

Abstract
One of the most critical problems we face in the study of biological systems is building accurate statistical descriptions of them. This problem has been particularly challenging because biological systems typically contain large numbers of interacting elements, which precludes the use of standard brute force approaches. Recently, though, several groups have reported that there may be an alternate strategy. The reports show that reliable statistical models can be built without knowledge of all the interactions in a system; instead, pairwise interactions can suffice. These findings, however, are based on the analysis of small subsystems. Here, we ask whether the observations will generalize to systems of realistic size, that is, whether pairwise models will provide reliable descriptions of true biological systems. Our results show that, in most cases, they will not. The reason is that there is a crossover in the predictive power of pairwise models: If the size of the subsystem is below the crossover point, then the results have no predictive power for large systems. If the size is above the crossover point, then the results may have predictive power. This work thus provides a general framework for determining the extent to which pairwise models can be used to predict the behavior of large biological systems. Applied to neural data, the size of most systems studied so far is below the crossover point.
Author Summary
Biological systems are exceedingly complicated: They consist of a large number of elements, those elements interact in nonlinear and highly unpredictable ways, and collective interactions typically play a critical role. It would seem surprising, then, that one could build a quantitative description of biological systems based only on knowledge of how pairs of elements interact. Yet, that is what a number of studies have found. Those studies, however, focused on relatively small systems. Here, we ask the question: Do their conclusions extend to large systems? We show that the answer depends on the size of the system relative to a crossover point: Below the crossover point the results on the small system have no predictive power for large systems; above the crossover point the results on the small system may have predictive power. Moreover, the crossover point can be computed analytically. This work thus provides a general framework for determining the extent to which pairwise models can be used to predict the behavior of large biological systems. It also provides a useful heuristic for designing experiments: If one is interested in understanding truly large systems via pairwise interactions, then make sure that the system one studies is above the crossover point.
Introduction
Many fundamental questions in biology are naturally treated in a probabilistic setting. For instance, deciphering the neural code requires knowledge of the probability of observing patterns of activity in response to stimuli [1]; determining which features of a protein are important for correct folding requires knowledge of the probability that a particular sequence of amino acids folds naturally [2],[3]; and determining the patterns of foraging of animals and their social and individual behavior requires knowledge of the distribution of food and species over both space and time [4]–[6].
Constructing these probability distributions is, however, hard. There are several reasons for this: i) biological systems are composed of large numbers of elements, and so can exhibit a huge number of configurations—in fact, an exponentially large number, ii) the elements typically interact with each other, making it impossible to view the system as a collection of independent entities, and iii) because of technological considerations, the descriptions of biological systems have to be built from very little data. For example, with current technology in neuroscience, we can record simultaneously from only about 100 neurons out of approximately 100 billion in the human brain. So, not only are we faced with the problem of estimating probability distributions in high dimensional spaces, we must do this based on a small fraction of the neurons in the network.
Despite these apparent difficulties, recent work has suggested that the situation may be less bleak than it seems, and that an accurate statistical description of systems can be achieved without having to examine all possible configurations [2], [3], [7]–[11]. One merely has to measure the probability distribution over pairs of elements and use those to build the full distribution. These “pairwise models” potentially offer a fundamental simplification, as the number of pairs is quadratic in the number of elements, not exponential. However, support for the efficacy of pairwise models has, necessarily, come from relatively small subsystems—small enough that the true probability distribution could be measured experimentally [7]–[9],[11]. While these studies have provided a key first step, a critical question remains: will the results from the analysis of these small subsystems extrapolate to large ones? That is, if a pairwise model predicts the probability distribution for a subset of the elements in a system, will it also predict the probability distribution for the whole system? Here we find that, for a biologically relevant class of systems, this question can be answered quantitatively and, importantly, generically—independent of many of the details of the biological system under consideration. And the answer is, generally, “no.” In this paper, we explain, both analytically and with simulations, why this is the case.
Results
The extrapolation problem
To gain intuition into the extrapolation problem, let us consider a specific
example: neuronal spike trains. Fig. 1A shows a typical spike train for a small population of
neurons. Although the raw spike times provide a complete description, they are
not a useful representation, as they are too high-dimensional. Therefore, we
divide time into bins and re-represent the spike train as 0 s and 1 s: 0 if
there is no spike in a bin; 1 otherwise (Fig. 1B) [7]–[9],[11]. For
now we assume that the bins are independent (an assumption whose validity we
discuss below, and in more detail in the section “Is there anything
wrong with using small time bins?”). The problem, then, is to find 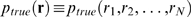 where
where 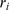 is a binary variable indicating no spike (
is a binary variable indicating no spike (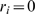 ) or one or more spikes (
) or one or more spikes ( ) on neuron
) on neuron  . Since this, too, is a high dimensional problem (though less
so than the original spike time representation), suppose that we instead
construct a pairwise approximation to
. Since this, too, is a high dimensional problem (though less
so than the original spike time representation), suppose that we instead
construct a pairwise approximation to  , which we denote
, which we denote  , for a population of size
, for a population of size  . (The pairwise model derives its name from the fact that it
has the same mean and pairwise correlations as the true model; see Eq. (15).)
Our question, then, is: if is close to for small , what can we say about how close the two distributions are for
large ?
. (The pairwise model derives its name from the fact that it
has the same mean and pairwise correlations as the true model; see Eq. (15).)
Our question, then, is: if is close to for small , what can we say about how close the two distributions are for
large ?
Figure 1. Transforming spike trains to spike count.

(A) Spike rasters. Tick marks indicate spike times; different rows correspond to different neurons. The horizontal dashed lines are the bin boundaries. (B) Spike count in each bin. In this example the bins are small enough that there is at most one spike per bin, but this is not necessary—one could use bigger bins and have larger spike counts.
To answer this question quantitatively, we need a measure of distance. The
measure we use, denoted  , is defined in Eq. (3) below, but all we need to know about it
for now is that if
, is defined in Eq. (3) below, but all we need to know about it
for now is that if 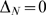 then
then  , and if is near one then is far from . In terms of , our main results are as follows. First, for small , in what we call the perturbative regime, is proportional to
, and if is near one then is far from . In terms of , our main results are as follows. First, for small , in what we call the perturbative regime, is proportional to  . In other words, as the population size increases, the
pairwise model becomes a worse and worse approximation to the true distribution.
Second, this behavior is entirely generic: for small , increases linearly, no matter what the true distribution is.
This is illustrated schematically in Fig. 2, which shows the generic behavior of . The solid red part of the curve is the perturbative regime,
where is a linearly increasing function of ; the dashed curves show possible behavior beyond the
perturbative regime.
. In other words, as the population size increases, the
pairwise model becomes a worse and worse approximation to the true distribution.
Second, this behavior is entirely generic: for small , increases linearly, no matter what the true distribution is.
This is illustrated schematically in Fig. 2, which shows the generic behavior of . The solid red part of the curve is the perturbative regime,
where is a linearly increasing function of ; the dashed curves show possible behavior beyond the
perturbative regime.
Figure 2. Cartoon illustrating the dependence of on .

For small there is always a perturbative regime in which increases linearly with (solid red line). When becomes large, may continue increasing with (red and black dashed lines) or it may plateau (cyan
dashed line), depending on . The observation that increases linearly with does not, therefore, provide much, if any information
about the large behavior.
These results have an important corollary: if one does an experiment and finds
that is increasing linearly with , then one has no information at all about the true
distribution. The flip side of this is more encouraging: if one can measure the
true distribution for sufficiently large that saturates, as for the dashed blue line in Fig. 2, then there is a chance that
extrapolation to large is meaningful. The implications for the interpretation of
experiments is, therefore, that one can gain information about large behavior only if one can analyze data past the perturbative
regime.
Under what conditions is a subsystem in the perturbative regime? The answer turns
out to be simple: the size of the system, , times the average probability of observing a spike in a bin,
must be small compared to 1. For example, if the average probability is 1/100,
then a system will be in the perturbative regime if the number of neurons is
small compared to 100. This last observation would seem to be good news: if we
divide the spike trains into sufficiently small time bins and ignore temporal
correlations, then we can model the data very well with a pairwise distribution.
The problem with this, though, is that temporal correlations can be ignored only
when time bins are large compared to the autocorrelation time. This leads to a
kind of catch-22: pairwise models are guaranteed to work well (in the sense that
they describe spike trains in which temporal correlations are ignored) if one
uses small time bins, but small time bins is the one regime where ignoring
temporal correlations is not a valid approximation.
In the next several sections we quantify the qualitative picture presented above:
we write down an explicit expression for , explain why it increases linearly with when is small, and provide additional tests, besides assessing the
linearity of , to determine whether or not one is in the perturbative
regime.
Quantifying how well the pairwise model explains the data
A natural measure of the distance between and is the Kullback-Leibler (KL) divergence [12], denoted  and defined as
and defined as
| (1) |
The KL divergence is zero if the two distributions are equal; otherwise it is nonzero.
Although the KL divergence is a very natural measure, it is not easy to interpret
(except, of course, when it is exactly zero). That is because a nonzero KL
divergence tells us that 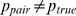 , but it does not give us any real handle on how good, or bad,
the pairwise model really is. To make sense of the KL divergence, we need
something to compare it to. A reasonable reference quantity, used by a number of
authors [7]–[9], is the KL divergence
between the true distribution and the independent one, the latter denoted
, but it does not give us any real handle on how good, or bad,
the pairwise model really is. To make sense of the KL divergence, we need
something to compare it to. A reasonable reference quantity, used by a number of
authors [7]–[9], is the KL divergence
between the true distribution and the independent one, the latter denoted  . The independent distribution, as its name suggests, is a
distribution in which the variables are taken to be independent,
. The independent distribution, as its name suggests, is a
distribution in which the variables are taken to be independent,
| (2) |
where  is the distribution of the response of the
is the distribution of the response of the  neuron,
neuron,  . With this choice for a comparison, we define a normalized
distance measure—a measure of how well the pairwise model explains the data—as
. With this choice for a comparison, we define a normalized
distance measure—a measure of how well the pairwise model explains the data—as
| (3) |
Note that the denominator in this expression, 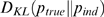 , is usually referred to as the multi-information [7],[13],[14].
, is usually referred to as the multi-information [7],[13],[14].
The quantity lies between 0 and 1, and measures how well a pairwise model
does relative to an independent model. If it is 0, the pairwise model is equal
to the true model (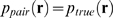 ); if it is near 1, the pairwise model offers little
improvement over the independent model; and if it is exactly 1, the pairwise
model is equal to the independent model (
); if it is near 1, the pairwise model offers little
improvement over the independent model; and if it is exactly 1, the pairwise
model is equal to the independent model ( ), and so offers no improvement.
), and so offers no improvement.
How do we attach intuitive meaning to the two divergences  and
and 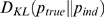 ? For the latter, we use the fact that, as is easy to show,
? For the latter, we use the fact that, as is easy to show,
| (4) |
where  and
and  are the entropies [15],[16] of
are the entropies [15],[16] of  and , respectively, defined, as usual, to be
and , respectively, defined, as usual, to be 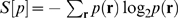 . For the former, we use the definition of the KL divergence to write
. For the former, we use the definition of the KL divergence to write
 |
(5) |
The quantity  has the flavor of an entropy, although it is a true entropy
only when is maximum entropy as well as pairwise (see Eq. (6) below).
For other pairwise distributions, all we need to know is that
has the flavor of an entropy, although it is a true entropy
only when is maximum entropy as well as pairwise (see Eq. (6) below).
