

Summary
The microtubule (MT) cytoskeleton is reorganized during myogenesis as individual myoblasts fuse into multinucleated myotubes. Although this reorganization has long been observed in cell culture, these findings have not been validated during development, and proteins that regulate this process are largely unknown. We have identified a novel postmitotic function for the cytokinesis proteins RacGAP50C (Tumbleweed) and Pavarotti as essential regulators of MT organization during Drosophila myogenesis. We show that the localization of the MT nucleator γ-tubulin changes from diffuse cytoplasmic staining in mononucleated myoblasts to discrete cytoplasmic puncta at the nuclear periphery in multinucleated myoblasts, and that this change in localization depends on RacGAP50C. RacGAP50C and γ-tubulin colocalize at perinuclear sites in myotubes, and in RacGAP50C mutants γ-tubulin remains dispersed throughout the cytoplasm. Furthermore, we show that the mislocalization of RacGAP50C in pavarotti mutants is sufficient to redistribute γ-tubulin to the muscle fiber ends. Finally, myotubes in RacGAP50C mutants have MTs with non-uniform polarity, resulting in multiple guidance errors. Taken together, these findings provide strong evidence that the reorganization of the MT network that has been observed in vitro plays an important role in myotube extension and muscle patterning in vivo, and also identify two molecules crucial for this process.
Keywords: Drosophila, Pavarotti, RacGAP50C, Gamma-tubulin, Microtubule, Myotube guidance
INTRODUCTION
Muscle differentiation is a multi-step process that involves myoblast specification, fusion, bidirectional elongation of the myotube and subsequent attachment of the muscle fiber. These processes are well conserved and are observed in cultured myocytes from diverse organisms including human, mouse and Drosophila (Bugnard et al., 2005; Cottam et al., 2006; Musa et al., 2003; Pizon et al., 2005; Tassin et al., 1985). However, little is known about the molecular mechanisms that drive these morphological changes during muscle development in vivo. The formation of the Drosophila embryonic somatic musculature provides a tractable system for the study of muscle development. The somatic musculature forms a repeated pattern of thirty muscles per abdominal hemisegment (see Fig. 1C). The development of these muscles requires the specification of founder cells (FCs) that express a transcriptional profile coding for each muscle identity, and fusion-competent myoblasts (FCMs) that add to muscle size by fusing to the FCs and adopting the muscle identity of the FC to which they fuse (Dohrmann et al., 1990; Rushton et al., 1995). As they fuse, the myotubes elongate along a linear axis towards specific sites of attachment in the epidermis called tendon cells (Volk, 1999). Although many of the molecular mechanisms underlying FC specification and myoblast fusion are well understood (Baylies and Michelson, 2001; Frasch, 1999; Richardson et al., 2008), less is known about the molecules that function during the migratory phase of muscle development. Several proteins required for matching specific subsets of muscles to their tendon cells have been identified. The Derailed receptor tyrosine kinase is expressed by a small subset of muscles and is required for their ventral attachments (Callahan et al., 1996). The extracellular matrix molecule Slit has been identified as an attractive guidance cue for a subset of muscles expressing the Roundabout receptor (Kramer et al., 2001). Additional studies have shown that Grip, Echinoid and Kon-Tiki (Perdido) also play a role in targeting subsets of muscle fibers to their attachments (Estrada et al., 2007; Schnorrer et al., 2007; Swan et al., 2006; Swan et al., 2004). However, although these molecules have given insight into how cell-surface and extracellular guidance molecules influence muscle attachment site (MAS) selection, little is known about the cytoskeletal changes that occur inside the myotube to allow for bidirectional myotube extension.
Fig. 1.

Drosophila RacGAP mutants have defects in somatic muscle patterning. Stage 16 embryos stained for Muscle myosin. Muscle patterning is shown at low (A,B) and high (D-I) magnification in wild-type (wt) (A,D,F,H) or RacGAP50CDH15 (B,E,G,I) embryos. (A) Muscle pattern as observed in the whole embryo. (B) No significant loss of muscle tissue or unfused myoblasts are observed in RacGAP50CDH15 mutants. (D,E) Arrows in D mark normal attachments of DO1. In RacGAP50CDH15, DO1 is in the incorrect position (E, arrows). (F,G) Arrowheads in F mark muscle 22 (LT2). Arrowheads in G mark an LT muscle of an abnormal shape. The arrow marks a rounded muscle that fails to migrate. (H,I) Arrowheads in H mark normal attachments for VO muscles. In RacGAP50CDH15 mutants, these muscles fail to fully extend (I, arrowheads). (C) Schematic of the wild-type muscle pattern of a single abdominal hemisegment, showing the names and numbers for muscles as referred to throughout this work. DA, dorsal acute; DO, dorsal oblique; LT, lateral transverse; VL, ventral lateral; VO, ventral oblique.
The cytoskeleton undergoes dramatic changes during the transition from the myoblast fusion phase of muscle development to the bidirectional elongation of the multinucleated myotube. Studies using mammalian muscle cell culture have shown that as proliferating myoblasts fuse into myotubes, the microtubule (MT) network changes from a radial, centrosomal array in individual myoblasts to a non-centrosomal array in multinucleated myotubes. This non-centrosomal MT array is based at the nuclear periphery and extends out in the longitudinal axis of the myotube (Bugnard et al., 2005; Musa et al., 2003; Pizon et al., 2005; Tassin et al., 1985) (reviewed by Bartolini and Gundersen, 2006). In addition, studies in cultured myotubes show that γ-tubulin, a minus-end anchoring protein essential for MT nucleation at the centrosome (Gunawardane et al., 2000; Stearns et al., 1991), may also be able to promote nucleation from non-centrosomal sites along the nuclear membrane and throughout the cytoplasm (Bugnard et al., 2005; Cottam et al., 2006; Musa et al., 2003). Although the reorganization of the MT cytoskeleton during muscle differentiation has been well described in cell culture, little is known about the role of this reorganization during muscle development. Furthermore, the mechanisms underlying the redistribution of γ-tubulin to the nuclear membrane following myoblast fusion remain to be addressed.
We identified RacGAP50C (RacGAP; also known as tumbleweed) as an essential component of the muscle migratory machinery in a screen for genes involved in somatic muscle patterning. RacGAP is conserved in C. elegans, Drosophila and humans (Jantsch-Plunger et al., 2000; Sotillos and Campuzano, 2000; Toure et al., 1998), and has been well studied for its function in cytokinesis, where it is required to coordinate anti-parallel central spindle MTs with the actomyosin cleavage furrow (D'Avino et al., 2006; Jantsch-Plunger et al., 2000; Mishima et al., 2002; Somers and Saint, 2003; Zavortink et al., 2005). RacGAP is localized at the cleavage furrow through binding to the Pavarotti kinesin-like protein (Pav), a plus-end MT motor, and this interaction also serves to bundle spindle MTs (Adams et al., 1998; Mishima et al., 2002; Somers and Saint, 2003). In addition to Pav, RacGAP binds to Anillin (Scraps - FlyBase), an actin- and myosin-binding protein (D'Avino et al., 2008; Field and Alberts, 1995; Gregory et al., 2008; Straight et al., 2005), and Pebble, a RhoGEF (Hime and Saint, 1992; Somers and Saint, 2003). In this way, RacGAP acts as an adaptor that coordinates the central spindle and actomyosin contractile ring for cleavage furrow formation and cell division (D'Avino et al., 2006; Zavortink et al., 2005).
Here, we describe a novel role for RacGAP and Pav in organizing the MT network during Drosophila myogenesis. Embryos mutant for either RacGAP or pav have similar defects in somatic muscle patterning, whereby muscle fibers are abnormally shaped and display a variety of guidance errors. RacGAP localizes to discrete cytoplasmic puncta concentrated at the nuclear periphery in multinucleated myotubes and this localization is dependent on the presence of Pav. The RacGAP-containing cytoplasmic puncta co-stain with the MT nucleating protein γ-tubulin, and γ-tubulin localization depends on the presence of RacGAP, suggesting that the function of RacGAP is to promote non-centrosomal MT organization at the nuclear periphery through an interaction with γ-tubulin. Consistent with this hypothesis, myotubes in RacGAP mutants show non-uniform polarity within the MT array. Our study provides strong evidence that the reorganization of the MT network that has been observed in vitro plays an important role in muscle patterning in vivo, and also identifies two molecules that are crucial for this process.
MATERIALS AND METHODS
Screening and mapping of the E414 mutant line
The E414 mutant line was obtained from a standard EMS screen of the second chromosome containing a CD8-sh-GFP insertion (Parnas et al., 2001). We discarded lines with general patterning defects or unfused myoblasts to avoid mutations in genes involved in muscle specification or fusion. Screening was performed at stage 16, prior to contraction (Pereanu et al., 2007), to avoid mutations in genes involved with attachment stabilization. For mapping details, see Fig. S1 in the supplementary material.
