DOI] [PubMed] [Google Scholar]
- Michan S, Sinclair D. Sirtuins in mammals: insights into their biological function. Biochem J. 2007;404:1–13. doi: 10.1042/BJ20070140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403:795–800. doi: 10.1038/35001622. [DOI] [PubMed] [Google Scholar]
- Tanny JC, Dowd GJ, Huang J, Hilz H, Moazed D. An enzymatic activity in the yeast Sir2 protein that is essential for gene silencing. Cell. 1999;99:735–745. doi: 10.1016/s0092-8674(00)81671-2. [DOI] [PubMed] [Google Scholar]
- Merrick CJ, Duraisingh MT. Plasmodium falciparum Sir2: an unusual sirtuin with dual histone deacetylase and ADP-ribosyltransferase activity. Eukaryot Cell. 2007;6:2081–2091. doi: 10.1128/EC.00114-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mancio-Silva L, Rojas-Meza AP, Vargas M, Scherf A, Hernandez-Rivas R. Differential association of Orc1 and Sir2 proteins to telomeric domains in Plasmodium falciparum . J Cell Sci. 2008;121:2046–2053. doi: 10.1242/jcs.026427. [DOI] [PubMed] [Google Scholar]
- Frye RA. Phylogenetic classification of prokaryotic and eukaryotic Sir2-like proteins. Biochem Biophys Res Commun. 2000;273:793–798. doi: 10.1006/bbrc.2000.3000. [DOI] [PubMed] [Google Scholar]
- Duraisingh MT, Triglia T, Cowman AF. Negative selection of Plasmodium falciparum reveals targeted gene deletion by double crossover recombination. Int J Parasitol. 2002;32:81–89. doi: 10.1016/s0020-7519(01)00345-9. [DOI] [PubMed] [Google Scholar]
- Rubio JP, Triglia T, Kemp DJ, de Bruin D, Ravetch JV, et al. A YAC contig map of Plasmodium falciparum chromosome 4: characterization of a DNA amplification between two recently separated isolates. Genomics. 1995;26:192–198. doi: 10.1016/0888-7543(95)80200-6. [DOI] [PubMed] [Google Scholar]
- Marti M, Good RT, Rug M, Knuepfer E, Cowman AF. Targeting malaria virulence and remodeling proteins to the host erythrocyte. Science. 2004;306:1930–1933. doi: 10.1126/science.1102452. [DOI] [PubMed] [Google Scholar]
- Hiller NL, Bhattacharjee S, van Ooij C, Liolios K, Harrison T, et al. A host-targeting signal in virulence proteins reveals a secretome in malarial infection. Science. 2004;306:1934–1937. doi: 10.1126/science.1102737. [DOI] [PubMed] [Google Scholar]
- Horrocks P, Pinches R, Christodoulou Z, Kyes SA, Newbold CI. Variable var transition rates underlie antigenic variation in malaria. Proc Natl Acad Sci U S A. 2004;101:11129–11134. doi: 10.1073/pnas.0402347101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, et al. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
- Beeson JG, Mann EJ, Byrne TJ, Caragounis A, Elliott SR, et al. Antigenic differences and conservation among placental Plasmodium falciparum-infected erythrocytes and acquisition of variant-specific and cross-reactive antibodies. J Infect Dis. 2006;193:721–730. doi: 10.1086/500145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott SR, Duffy MF, Byrne TJ, Beeson JG, Mann EJ, et al. Cross-reactive surface epitopes on chondroitin sulfate A-adherent Plasmodium falciparum-infected erythrocytes are associated with transcription of var2csa. Infect Immun. 2005;73:2848–2856. doi: 10.1128/IAI.73.5.2848-2856.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duffy MF, Byrne TJ, Elliott SR, Wilson DW, Rogerson SJ, et al. Broad analysis reveals a consistent pattern of var gene transcription in Plasmodium falciparum repeatedly selected for a defined adhesion phenotype. Mol Microbiol. 2005;56:774–788. doi: 10.1111/j.1365-2958.2005.04577.x. [DOI] [PubMed] [Google Scholar]
- Richards EJ, Elgin SC. Epigenetic codes for heterochromatin formation and silencing: rounding up the usual suspects. Cell. 2002;108:489–500. doi: 10.1016/s0092-8674(02)00644-x. [DOI] [PubMed] [Google Scholar]
- Moazed D, Rudner AD, Huang J, Hoppe GJ, Tanny JC. A model for step-wise assembly of heterochromatin in yeast. Novartis Found Symp. 2004;259:48–56. 56–62, 163–169. discussion. [PubMed] [Google Scholar]