For other pairwise distributions, all we need to know is that  lies between and . A plot illustrating the relationship between , the two entropies and , and the entropy-like quantity
lies between and . A plot illustrating the relationship between , the two entropies and , and the entropy-like quantity  , is shown in Fig.
3.
, is shown in Fig.
3.
Figure 3. Schematic plot of (black line),  (cyan line) and (red line).
(cyan line) and (red line).

The better the pairwise model, the closer  is to . This is reflected in the normalized distance measure, , which is the distance between the red and cyan lines
divided by the distance between the red and black lines.
is to . This is reflected in the normalized distance measure, , which is the distance between the red and cyan lines
divided by the distance between the red and black lines.
Note that for pairwise maximum entropy models (or maximum entropy models for
short), has a particularly simple interpretation, since in this case  really is an entropy. Using
really is an entropy. Using  to denote the pairwise entropy of a maximum entropy model, for
this case we have
to denote the pairwise entropy of a maximum entropy model, for
this case we have
| (6) |
as is easy to see by inserting Eqs. (4) and (5) into (3). This expression has been used previously by a number of authors [7],[9].
in the perturbative regime
The extrapolation problem discussed above is the problem of determining in the large limit. This is hard to do in general, but there is a
perturbative regime in which it is possible. The small parameter that defines
this regime is the average number of spikes produced by the whole population of
neurons in each time bin. It is given quantitatively by 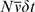 where
where 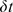 is the bin size and
is the bin size and 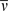 the average firing rate,
the average firing rate,
| (7) |
with  the firing rate of neuron
the firing rate of neuron  .
.
The first step in the perturbation expansion is to compute the two quantities
that make up :  and
and  . As we show in the section “Perturbative
Expansion” (Methods), these are
given by
. As we show in the section “Perturbative
Expansion” (Methods), these are
given by
| (8a) |
| (8b) |
where
| (9a) |
| (9b) |
Here and in what follows we use  to denote terms that are proportional to
to denote terms that are proportional to  in the limit
in the limit  . The
. The  in Eq. (9a) has been noted previously [7], although the
authors did not compute the prefactor,
in Eq. (9a) has been noted previously [7], although the
authors did not compute the prefactor, 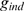 .
.
The prefactors and  , which are given explicitly in Eqs. (42) and (44), depend on
the low order statistics of the spike trains: depends on the second order normalized correlation
coefficients, depends on the second and third order normalized correlation
coefficients (the normalized correlation coefficients are defined in Eq. (16)
below), and both depend on the firing rates of the individual cells. The details
of that dependence, however, are not important for now; what is important is
that and are independent of and
, which are given explicitly in Eqs. (42) and (44), depend on
the low order statistics of the spike trains: depends on the second order normalized correlation
coefficients, depends on the second and third order normalized correlation
coefficients (the normalized correlation coefficients are defined in Eq. (16)
below), and both depend on the firing rates of the individual cells. The details
of that dependence, however, are not important for now; what is important is
that and are independent of and  (at least on average; see next section).
(at least on average; see next section).
Inserting Eq. (8) into Eq. (3) (into the definition of ) and using Eq. (9), we arrive at our main result,
| (10a) |
| (10b) |
Note that in the regime  , is necessarily small. This explains why, in an analytic study
of non-pairwise model in which
, is necessarily small. This explains why, in an analytic study
of non-pairwise model in which  was small, Shlens et al. found that was rarely greater than 0.1 [8].
was small, Shlens et al. found that was rarely greater than 0.1 [8].
We refer to quantities with a superscript zero as “zeroth order.” Note that, via Eqs. (4) and (5), we can also define zeroth order entropies,
| (11a) |
| (11b) |
These quantities are important primarily because differences between them and the actual entropies indicate a breakdown of the perturbation expansion (see in particular Fig. 4 below).
Figure 4. Cartoon showing extrapolations of the zeroth order KL divergences and entropies (see Eqs. (9) and (11)).

These extrapolations illustrate why the two natural quantities derived
from them,  and
and 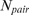 , occur beyond the point at which the extrapolation is
meaningful. (A) Extrapolations on a log-log scale. Black: ; green:
, occur beyond the point at which the extrapolation is
meaningful. (A) Extrapolations on a log-log scale. Black: ; green:  ; cyan:
; cyan:  . The red points are the data. The points and label the intersections of the two extrapolations with
the independent entropy, . (B) Extrapolation of the entropies rather than the KL
divergences, plotted on a linear-linear scale. The data, again shown in
red, is barely visible in the lower left hand corner. Black: ; solid orange:
. The red points are the data. The points and label the intersections of the two extrapolations with
the independent entropy, . (B) Extrapolation of the entropies rather than the KL
divergences, plotted on a linear-linear scale. The data, again shown in
red, is barely visible in the lower left hand corner. Black: ; solid orange:  ; solid maroon:
; solid maroon:  . The dashed orange and maroon lines are the
extrapolations of the true entropy and true pairwise
“entropy”, respectively.
. The dashed orange and maroon lines are the
extrapolations of the true entropy and true pairwise
“entropy”, respectively.
Assuming, as discussed in the next section, that and are approximately independent of , , and , Eq. (10) tells us that scales linearly with in the perturbative regime—the regime in which  . The key observation about this scaling is that it is
independent of the details of the true distribution, . This has a very important consequence, one that has major
implications for experimental data: if one does an experiment with small
. The key observation about this scaling is that it is
independent of the details of the true distribution, . This has a very important consequence, one that has major
implications for experimental data: if one does an experiment with small  and finds that is proportional to
and finds that is proportional to  , then the system is, with very high probability, in the
perturbative regime, and one does not know whether will remain close to as increases. What this means in practical terms is that if one
wants to know whether a particular pairwise model is a good one for large
systems, it is necessary to consider values of that are significantly greater than
, then the system is, with very high probability, in the
perturbative regime, and one does not know whether will remain close to as increases. What this means in practical terms is that if one
wants to know whether a particular pairwise model is a good one for large
systems, it is necessary to consider values of that are significantly greater than  , where
, where
| (12) |
We interpret  as the value at which there is a crossover in the behavior of
the pairwise model. Specifically, if
as the value at which there is a crossover in the behavior of
the pairwise model. Specifically, if  , the system is in the perturbative regime and the pairwise
model is not informative about the large behavior, whereas if
, the system is in the perturbative regime and the pairwise
model is not informative about the large behavior, whereas if  , the system is in a regime in which it may be possible to make
inferences about the behavior of the full system.
, the system is in a regime in which it may be possible to make
inferences about the behavior of the full system.
The prefactors, and
As we show in Methods (see in particular
Eqs. (42) and (44)), the prefactors and depend on which neurons out of the full population are used.
Consequently, these quantities fluctuate around their true values (in the sense
that different subpopulations produce different values of and ), where “true” refers to an average over
all possible  sub-populations. Here we assume that the neurons are chosen randomly from the full population, so any
set of neurons provides unbiased estimates of and . In our simulations, the fluctuations were small, as indicated
by the small error bars on the blue points in Fig. 5. However, in general the size of the
fluctuations is determined by the range of firing rates and correlation
coefficients, with larger ranges producing larger fluctuations.
sub-populations. Here we assume that the neurons are chosen randomly from the full population, so any
set of neurons provides unbiased estimates of and . In our simulations, the fluctuations were small, as indicated
by the small error bars on the blue points in Fig. 5. However, in general the size of the
fluctuations is determined by the range of firing rates and correlation
coefficients, with larger ranges producing larger fluctuations.
Figure 5. The dependence of the KL divergences and the normalized
distance measure, .

Data was generated from a third order model, as explained in the section
“Generating synthetic data” (Methods), and fit to pairwise maximum entropy models
and independent models. All data points correspond to averages over
marginalizations of the true distribution (see text for details). The
red points were computed directly using Eqs. (1), (3) and (4); the blue
points are the zeroth order estimates,  ,
,  , and
, and  , in rows 1, 2 and 3, respectively. The three columns
correspond to
, in rows 1, 2 and 3, respectively. The three columns
correspond to  , 0.029, and 0.037, from left to right. (A, B, C) (
, 0.029, and 0.037, from left to right. (A, B, C) ( ). Predictions from the perturbative expansion are in
good agreement with the measurements up to
). Predictions from the perturbative expansion are in
good agreement with the measurements up to  , indicating that the data is in the perturbative
regime. (D, E, F) (
, indicating that the data is in the perturbative
regime. (D, E, F) ( ). Predictions from the perturbative expansion are in
good agreement with the measurements up to
). Predictions from the perturbative expansion are in
good agreement with the measurements up to  , indicating that the data is only partially in the
perturbative regime. (G, H, I) (
, indicating that the data is only partially in the
perturbative regime. (G, H, I) (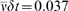 ). Predictions from the perturbative expansion are not
in good agreement with the measurements, even for small , indicating that the data is outside the perturbative
regime.
). Predictions from the perturbative expansion are not
in good agreement with the measurements, even for small , indicating that the data is outside the perturbative
regime.
Because does not affect the mean values of and , it is reasonable to think of these quantities—or at
least their true values—as being independent of . They are also independent of , again modulo fluctuations. Finally, as we show in the section
“Bin size and the correlation coefficients” (Methods), and are independent of in the limit that is small compared to the width of the temporal correlations
among neurons. We will assume this limit applies here. In sum, then, to first
approximation, and are independent of our three important quantities: , , and . Thus, we treat them as effectively constant throughout our
analysis.
The dangers of extrapolation
Although the behavior of in the perturbative regime does not tell us much about its
behavior at large , it is possible that other quantities that can be calculated
in the perturbative regime, , , and (the last one exactly), are informative, as others have
suggested [7]. Here we show that this is not the
case—they also are uninformative.
The easiest way to relate the perturbative regime to the large regime is to ignore the corrections in Eqs. (8a) and (8b),
extrapolate the expressions for the zeroth order terms, and ask what their large behavior tells us. Generic versions of these extrapolations,
plotted on a log-log scale, are shown in Fig. 4A, along with a plot of the independent
entropy, (which is necessarily linear in ). The first thing we notice about the extrapolations is that
they do not, technically, have a large behavior: one terminates at the point labeled , which is where 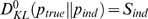 (and thus, via Eq. (0a),
(and thus, via Eq. (0a), 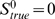 ; continuing the extrapolation implies negative true zeroth
order entropy); the other at the point labeled , which is where
; continuing the extrapolation implies negative true zeroth
order entropy); the other at the point labeled , which is where  (and thus, via Eq. (5) and the fact that
(and thus, via Eq. (5) and the fact that 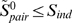 ,
,  ).
).
Despite the fact that the extrapolations end abruptly, they still might provide
information about the large regime. For example, and/or might be values of at which something interesting happens. To see if this is the
case, in Fig. 4B we plot the
naive extrapolations of  and (that is, the zeroth order quantities given in Eq. (11),
and (that is, the zeroth order quantities given in Eq. (11), 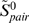 and
and  ), on a linear-linear plot, along with . This plot contains no new information compared to Fig. 4A, but it does elucidate
the meaning of the extrapolations. Perhaps its most striking feature is that the
naive extrapolation of has a decreasing portion. As is easy to show mathematically,
entropy cannot decrease with (intuitively, that is because observing one additional neuron
cannot decrease the entropy of previously observed neurons). Thus, , which occurs well beyond the point where the naive
extrapolation of is decreasing, has essentially no meaning, something that has
been pointed out previously by Bethge and Berens [10]. The other
potentially important value of is . This, though, suffers from a similar problem: when
), on a linear-linear plot, along with . This plot contains no new information compared to Fig. 4A, but it does elucidate
the meaning of the extrapolations. Perhaps its most striking feature is that the
naive extrapolation of has a decreasing portion. As is easy to show mathematically,
entropy cannot decrease with (intuitively, that is because observing one additional neuron
cannot decrease the entropy of previously observed neurons). Thus, , which occurs well beyond the point where the naive
extrapolation of is decreasing, has essentially no meaning, something that has
been pointed out previously by Bethge and Berens [10]. The other
potentially important value of is . This, though, suffers from a similar problem: when  ,
,  is negative.
is negative.