Drosophila strains and genetics
w1118 was used as wild type. The enhancer trap P1618 (sr-lacZ, gift from M. Baylies, Sloan-Kettering Institute, NY, USA) labels all epidermal tendon cells. 5053A-GAL4/TM6B,Tb+ (BL-2702) and UAS-lacZ.NZ (BL-3956) were used to label muscle 12 (VL1) nuclei. Rescue experiments were carried out using epidermal drivers 69B-GAL4 (BL-1774) and prd-GAL4/TM3,Sb1 (BL-1947), or muscle driver G14-GAL4/CyOWgβgal (Shishido et al., 1998) with wild-type RacGAP, UAS-tum::myc [gift from Robert Saint (Zavortink et al., 2005)]. G14-GAL4,RacGAPDH15/CyOWgβgal flies were created using standard recombination. Mef2-GAL4 or G14-GAL4 drivers were crossed to w1118;UAS-GFP::pav [gift from David M. Glover (Minestrini et al., 2003)] to show Pav localization in muscles of wild type or RacGAPDH15 mutants. G14-GAL4 was used to drive UAS-nod::GFP (BL-9282 and BL-9283) to label MT minus ends. Loss-of-function (LOF) alleles used were scraps8,cn1,bw1,sp1/CyO (BL-4286) and y1w1;64A-lacZ,pavB200,th1,st1,cu1,sr1,es,ca1/TM6B,iab-2-lacZ,Tb1 (BL-4384).
Immunohistochemistry and imaging
Embryo collection and fixation were carried out as described (Patel, 1994). For Phalloidin labeling, embryos were fixed for 6 minutes in 33% formaldehyde and 10% EGTA, and de-vitellinized by hand. Antibodies were used as follows: mouse anti-Muscle myosin [1:20, MAb FMM5C8 (Kramer et al., 2001)], rabbit anti-β-galactosidase (1:1000, MP Biomedicals), mouse anti-αPS2 integrin, mouse anti-Even skipped and mouse anti-α-Spectrin (1:10) (all from the Developmental Studies Hybridoma Bank), guinea pig anti-Runt (1:2000, preabsorbed, gift from J. Reinitz, Stony Brook University, Stony Brook, NY, USA), rabbit anti-Vestigial (1:50, gift from S. Carroll, HHMI, University of Wisconsin, Madison, WI, USA), rabbit anti-RacGAP50C (1:500, gift from D. Glover, University of Cambridge, Cambridge, UK), rat anti-RacGAP50C (1:500, gift from R. Saint, Australian National University, Canberra, ACT, Australia), rabbit anti-β3-tubulin (1:5000, gift from D. Buttgereit, Philipps-Universitat Marburg, Marburg, Germany), mouse anti-Roundabout (1:10), mouse anti-γ-tubulin (1:500, Sigma, clone GTU-88), rabbit anti-Mef2 (1:2000, gift from B. Paterson, National Cancer Institute, NIH, Bethesda, MD, USA), Alexa Fluor 555-conjugated rabbit anti-GFP (1:250), Alexa Fluor 488-conjugated Phalloidin (1:500), Alexa Fluor 488-conjugated goat anti-rabbit, -mouse and -guinea pig (1:500), Alexa Fluor 555-conjugated goat anti-rabbit and -mouse (1:500) (all Alexa Fluors from Molecular Probes), and Cy3-conjugated goat anti-mouse (1:500, Jackson Laboratories).
Embryos were cleared in 50% or 60% glycerol for whole-mount or fillets. Fillets were prepared for Roundabout, RacGAP50C and γ-tubulin stainings using tungsten needles. Confocal z-sections were collected using an Olympus IX81 inverted microscope with a CARV Nipkow disc confocal unit and 20× phase (0.40NA), 40× water immersion (1.15NA) and 60× water immersion (1.20NA) objectives. Images were collected with SensiCam QE or Hamamatsu Orca EM-CCD cameras. To improve the γ-tubulin signal, the wild-type, pavB200 and RacGAPDH15 embryos in Fig. 8D,D′,E,E′,F,F′,G were imaged with increased gain using electron multiplication on the Hamamatsu Orca EM-CCD camera. Imaging analysis and processing were performed using IPLab software (SensiCam QE) or iVision software (Hamamatsu Orca EM-CCD) and Photoshop CS2 (Adobe).
Fig. 8.

RacGAP controls MT polarity through interaction with γ-tubulin. Yellow arrows mark segment borders, where myotubes from adjacent segments make attachments. (A,B) In wild type, γ-tubulin (A) and RacGAP (B) have a similar localization pattern in cytoplasmic puncta that are concentrated towards the center of the myotube. (C,C′) Merge of RacGAP (red) and γ-tubulin (green) shows that γ-tubulin colocalizes with RacGAP puncta at the nuclear periphery. (D-G) Gain was increased for γ-tubulin imaging in D,D′,E,E′,F,F′,G. The segment borders indicated by the yellow arrows in D, E and F are magnified in D′, E′ and F′, respectively. D″, E″ and F″ are schematics of γ-tubulin localization within extending myotubes at a segment border in the wild-type, pav and RacGAP backgrounds, respectively. In wild-type mononucleated myoblasts (G), γ-tubulin (green) is present diffusely throughout the cytoplasm and is absent from the nucleus (magenta, labeled with anti-Mef2). In wild-type myotubes (D,D′), γ-tubulin is found in the cytoplasm (D), with puncta concentrated at the nuclear periphery (D′, arrowheads), but is absent from the muscle ends (yellow arrow). In pavB200 mutants (E,E′), γ-tubulin is not as highly concentrated around nuclei (E′, n), but is localized to patches at muscle ends (E′, arrowhead). In RacGAPDH15 mutants (F,F′), γ-tubulin fails to localize to perinuclear sites and instead is diffusely localized throughout the muscle cytoplasm, similar to its pattern in mononucleated myoblasts (G).
RESULTS
Drosophila RacGAP mutants have defects in somatic muscle morphology and patterning
To identify proteins involved in somatic MAS selection, we screened the second chromosome for mutants that showed somatic muscle defects in late stage embryos (see Materials and methods). From this screen, we identified a mutant line that we refer to as E414. Embryos homozygous for E414 have muscle-patterning defects at stage 16 as visualized by staining with an antibody against Drosophila Muscle myosin (see Fig. S1B,B′ in the supplementary material). We observed muscle defects in 93% (n=46) of E414-/- embryos, and these defects were present in all segments. We used recombination and deficiency mapping to map the mutation in E414 to the RacGAP locus (see Fig. S1D-F in the supplementary material). Because we have not been able to determine the molecular nature of the RacGAP mutation in E414, we have chosen to use the well-characterized LOF allele RacGAPDH15 (Jones and Bejsovec, 2005) for all subsequent experiments in this study.
To understand the role that RacGAP plays in muscle patterning, we examined in detail the muscle phenotype in the wild type (Fig. 1A,D,F,H) and in RacGAPDH15 mutants (Fig. 1B,E,G,I). In RacGAPDH15 homozygous embryos, we observed significant defects in muscle morphology and MAS selection. Muscles were often curved compared with wild type. In addition, we observed myotubes with ectopic extensions that often attached to the wrong tendon cell or failed to extend towards their attachment sites. These defects were not limited to a specific subset of muscles, but rather could be seen throughout the dorsal (Fig. 1E), lateral (Fig. 1G) and ventral (Fig. 1I) body wall. These results suggest that RacGAP plays an important role in the development of the embryonic somatic musculature.
In the absence of RacGAP, muscles stably attach to tendon cells
During MAS selection, migrating myotubes elongate towards specific sites of attachment in the epidermis called tendon cells, where they form stable attachments in the form of myotendinous junctions (Volk, 1999). If these attachments are not stabilized, the muscles often pull away from the tendon cells upon contraction and the resulting phenotype may resemble that seen in embryos with muscle migration defects (Brown et al., 2000; Chanana et al., 2007). To rule out the possibility that the muscle defects are due to a failure of tendon cell differentiation or of the ability of muscle fibers to form stable attachments, we examined both tendon cells and myotendinous junctions in RacGAPDH15 embryos. Tendon cells were labeled with the P1618 enhancer trap (Becker et al., 1997), which expresses lacZ under the control of the stripe (sr) promoter. In wild-type embryos, the tendon cells were mainly located along the segment borders, with a few additional tendon cell clusters in the intrasegmental region (Fig. 2A). In RacGAPDH15 embryos, the tendon cells were present, differentiated and in their proper locations (Fig. 2B). However, we often saw muscle fibers that attached to the wrong tendon cell (Fig. 2B). The myotendonous junction is marked by αPS2 integrin (Inflated - FlyBase) at the ends of all muscles at stage 16 (Devenport et al., 2007) (Fig. 2C). RacGAPDH15 mutant muscles formed stable attachments as revealed by αPS2 integrin staining (Fig. 2D). This was further supported by the observation that at the onset of muscle contraction during late stage 17a/b (Pereanu et al., 2007), the muscles remained attached (data not shown).
Fig. 2.