- Kyes SA, Rowe JA, Kriek N, Newbold CI. Rifins: a second family of clonally variant proteins expressed on the surface of red cells infected with Plasmodium falciparum . Proc Natl Acad Sci U S A. 1999;96:9333–9338. doi: 10.1073/pnas.96.16.9333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tham WH, Payne PD, Brown GV, Rogerson SJ. Identification of basic transcriptional elements required for rif gene transcription. Int J Parasitol. 2007;37:605–615. doi: 10.1016/j.ijpara.2006.11.006. [DOI] [PubMed] [Google Scholar]
- Joannin N, Abhiman S, Sonnhammer EL, Wahlgren M. Sub-grouping and sub-functionalization of the RIFIN multi-copy protein family. BMC Genomics. 2008;9:19. doi: 10.1186/1471-2164-9-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sam-Yellowe TY, Florens L, Johnson JR, Wang T, Drazba JA, et al. A Plasmodium gene family encoding Maurer's cleft membrane proteins: structural properties and expression profiling. Genome Res. 2004;14:1052–1059. doi: 10.1101/gr.2126104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sargeant TJ, Marti M, Caler E, Carlton JM, Simpson K, et al. Lineage-specific expansion of proteins exported to erythrocytes in malaria parasites. Genome Biol. 2006;7:R12. doi: 10.1186/gb-2006-7-2-r12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spielmann T, Fergusen DJ, Beck HP. etramps, a new Plasmodium falciparum gene family coding for developmentally regulated and highly charged membrane proteins located at the parasite-host cell interface. Mol Biol Cell. 2003;14:1529–1544. doi: 10.1091/mbc.E02-04-0240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hediger F, Gasser SM. Nuclear organization and silencing: putting things in their place. Nat Cell Biol. 2002;4:E53–E55. doi: 10.1038/ncb0302-e53. [DOI] [PubMed] [Google Scholar]
- Palladino F, Laroche T, Gilson E, Axelrod A, Pillus L, et al. SIR3 and SIR4 proteins are required for the positioning and integrity of yeast telomeres. Cell. 1993;75:543–555. doi: 10.1016/0092-8674(93)90388-7. [DOI] [PubMed] [Google Scholar]
- Ralph SA, Scherf A. The epigenetic control of antigenic variation in Plasmodium falciparum . Curr Opin Microbiol. 2005;8:434–440. doi: 10.1016/j.mib.2005.06.007. [DOI] [PubMed] [Google Scholar]
- Tham WH, Zakian VA. Transcriptional silencing at Saccharomyces telomeres: implications for other organisms. Oncogene. 2002;21:512–521. doi: 10.1038/sj.onc.1205078. [DOI] [PubMed] [Google Scholar]
- Millar CB, Grunstein M. Genome-wide patterns of histone modifications in yeast. Nat Rev Mol Cell Biol. 2006;7:657–666. doi: 10.1038/nrm1986. [DOI] [PubMed] [Google Scholar]
- Zhao K, Chai X, Clements A, Marmorstein R. Structure and autoregulation of the yeast Hst2 homolog of Sir2. Nat Struct Biol. 2003;10:864–871. doi: 10.1038/nsb978. [DOI] [PubMed] [Google Scholar]
- Pisano S, Galati A, Cacchione S. Pisano S, Galati A, Cacchione S (2008) Telomeric nucleosomes: forgotten players at chromosome ends; 2008. Cell Mol Life Sci. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Figueiredo LM, Pirrit LA, Scherf A. Genomic organisation and chromatin structure of Plasmodium falciparum chromosome ends. Mol Biochem Parasitol. 2000;106:169–174. doi: 10.1016/s0166-6851(99)00199-1. [DOI] [PubMed] [Google Scholar]
- Figueiredo L, Scherf A. Plasmodium telomeres and telomerase: the usual actors in an unusual scenario. Chromosome Res. 2005;13:517–524. doi: 10.1007/s10577-005-0996-3. [DOI] [PubMed] [Google Scholar]
- Figueiredo LM, Rocha EP, Mancio-Silva L, Prevost C, Hernandez-Verdun D, et al. The unusually large Plasmodium telomerase reverse-transcriptase localizes in a discrete compartment associated with the nucleolus. Nucleic Acids Res. 2005;33:1111–1122. doi: 10.1093/nar/gki260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharyya MK, Lustig AJ. Telomere dynamics in genome stability. Trends Biochem Sci. 2006;31:114–122. doi: 10.1016/j.tibs.2005.12.001. [DOI] [PubMed] [Google Scholar]