How do the naively extrapolated entropies—the solid lines in Fig. 4B—compare to
the actual entropies? To answer this, in Fig. 4B we show the true behavior of and  versus (dashed lines). Note that is asymptotically linear in , even though the neurons are correlated, a fact that forces
versus (dashed lines). Note that is asymptotically linear in , even though the neurons are correlated, a fact that forces 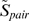 to be linear in , as it is sandwiched between and . (The asymptotically linear behavior of is typical, even in highly correlated systems. Although this
is not always appreciated, it is easy to show; see the section “The
behavior of the true entropy in the large N limit,”
Methods.) Comparing the dashed and solid lines, we see that the naively
extrapolated and true entropies, and thus the naively extrapolated and true
values of , have extremely different behavior. This further suggests that
there is very little connection between the perturbative and large regimes.
to be linear in , as it is sandwiched between and . (The asymptotically linear behavior of is typical, even in highly correlated systems. Although this
is not always appreciated, it is easy to show; see the section “The
behavior of the true entropy in the large N limit,”
Methods.) Comparing the dashed and solid lines, we see that the naively
extrapolated and true entropies, and thus the naively extrapolated and true
values of , have extremely different behavior. This further suggests that
there is very little connection between the perturbative and large regimes.
In fact, these observations follow directly from the fact that and depend only on correlation coefficients up to third order (see
Eqs. (42) and (44)) whereas the large behavior depends on correlations at all orders. Thus, since and tell us very little, if anything, about higher order
correlations, it is not surprising that they tell us very little about the
behavior of in the large limit.
Numerical simulations
To check that our perturbation expansions, Eqs. (8–10), are correct,
and to investigate the regime in which they are valid, we performed numerical
simulations. We generated, from synthetic data, a set of true distributions,
computed the true distance measures,  ,
,  , and numerically, and compared them to the zeroth order ones,
, and numerically, and compared them to the zeroth order ones,  ,
,  , and
, and  . If the perturbation expansion is valid, then the true values
should be close to the zeroth order values whenever
. If the perturbation expansion is valid, then the true values
should be close to the zeroth order values whenever  is small. The results are shown in Fig. 5, and that is, indeed, what we
observed. Before discussing that figure, though, we explain our procedure for
constructing true distributions.
is small. The results are shown in Fig. 5, and that is, indeed, what we
observed. Before discussing that figure, though, we explain our procedure for
constructing true distributions.
The set of true distributions we used were generated from a third order model (so named because it includes up to third order interactions). This model has the form
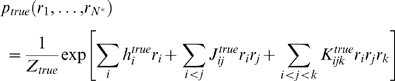 |
(13) |
where  is a normalization constant, chosen to ensure that the
probability distribution sums to 1, and the sums over
is a normalization constant, chosen to ensure that the
probability distribution sums to 1, and the sums over  ,
, 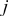 and
and 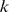 run from 1 to
run from 1 to 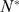 . The parameters
. The parameters  and
and  were chosen by sampling from distributions (see the section
“Generating synthetic data,” Methods), which allowed us to
generate many different true distributions. In all of our simulations we
calculate the relevant quantities directly from Eq. (13) . Consequently, we do
not have to worry about issues of finite data, as would be the case in realistic
experiments.
were chosen by sampling from distributions (see the section
“Generating synthetic data,” Methods), which allowed us to
generate many different true distributions. In all of our simulations we
calculate the relevant quantities directly from Eq. (13) . Consequently, we do
not have to worry about issues of finite data, as would be the case in realistic
experiments.
For a particular simulation (corresponding to a column in Fig. 5), we generated a true distribution
with 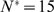 , randomly chose 5 neurons, and marginalized over them. This
gave us a 10-neuron true distribution. True distributions with
, randomly chose 5 neurons, and marginalized over them. This
gave us a 10-neuron true distribution. True distributions with 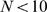 were constructed by marginalizing over additional neurons
within our 10-neuron population. To achieve a representative sample, we
considered all possible marginalizations (of which there are 10 choose , or
were constructed by marginalizing over additional neurons
within our 10-neuron population. To achieve a representative sample, we
considered all possible marginalizations (of which there are 10 choose , or  ). The results in Fig. 5 are averages over these marginalizations.
). The results in Fig. 5 are averages over these marginalizations.
For neural data, the most commonly used pairwise model is the maximum entropy model. Therefore, we use that one here. To emphasize the maximum entropy nature of this model, we replace the label “pair” that we have been using so far with “maxent.” The maximum entropy distribution has the form
 |
(14) |
Fitting this distribution requires that we choose the 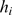 and
and 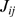 so that the first and second moments match those of the true
distribution. Quantitatively, these conditions are
so that the first and second moments match those of the true
distribution. Quantitatively, these conditions are
| (15a) |
| (15b) |
where the angle brackets,  and
and  , represent averages with respect to
, represent averages with respect to  and , respectively. Once we have
and , respectively. Once we have  and
and  that satisfy Eq. (15), we calculate the KL divergences, Eqs.
(1) and (4), and use those to compute .
that satisfy Eq. (15), we calculate the KL divergences, Eqs.
(1) and (4), and use those to compute .
The results are shown in Fig.
5. The rows correspond to our three quantities of interest: 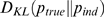 ,
,  , and (top to bottom). The columns correspond to different values of
, and (top to bottom). The columns correspond to different values of  , with the smallest
, with the smallest 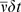 on the left and the largest on the right. Red circles are the
true values of these quantities; blue ones are the zeroth order predictions from
Eqs. (9) and (10b).
on the left and the largest on the right. Red circles are the
true values of these quantities; blue ones are the zeroth order predictions from
Eqs. (9) and (10b).
As suggested by our perturbation analysis, the smaller the value of 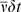 , the larger the value of for which agreement between the true and zeroth order values
is good. Our simulations corroborate this: the left column of Fig. 5 has
, the larger the value of for which agreement between the true and zeroth order values
is good. Our simulations corroborate this: the left column of Fig. 5 has  , and agreement is almost perfect out to
, and agreement is almost perfect out to  ; the middle column has
; the middle column has  , and agreement is almost perfect out to
, and agreement is almost perfect out to  ; and the right column has
; and the right column has 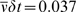 , and agreement is not good for any value of . Note that the perturbation expansion breaks down for values
of well below
, and agreement is not good for any value of . Note that the perturbation expansion breaks down for values
of well below  (defined in Eq.(12)): in the middle column of Fig. 5 it breaks down when
(defined in Eq.(12)): in the middle column of Fig. 5 it breaks down when  , and in the right column it breaks down when
, and in the right column it breaks down when 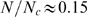 . This is not, however, especially surprising, as the
perturbation expansion is guaranteed to be valid only if
. This is not, however, especially surprising, as the
perturbation expansion is guaranteed to be valid only if  .
.
These results validate the perturbation expansions in Eqs. (8) and (10), and show
that those expansions provide sensible predictions—at least for some
parameters. They also suggest a natural way to assess the significance of
one's data: plot  ,
,  , and versus , and look for agreement with the predictions of the
perturbation expansion. If agreement is good, as in the left column of Fig. 5, then one is in the
perturbative regime, and one knows very little about the true distribution. If,
on the other hand, agreement is bad, as in the right column, then one is out of
the perturbative regime, and it may be possible to extract meaningful
information about the relationship between the true and pairwise models.
, and versus , and look for agreement with the predictions of the
perturbation expansion. If agreement is good, as in the left column of Fig. 5, then one is in the
perturbative regime, and one knows very little about the true distribution. If,
on the other hand, agreement is bad, as in the right column, then one is out of
the perturbative regime, and it may be possible to extract meaningful
information about the relationship between the true and pairwise models.
That said, the qualifier “at least for some parameters” is an
important one. This is because the perturbation expansion is essentially an
expansion that depends on the normalized correlation coefficients, and there is
an underlying assumption that they don't exhibit pathological behavior.
The  order normalized correlation coefficient for the distribution
order normalized correlation coefficient for the distribution  , denoted
, denoted  , is written
, is written
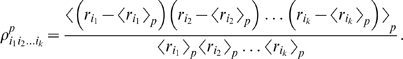 |
(16) |
A potentially problematic feature of the correlation coefficients
is that the denominator is a product over mean activities. If the mean
activities are small, the denominator can become very small, leading to very
large correlation coefficients. Although our perturbation expansion is always
valid for sufficiently small time bins (because the correlation coefficients
eventually becomes independent of bin size; see the section “Bin size
and the correlation coeffcients,” Methods), “sufficiently
small” can depend in detail on the parameters. For instance, at the
maximum population size tested ( ) and for the true distributions that had
) and for the true distributions that had 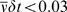 , the absolute error of the prediction had a median of
approximately 16%. However, about 11% of the runs had
errors larger than 60%. Thus, the exact size of the small parameter
at which the perturbative expansion breaks down can depend on the details of the
true distribution.
, the absolute error of the prediction had a median of
approximately 16%. However, about 11% of the runs had
errors larger than 60%. Thus, the exact size of the small parameter
at which the perturbative expansion breaks down can depend on the details of the
true distribution.
External fields and pairwise couplings have a simple dependence on firing rates and correlation coefficients in the perturbative regime
Estimation of the KL divergences and from real data can be hard, in the sense that it takes a large
amount of data for them to converge to their true values. In addition, as
discussed above, in the section “The prefactors
gind and
gpair”, there are fluctuations in associated with finite subsampling of the full population of
neurons. Those fluctuations tend to keep from being purely linear, as can seen, for example, in the
blue points in Fig. 5F and
5I. We therefore provide a second set of relationships that can be used
to determine whether or not a particular data set is in the perturbative regime.
These relationships are between the parameters of the maximum entropy model, the 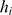 and
and 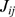 , and the mean activity and normalized second order correlation
coefficient (the latter defined in Eq. (19) below).
, and the mean activity and normalized second order correlation
coefficient (the latter defined in Eq. (19) below).
Since the quantity 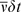 plays a central role in our analysis, we replace it with a
single parameter, which we denote
plays a central role in our analysis, we replace it with a
single parameter, which we denote  ,
,
| (17) |
In terms of this parameter, we find (using the same perturbative approach that led us to Eqs. (8–10); see the section “External fields, pairwise couplings and moments,” Methods), that
| (18a) |
| (18b) |
where 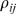 , the normalized second order correlation coefficient, is
defined in Eq. (16) with
, the normalized second order correlation coefficient, is
defined in Eq. (16) with 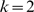 ; it is given explicitly by
; it is given explicitly by
| (19) |
(We don't need a superscript on  or a subscript on the angle brackets because the first and
second moments are the same under the true and pairwise distributions.) Equation
(18a) can be reconstructed from the low firing rate limit of analysis carried
out by Sessak and Monasson [17], as can the first three terms in the
expansion of the log in Eq. (18b).
or a subscript on the angle brackets because the first and
second moments are the same under the true and pairwise distributions.) Equation
(18a) can be reconstructed from the low firing rate limit of analysis carried
out by Sessak and Monasson [17], as can the first three terms in the
expansion of the log in Eq. (18b).