RacGAPDH15 mutants can form stable muscle attachments. Wild-type (A,C) or RacGAPDH15 mutant (B,D) stage 16 Drosophila embryos. To the left is a schematic of the wild-type muscle pattern (magenta) and tendon cells (green). (A,B) Muscle myosin and β-galactosidase staining of embryos with the sr-lacZ reporter labels muscles (magenta) and tendon cells (green). Tendon cells are specified in RacGAPDH15 mutants (B), but muscles often select the wrong attachment sites. Arrowheads in A mark normal attachments for LT muscles, as compared with arrowheads in B that mark muscles making inappropriate attachments in mutants. Asterisks mark tendon cells to which muscles should be attached. (C) αPS2 integrin staining in the wild-type labels muscle ends at their contact site with tendon cells. (D) RacGAPDH15 mutants make stable attachments as indicated by αPS2 integrin staining.
RacGAP is required within migrating myotubes for muscle patterning
RacGAP is uniformly expressed at low levels during late embryogenesis (Jones and Bejsovec, 2005) and it is unclear whether RacGAP is required in epidermal tendon cells for signaling to migrating myotubes, or in the muscle fiber itself to facilitate myotube extension. In addition, we and others (Jones and Bejsovec, 2005; Zavortink et al., 2005) have observed binucleate epidermal cells in RacGAP LOF embryos resulting from failures in cytokinesis (data not shown) that could indirectly lead to MAS selection defects. To identify whether RacGAP is required in tendon cells or muscles, we performed tissue-specific rescue experiments using the GAL4-UAS system (Brand and Perrimon, 1993).
The prd-GAL4 driver expresses GAL4 in stripes of the epidermis, including tendon cells, and has been used in previous studies to rescue RacGAP epidermal cytokinesis defects (Zavortink et al., 2005). We expressed UAS-tum::myc, a wild-type RacGAP transgene, in the epidermis of RacGAPDH15 homozygous embryos using the prd-GAL4 driver. Although we were able to rescue epidermal cytokinesis defects, the muscle-patterning defects remained (data not shown). The pan-epidermal 69B-GAL4 driver also failed to rescue MAS selection defects (Fig. 3C). These results argue that RacGAP is not required in the epidermis for muscle patterning, and that the binucleate tendon cells we observe in RacGAPDH15 mutants are not the primary cause of the muscle defects.
Fig. 3.

RacGAP functions cell-autonomously in muscles. Muscle myosin staining in stage 16 Drosophila embryos. Brackets mark LT muscles in two adjacent segments. (A) RacGAPDH15 homozyogotes have muscle attachment site (MAS) selection and morphology defects. Shown is a LT muscle of abnormal shape, extending past its attachment site (top right bracket). (B) G14-GAL4/+;UAS-tum::myc/+ embryo showing that overexpression of RacGAP in muscles does not cause an abnormal phenotype. (C) RacGAPDH15;69B-GAL4/UAS-tum::myc embryo showing that muscle defects are not rescued by RacGAP expression in tendon cells. (D) G14-GAL4, RacGAPDH15/+,RacGAPDH15;UAS-tum::myc/+ embryo showing that muscle defects are significantly rescued by expressing RacGAP specifically in muscles. LT muscles are now properly positioned.
Finally, we used the G14-GAL4 driver (Shishido et al., 1998) to express UAS-tum::myc in all muscle tissue beginning at stage 12. In w1118 embryos, expression of UAS-tum::myc by G14-GAL4 does not cause an overexpression phenotype (Fig. 3B). However, RacGAP expression in muscles of RacGAPDH15 homozygous embryos (Fig. 3D) significantly improved muscle morphology and rescued the muscle-patterning defects, as compared with RacGAPDH15 embryos (Fig. 3A). These results show that RacGAP functions cell-autonomously in muscles for their patterning.
Muscle specification and myoblast fusion are not impaired in RacGAP mutants
RacGAP has been well characterized with respect to its role in cytokinesis (D'Avino et al., 2006; Jantsch-Plunger et al., 2000; Mishima et al., 2002; Somers and Saint, 2003; Zavortink et al., 2005). Previous studies in RacGAP zygotic mutants have shown that cytokinesis defects in the epidermis are observed during late stage 11, when maternal contributions of RacGAP are depleted (Zavortink et al., 2005). Since the majority of mesodermal cells complete their final round of cell division around this time (Campos-Ortega and Hartenstein, 1985) and FC specification depends on proper asymmetric cell division (Carmena et al., 1998; Ruiz Gomez and Bate, 1997), we tested whether the muscle defects in RacGAP mutants could be indirectly related to a failure in early mesodermal cytokinesis and specification.
In Drosophila, each muscle fiber is formed from one muscle FC, which specifies the identity of the developing muscle, and several FCMs, which fuse with the FC and contribute to the growth of the multinucleated myotube (Bate, 1990; Baylies et al., 1998; Frasch, 1999). To assess muscle specification in RacGAPDH15 mutants, we stained with antibodies against several FC-specific markers. Even skipped (Eve) is a transcription factor that is expressed in the FC for muscle 1 (DA1) and in two pericardial cells at stage 12 (Carmena et al., 2002; Halfon et al., 2000) (Fig. 4A). In RacGAPDH15 embryos, Eve-positive cells were properly specified in all segments (Fig. 4B). We also obtained similar results when we stained for the transcription factor Runt, which labels FCs for muscles 10 (DO2), 15 (VO4) and 28 (VO3) (Carmena et al., 2002; Beckett and Baylies, 2007) (Fig. 4C). In RacGAPDH15 mutants, Runt-positive FCs were properly specified (Fig. 4D), although we occasionally saw an extra Runt-positive muscle 10 nucleus. These results provide strong evidence that FC specification is not altered in RacGAPDH15 mutants.
Fig. 4.

FC specification and myoblast fusion are not affected in RacGAP mutants. (A,B) In wild-type Drosophila embryos at stage 12, Eve is expressed in the founder cell (FC) for muscle 1 (DA1) and two pericardial cells (A). These cells are properly specified in RacGAPDH15 embryos (B). (C) Runt staining labels FCs for muscles 10 (DO2), 28 (VO3) and 15 (VO4) at stage 12. (D) In RacGAPDH15 embryos, Runt-positive FCs are properly specified, although some segments contain an extra Runt-positive muscle 10 nucleus (arrow), whereas FCs for muscles 28 and 15 are unaffected (arrowhead). (E,F) Anti-Vestigial labels a subset of FCs at stage 12 in the wild type (E), and this staining is normal in RacGAPDH15 mutants (F). (G,H) By stage 14, FCs have fused with fusion-competent myoblasts (FCMs), showing an increase in Vestigial-positive nuclei of wild-type dorsal muscles (G). Fusion also occurs in RacGAPDH15 mutants (H). (I,J) 5053A-GAL4/UAS-lacZ embryo stained for Muscle myosin to label all muscles (magenta) and for β-galactosidase to label nuclei (green) of muscle 12 (VL1). In the wild type (I), muscle 12 spans each hemisegment and contains ∼11 nuclei. In RacGAPDH15 mutants (J), muscle 12 is specified and contains the correct number of nuclei, but is mispositioned in 6% of hemisegments (n=52) (brackets). Asterisks mark intestinal cells that are also labeled with the 5053-GAL4 driver.
During myoblast fusion, FCMs assume the fate of the FC and begin to express FC markers (Dohrmann et al., 1990). To determine whether myoblast fusion was impaired in RacGAPDH15 mutants, we examined the expression of Vestigial (Vg), another FC marker (Bate and Rushton, 1993; Rushton et al., 1995; Williams et al., 1991), before and after myoblast fusion. At stage 12, expression of Vg was identical in wild-type and RacGAPDH15 mutant embryos (Fig. 4E,F), indicating that the specification of Vg-positive FCs occurs normally. Furthermore, the increase in Vg-positive nuclei in stage 14 mutant embryos showed that fusion also occurs properly (Fig. 4G,H).
Finally, we used a UAS-lacZ reporter with 5053A-GAL4, a driver specific to muscle 12 (VL1) (Jarecki et al., 1999; Schnorrer et al., 2007), to examine specification and fusion in late stage embryos. Wild-type and RacGAPDH15 embryos were stained for Muscle myosin and β-galactosidase to visualize somatic muscles and muscle 12 nuclei, respectively (Fig. 4I,J). In RacGAPDH15 embryos, muscle 12 was specified properly, although we occasionally observed mispositioning of this muscle (Fig. 4J). We also examined the number of nuclei in lacZ-positive muscles, as it has been shown that each multinucleated myotube has a predictable number of nuclei within a given range (Beckett and Baylies, 2007). In the wild type (Fig. 4I), we counted 11±1 nuclei (n=42) in muscle 12. In RacGAPDH15 embryos (Fig. 4J), we counted 9±3 (n=52), indicating no significant differences in nuclei number. Furthermore, the lack of unfused myoblasts in RacGAPDH15 mutants (Fig. 1) further demonstrated that myoblast fusion was not disrupted.