- Jensen AT, Magistrado P, Sharp S, Joergensen L, Lavstsen T, et al. Plasmodium falciparum associated with severe childhood malaria preferentially expresses PfEMP1 encoded by group A var genes. J Exp Med. 2004;199:1179–1190. doi: 10.1084/jem.20040274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyriacou HM, Stone GN, Challis RJ, Raza A, Lyke KE, et al. Differential var gene transcription in Plasmodium falciparum isolates from patients with cerebral malaria compared to hyperparasitaemia. Mol Biochem Parasitol. 2006;150:211–218. doi: 10.1016/j.molbiopara.2006.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felsenstein J. Inferring phylogenies from protein sequences by parsimony, distance, and likelihood methods. Methods Enzymol. 1996;266:418–427. doi: 10.1016/s0076-6879(96)66026-1. [DOI] [PubMed] [Google Scholar]
- Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. 2003;52:696–704. doi: 10.1080/10635150390235520. [DOI] [PubMed] [Google Scholar]
- Wu Y, Sifri CD, Lei H-H, Su X-S, Wellems TE. Transfection of Plasmodium falciparum within human red blood cells. Proc Natl Acad Sci U S A. 1995;92:973–977. doi: 10.1073/pnas.92.4.973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crabb BS, Cowman AF. Characterization of promoters and stable transfection by homologous and nonhomologous recombination in Plasmodium falciparum . Proc Natl Acad Sci U S A. 1996;93:7289–7294. doi: 10.1073/pnas.93.14.7289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyes S, Pinches R, Newbold C. A simple RNA analysis method shows var and rif multigene family expression patterns in Plasmodium falciparum . Mol Biochem Parasitol. 2000;105:311–315. doi: 10.1016/s0166-6851(99)00193-0. [DOI] [PubMed] [Google Scholar]
- Cortes A, Carret C, Kaneko O, Yim Lim BY, Ivens A, et al. Epigenetic silencing of Plasmodium falciparum genes linked to erythrocyte invasion. PLoS Pathog. 2007;3:e107. doi: 10.1371/journal.ppat.0030107. doi: 10.1371/journal.ppat.0030107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentleman RC, Carey VJ, Bates DM, Bolstad B, Dettling M, et al. Bioconductor: open software development for computational biology and bioinformatics. Genome Biol. 2004;5:R80. doi: 10.1186/gb-2004-5-10-r80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gautier L, Cope L, Bolstad BM, Irizarry RA. affy–analysis of Affymetrix GeneChip data at the probe level. Bioinformatics. 2004;20:307–315. doi: 10.1093/bioinformatics/btg405. [DOI] [PubMed] [Google Scholar]
- Irizarry RA, Bolstad BM, Collin F, Cope LM, Hobbs B, et al. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003;31:e15. doi: 10.1093/nar/gng015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waterkeyn JG, Cowman AF, Cooke BM. Plasmodium falciparum: gelatin enrichment selects for parasites with full-length chromosome 2. implications for cytoadhesion assays. Exp Parasitol. 2001;97:115–118. doi: 10.1006/expr.2000.4593. [DOI] [PubMed] [Google Scholar]
- Beeson JG, Rogerson SJ, Cooke BM, Reeder JC, Chai W, et al. Adhesion of Plasmodium falciparum-infected erythrocytes to hyaluronic acid in placental malaria. Nature Med. 2000;6:86–90. doi: 10.1038/71582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michishita E, McCord RA, Berber E, Kioi M, Padilla-Nash H, et al. SIRT6 is a histone H3 lysine 9 deacetylase that modulates telomeric chromatin. Nature. 2008;452:492–496. doi: 10.1038/nature06736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mostoslavsky R, Chua KF, Lombard DB, Pang WW, Fischer MR, et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell. 2006;124:315–329. doi: 10.1016/j.cell.2005.11.044. [DOI] [PubMed] [Google Scholar]
- Ford E, Voit R, Liszt G, Magin C, Grummt I, et al. Mammalian Sir2 homolog SIRT7 is an activator of RNA polymerase I transcription. Genes Dev. 2006;20:1075–1080. doi: 10.1101/gad.1399706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlton JM, Adams JH, Silva JC, Bidwell SL, Lorenzi H, et al. Comparative genomics of the neglected human malaria parasite Plasmodium vivax . Nature. 2008;455:757–763. doi: 10.1038/nature07327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall N, Karras M, Raine JD, Carlton JM, Kooij TW, et al. A comprehensive survey of the Plasmodium life cycle by genomic, transcriptomic, and proteomic analyses. Science. 2005;307:82–86. doi: 10.1126/science.1103717. [DOI] [PubMed] [Google Scholar]