Equation (18) tells us that the 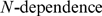 of the
of the 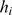 and
and  , the external fields and pairwise couplings, is very weak. In
Fig. 6 we confirm this
through numerical simulations. Equation (18b) also provides additional
information—it gives us a functional relationship between the pairwise
couplings and the normalized pairwise correlations function, . In Fig.
7A–C we plot the pairwise couplings,
, the external fields and pairwise couplings, is very weak. In
Fig. 6 we confirm this
through numerical simulations. Equation (18b) also provides additional
information—it gives us a functional relationship between the pairwise
couplings and the normalized pairwise correlations function, . In Fig.
7A–C we plot the pairwise couplings,  , versus the normalized pairwise correlation coefficient, (blue dots), along with the prediction from Eq. (18b) (black
line). Consistent with our predictions, the data in Fig. 7A–C essentially follows a
line—the line given by Eq. (18b).
, versus the normalized pairwise correlation coefficient, (blue dots), along with the prediction from Eq. (18b) (black
line). Consistent with our predictions, the data in Fig. 7A–C essentially follows a
line—the line given by Eq. (18b).
Figure 6. The true external fields and pairwise interactions compared with the predictions of the perturbation expansion.

The top row shows the true external fields, 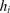 , versus those predicted from Eq. (18a), and the bottom
row shows the true pairwise interaction,
, versus those predicted from Eq. (18a), and the bottom
row shows the true pairwise interaction,  , versus those predicted from Eq. (18b). Values of ranging from 5 to 10 are shown, with different colors
corresponding to different
, versus those predicted from Eq. (18b). Values of ranging from 5 to 10 are shown, with different colors
corresponding to different  . For each value of , data is shown for 45 realization of the true
distribution. Insets show the
. For each value of , data is shown for 45 realization of the true
distribution. Insets show the  of the mean external fields (top) and mean pairwise
interactions (bottom). The three columns correspond exactly to the
columns in Fig. 5.
(A, B) (
of the mean external fields (top) and mean pairwise
interactions (bottom). The three columns correspond exactly to the
columns in Fig. 5.
(A, B) ( ). There is a very good match between the true and
predicted values of both external fields and pairwise interactions. (C,
D) (
). There is a very good match between the true and
predicted values of both external fields and pairwise interactions. (C,
D) ( ). Even though
). Even though 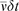 has increased, the match is still good. (E, F) (
has increased, the match is still good. (E, F) ( ). The true and predicted external fields and pairwise
interactions do not match as well as the cases shown in (A, B, C, D).
There is also now a stronger
). The true and predicted external fields and pairwise
interactions do not match as well as the cases shown in (A, B, C, D).
There is also now a stronger  in the mean external fields compared to (A) and (B).
The
in the mean external fields compared to (A) and (B).
The  of the pairwise interactions in (F) is weaker than
that of the external fields, but still notably stronger than the ones in
(B) and (D). This indicates that the perturbative expansion is starting
to break down.
of the pairwise interactions in (F) is weaker than
that of the external fields, but still notably stronger than the ones in
(B) and (D). This indicates that the perturbative expansion is starting
to break down.
Figure 7. The relation between pairwise couplings and pairwise correlations.

This figure shows that there is a simple relation between  and , but not between
and , but not between  and
and 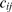 . (A, C, E)
. (A, C, E)  versus the normalized coefficients, (blue points), along with the predicted relationship,
via Eq. (18b) (black line). (B, D, F)
versus the normalized coefficients, (blue points), along with the predicted relationship,
via Eq. (18b) (black line). (B, D, F)  versus the Pearson correlation coefficients, , Eq. (26) (blue points). The three columns correspond
exactly to the columns in Fig. 5 from left to right; that is,
versus the Pearson correlation coefficients, , Eq. (26) (blue points). The three columns correspond
exactly to the columns in Fig. 5 from left to right; that is,  for (A, B),
for (A, B), 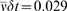 for (C, D), and
for (C, D), and  for (E, F). The prediction in the top row (black line)
matches the data well, even in the rightmost column.
for (E, F). The prediction in the top row (black line)
matches the data well, even in the rightmost column.
A relationship between the pairwise couplings and the correlations coefficients
has been sought previously, but for the more standard Pearson correlation
coefficient [7],[9],[11]. Our analysis explains why it was not found.
The Pearson correlation coefficient, denoted , is given by
 |
(20) |
In the small  limit—the limit of interest—the right hand
side of Eq. (20) is approximately equal to
limit—the limit of interest—the right hand
side of Eq. (20) is approximately equal to 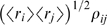 . Because
. Because  depends on the external fields,
depends on the external fields, 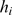 and
and  (see Eq. (18a)) and there is a one-to-one
relationship between and
(see Eq. (18a)) and there is a one-to-one
relationship between and  (Eq. (18b)), there can't be a one-to-one relationship
between and
(Eq. (18b)), there can't be a one-to-one relationship
between and 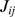 . We verify the lack of a relationship in Fig. 7D and 7E, where we again plot
. We verify the lack of a relationship in Fig. 7D and 7E, where we again plot  , but this time versus the standard correlation coefficient, . As predicted, the data in Fig. 7D and 7E is scattered over two
dimensions. This suggests that , not , is the natural measure of the correlation between two neurons
when they have a binary representation, something that has also been suggested
by Amari based on information-geometric arguments [18].
, but this time versus the standard correlation coefficient, . As predicted, the data in Fig. 7D and 7E is scattered over two
dimensions. This suggests that , not , is the natural measure of the correlation between two neurons
when they have a binary representation, something that has also been suggested
by Amari based on information-geometric arguments [18].
Note that the lack of a simple relationship between the pairwise couplings and
the standard correlation coefficient has been a major motivation in building
maximum entropy models [7],[11]. This is for good reason: if there is a
simple relationship, knowing the  adds essentially nothing. Thus, plotting
adds essentially nothing. Thus, plotting  versus (but not ) is an important test of one's data, and if the two
quantities fall on the curve predicted by Eq. (18b), the maximum entropy model
is adding very little information, if any.
versus (but not ) is an important test of one's data, and if the two
quantities fall on the curve predicted by Eq. (18b), the maximum entropy model
is adding very little information, if any.
As an aside, we should point out that the 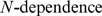 is a function of the variables used to represent the firing
patterns. Here we use 0 for no spike and 1 for one or more spikes, but another,
possibly more common, representation, derived from the Ising model and used in a
number of studies [7],[9],[11], is to use −1 and +1
rather than 0 and 1. This amounts to making the change of variables
is a function of the variables used to represent the firing
patterns. Here we use 0 for no spike and 1 for one or more spikes, but another,
possibly more common, representation, derived from the Ising model and used in a
number of studies [7],[9],[11], is to use −1 and +1
rather than 0 and 1. This amounts to making the change of variables 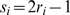 . In terms of
. In terms of  , the maximum entropy model has the form
, the maximum entropy model has the form 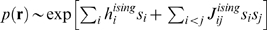 where
where 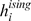 and
and  are given by
are given by
| (21a) |
| (21b) |
The second term on the right side of Eq. (21a) is proportional to  , which means the external fields in the Ising representation
acquire a linear
, which means the external fields in the Ising representation
acquire a linear 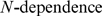 that was not present in our 0/1 representation. The two
studies that reported the
that was not present in our 0/1 representation. The two
studies that reported the  of the external fields [7],[9] used
this representation, and, as predicted by our analysis, the external fields in
those studies had a component that was linear in .
of the external fields [7],[9] used
this representation, and, as predicted by our analysis, the external fields in
those studies had a component that was linear in .
Is there anything wrong with using small time bins?
An outcome of our perturbative approach is that our normalized distance measure, , is linear in bin size (see Eq. (10b)). This suggests that one
could make the pairwise model look better and better simply by making the bin
size smaller and smaller. Is there anything wrong with this? The answer is yes,
for reasons discussed above (see the the section “The extrapolation
problem”); here we emphasize and expand on this issue, as it is an
important one for making sense of experimental results.
The problem arises because what we have been calling the “true” distribution is not really the true distribution of spike trains. It is the distribution assuming independent time bins, an assumption that becomes worse and worse as we make the bins smaller and smaller. (We use this potentially confusing nomenclature primarily because all studies of neuronal data carried out so far have assumed temporal independence, and compared the pairwise distribution to the temporally independent—but still correlated across neurons—distribution [7]–[9],[11]. In addition, the correct name “true under the assumption of temporal independence,” is unwieldy.) Here we quantify how much worse. In particular, we show that if one uses time bins that are small compared to the characteristic correlation time in the spike trains, the pairwise model will not provide a good description of the data. Essentially, we show that, when the time bins are too small, the error one makes in ignoring temporal correlations is larger than the error one makes in ignoring correlations across neurons.
As usual, we divide time into bins of size . However, because we are dropping the independence assumption,
we use 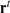 , rather than
, rather than  , to denote the response in bin
, to denote the response in bin  . The full probability distribution over all time bins is
denoted
. The full probability distribution over all time bins is
denoted 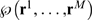 . Here
. Here 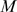 is the number of bins; it is equal to
is the number of bins; it is equal to  for spike trains of length
for spike trains of length 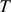 . If time bins are approximately independent and the
distribution of
. If time bins are approximately independent and the
distribution of  is the same for all
is the same for all  (an assumption we make for convenience only, but do not need;
see the section “Extending the normalized distance measure to the time
domain,” Methods), we can write
(an assumption we make for convenience only, but do not need;
see the section “Extending the normalized distance measure to the time
domain,” Methods), we can write
| (22) |
Furthermore, if the pairwise model is a good one, we have
| (23) |
Combining Eqs. (22) and Eq. (23) then gives us an especially simple expression
for the full probability distribution:  .
.
The problem with small time bins lies in Eq. (22): the right hand side is a good
approximation to the true distribution when the time bins are large compared to
the spike train correlation time, but it is a bad approximation when the time
bins are small (because adjacent time bins become highly correlated). To
quantify how bad, we compare the error one makes assuming independence across
time to the error one makes assuming independence across neurons. The ratio of
those two errors, denoted  , is given by
, is given by
 |
(24) |
It is relatively easy to compute in the limit of small time bins (see the section
“Extending the normalized distance measure to the time
domain,” Methods), and we find that
| (25) |
As expected, this reduces to our old result, , when there is only one time bin ( ). When is larger than 1, however, the second term is always at least
one, and for small bin size, the third term is much larger than one.
Consequently, if we use bins that are small compared to the temporal correlation
time of the spike trains, the pairwise model will do a very bad job describing
the full, temporally correlated spike trains.
). When is larger than 1, however, the second term is always at least
one, and for small bin size, the third term is much larger than one.
Consequently, if we use bins that are small compared to the temporal correlation
time of the spike trains, the pairwise model will do a very bad job describing
the full, temporally correlated spike trains.
Discussion
Probability distributions over the configurations of biological systems are extremely important quantities. However, because of the large number of interacting elements comprising such systems, these distributions can almost never be determined directly from experimental data. Using parametric models to approximate the true distribution is the only existing alternative. While such models are promising, they are typically applied only to small subsystems, not the full system. This raises the question: are they good models of the full system?