RacGAP localizes to perinuclear cytoplasmic puncta in somatic muscles
Our genetic experiments demonstrate that RacGAP functions in muscle tissue during the later stages of myogenesis. We examined RacGAP localization in multinucleated myotubes in order to gain a better understanding of its function at this time. By double staining with Muscle myosin antibody and a rabbit polyclonal against RacGAP (D'Avino et al., 2006), we found that RacGAP is localized diffusely throughout myotubes, with discrete cytoplasmic puncta that cluster at the center of each fiber (Fig. 5A-C). This is in contrast to the epidermal cells, where RacGAP was localized to the nucleus (Fig. 5G). Closer examination of muscle RacGAP staining showed that the cytoplasmic puncta were concentrated at the nuclear periphery and that this localization was visible as early as stage 14 (Fig. 5C,C′; see Fig. S2A,B in the supplementary material). This muscle RacGAP staining was weak and only detectible when the embryos were prepared as fillets (see Materials and methods).
Fig. 5.

Localization of RacGAP and Pavarotti in muscle fibers. Stage 16 Drosophila embryos. The schematic on the left shows interior muscles 6, 7, 12 and 13 (VL1-4), which are exposed by fillet preparation performed for embryos stained for RacGAP, Roundabout and γ-tubulin. (A,B,G) RacGAP (A,G) and Muscle myosin (B) staining shows RacGAP throughout all muscle fibers and in discrete cytoplasmic puncta, in contrast to RacGAP nuclear localization in the epidermis (G, arrowheads mark epidermal cells). (C,C′) Merge of RacGAP (magenta) and Muscle myosin (green) in wild type showing discrete puncta of RacGAP (C′, arrowheads) at the periphery of the nuclei (C′, n). (D-F′) MEF2-GAL4/UAS-GFP::pav embryo stained for RacGAP (D) and GFP (E) and merged (F,F′). Pav is nuclear in all muscle fibers (E-F′ green), and co-staining of RacGAP (magenta) shows an increase in RacGAP concentration (F′, arrowhead) at the nuclear periphery. (H) pavB200 mutants have a similar muscle phenotype to RacGAPDH15 (arrowheads mark muscles making inappropriate attachments). (I) RacGAP is mislocalized to the ends of muscle fibers in pavB200 mutants (arrowheads). (J) In RacGAPDH15 mutants, Pav (green) still localizes to the nuclei of the muscle fibers (magenta). (K,L) w1118 (K) and scraps8 (L) mutants have normal muscle patterning. (M,M′) α-Spectrin (magenta) and DAPI (green) staining shows that scraps8 mutants have cytokinesis defects in the epidermis. The boxed region in M is shown at high magnification in M′, where arrowheads indicate binucleate cells.
Mutants for Pav, but not Anillin, show defects in muscle guidance, and Pav is essential for RacGAP localization
During cell division, the direct interaction of MT-bound RacGAP with Pav is crucial for cytokinesis (D'Avino et al., 2006; Somers and Saint, 2003). Furthermore, studies of CYK-4 and ZEN-4 (CeMKLP1), the C. elegans orthologs of RacGAP and Pav, have shown that their localization is mutually dependent (Jantsch-Plunger et al., 2000) and that a multimeric complex between CYK-4 and ZEN-4 is sufficient to bundle MTs in vitro (Mishima et al., 2002). Based upon these results, we tested whether Pav also functions with RacGAP during muscle migration. To examine the relationship between RacGAP and Pav localization, we expressed UAS::GFP-pav (Minestrini et al., 2003) in the muscles using the Mef2-GAL4 driver and immunostained with antibodies against RacGAP and GFP. The UAS::GFP-pav transgene was used in previous studies to examine Pav localization (Minestrini et al., 2003). We observed that GFP-Pav is localized primarily to the nucleus in muscles (Fig. 5E,F). This is in contrast to RacGAP, which was mainly cytoplasmic, with discrete perinuclear puncta (Fig. 5A,C,C′; see Fig. S2 in the supplementary material). Furthermore, in embryos expressing GFP-pav, we observed an increase in RacGAP staining (Fig. 5D, compare with 5A). Closer examination of these embryos revealed that the increase in RacGAP staining occurred at the nuclear periphery (Fig. 5F′, compare with 5C′). These results suggest that perinuclear RacGAP localization is likely to occur through an interaction with Pav.
Examination of pavB200, a zygotic LOF mutant, revealed a similar muscle phenotype to that seen in RacGAPDH15 homozygotes (Fig. 5H, compare with Fig. 1G). Furthermore, when we examined RacGAP localization in pavB200 homozygotes, we found significant changes in the RacGAP localization pattern. Specifically, RacGAP became enriched at the ends of the muscle fiber (Fig. 5I) and was not as highly concentrated in perinuclear cytoplasmic puncta (Fig. 5A,C,C′). By contrast, Pav still localized to the nucleus in RacGAPDH15 mutants (Fig. 5J). These results provide strong evidence that muscle patterning requires RacGAP localization to discrete cytoplasmic puncta that are concentrated at the nuclear periphery, and that Pav is essential for this localization.
Anillin is an actin-interacting protein known to be essential for cytokinesis in Drosophila and vertebrates (Field and Alberts, 1995; Oegema et al., 2000; Piekny and Glotzer, 2008; Straight et al., 2005). Recent studies have shown that RacGAP is necessary for Anillin accumulation at the cleavage furrow and that they colocalize in vivo and interact in vitro (D'Avino et al., 2008; Gregory et al., 2008). Anillin is encoded by scraps, and scraps zygotic mutants are embryonic lethal indicating that maternally contributed Anillin is depleted before the end of embryogenesis (Heitzler et al., 1993). We examined both the epidermis and muscles in scraps8 embryos and found cytokinesis defects in the epidermis (Fig. 5M,M′). However, there were no significant defects in the muscle pattern (Fig. 5L) as compared with the wild type (Fig. 5K). Although we cannot rule out the possibility that the maternal contribution of Anillin persists in the mesoderm, our results argue for a model in which RacGAP and Pav function together during muscle development independently of Anillin.
RacGAP mutants do not have general defects in the actin or MT cytoskeleton
Because of the established role of RacGAP in cytoskeletal organization during cytokinesis, we hypothesized that the essential function for RacGAP and Pav in somatic muscles could be for the reorganization of the actin and/or MT cytoskeleton during muscle development. We first investigated whether there were any general defects in the actin or MT networks in RacGAPDH15 myotubes, using Alexa 488-conjugated Phalloidin to label the actin cytoskeleton. Both wild-type and RacGAPDH15 mutant muscles fibers stained strongly for actin, and extended multiple actin-based filopodia at their ends during myotube migration (Fig. 6A-B′,E-F′). During the attachment phase, actin begins to accumulate at the muscle ends to support the forming attachment (Gregory and Brown, 1998) (Fig. 6C,D). In RacGAPDH15 mutants, actin accumulated normally at the sites of attachment, even when they formed at the wrong location (Fig. 6G,H). These results, along with the lack of muscle-patterning defects in scraps mutants (Fig. 5L), provide evidence that organization of the actin cytoskeleton is not the primary function of RacGAP during muscle development. This was further supported by our finding that myoblast fusion, another actin-based process (Richardson et al., 2008), was not disrupted in RacGAPDH15 mutants (Fig. 4H,J).
Fig. 6.

RacGAP mutants do not have general defects in the actin or microtubule cytoskeleton. (A-H) Phalloidin labeling showing LT (A-B′,E-F′) and ventral (C,D,G,H) muscles. During MAS selection, F-actin concentrates at the ends of myotubes in the wild type (A-C) and in RacGAPDH15 mutants (E-G). Magnification of LT ends shows multiple filopodia in both wild type (B,B′) and RacGAPDH15 (F,F′). At stage 16, F-actin continues to accumulate at muscle ends (D). In RacGAPDH15 mutants, F-actin accumulation occurs even when muscles attach to the wrong site (H, arrowhead) or contain ectopic extensions (H, arrow). (I-L) Anti-β3-tubulin labels microtubules (MTs) in somatic muscles. At stage 13 during myoblast fusion, wild type (I) and RacGAPDH15 (K) have a similar β3-tubulin pattern. By stage 16, wild-type MTs are organized along the longitudinal axis of each muscle (J). In RacGAPDH15 mutants, MTs are present, but muscles make inappropriate attachments (L, arrowheads).
We also examined the MT network in wild-type and RacGAPDH15 mutant myotubes using an antibody against β3-tubulin, a muscle-specific tubulin isoform (Buttgereit et al., 1996). In RacGAPDH15 mutants, we did not observe any significant changes in the β3-tubulin pattern as compared with the wild type at stage 13, as myoblasts fuse (Fig. 6I,K), or at later stages as myotubes extend and make attachments (Fig. 6J,L).