- Pain A, Renauld H, Berriman M, Murphy L, Yeats CA, et al. Genome of the host-cell transforming parasite Theileria annulata compared with T. parva . Science. 2005;309:131–133. doi: 10.1126/science.1110418. [DOI] [PubMed] [Google Scholar]
- Xu P, Widmer G, Wang Y, Ozaki LS, Alves JM, et al. The genome of Cryptosporidium hominis . Nature. 2004;431:1107–1112. doi: 10.1038/nature02977. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Alignment of the Catalytic Domain of Sir2A and B Paralogues with Other Sirtuins
Shading level denotes level of conservation within aligned sequences. Boxes 1, 2, and 3 form the “Rossman fold” and are the binding pocket for NAD+ . Residues that are considered critical in Hst2 for the catalytic activity of the NAD+ binding pocket are marked with “*” and these are completely conserved in PfSir2B. “#” marks the cystine residues that form a triad for binding zinc ions and these are completely conserved in PfSir2B. Boxes 4 and 5 denote the region that binds the acetylated substrate and this shows diversity between PfSir2A and PfSir2B and other sirtuin members. “*” residues highlighted in boxes 4 and 5 are those that make contact with the acetylated substrate in Hst2 and these also show differences. Hs, Homo sapiens; Pf, P. falciparum; Pv, P. vivax; Sc, Saccharomyces cerevisiae.
(1.11 MB TIF)
Genomic Characterisation of PfSir2 Mutants
(A) Pulse field gel and southern blot using plasmid backbone as a probe. Membrane shows integration of plasmid into two high molecule weight chromosomes consistent with Chromosomes 13 and 14 and proves the identity of these parasites for further analysis.
(B) Rep20 fingerprinting of parasite lines signifying that they are all 3D7. Var genes and subtelomeric ends are highly polymorphic between strains and thus Rep20 fingerprinting provides evidence that comparisons of var (and other genes) within subtelomeric regions is comparable between all these cell lines. Arrow points to an RFLP between ΔPfSir2A and 3D7.
(905 KB TIF)
Physical Map of Subtelomeric Gene Expression in Sir2-Disrupted Lines
Physical map of subtelomeric and colour-coded expression of each gene denotes expression level and Sir2-disrupted line it is expressed in. Subtelomeric ends of each of the 14 P. falciparum chromosomes are accompanied by graphs of the expression of subtelomeric genes. Expression profile of every biological replicate for each individual gene across the three intraerythrocytic time points is shown as a dot while the average expression across the replicates is shown as a line: red, ΔPfSir2A; blue, ΔPfSir2B; and grey, wildtype 3D7 parasites.
(3.73 MB PDF)
Raw Expression Values of var Genes in 3D7 and Sir2-Disrupted Parasites
Var genes were grouped according to the conserved promoter type that drives their expression and plotted on a graph as a log2 values. Expression values of var genes in 3D7 are in green, ΔPfSir2A in red, and ΔPfSir2B in blue. Highly statistically different values from 3D7 are marked with a “*.” As seen with Gaussian self-clustering disruption of PfSir2A has a greater effect on the derepression of UpsA and UpsC-driven var genes suggesting a role for this molecule in their suppression, whereas UpsB var gene suppression is controlled by PfSir2B. This Affymetrix expression data is corroborated by qPCR expression values outlined in Table S2.
(576 KB TIF)
Analysis of var Genes by qPCR on CSA Selected Parasite Lines
In standard culture conditions 3D7 parasites express a standard set of var genes and largely do not express var2CSA. Upon selection of infected red blood cells to bind to CSA parasites expressing var2CSA are highly enriched and as a consequence of mutual exclusion the expression of all other var genes in these parasites are suppressed as they drop to near basal levels.
(501 KB TIF)
Log2 Ratio of All Genes Differentially Expressed at Any Stages in ΔSir2A and ΔSir2B Parasite Lines
(54 KB XLS)
Expression of 3D7 var Repertoire in PfSir2A, PfSir2B, and Wildtype Parasites before and after Selection on CSA
(68 KB XLS)
(52 KB DOC)