We answered this question for a class of parametric models known as pairwise models. We focused on a particular application, neuronal spike trains, and our main result is as follows: if one were to record spikes from multiple neurons, use sufficiently small time bins and a sufficiently small number of cells, and assume temporal independence, then a pairwise model will almost always succeed in matching the true (but temporally independent) distribution—whether or not it would match the true (but still temporally independent) distribution for large time bins or a large number of cells. In other words, pairwise models in the “sufficiently small” regime, what we refer to as the perturbative regime, have almost no predictive value for what will happen with large populations. This makes extrapolation from small to large systems dangerous.
This observation is important because pairwise models, and in particular pairwise
maximum entropy models, have recently attracted a great deal of attention: they have
been applied to salamander and guinea pig retinas [7], primate retina
[8],
primate cortex [9], cultured cortical networks [7], and cat visual
cortex [11].
These studies have mainly operated close to the perturbative regime. For example,
Schneidman et al. [7] had  (for the data set described in their Fig. 5), Tang et al. [9] had
(for the data set described in their Fig. 5), Tang et al. [9] had 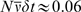 to 0.4 (depending on the preparation), and Yu et al. [11] had
to 0.4 (depending on the preparation), and Yu et al. [11] had 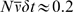 . For these studies, then, it would be hard to justify
extrapolating to large populations.
. For these studies, then, it would be hard to justify
extrapolating to large populations.
The study by Shlens et al. [8], on the other hand, might be more amenable to
extrapolation. This is because spatially localized visual patterns were used to
stimulate retinal ganglion cells, for which a nearest neighbor maximum entropy
models provided a good fit to their data. (Nearest neighbor means  is zero unless neuron
is zero unless neuron  and neuron are adjacent.) Our analysis still applies, but, since all but the
nearest neighbor correlations are zero, many of the terms that make up and vanish (see Eqs. (42) and (44)). Consequently, the small parameter
in the perturbative expansion becomes
and neuron are adjacent.) Our analysis still applies, but, since all but the
nearest neighbor correlations are zero, many of the terms that make up and vanish (see Eqs. (42) and (44)). Consequently, the small parameter
in the perturbative expansion becomes  (rather than
(rather than 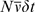 ), where
), where  is the number of nearest neighbors. Since
is the number of nearest neighbors. Since  is fixed, independent of the population size, the small parameter
will not change as the population size increases. Thus, Shlens et al.may have tapped
into the large population behavior even though they considered only a few cells at a
time in their analysis. Indeed, they have recently extended their analysis to more
than 100 neurons, and they still find that nearest neighbor maximum entropy models
provide very good fits to the data [19].
is fixed, independent of the population size, the small parameter
will not change as the population size increases. Thus, Shlens et al.may have tapped
into the large population behavior even though they considered only a few cells at a
time in their analysis. Indeed, they have recently extended their analysis to more
than 100 neurons, and they still find that nearest neighbor maximum entropy models
provide very good fits to the data [19].
Time bins and population size
That the pairwise model is always good if 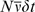 is sufficiently small has strong implications: if we want to
build a good model for a particular , we can simply choose a bin size that is small compared to
is sufficiently small has strong implications: if we want to
build a good model for a particular , we can simply choose a bin size that is small compared to 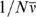 . However, one of the assumptions in all pairwise models used
on neural data is that bins at different times are independent. This produces a
tension between small time bins and temporal independence: small time bins
essentially ensure that a pairwise model will provide a close approximation to a
model with independent bins, but they make adjacent bins highly correlated.
Large time bins come with no such assurance, but they make adjacent bins
independent. Unfortunately, this tension is often unresolvable in large
populations, in the sense that pairwise models are assured to work only up to
populations of size
. However, one of the assumptions in all pairwise models used
on neural data is that bins at different times are independent. This produces a
tension between small time bins and temporal independence: small time bins
essentially ensure that a pairwise model will provide a close approximation to a
model with independent bins, but they make adjacent bins highly correlated.
Large time bins come with no such assurance, but they make adjacent bins
independent. Unfortunately, this tension is often unresolvable in large
populations, in the sense that pairwise models are assured to work only up to
populations of size 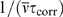 where τ
corr is the typical
correlation time. Given that is at least several Hz, for experimental paradigms in which
the correlation time is more than a few hundred ms,
where τ
corr is the typical
correlation time. Given that is at least several Hz, for experimental paradigms in which
the correlation time is more than a few hundred ms,  is about one, and this assurance does not apply to even
moderately sized populations of neurons.
is about one, and this assurance does not apply to even
moderately sized populations of neurons.
These observations are especially relevant for studies that use small time bins to model spike trains driven by natural stimuli. This is because the long correlation times inherent in natural stimuli are passed on to the spike trains, so the assumption of independence across time (which is required for the independence assumption to be valid) breaks badly. Knowing that these models are successful in describing spike trains under the independence assumption, then, does not tell us whether they will be successful in describing full, temporally correlated, spike trains. Note that for studies that use stimuli with short correlation times (e.g., non-natural stimuli such as white noise), the temporal correlations in the spike trains are likely to be short, and using small time bins may be perfectly valid.
The only study that has investigated the issue of temporal correlations in
maximum entropy models does indeed support the above picture [9]: for
the parameters used in that study ( to 0.4), the pairwise maximum entropy model provided a good
fit to the data ( was typically smaller than 0.1), but it did not do a good job
modeling the temporal structure of the spike trains.
to 0.4), the pairwise maximum entropy model provided a good
fit to the data ( was typically smaller than 0.1), but it did not do a good job
modeling the temporal structure of the spike trains.
Other systems—Protein folding
As mentioned in the Introduction, in addition to the studies on neuronal data,
studies on protein folding have also emphasized the role of pairwise
interactions [2],[3]. Briefly, proteins
consist of strings of amino acids, and a major question in structural biology
is: what is the probability distribution of amino acid strings in naturally
folding proteins? One way to answer this is to approximate the full probability
distribution of naturally folding proteins from knowledge of single-site and
pairwise distributions. One can show that there is a perturbative regime for
proteins as well. This can be readily seen using the celebrated HP protein model
[20], where a protein is composed of only two types of
amino acids. If, at each site, one amino acid type is preferred and occurs with
high probability, say  with
with 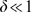 , then a protein of length shorter than
, then a protein of length shorter than 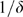 will be in the perturbative regime, and, therefore, a good
match between the true distribution and the pairwise distribution for such a
protein is virtually guaranteed.
will be in the perturbative regime, and, therefore, a good
match between the true distribution and the pairwise distribution for such a
protein is virtually guaranteed.
Fortunately, the properties of real proteins generally prevent this from
happening: at the majority of sites in a protein, the distribution of amino
acids is not sharply peaked around one amino acid. Even for
those sites that are sharply peaked (the evolutionarily-conserved sites), the
probability of the most likely amino acid,  , rarely exceeds 90% [21],[22]. This puts proteins consisting of only a few
amino acids out of the perturbative regime, and puts longer
proteins—the ones usually studied using pairwise models—well
out of it.
, rarely exceeds 90% [21],[22]. This puts proteins consisting of only a few
amino acids out of the perturbative regime, and puts longer
proteins—the ones usually studied using pairwise models—well
out of it.
This difference is fundamental: because many of the studies that have been carried out on neural data were in the perturbative regime, the conclusions of those studies—specifically, the conclusion that pairwise models provide accurate descriptions of large populations of neurons—is not yet supported. This is not the case for the protein studies, because they are not in the perturbative regime. Thus, the evidence that pairwise models provide accurate descriptions of protein folding remain strong and exceedingly promising.
Open questions
In our analysis, we sidestepped two issues of practical importance: finite sampling and alternative measures for assessing the quality of the pairwise model. These issues are beyond the scope of this paper, but in our view, they are natural next steps in the analysis of pairwise models. Below we briefly expand on them.
Finite sampling refers to the fact that in any real experiment, one has access to only a finite amount of data, and so does not know the true probability distribution of the spike trains. In our analysis, however, we assumed that one did have full knowledge of the true probability distribution. Since a good estimate of the probability distribution is crucial for assessing whether the pairwise model can be extrapolated to large populations, it is important to study how such estimates are affected by finite data. Future work is needed to address this issue, and to find ways to overcome data limitation—for example, by finding efficient methods for removing the finite data bias that affects information theoretic quantities such as the Kullback-Leibler divergence.
There are always many possible ways to assess the quality of a model. Our choice
of was motivated by two considerations: it is based on the
Kullback-Leibler divergence, which is a standard measure of
“distance” between probability distributions, and it is a
widely used measure in the field [7]–[10]. It
suffers, however, from a number of shortcomings. In particular, can be small even when the pairwise model assigns very
different probabilities to many of the configurations of the system. It would,
therefore, be important to study the quality of pairwise models using other
measures.
Methods
The behavior of the true entropy in the large limit
To understand how the true entropy behaves in the large limit, it is useful to express the difference of the entropies
as a mutual information. Using 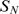 to denote the true entropy of neurons and
to denote the true entropy of neurons and 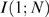 to denote the mutual information between one neuron and the
other neurons in a population of size
to denote the mutual information between one neuron and the
other neurons in a population of size  , we have
, we have
| (26) |
If knowing the activity of neurons does not fully constrain the firing of neuron  , then the single neuron entropy,
, then the single neuron entropy,  , will exceed the mutual information,
, will exceed the mutual information,  , and the entropy will be an increasing function of . For the entropy to be linear in , all we need is that the mutual information saturates with . Because of synaptic noise, this is a reasonable assumption
for networks of neurons: even if we observed all the spikes from all the
neurons, there would still be residual noise associated with synaptic failures,
jitter in release time, variability in the amount of neurotransmitter released,
stochastic channel dynamics, etc. Consequently, in the large limit, we may replace
, and the entropy will be an increasing function of . For the entropy to be linear in , all we need is that the mutual information saturates with . Because of synaptic noise, this is a reasonable assumption
for networks of neurons: even if we observed all the spikes from all the
neurons, there would still be residual noise associated with synaptic failures,
jitter in release time, variability in the amount of neurotransmitter released,
stochastic channel dynamics, etc. Consequently, in the large limit, we may replace 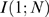 by its average, denoted
by its average, denoted  . Also replacing
. Also replacing  by its average, denoted
by its average, denoted 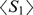 , we see that for large , the difference between
, we see that for large , the difference between  and
and  approaches a constant. Specifically,
approaches a constant. Specifically,
| (27) |
where this expression is valid in the large limit and the corrections are sublinear in .
Perturbative expansion
Our main quantitative result, given in Eqs. (8–10), is that the KL
divergence between the true distribution and both the independent and pairwise
distributions can be computed perturbatively as an expansion in powers of  in the limit
in the limit  . Here we carry out this expansion, and derive explicit
expressions for the quantities and .
. Here we carry out this expansion, and derive explicit
expressions for the quantities and .
To simplify our notation, here we use for the true distribution. The critical step in computing the
KL divergences perturbatively is to use the Sarmanov-Lancaster expansion [23]–[28] for ,
| (28) |
where
| (29a) |
| (29b) |
| (29c) |
| (29d) |
This expansion has a number of important, but not immediately obvious, properties. First, as can be shown by a direct calculation,
| (30) |
where the subscripts  and
and  indicate an average with respect to and
indicate an average with respect to and  , respectively. This has an immediate corollary,
, respectively. This has an immediate corollary,
This last relationship is important, because it tells us that if a product of  contains any terms linear in one of the
contains any terms linear in one of the  , the whole product averages to zero under the independent
distribution. This can be used to show that
, the whole product averages to zero under the independent
distribution. This can be used to show that
| (31) |
from which it follows that
Thus, is properly normalized. Finally, a slightly more involved
calculations provides us with a relationship between the parameters of the model
and the moments: for  ,
,
| (32a) |
| (32b) |
Virtually identical expressions hold for higher order moments. It is this last set of relationships that make the Sarmanov-Lancaster expansion so useful.