RacGAP is required for uniform MT polarity
To further characterize MTs in RacGAPDH15 mutants, we examined MT polarity. In multinucleated myotubes, the minus ends of MTs are found near the center of the fiber and the plus ends are directed outward as muscles extend towards their attachment sites (Clark et al., 1997). To assess MT polarity in RacGAPDH15 myotubes, we used a UAS-nod::GFP transgene (Andersen et al., 2005) with the G14-GAL4 driver. Nod::GFP is a chimera that localizes to MT minus ends in multiple cell types and is commonly used as a MT minus-end marker (Clark et al., 1997; Whited et al., 2004; Zheng et al., 2008). As shown previously for Nod::β-Gal (Clark et al., 1997), Nod::GFP was detectable at high levels in the interior region of the muscle fibers and was notably absent from muscle ends at stage 16 (Fig. 7D,G). This localization pattern was also visible at stage 14/15 as the myotubes migrate to their attachment sites (Fig. 7A,C), providing evidence for the establishment of MT polarity during the migratory phase of muscle development. In RacGAPDH15 mutants, Nod::GFP frequently became enriched at muscle ends in stage 16 embryos (Fig. 7E,H). The mislocalization of Nod::GFP was also visible at stage 14/15, when Nod:GFP was distributed throughout the cytoplasm instead of near the interior of the myotube (Fig. 7B,C′). Mislocalization of Nod::GFP was also observed in pavB200 embryos (Fig. 7I). Co-expression of UAS-tum::myc and UAS-nod::GFP in RacGAPDH15 muscles significantly rescued guidance defects and also restored the wild-type localization of Nod::GFP (Fig. 7F). These results are consistent with the idea that RacGAP plays an important role in MT organization in the muscle fiber, and without RacGAP the MTs are no longer uniform in their polarity. Furthermore, these results suggest that the disorganization of MTs in RacGAPDH15 mutants is the cause of the muscle guidance defects we observe. Overall muscle polarity and trafficking were not affected in RacGAPDH15 mutants, as shown by staining with an antibody against the transmembrane receptor Roundabout (Kramer et al., 2001), which localized to muscle ends in wild-type and RacGAPDH15 embryos (see Fig. S3A,B in the supplementary material).
Fig. 7.

RacGAP is required for uniform MT polarity. (A-I) G14-GAL4 driving UAS-nod::GFP labels MT minus ends. Brackets mark individual muscles, arrowheads mark muscle ends. As wild-type muscles extend (A,C) and make attachments (D,G), Nod::GFP accumulates towards the center of the fiber (arrows) and is notably absent from muscle ends. In RacGAPDH15 (B,C′,E,H) and pavB200 (I) mutants, Nod::GFP is present throughout the entire muscle, which shows a disruption of MT polarity. C and C′ are higher magnifications of stage 14/15 Nod::GFP expression in wild type and RacGAPDH15 mutants. (F) Co-expression of UAS-tum::myc and UAS-nod::GFP in RacGAPDH15 mutants results in significant rescue of muscle patterning and restoration of MT polarity, as shown by the absence of Nod::GFP at muscle ends. G and H are higher magnifications of D and E, respectively, showing Nod::GFP at the interior of muscle fibers in the wild type (G) and at the ends of muscles in RacGAPDH15 (H).
γ-tubulin requires RacGAP for its localization to the nuclear periphery
Studies in cultured myotubes have shown that γ-tubulin, a minus-end anchoring protein essential for MT nucleation at the centrosome (Gunawardane et al., 2000; Stearns et al., 1991), may also be able to promote nucleation from non-centrosomal sites along the nuclear membrane and throughout the cytoplasm (Bugnard et al., 2005; Cottam et al., 2006; Musa et al., 2003). Based upon the previously defined role for RacGAP in MT organization during cell division, as well as our current findings that MT polarity in myotubes is altered in RacGAP and pav mutants (Fig. 7B,E,I), we decided to examine γ-tubulin localization in wild-type, RacGAPDH15 and pavB200 embryos. In wild-type mononucleated myoblasts, γ-tubulin is localized diffusely throughout the cytoplasm (Fig. 8G). This pattern is consistent with that recently reported for multiple Drosophila cell types during interphase, which lack functional centrosomes and organize MTs independently of MTOCs (Rogers et al., 2008). However, in multinucleated myotubes, the pattern of γ-tubulin was significantly different. By examining fillet preparations, we found that similar to in cultured myotubes, γ-tubulin was weakly detected and concentrated in discrete cytoplasmic puncta that accumulated towards the interior of the muscle (Fig. 8A). This pattern of γ-tubulin was strikingly similar to that observed for RacGAP (Fig. 5A), suggesting that these proteins might colocalize in myotubes.
Immunostaining for both RacGAP and γ-tubulin revealed significant colabeling of these proteins in muscle fibers (Fig. 8A-C′). Further examination of γ-tubulin staining revealed that γ-tubulin accumulates at high concentration at the nuclear periphery (Fig. 8D,D′). Furthermore, the pattern of γ-tubulin localization was significantly altered in muscle fibers of RacGAPDH15 and pavB200 mutants. In pavB200 embryos, in which RacGAP protein is enriched at the ends of the muscle fiber (Fig. 5I), we saw a decrease of γ-tubulin at the nuclear periphery and often observed patches of γ-tubulin near the ends of the muscle fiber (Fig. 8E,E′). By contrast, γ-tubulin staining was more diffusely dispersed throughout the cytoplasm in RacGAPDH15 embryos (Fig. 8F,F′). This diffuse cytoplasmic pattern of γ-tubulin is similar to that seen in wild-type mononucleated myoblasts (Fig. 8G). These results show that RacGAP colocalizes with γ-tubulin, and that RacGAP and Pav are required for the reorganization of the MT array in multinucleated myotubes by localizing γ-tubulin to the nuclear periphery.
DISCUSSION
The reorganization of the actin and MT cytoskeletons during myogenesis has long been observed in cell culture as individual myoblasts fuse into multinucleated myotubes (Bugnard et al., 2005; Cottam et al., 2006; Musa et al., 2003; Pizon et al., 2005; Tassin et al., 1985). Although the actin cytoskeleton has been shown to be indispensable for mediating myoblast fusion (Kim et al., 2007; Massarwa et al., 2007; Peckham, 2008; Richardson et al., 2007), little is known about how the MT network is remodeled during muscle development or the developmental significance of this event. In this study, we provide evidence that RacGAP, a known regulator of the MT cytoskeleton during cytokinesis, and Pav, a kinesin-like RacGAP-binding protein, play a novel and important role in MT organization in vivo by localizing γ-tubulin to perinuclear sites in myotubes (Fig. 9). Furthermore, we show that the organization of the MT network in multinucleated myotubes is important for MAS selection. Muscles that are mutant for RacGAP or pav have defects in MT polarity and fail to properly extend towards their attachment sites, resulting in defects in somatic muscle patterning.
Fig. 9.
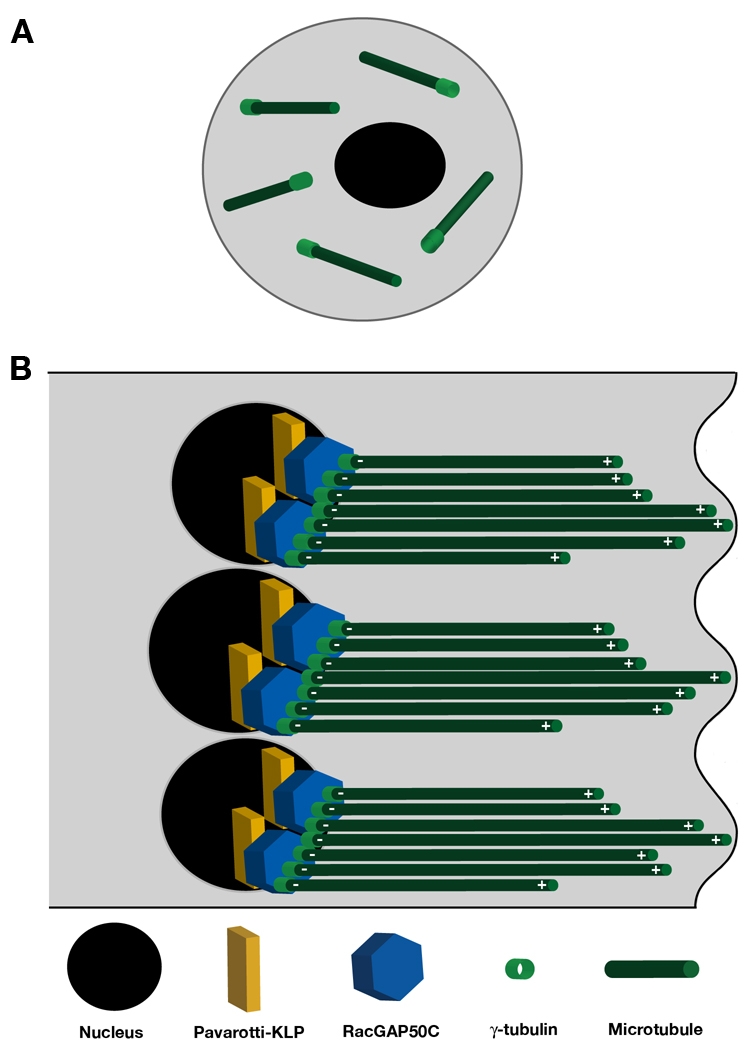
Model of RacGAP and Pav function during myotube extension in Drosophila. (A) Mononucleate myoblasts have MTs dispersed throughout the cytoplasm and diffuse cytoplasmic γ-tubulin localization. (B) One end of an elongating myotube. In multinucleated myotubes, the MT array must be reorganized in the longitudinal axis of the muscle to allow for elongation and extension. This organization requires MT polarity, characterized by minus ends at the interior of the myotube and plus ends at the periphery, to drive extension. RacGAP, Pavarotti (KLP, kinesin-like protein) and γ-tubulin are required to establish the proper MT array in migrating myotubes. Pav localizes RacGAP to discrete cytoplasmic puncta at the nuclear periphery, and RacGAP localization determines γ-tubulin distribution. RacGAP may transport incoming MTs to the nuclear periphery after myoblast fusion and/or promote the nucleation of new MTs in the appropriate orientation by increasing γ-tubulin at the nuclear periphery, thus establishing the MT array required for myotube extension.