Note that Eqs. (32a) and (32b), along with the expression for the normalized correlation coefficients given in Eq. (16), imply that
| (33a) |
| (33b) |
These identities will be extremely useful for simplifying expressions later on.
Because the moments are so closely related to the parameters of the distribution,
moment matching is especially convenient: to construct a distribution whose
moments match those of up to some order, one simply needs to ensure that the
parameters of that distribution,  ,
,  ,
, 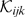 , etc., are identical to those of the true distributions up to
the order of interest. In particular, let us write down a new distribution,
, etc., are identical to those of the true distributions up to
the order of interest. In particular, let us write down a new distribution,  ,
,
| (34a) |
| (34b) |
We can recover the independent distribution by letting  , and we can recover the pairwise distribution by letting
, and we can recover the pairwise distribution by letting 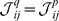 . This allows us to compute
. This allows us to compute 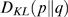 in the general case, and then either set
in the general case, and then either set  to zero or set
to zero or set 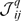 to
to 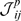 .
.
Expressions analogous to those in Eqs. (31–33) exist for averages with
respect to ; the only difference is that 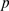 is replaced by
is replaced by  .
.
The KL divergence in the Sarmanov-Lancaster representation
Using Eqs. (28) and (34a) and a small amount of algebra, the KL divergence
between and may be written
| (35) |
where
| (36) |
To derive Eq. (35), we used the fact that  (see Eq. (31)). The extra term
(see Eq. (31)). The extra term 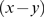 was included to ensure that
was included to ensure that 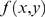 and its first derivatives vanish at
and its first derivatives vanish at  , something that greatly simplifies our analysis later on.
, something that greatly simplifies our analysis later on.
Our approach is to Taylor expand the right hand side of Eq. (35) around 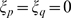 , compute each term, and then sum the whole
series (we do not assume that either
, compute each term, and then sum the whole
series (we do not assume that either  or
or  is small). Using
is small). Using 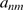 to denote the coefficients of the Taylor series, we have
to denote the coefficients of the Taylor series, we have
| (37) |
Note that because and its first derivatives vanish at 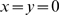 , all terms in this sum have
, all terms in this sum have  .
.
Because both 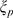 and
and  are themselves sums, an exact calculation of the terms in Eq.
(37) would be difficult. However, as we show below, in the section
“Averages of powers of ξp and
ξq” (see in particular
Eqs. (52) and (54)), they can be computed as perturbation expansions in powers
of , and the result is
are themselves sums, an exact calculation of the terms in Eq.
(37) would be difficult. However, as we show below, in the section
“Averages of powers of ξp and
ξq” (see in particular
Eqs. (52) and (54)), they can be computed as perturbation expansions in powers
of , and the result is
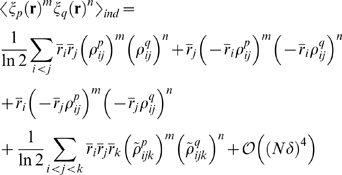 |
(38) |
where  and
and  are given by
are given by
| (39) |
 . The last equality in Eq. (39) follows from a small amount of
algebra and the definition of the correlation coefficients given in Eq. (16).
Equation (38) is valid only when
. The last equality in Eq. (39) follows from a small amount of
algebra and the definition of the correlation coefficients given in Eq. (16).
Equation (38) is valid only when  , which is the case of interest to us (since the Taylor
expansion of has only terms with
, which is the case of interest to us (since the Taylor
expansion of has only terms with 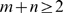 ).
).
The important point about Eq. (38) is that the  and
and  dependence follows that of the original Taylor expansion.
Thus, when we insert this equation back into Eq. (37), we recover our original
function—all we have to do is interchange the sums. For example,
consider inserting the first term in Eq. (38) into Eq. (37),
dependence follows that of the original Taylor expansion.
Thus, when we insert this equation back into Eq. (37), we recover our original
function—all we have to do is interchange the sums. For example,
consider inserting the first term in Eq. (38) into Eq. (37),
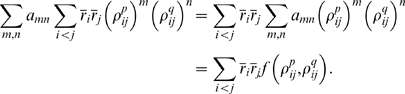 |
Performing the same set of manipulations on all of Eq. (38) leads to
 |
(40) |
This expression is true in general (except for some technical considerations; see
the section “Averages of powers of
ξp and
ξq”); to restrict it to the KL
divergences of interest we set to  and to either
and to either  or
or  . In the first case ( set to
. In the first case ( set to  ),
),  , which implies that
, which implies that  , and thus
, and thus  . Because has a quadratic minimum at
. Because has a quadratic minimum at  , when
, when  , the second two terms on the right hand side of Eq. (40) are
, the second two terms on the right hand side of Eq. (40) are  . We thus have, to lowest nonvanishing order in ,
. We thus have, to lowest nonvanishing order in ,
| (41) |
with the 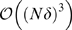 correction coming from the last sum in Eq. (40). Defining
correction coming from the last sum in Eq. (40). Defining
| (42) |
where, recall  , and inserting Eq. (42) into Eq. (41), we recover Eq. (8a).
, and inserting Eq. (42) into Eq. (41), we recover Eq. (8a).
In the second case ( set to  ), the first and second moments of
), the first and second moments of  and
and  are equal. This implies, using Eq. (32), that
are equal. This implies, using Eq. (32), that 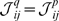 , and thus
, and thus 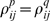 . Because
. Because 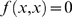 (see Eq. (36)), the first three terms on the right hand side
of Eq. (40)—those involving
(see Eq. (36)), the first three terms on the right hand side
of Eq. (40)—those involving  and but not —vanish. The next order term does not vanish, and yields
and but not —vanish. The next order term does not vanish, and yields
| (43) |
Defining
| (44) |
and inserting this expression into Eq. (43), we recover Eq. (8b).
External fields, pairwise couplings and moments
In this section we derive, to leading order in , expressions relating the external fields and pairwise
couplings of the maximum entropy model,  and
and 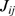 , to the first and second moments; these are the expressions
reported in Eq. (18). The calculation proceeds along the same lines as in the
previous section. There is, though, one extra step associated with the fact that
the quadratic term in the maximum entropy distribution given in Eq. (14) is
proportional to
, to the first and second moments; these are the expressions
reported in Eq. (18). The calculation proceeds along the same lines as in the
previous section. There is, though, one extra step associated with the fact that
the quadratic term in the maximum entropy distribution given in Eq. (14) is
proportional to 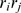 , not
, not  . However, to lowest order in , this doesn't matter. That's because
. However, to lowest order in , this doesn't matter. That's because
where 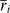 is defined as in Eq. (29d) except with
is defined as in Eq. (29d) except with 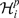 replaced by
replaced by 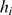 , and we used the fact that
, and we used the fact that  . The second term introduces a correction to the external
fields,
. The second term introduces a correction to the external
fields,  . However, the correction is
. However, the correction is  , so we drop it. We should keep in mind, though, that our final
expression for
, so we drop it. We should keep in mind, though, that our final
expression for  will have corrections of this order.
will have corrections of this order.
Using Eq. (14), but with  replaced by where it appears with
replaced by where it appears with 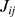 , we may write (after a small amount of algebra)
, we may write (after a small amount of algebra)
| (45) |
where  is the same as the function
is the same as the function  defined in Eq. (29a) except that
defined in Eq. (29a) except that  is replaced by
is replaced by 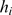 , the subscript “ind”
indicates, as usual, an average with respect to
, the subscript “ind”
indicates, as usual, an average with respect to 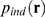 , and the two functions
, and the two functions 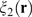 and
and 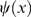 are defined by
are defined by
| (46) |
and
| (47) |
Given this setup, we can use Eqs. (55) and (56) below to compute the moments
under the maximum entropy model. That's because both 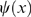 and its first derivative vanish at
and its first derivative vanish at  , which are the two conditions required for these equations to
be valid. Using also the fact that
, which are the two conditions required for these equations to
be valid. Using also the fact that 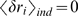 , Eqs. (55) and (56) imply that
, Eqs. (55) and (56) imply that
| (48a) |
| (48b) |
| (48c) |
where the first term in Eq. (48b) came from Eq. (29d) with  replaced by
replaced by  , the term “
, the term “ ” in Eq. (48c) came from
” in Eq. (48c) came from  , and for the second two expressions we used the fact that, to
lowest order in , the denominator in Eq. (45) is equal to 1.
, and for the second two expressions we used the fact that, to
lowest order in , the denominator in Eq. (45) is equal to 1.
Finally, using Eq. (19) for the normalized correlation coefficient, dropping the subscript “maxent” (since the first and second moments are the same under the maxent and true distributions), and inverting Eqs. (48b) and (48c) to express the external fields and coupling coefficients in terms of the first and second moments, we arrive at Eq (18).
Averages of powers of  and
and 
Here we compute  , which, as can be seen in Eq. (37), is the key quantity in our
perturbation expansion. Our starting point is to (formally) expand the sums that
make up
, which, as can be seen in Eq. (37), is the key quantity in our
perturbation expansion. Our starting point is to (formally) expand the sums that
make up 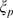 and
and 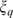 (see Eqs. (29b) and (34b)), which yields
(see Eqs. (29b) and (34b)), which yields
 |
(49) |
The sum over  is a sum over all possible configurations of the
is a sum over all possible configurations of the 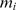 . The coefficient
. The coefficient  are complicated functions of the
are complicated functions of the  , etc. Computing these functions is straightforward, although
somewhat tedious, especially when
, etc. Computing these functions is straightforward, although
somewhat tedious, especially when  is large; below we compute them only for
is large; below we compute them only for  and 3. The reason
and 3. The reason 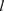 starts at 2 is that
starts at 2 is that 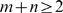 ; see Eq. (37).
; see Eq. (37).
We first show that all terms with superscript  are
are  . To do this, we note that, because the right hand side of Eq.
(49) is an average with respect to the independent distribution, the average of
the product is the product of the averages,
. To do this, we note that, because the right hand side of Eq.
(49) is an average with respect to the independent distribution, the average of
the product is the product of the averages,
| (50) |
Then, using the fact that 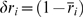 with probability
with probability  and
and  with probability
with probability  (see Eq. (29c)), we have
(see Eq. (29c)), we have
 |
(51) |
The significance of this expression is that, for  ,
,  , independent of
, independent of  . Consequently, if all the
. Consequently, if all the  in Eq. (50) are greater than 1, then the right hand side is
in Eq. (50) are greater than 1, then the right hand side is 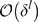 . This shows that, as promised above, the superscript
. This shows that, as promised above, the superscript  labels the order of the terms.
labels the order of the terms.
As we saw in the section “The KL divergence in the Sarmanov-Lancaster
representation”, we need to go to third order in , which means we need to compute the terms on the right hand
side of Eq. (49) with  and 3. Let us start with
and 3. Let us start with 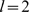 , which picks out only those terms with two unique indices.
Examining the expressions for
, which picks out only those terms with two unique indices.
Examining the expressions for 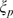 and
and  given in Eqs. (29b) and (34b), we see that we must keep only
terms involving , since has three indices, and higher order terms have more. Thus, the
given in Eqs. (29b) and (34b), we see that we must keep only
terms involving , since has three indices, and higher order terms have more. Thus, the  contribution to the average in Eq. (49), which we denote
contribution to the average in Eq. (49), which we denote  , is given by
, is given by
Pulling  and
and  out of the averages, using Eq. (33a) to eliminate
out of the averages, using Eq. (33a) to eliminate  and
and  in favor of
in favor of  and
and  , and applying Eq. (51) (while throwing away some of the terms
in that equation that are higher than second order in ), the above expression may be written
, and applying Eq. (51) (while throwing away some of the terms
in that equation that are higher than second order in ), the above expression may be written
 |
(52) |
Note that we were not quite consistent in our ordering with respect to , in the sense that we kept some higher order terms and not
others. We did this so that we could use rather than , as the former is directly observable.
For 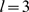 the calculation is more involved, but not substantially so.
Including terms with exactly three unique indices in the sum on the right hand
side of Eq. (49) gives us
the calculation is more involved, but not substantially so.
Including terms with exactly three unique indices in the sum on the right hand
side of Eq. (49) gives us
 |
(53) |
This expression is not quite correct, since some of the terms contain only two
unique indices—these are the terms proportional to  —whereas it should contain only terms with exactly
three unique indices. Fortunately, this turns out not to matter, for reasons we
discuss at the end of the section.
—whereas it should contain only terms with exactly
three unique indices. Fortunately, this turns out not to matter, for reasons we
discuss at the end of the section.
To perform the averages in Eq. (53), we would need to use multinomial expansions,
and then average over the resulting powers of 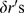 . For the latter, we can work to lowest order in , which means we only take the first term in Eq. (51). This
amounts to replacing every with
. For the latter, we can work to lowest order in , which means we only take the first term in Eq. (51). This
amounts to replacing every with 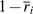 (and similarly for and ), and in addition multiplying the whole expression by an
overall factor of
(and similarly for and ), and in addition multiplying the whole expression by an
overall factor of  . For example, if
. For example, if  and
and 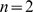 , one of the terms in the multinomial expansion is
, one of the terms in the multinomial expansion is 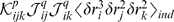 . This average would yield, using Eq. (51) and considering only
the lowest order term,
. This average would yield, using Eq. (51) and considering only
the lowest order term,  .
.
This procedure also is not quite correct, since terms with only one factor of , which average to zero, are replaced with 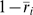 . This also turns out not to matter; again, we discuss why at
the end of the section.
. This also turns out not to matter; again, we discuss why at
the end of the section.
We can, then, go ahead and use the above “replace blindly”
algorithm. Note that the factors of  ,
, 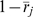 and
and 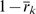 turn and into normalized correlation coefficients (see Eq. (33)), which
considerably simplifies our equations. Using also Eq. (39) for
turn and into normalized correlation coefficients (see Eq. (33)), which
considerably simplifies our equations. Using also Eq. (39) for  , Eq. (53) becomes
, Eq. (53) becomes
| (54) |
We can now combine Eqs. (52) and (54), and insert them into Eq. (49). This gives
us the first two terms in the perturbative expansion of  ; the result is written down in Eq. (38) above.
; the result is written down in Eq. (38) above.
Why can we ignore the overcounting associated with terms in which an index
appears exactly zero or one times? We clearly can't do this in general,
because for such terms, replacing with  fails—either because the terms didn't exist
in the first place (when one of the indices never appeared) or because they
averaged to zero (when an index appeared exactly once). In our case, however,
such terms do not appear in the Taylor expansion. To see why, note first of all
that, because of the form of , its Taylor expansion can be written
fails—either because the terms didn't exist
in the first place (when one of the indices never appeared) or because they
averaged to zero (when an index appeared exactly once). In our case, however,
such terms do not appear in the Taylor expansion. To see why, note first of all
that, because of the form of , its Taylor expansion can be written  where
where 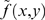 is finite at
is finite at  (see Eq. (36)). Consequently, the original Taylor expansion of
(see Eq. (36)). Consequently, the original Taylor expansion of  , Eq. (37), should contain a factor of
, Eq. (37), should contain a factor of  ; i.e.,
; i.e.,
where the  are the coefficients of the Taylor expansion of
are the coefficients of the Taylor expansion of 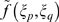 . The factor
. The factor  , when expanded, has the form
, when expanded, has the form
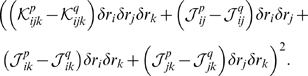 |
As we saw in the previous section, we are interested in the third order term only
to compute  , for which
, for which  . Therefore, the above multiplicative factor reduces to
. Therefore, the above multiplicative factor reduces to 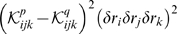 . It is that last factor of
. It is that last factor of 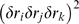 that is important, since it guarantees that for every term in
the Taylor expansion, all indices appear at least twice. Therefore, although Eq.
(53) is not true in general, it is valid for our analysis.
that is important, since it guarantees that for every term in
the Taylor expansion, all indices appear at least twice. Therefore, although Eq.
(53) is not true in general, it is valid for our analysis.
We end this section by pointing out that there is a very simple procedure for
computing averages to second order in . Consider a function 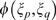 that has a minimum at
that has a minimum at  and also obeys
and also obeys 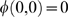 . Then, based on the above analysis, we have
. Then, based on the above analysis, we have
| (55) |
Two easy corollaries of this are: for and  positive integers,
positive integers,
| (56a) |
| (56b) |
where the sum in Eq. (56a) runs over only, and we used the fact that both  and
and  are symmetric with respect to the interchange of
are symmetric with respect to the interchange of  and .
and .
Generating synthetic data
As can be seen in Eq. (13), the synthetic data depends on three sets of
parameters: 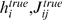 , and . Here we describe how they were generated.
, and . Here we describe how they were generated.
To generate the 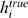 , we draw a set of firing rates,
, we draw a set of firing rates,  , from an exponential distribution with mean 0.02 (recall that
, from an exponential distribution with mean 0.02 (recall that  , which we set to 15, is the number of neurons in our base
distribution). From this we chose the external field according to Eq. (18a),
, which we set to 15, is the number of neurons in our base
distribution). From this we chose the external field according to Eq. (18a),
In the perturbative regime, a distribution generated with these values of the
external fields has firing rates approximately equal to the 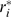 ; see Eq. (18a) and Fig. 6.
; see Eq. (18a) and Fig. 6.
To generate the  and , we drew them from Gaussian distributions with means equal to
0.05 and 0.02 and standard deviations of 0.8 and 0.5, respectively. Using
non-zero values for means that the true distribution is not pairwise.
and , we drew them from Gaussian distributions with means equal to
0.05 and 0.02 and standard deviations of 0.8 and 0.5, respectively. Using
non-zero values for means that the true distribution is not pairwise.
Bin size and the correlation coefficients
One of our main claims is that is linear in bin size, . This is true, however, only if and are independent of , as can be seen from Eq. (10b). In this section we show that
independence is satisfied if  is smaller than the typical correlation time of the responses.
For larger than such correlation times, and do depend on , and is no longer linear in . Note, though, that the correlation time is always finite, so
there will always be a bin size below which the linear relationship,
is smaller than the typical correlation time of the responses.
For larger than such correlation times, and do depend on , and is no longer linear in . Note, though, that the correlation time is always finite, so
there will always be a bin size below which the linear relationship,  , is guaranteed.
, is guaranteed.
Examining Eqs. (42) and (44), we see that and depend on the normalized correlation coefficients, and (we drop superscripts, since our discussion will be generic).
Thus, to understand how and depend on bin size, we need to understand how the normalized
correlation coefficients depend on bin size. To do that, we express them in
terms of standard cross-correlograms, as the cross-correlograms contain, in a
very natural way, information about the temporal timescales in the spike train.
We start with the second order correlation coefficient, since it is simplest. The
corresponding cross-correlogram, which we denote  , is given by
, is given by
| (57) |
where  is the time of the k
th spike on
neuron
is the time of the k
th spike on
neuron  (and similarly for
(and similarly for  ), and
), and  is the Dirac
is the Dirac  . The normalization in Eq. (57) is slightly
non-standard—more typical is to divide by something with units of
firing rate (
. The normalization in Eq. (57) is slightly
non-standard—more typical is to divide by something with units of
firing rate (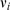 ,
,  or
or  ), to give units of spikes/s. The normalization we use is
convenient, however, because in the limit of large
), to give units of spikes/s. The normalization we use is
convenient, however, because in the limit of large  , approaches one.
, approaches one.
It is slightly tedious, but otherwise straightforward, to show that when is sufficiently small that only one spike can occur in a time
bin, is related to via
| (58) |
The (unimportant) factor  comes from the fact that the spikes occur at random locations
within a bin.
comes from the fact that the spikes occur at random locations
within a bin.
Equation (58) has a simple interpretation: is the average height of the central peak of the
cross-correlogram relative to baseline. How strongly depends on is thus determined by the shape of the cross-correlogram. If
it is smooth, then approaches a constant as becomes small. If, on the other hand, there is a sharp peak at 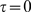 , then
, then 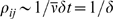 for small , so long as is larger than the width of the peak. (The factor of included in the scaling is approximate; it is a placeholder
for an effective firing rate that depends on the indices
for small , so long as is larger than the width of the peak. (The factor of included in the scaling is approximate; it is a placeholder
for an effective firing rate that depends on the indices  and . It is, however, sufficiently accurate for our purposes.) A
similar relationship exists between the third order correlogram and the
correlation coefficient. Thus, is also independent of in the small limit, whereas if the central peak is sharp it scales as
and . It is, however, sufficiently accurate for our purposes.) A
similar relationship exists between the third order correlogram and the
correlation coefficient. Thus, is also independent of in the small limit, whereas if the central peak is sharp it scales as  .
.
The upshot of this analysis is that the shape of the cross-correlogram has a
profound effect on the correlation coefficients and, therefore, on . What is the shape in real networks? The answer typically
depends on the physical distance between cells. If two neurons are close, they
are likely to receive common input and thus exhibit a narrow central peak in
their cross-correlogram. Just how narrow depends on the area. Early in the
sensory pathways, such as retina [29]–[31] and LGN [32], peaks
can be very narrow—on the order of milliseconds. Deeper into cortex,
however, peaks tend to broaden, to at least tens of milliseconds [33],[34].
If, on the other hand, the neurons are far apart, they are less likely to
receive common input. In this case, the correlations come from external stimuli,
so the central peak tends to have a characteristic width given by the temporal
correlation time of the stimulus, typically 100 s of milliseconds.
Although clearly both kinds of cross-correlograms exist in any single population
of neurons, it is convenient to analyze them separately. We have already
considered networks in which the cross-correlograms were broad and perfectly
flat, so that the correlation coefficients were strictly independent of bin
size. We can also consider the opposite extreme: networks in which the the
cross-correlograms (both second and higher order) among nearby neurons exhibit
sharp peaks while those among distant neurons are uniformly equal to 1. In this
regime, the correlation coefficients depend on : as discussed above, the second order ones scale as  and the third as
and the third as  . This means that the arguments of
. This means that the arguments of 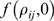 and
and 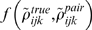 are large (see Eqs. (42) and (44)). From the definition of in Eq. (36), in this regime both are approximately linear in
their arguments (ignoring log corrections). Consequently,
are large (see Eqs. (42) and (44)). From the definition of in Eq. (36), in this regime both are approximately linear in
their arguments (ignoring log corrections). Consequently,  and
and  . This implies that and scale as and
. This implies that and scale as and  , respectively, and so
, respectively, and so  , independent of . Thus, if the bin size is large compared to the correlation
time, will be approximately independent of bin size.