The current model for MT organization in differentiated myotubes has come primarily from cell culture studies, which describe MTs that run parallel to the long axis of the cell and do not appear to be directly associated with any one organizing center (Tassin et al., 1985). Further studies have demonstrated that proteins involved in MT organization, such as γ-tubulin, are redistributed from the centrosome of individual myoblasts to discrete cytoplasmic puncta as well as along the nuclear membrane in multinucleated myotubes and that these sites are associated with MT growth (Bugnard et al., 2005; Musa et al., 2003). The diffuse cytoplasmic distribution of γ-tubulin that we observe in Drosophila myoblasts (Fig. 8G) differs from that in cultured vertebrate myoblasts, in which γ-tubulin is associated with centrosomes. Nonetheless, in both cases, the MT cytoskeleton must be reorganized from either a centrosomal or broadly distributed array in individual myoblasts, to a parallel array in multinucleated myotubes with the plus ends directed outwards (Fig. 9). In this study, we show that RacGAP plays a crucial role in this reorganization. In the absence of RacGAP, MTs are not uniform in their polarity and γ-tubulin remains dispersed throughout the cytoplasm rather than accumulating at the nuclear periphery of multinucleated myotubes. Furthermore, in pav mutants, mislocalization of RacGAP is sufficient to redistribute γ-tubulin to the ends of myotubes.
To date, the perinuclear localization of γ-tubulin in myotubes has only been weakly detected in vitro (Bugnard et al., 2005). In this study, we show that the association of γ-tubulin with the nucleus also occurs in vivo and is dependent at least in part on RacGAP. What is the function of γ-tubulin localization to the nuclear periphery in myotubes? One likely possibility is to anchor MT minus ends. Because the nuclei in multinucleated myotubes cluster in the interior of the myotube, this would allow for the polarization of the MT network, which is aligned along the axis of cell migration, with the plus ends at the leading edge. What is the purpose of MT polymerization at the ends of myotubes? Although, conventionally, the driving force for cell motility has been thought to be provided mainly by the reorganization of the actin cytoskeleton, there is increasing evidence that MTs are indispensable for cell migration (Ridley, 2001; Watanabe et al., 2005). It has been hypothesized that MTs form longitudinal arrays in bipolar myotubes in order to facilitate elongation by `active crawling' of the two ends of the myotube during MAS selection (Musa et al., 2003). Our data point to a MT-based mechanism for myotube extension and MAS selection. In the absence of RacGAP or Pav, the MT network shows non-uniform polarity and many muscle fibers are abnormally shaped and display guidance errors. The effect of RacGAP and pav mutations on muscle morphology is consistent with previous findings in which both RacGAP and Pav have been implicated in regulating axonal outgrowth and maintaining dendritic morphology. RacGAP was identified in a genetic screen by the increased dendritic branching phenotype observed in tum mutants (Gao et al., 1999). RacGAP and Pav have also been shown to play a role in regulating the morphogenesis of postmitotic mushroom body neurons in the Drosophila brain (Goldstein et al., 2005). In addition, disruption of the mammalian form of Pav, KIF23 (CHO1; MKLP1), in postmitotic cultured neurons resulted in the rearrangement of MT polarity and in the disruption of dendrite morphology (Yu et al., 2000).
There is increasing evidence that morphological processes require regulated coordination of the cytoskeleton by linking actin and MTs. For example, in Drosophila the Rho activator RhoGEF2 is implicated both in Myosin II localization and MT organization via the localization of the plus-end protein Eb1 (Dawes-Hoang et al., 2005; Rogers et al., 2004). Likewise, RacGAP provides a connection between the actomyosin ring and the peripheral central-spindle MTs during cytokinesis via its interaction with the actin-binding protein Anillin (D'Avino et al., 2008; Gregory et al., 2008). In addition, proper formation of the cleavage furrow is dependent on a complex between RacGAP, the Rho activator Pebble, and the plus-end-directed MT protein Pav (Somers and Saint, 2003; Zavortink et al., 2005). Our data show that similar to its function during cytokinesis, the function of RacGAP in postmitotic myotubes depends on its association with the MT-binding protein Pav. However, the role of RacGAP in regulating γ-tubulin distribution appears to be independent of its interaction with Anillin and the actin cytoskeleton. scraps mutants do not show defects in muscle patterning (Fig. 5L). Furthermore, the organization of the actin cytoskeleton (Fig. 6) and two known actin-dependent processes, myoblast fusion (Fig. 4) and muscle attachment (Fig. 2), are not significantly affected in RacGAP mutants. These findings demonstrate a newly described function for RacGAP that is restricted to the modulation of MTs, but not the actin cytoskeleton, in postmitotic cells.
What is the developmental significance of the actin-independent function of RacGAP in myotube extension? The answer might lie in the complex process of myogenesis itself. Building a mature muscle fiber requires the coordination of many morphological processes, including myoblast fusion, myotube extension and muscle attachment. The uncoupling of actin- and MT-based cytoskeletal processes might allow for actin-based myoblast fusion and MT-based myotube elongation to occur simultaneously. This idea is supported by previous findings showing that myoblasts continue to fuse as the myotube elongates to find its attachment sites (Schnorrer and Dickson, 2004). In addition, fusion-defective mutant FCs have been observed to extend and attempt to migrate to their targets (Ruiz-Gomez et al., 2000; Rushton et al., 1995), demonstrating that the migration machinery is not perturbed in mutants in which fusion is disrupted.
It is not yet clear what serves as the trigger for MT reorganization upon myoblast fusion or how RacGAP is recruited for this process. It also remains to be determined whether RacGAP promotes the nucleation of new MTs at the nuclear periphery, or reorganizes existing MTs from fusing myoblasts. Changes in MT architecture could be regulated through a direct physical interaction between RacGAP and γ-tubulin, or indirectly through a complex with downstream targets of the GAP domain of RacGAP.
Supplementary material
Supplementary material for this article is available at http://dev.biologists.org/cgi/content/full/136/9/1411/DC1
Supplementary Material
Acknowledgments
We thank Dorit Parnas for initiating the chromosome II screen; Corey S. Goodman in whose laboratory this screen was initially conducted; Robert Connacher and Neil Rodriguez for technical assistance; Brian McCabe for muscle illustration; Mary Baylies, Joe Kramer and Ruth Steward for critical insights; Nadine Soplop, Edgardo Santiago-Martínez and Donald Winkelmann for helpful discussions and reading of the manuscript; and Mary Baylies, Detlev Buttgereit, David M. Glover, John Reinitz and Robert Saint for generously providing reagents.
This work was supported by National Institutes of Health grant AR054482 to S.G.K. Deposited in PMC for release after 12 months.