, independent of . Thus, if the bin size is large compared to the correlation
time, will be approximately independent of bin size.
Extending the normalized distance measure to the time domain
In this section we derive the expression for given in Eq. (25). Our starting point is its definition, Eq.
(24). It is convenient to define  to be a concatenation of the responses in time bins,
to be a concatenation of the responses in time bins,
| (59) |
where, as in the section “Is there anything wrong with
using small time bins?”, the superscript labels time, so  is the full, temporally correlated, distribution.
is the full, temporally correlated, distribution.
With this definition, we may write the numerator in Eq. (24) as
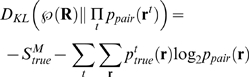 |
(60) |
where  is the entropy of , the last sum follows from a marginalization over all but one
element of , and
is the entropy of , the last sum follows from a marginalization over all but one
element of , and  is the true distribution at time
is the true distribution at time  (unlike in the section “Is there anything wrong with
using small time bins?”, here we do not assume that the true
distribution is the same in all time bins). Note that
(unlike in the section “Is there anything wrong with
using small time bins?”, here we do not assume that the true
distribution is the same in all time bins). Note that  is independent of time, since it is computed from a time
average of the true distribution. That time average, which we call
is independent of time, since it is computed from a time
average of the true distribution. That time average, which we call  , is given in terms of
, is given in terms of  as
as
Inserting this definition into Eq. (60) eliminates the sum over  , and replaces it with
, and replaces it with 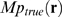 . For simplicity we consider the maximum entropy pairwise
model. In this case, because
. For simplicity we consider the maximum entropy pairwise
model. In this case, because 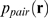 is in the exponential family, and the first and second moments
are the same under the true and maximum entropy distributions, we can replace
is in the exponential family, and the first and second moments
are the same under the true and maximum entropy distributions, we can replace  with
with  . Consequently, Eq. (60) becomes
. Consequently, Eq. (60) becomes
This gives us the numerator in the expression for (Eq. (24)); using Eq. (4) to write 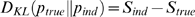 , the full expression for becomes
, the full expression for becomes
| (61) |
where we added and subtracted  to the numerator.
to the numerator.
The first term on the right hand side of Eq. (61) we recognize, from Eq. (6), as . To cast the second into a reasonable form, we define  to be the entropy of the distribution that retains the
temporal correlations within each neuron but is independent across neurons.
Then, adding and subtracting this quantity to the numerator in Eq. (61), and
also adding and subtracting
to be the entropy of the distribution that retains the
temporal correlations within each neuron but is independent across neurons.
Then, adding and subtracting this quantity to the numerator in Eq. (61), and
also adding and subtracting 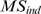 , we have
, we have
| (62) |
The key observation is that if  , then
, then
Comparing this with Eqs. (8a) and (9a), we see that 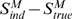 is a factor of
is a factor of  times larger than
times larger than  . We thus have
. We thus have
| (63) |
Again assuming  , and defining
, and defining 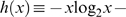
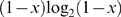 , the last term in this expression may be written
, the last term in this expression may be written
 |
(64) |
Inserting this into Eq. (63) and using Eqs. (4), (8a) and (9a) yields Eq. (25).
We have assumed here that  ; what happens when
; what happens when  , or larger? To answer this, we rewrite Eq. (61) as
, or larger? To answer this, we rewrite Eq. (61) as
| (65) |
We argue that in general, as increases,  becomes increasingly different from
becomes increasingly different from 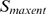 , since the former was derived under the assumption that the
responses at different time bins were independent. Thus, Eq. (25) should be
considered a lower bound on .
, since the former was derived under the assumption that the
responses at different time bins were independent. Thus, Eq. (25) should be
considered a lower bound on .
Footnotes
The authors have declared that no competing interests exist.
YR and PEL were supported by the Gatsby Charitable Foundation (http://www.gatsby.org.uk) and by the US National Institute of Mental Health grant R01 MH62447. SN was supported by the US National Eye Institute grant R01 EY12978. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Rieke F, Warland D, de Ruyter van Steveninck R, Bialek W. Spikes: exploring the neural code. Cambridge, MA: MIT Press; 1997. [Google Scholar]
- 2.Russ W, Lowery D, Mishra P, Yaffe M, Ranganathan R. Natural-like function in artificial WW domains. Nature. 2005;437:579–583. doi: 10.1038/nature03990. [DOI] [PubMed] [Google Scholar]
- 3.Socolich M, Lockless S, Russ W, Lee H, Gardner K, et al. Evolutionary information for specifying a protein fold. Nature. 2005;437:512–518. doi: 10.1038/nature03991. [DOI] [PubMed] [Google Scholar]
- 4.Oates J. Food distribution and foraging behavior. In: Smuts B, Cheney D, Seyfarth R, Wrangham R, Struhsaker T, editors. Primate societies. Chicago: University of Chicago Press; 1987. pp. 197–209. [Google Scholar]
- 5.Wrangham R. Evolution of social structure. In: Smuts B, Cheney D, Seyfarth R, Wrangham R, Struhsaker T, editors. Primate societies. Chicago: University of Chicago Press; 1987. pp. 282–298. [Google Scholar]
- 6.Eisenberg J, Muckenhirn N, Rundran R. The relation between ecology a social structure in primates. Science. 1972;176:863–874. doi: 10.1126/science.176.4037.863. [DOI] [PubMed] [Google Scholar]
- 7.Schneidman E, Berry M, Segev R, Bialek W. Weak pairwise correlations imply strongly correlated network states in a neural population. Nature. 2006;440:1007–1012. doi: 10.1038/nature04701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shlens J, Field G, Gauthier J, Grivich M, Petrusca D, et al. The structure of multi-neuron firing patterns in primate retina. J Neurosci. 2006;26:8254–8266. doi: 10.1523/JNEUROSCI.1282-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tang A, Jackson D, Hobbs J, Chen W, Smith J, et al. A maximum entropy model applied to spatial and temporal correlations from cortical networks in vitro. J Neurosci. 2008;28:505–518. doi: 10.1523/JNEUROSCI.3359-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bethge M, Berens P. Near-maximum entropy models for binary neural representations of natural images. In: Platt J, Koller D, Singer Y, Roweis S, editors. Advances in Neural Information Processing Systems 20. Cambridge, MA: MIT Press; 2008. pp. 97–104. [Google Scholar]
- 11.Yu S, Huang D, Singer W, Nikolic D. A small world of neuronal synchrony. Cereb Cortex. 2008;18:2891–2901. doi: 10.1093/cercor/bhn047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kullback S, Leibler R. On information and sufficiency. Ann Math Stat. 1951;22:79–86. [Google Scholar]
- 13.Friedman N, Mosenzon O, Slonim N, Tishby N. Proc. of Uncertainty in Artificial Intelligence (UAI-17) San Mateo, CA: Morgan Kaufmann Publishers; 2001. Multivariate information bottleneck. pp. 152–161. [Google Scholar]
- 14.Slonim N, Friedman N, Tishby N. Multivariate information bottleneck. Neural Comput. 2006;18:1739–1789. doi: 10.1162/neco.2006.18.8.1739. [DOI] [PubMed] [Google Scholar]
- 15.Shannon C, Weaver W. The mathematical theory of communication. Urbana, Illinois: University of Illinois Press; 1949. [Google Scholar]
- 16.Cover T, Thomas J. Elements of information theory. New York, NY: John Wiley & Sons; 1991. [Google Scholar]
- 17.Sessak V, Monasson R. Small-correlation expansions for the inverse ising problem. J Phys A. 2009;42:055001. [Google Scholar]
- 18.Amari S. Measure of correlation orthogonal to changing in firing rate. Neural Comput. 2009;21:960–972. doi: 10.1162/neco.2008.03-08-729. [DOI] [PubMed] [Google Scholar]
- 19.Shlens J, Field G, Gauthier J, Greschner M, Sher A, et al. Spatial organization of large-scale concerted activity in the primate retina. J Neurosci. In Press 2009 [Google Scholar]
- 20.Dill K. Theory for the folding and stability of globular proteins. Biochemistry. 1985;24:1501–1509. doi: 10.1021/bi00327a032. [DOI] [PubMed] [Google Scholar]
- 21.Lockless S, Ranganathan R. Evolutionarily conserved pathways of energetic connectivity in protein families. Science. 1999;286:295–299. doi: 10.1126/science.286.5438.295. [DOI] [PubMed] [Google Scholar]
- 22.Vargas-Madrazo E, Lara-Ochoa F, Jiménez-Montaño M. A skewed distribution of amino acids at recognition sites of the hypervariable region of immunoglobulins. J Mol Evol. 1994;38:100–104. doi: 10.1007/BF00175497. [DOI] [PubMed] [Google Scholar]
- 23.Sarmanov O. Maximum correlation coeffcient (nonsymmetric case). Selected Translations in Mathematical Statistics and Probability. Amer. Math. Soc. Volume 2. 1962. pp. 207–210.
- 24.Sarmanov O. Maximum correlation coefficient (nonsymmetric case). Selected Translations in Mathematical Statistics and Probability. Amer. Math. Soc. Volume 4. 1963. pp. 271–275.
- 25.Lancaster H. The structure of bivariate distributions. Ann Math Stat. 1958;29:719–736. [Google Scholar]
- 26.Lancaster H. Correlation and complete dependence of random variables. Ann Math Stat. 1963;34:1315–1321. [Google Scholar]
- 27.Bahadur R. A representation of the joint distribution of responses to n dichotomous items. In: Solomon H, editor. Studies in Item Analysis and Prediction. Stanford University Press; 1961. pp. 158–168. [Google Scholar]
- 28.Johnson D, Goodman I. Inferring the capacity of the vector Poisson channel with a Bernoulli model. Network. 2008;19:13–33. doi: 10.1080/09548980701656798. [DOI] [PubMed] [Google Scholar]
- 29.Mastronarde D. Correlated firing of cat retinal ganglion cells. I. spontaneously active inputs to X- and Y-cells. J Neurophysiol. 1983;49:303–324. doi: 10.1152/jn.1983.49.2.303. [DOI] [PubMed] [Google Scholar]
- 30.DeVries S. Correlated firing in rabbit retinal ganglion cell. J Neurophysiol. 1999;81:908–920. doi: 10.1152/jn.1999.81.2.908. [DOI] [PubMed] [Google Scholar]
- 31.Nirenberg S, Carcieri S, Jacobs A, Latham P. Retinal ganglion cells act largely as independent encoders. Nature. 2001;411:698–701. doi: 10.1038/35079612. [DOI] [PubMed] [Google Scholar]
- 32.Dan Y, Alonso J, Usrey W, Reid R. Coding of visual information by precisely correlated spikes in the lateral geniculate nucleus. Nat Neurosci. 1998;1:501–507. doi: 10.1038/2217. [DOI] [PubMed] [Google Scholar]
- 33.Ts'o D, Gilbert C, Wiesel T. Relationships between horizontal interactions and functional architecture in cat striate cortex as revealed by cross-correlation analysis. J Neurosci. 1986;6:1160–1170. doi: 10.1523/JNEUROSCI.06-04-01160.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nelson J, Salin P, Munk M, Arzi M, Bullier J. Spatial and temporal coherence in cortico-cortical connections: a cross-correlation study in areas 17 and 18 in the cat. Vis Neurosci. 1992;9:21–37. doi: 10.1017/s0952523800006349. [DOI] [PubMed] [Google Scholar]