References
- Adams, R. R., Tavares, A. A., Salzberg, A., Bellen, H. J. and Glover, D. M. (1998). pavarotti encodes a kinesin-like protein required to organize the central spindle and contractile ring for cytokinesis. Genes Dev. 12, 1483-1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen, R., Li, Y., Resseguie, M. and Brenman, J. E. (2005). Calcium/calmodulin-dependent protein kinase II alters structural plasticity and cytoskeletal dynamics in Drosophila. J. Neurosci. 25, 8878-8888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartolini, F. and Gundersen, G. G. (2006). Generation of noncentrosomal microtubule arrays. J. Cell Sci. 119, 4155-4163. [DOI] [PubMed] [Google Scholar]
- Bate, M. (1990). The embryonic development of larval muscles in Drosophila. Development 110, 791-804. [DOI] [PubMed] [Google Scholar]
- Bate, M. and Rushton, E. (1993). Myogenesis and muscle patterning in Drosophila. CR Acad. Sci. III 316, 1047-1061. [PubMed] [Google Scholar]
- Baylies, M. K. and Michelson, A. M. (2001). Invertebrate myogenesis: looking back to the future of muscle development. Curr. Opin. Genet. Dev. 11, 431-439. [DOI] [PubMed] [Google Scholar]
- Baylies, M. K., Bate, M. and Ruiz Gomez, M. (1998). Myogenesis: a view from Drosophila. Cell 93, 921-927. [DOI] [PubMed] [Google Scholar]
- Becker, S., Pasca, G., Strumpf, D., Min, L. and Volk, T. (1997). Reciprocal signaling between Drosophila epidermal muscle attachment cells and their corresponding muscles. Development 124, 2615-2622. [DOI] [PubMed] [Google Scholar]
- Beckett, K. and Baylies, M. K. (2007). 3D analysis of founder cell and fusion competent myoblast arrangements outlines a new model of myoblast fusion. Dev. Biol. 309, 113-125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118, 401-415. [DOI] [PubMed] [Google Scholar]
- Brown, N. H., Gregory, S. L. and Martin-Bermudo, M. D. (2000). Integrins as mediators of morphogenesis in Drosophila. Dev. Biol. 223, 1-16. [DOI] [PubMed] [Google Scholar]
- Bugnard, E., Zaal, K. J. and Ralston, E. (2005). Reorganization of microtubule nucleation during muscle differentiation. Cell Motil. Cytoskeleton 60, 1-13. [DOI] [PubMed] [Google Scholar]
- Buttgereit, D., Paululat, A. and Renkawitz-Pohl, R. (1996). Muscle development and attachment to the epidermis is accompanied by expression of beta 3 and beta 1 tubulin isotypes, respectively. Int. J. Dev. Biol. 40, 189-196. [PubMed] [Google Scholar]
- Callahan, C. A., Bonkovsky, J. L., Scully, A. L. and Thomas, J. B. (1996). derailed is required for muscle attachment site selection in Drosophila. Development 122, 2761-2767. [DOI] [PubMed] [Google Scholar]
- Campos-Ortega, J. A. and Hartenstein, V. (1985). The Embryonic Development of Drosophila Melanogaster. New York: Springer Verlag.
- Carmena, A., Murugasu-Oei, B., Menon, D., Jimenez, F. and Chia, W. (1998). Inscuteable and numb mediate asymmetric muscle progenitor cell divisions during Drosophila myogenesis. Genes Dev. 12, 304-315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmena, A., Buff, E., Halfon, M. S., Gisselbrecht, S., Jimenez, F., Baylies, M. K. and Michelson, A. M. (2002). Reciprocal regulatory interactions between the Notch and Ras signaling pathways in the Drosophila embryonic mesoderm. Dev. Biol. 244, 226-242. [DOI] [PubMed] [Google Scholar]
- Chanana, B., Graf, R., Koledachkina, T., Pflanz, R. and Vorbruggen, G. (2007). AlphaPS2 integrin-mediated muscle attachment in Drosophila requires the ECM protein Thrombospondin. Mech. Dev. 124, 463-475. [DOI] [PubMed] [Google Scholar]
- Clark, I. E., Jan, L. Y. and Jan, Y. N. (1997). Reciprocal localization of Nod and kinesin fusion proteins indicates microtubule polarity in the Drosophila oocyte, epithelium, neuron and muscle. Development 124, 461-470. [DOI] [PubMed] [Google Scholar]
- Cottam, D. M., Tucker, J. B., Rogers-Bald, M. M., Mackie, J. B., Macintyre, J., Scarborough, J. A., Ohkura, H. and Milner, M. J. (2006). Non-centrosomal microtubule-organising centres in cold-treated cultured Drosophila cells. Cell Motil. Cytoskeleton 63, 88-100. [DOI] [PubMed] [Google Scholar]
- D'Avino, P. P., Savoian, M. S., Capalbo, L. and Glover, D. M. (2006). RacGAP50C is sufficient to signal cleavage furrow formation during cytokinesis. J. Cell Sci. 119, 4402-4408. [DOI] [PubMed] [Google Scholar]
- D'Avino, P. P., Takeda, T., Capalbo, L., Zhang, W., Lilley, K. S., Laue, E. D. and Glover, D. M. (2008). Interaction between Anillin and RacGAP50C connects the actomyosin contractile ring with spindle microtubules at the cell division site. J. Cell Sci. 121, 1151-1158. [DOI] [PubMed] [Google Scholar]
- Dawes-Hoang, R. E., Parmar, K. M., Christiansen, A. E., Phelps, C. B., Brand, A. H. and Wieschaus, E. F. (2005). Folded gastrulation, cell shape change and the control of myosin localization. Development 132, 4165-4178. [DOI] [PubMed] [Google Scholar]
- Devenport, D., Bunch, T. A., Bloor, J. W., Brower, D. L. and Brown, N. H. (2007). Mutations in the Drosophila alphaPS2 integrin subunit uncover new features of adhesion site assembly. Dev. Biol. 308, 294-308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dohrmann, C., Azpiazu, N. and Frasch, M. (1990). A new Drosophila homeo box gene is expressed in mesodermal precursor cells of distinct muscles during embryogenesis. Genes Dev. 4, 2098-2111. [DOI] [PubMed] [Google Scholar]
- Estrada, B., Gisselbrecht, S. S. and Michelson, A. M. (2007). The transmembrane protein Perdido interacts with Grip and integrins to mediate myotube projection and attachment in the Drosophila embryo. Development 134, 4469-4478. [DOI] [PubMed] [Google Scholar]
- Field, C. M. and Alberts, B. M. (1995). Anillin, a contractile ring protein that cycles from the nucleus to the cell cortex. J. Cell Biol. 131, 165-178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frasch, M. (1999). Controls in patterning and diversification of somatic muscles during Drosophila embryogenesis. Curr. Opin. Genet. Dev. 9, 522-529. [DOI] [PubMed] [Google Scholar]
- Gao, F. B., Brenman, J. E., Jan, L. Y. and Jan, Y. N. (1999). Genes regulating dendritic outgrowth, branching, and routing in Drosophila. Genes Dev. 13, 2549-2561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein, A. Y., Jan, Y. N. and Luo, L. (2005). Function and regulation of Tumbleweed (RacGAP50C) in neuroblast proliferation and neuronal morphogenesis. Proc. Natl. Acad. Sci. USA 102, 3834-3839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gregory, S. L. and Brown, N. H. (1998). kakapo, a gene required for adhesion between and within cell layers in Drosophila, encodes a large cytoskeletal linker protein related to plectin and dystrophin. J. Cell Biol. 143, 1271-1282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gregory, S. L., Ebrahimi, S., Milverton, J., Jones, W. M., Bejsovec, A. and Saint, R. (2008). Cell division requires a direct link between microtubule-bound RacGAP and Anillin in the contractile ring. Curr. Biol. 18, 25-29. [DOI] [PubMed] [Google Scholar]
- Gunawardane, R. N., Lizarraga, S. B., Wiese, C., Wilde, A. and Zheng, Y. (2000). gamma-Tubulin complexes and their role in microtubule nucleation. Curr. Top. Dev. Biol. 49, 55-73. [DOI] [PubMed] [Google Scholar]
- Halfon, M. S., Carmena, A., Gisselbrecht, S., Sackerson, C. M., Jimenez, F., Baylies, M. K. and Michelson, A. M. (2000). Ras pathway specificity is determined by the integration of multiple signal-activated and tissue-restricted transcription factors. Cell 103, 63-74. [DOI] [PubMed] [Google Scholar]
- Heitzler, P., Coulson, D., Saenz-Robles, M. T., Ashburner, M., Roote, J., Simpson, P. and Gubb, D. (1993). Genetic and cytogenetic analysis of the 43A-E region containing the segment polarity gene costa and the cellular polarity genes prickle and spiny-legs in Drosophila melanogaster. Genetics 135, 105-115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hime, G. and Saint, R. (1992). Zygotic expression of the pebble locus is required for cytokinesis during the postblastoderm mitoses of Drosophila. Development 114, 165-171. [DOI] [PubMed] [Google Scholar]
- Jantsch-Plunger, V., Gonczy, P., Romano, A., Schnabel, H., Hamill, D., Schnabel, R., Hyman, A. A. and Glotzer, M. (2000). CYK-4: A Rho family GTPase activating protein (GAP) required for central spindle formation and cytokinesis. J. Cell Biol. 149, 1391-1404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarecki, J., Johnson, E. and Krasnow, M. A. (1999). Oxygen regulation of airway branching in Drosophila is mediated by branchless FGF. Cell 99, 211-220. [DOI] [PubMed] [Google Scholar]
- Jones, W. M. and Bejsovec, A. (2005). RacGap50C negatively regulates wingless pathway activity during Drosophila embryonic development. Genetics 169, 2075-2086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, S., Shilagardi, K., Zhang, S., Hong, S. N., Sens, K. L., Bo, J., Gonzalez, G. A. and Chen, E. H. (2007). A critical function for the actin cytoskeleton in targeted exocytosis of prefusion vesicles during myoblast fusion. Dev. Cell 12, 571-586. [DOI] [PubMed] [Google Scholar]
- Kramer, S. G., Kidd, T., Simpson, J. H. and Goodman, C. S. (2001). Switching repulsion to attraction: changing responses to slit during transition in mesoderm migration. Science 292, 737-740. [DOI] [PubMed] [Google Scholar]
- Massarwa, R., Carmon, S., Shilo, B. Z. and Schejter, E. D. (2007). WIP/WASp-based actin-polymerization machinery is essential for myoblast fusion in Drosophila. Dev. Cell 12, 557-569. [DOI] [PubMed] [Google Scholar]
- Minestrini, G., Harley, A. S. and Glover, D. M. (2003). Localization of Pavarotti-KLP in living Drosophila embryos suggests roles in reorganizing the cortical cytoskeleton during the mitotic cycle. Mol. Biol. Cell 14, 4028-4038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishima, M., Kaitna, S. and Glotzer, M. (2002). Central spindle assembly and cytokinesis require a kinesin-like protein/RhoGAP complex with microtubule bundling activity. Dev. Cell 2, 41-54. [DOI] [PubMed] [Google Scholar]
- Musa, H., Orton, C., Morrison, E. E. and Peckham, M. (2003). Microtubule assembly in cultured myoblasts and myotubes following nocodazole induced microtubule depolymerisation. J. Muscle Res. Cell Motil. 24, 301-308. [PMC free article] [PubMed] [Google Scholar]
- Oegema, K., Savoian, M. S., Mitchison, T. J. and Field, C. M. (2000). Functional analysis of a human homologue of the Drosophila actin binding protein anillin suggests a role in cytokinesis. J. Cell Biol. 150, 539-552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parnas, D., Haghighi, A. P., Fetter, R. D., Kim, S. W. and Goodman, C. S. (2001). Regulation of postsynaptic structure and protein localization by the Rho-type guanine nucleotide exchange factor dPix. Neuron 32, 415-424. [DOI] [PubMed] [Google Scholar]
- Patel, N. H. (1994). Imaging neuronal subsets and other cell types in whole-mount Drosophila embryos and larvae using antibody probes. Methods Cell Biol. 44, 445-487. [DOI] [PubMed] [Google Scholar]
- Peckham, M. (2008). Engineering a multi-nucleated myotube, the role of the actin cytoskeleton. J. Microsc. 231, 486-493. [DOI] [PubMed] [Google Scholar]
- Pereanu, W., Spindler, S., Im, E., Buu, N. and Hartenstein, V. (2007). The emergence of patterned movement during late embryogenesis of Drosophila. Dev. Neurobiol. 67, 1669-1685. [DOI] [PubMed] [Google Scholar]
- Piekny, A. J. and Glotzer, M. (2008). Anillin is a scaffold protein that links RhoA, actin, and myosin during cytokinesis. Curr. Biol. 18, 30-36. [DOI] [PubMed] [Google Scholar]
- Pizon, V., Gerbal, F., Diaz, C. C. and Karsenti, E. (2005). Microtubule-dependent transport and organization of sarcomeric myosin during skeletal muscle differentiation. EMBO J. 24, 3781-3792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson, B. E., Beckett, K., Nowak, S. J. and Baylies, M. K. (2007). SCAR/WAVE and Arp2/3 are crucial for cytoskeletal remodeling at the site of myoblast fusion. Development 134, 4357-4367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson, B., Beckett, K. and Baylies, M. (2008). Visualizing new dimensions in Drosophila myoblast fusion. BioEssays 30, 423-431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridley, A. J. (2001). Rho GTPases and cell migration. J. Cell Sci. 114, 2713-2722. [DOI] [PubMed] [Google Scholar]
- Rogers, G. C., Rusan, N. M., Peifer, M. and Rogers, S. L. (2008). A multicomponent assembly pathway contributes to the formation of acentrosomal microtubule arrays in interphase Drosophila cells. Mol. Biol. Cell 19, 3163-3178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers, S. L., Wiedemann, U., Hacker, U., Turck, C. and Vale, R. D. (2004). Drosophila RhoGEF2 associates with microtubule plus ends in an EB1-dependent manner. Curr. Biol. 14, 1827-1833. [DOI] [PubMed] [Google Scholar]
- Ruiz Gomez, M. and Bate, M. (1997). Segregation of myogenic lineages in Drosophila requires numb. Development 124, 4857-4866. [DOI] [PubMed] [Google Scholar]
- Ruiz-Gomez, M., Coutts, N., Price, A., Taylor, M. V. and Bate, M. (2000). Drosophila dumbfounded: a myoblast attractant essential for fusion. Cell 102, 189-198. [DOI] [PubMed] [Google Scholar]
- Rushton, E., Drysdale, R., Abmayr, S. M., Michelson, A. M. and Bate, M. (1995). Mutations in a novel gene, myoblast city, provide evidence in support of the founder cell hypothesis for Drosophila muscle development. Development 121, 1979-1988. [DOI] [PubMed] [Google Scholar]
- Schnorrer, F. and Dickson, B. J. (2004). Muscle building; mechanisms of myotube guidance and attachment site selection. Dev. Cell 7, 9-20. [DOI] [PubMed] [Google Scholar]
- Schnorrer, F., Kalchhauser, I. and Dickson, B. J. (2007). The transmembrane protein Kon-tiki couples to Dgrip to mediate myotube targeting in Drosophila. Dev. Cell 12, 751-766. [DOI] [PubMed] [Google Scholar]
- Shishido, E., Takeichi, M. and Nose, A. (1998). Drosophila synapse formation: regulation by transmembrane protein with Leu-rich repeats, CAPRICIOUS. Science 280, 2118-2121. [DOI] [PubMed] [Google Scholar]
- Somers, W. G. and Saint, R. (2003). A RhoGEF and Rho family GTPase-activating protein complex links the contractile ring to cortical microtubules at the onset of cytokinesis. Dev. Cell 4, 29-39. [DOI] [PubMed] [Google Scholar]
- Sotillos, S. and Campuzano, S. (2000). DRacGAP, a novel Drosophila gene, inhibits EGFR/Ras signalling in the developing imaginal wing disc. Development 127, 5427-5438. [DOI] [PubMed] [Google Scholar]
- Stearns, T., Evans, L. and Kirschner, M. (1991). Gamma-tubulin is a highly conserved component of the centrosome. Cell 65, 825-836. [DOI] [PubMed] [Google Scholar]
- Straight, A. F., Field, C. M. and Mitchison, T. J. (2005). Anillin binds nonmuscle myosin II and regulates the contractile ring. Mol. Biol. Cell 16, 193-201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swan, L. E., Wichmann, C., Prange, U., Schmid, A., Schmidt, M., Schwarz, T., Ponimaskin, E., Madeo, F., Vorbruggen, G. and Sigrist, S. J. (2004). A glutamate receptor-interacting protein homolog organizes muscle guidance in Drosophila. Genes Dev. 18, 223-237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swan, L. E., Schmidt, M., Schwarz, T., Ponimaskin, E., Prange, U., Boeckers, T., Thomas, U. and Sigrist, S. J. (2006). Complex interaction of Drosophila GRIP PDZ domains and Echinoid during muscle morphogenesis. EMBO J. 25, 3640-3651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tassin, A. M., Maro, B. and Bornens, M. (1985). Fate of microtubule-organizing centers during myogenesis in vitro. J. Cell Biol. 100, 35-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toure, A., Dorseuil, O., Morin, L., Timmons, P., Jegou, B., Reibel, L. and Gacon, G. (1998). MgcRacGAP, a new human GTPase-activating protein for Rac and Cdc42 similar to Drosophila rotund RacGAP gene product, is expressed in male germ cells. J. Biol. Chem. 273, 6019-6023. [DOI] [PubMed] [Google Scholar]
- Volk, T. (1999). Singling out Drosophila tendon cells: a dialogue between two distinct cell types. Trends Genet. 15, 448-453. [DOI] [PubMed] [Google Scholar]
- Watanabe, T., Noritake, J. and Kaibuchi, K. (2005). Regulation of microtubules in cell migration. Trends Cell Biol. 15, 76-83. [DOI] [PubMed] [Google Scholar]
- Whited, J. L., Cassell, A., Brouillette, M. and Garrity, P. A. (2004). Dynactin is required to maintain nuclear position within postmitotic Drosophila photoreceptor neurons. Development 131, 4677-4686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams, J. A., Bell, J. B. and Carroll, S. B. (1991). Control of Drosophila wing and haltere development by the nuclear vestigial gene product. Genes Dev. 5, 2481-2495. [DOI] [PubMed] [Google Scholar]
- Yu, W., Cook, C., Sauter, C., Kuriyama, R., Kaplan, P. L. and Baas, P. W. (2000). Depletion of a microtubule-associated motor protein induces the loss of dendritic identity. J. Neurosci. 20, 5782-5791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zavortink, M., Contreras, N., Addy, T., Bejsovec, A. and Saint, R. (2005). Tum/RacGAP50C provides a critical link between anaphase microtubules and the assembly of the contractile ring in Drosophila melanogaster. J. Cell Sci. 118, 5381-5392. [DOI] [PubMed] [Google Scholar]
- Zheng, Y., Wildonger, J., Ye, B., Zhang, Y., Kita, A., Younger, S. H., Zimmerman, S., Jan, L. Y. and Jan, Y. N. (2008). Dynein is required for polarized dendritic transport and uniform microtubule orientation in axons. Nat. Cell Biol. 10, 1172-1180. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.