

Abstract
AIM: To determine platelet activating factor (PAF) receptor expression in cirrhotic hepatic stellate cells.
METHODS: Hepatic stellate cells, isolated from the livers of control and CCl4-induced cirrhotic rats, were placed in serum-free medium after overnight culture. We determined the PAF receptor in hepatic stellate cells by saturation binding technique and semiquantitative reverse transcriptase polymerase chain reaction (RT-PCR), and the effects of PAF and its antagonist BN52021 on prostaglandin E2 (PGE2) release by stellate cells.
RESULTS: Scatchard analysis indicated the presence of PAF receptor with dissociation constant (Kd) of 4.66 nmol/L and maximum binding capacity (Bmax) of 24.65 fmol/μg in cirrhotic stellate cells. Compared with the control, the maximum PAF binding capacity increased significantly (Bmax: 24.65 ± 1.96 fmol/μg. DNA, R = 0.982 vs 5.74 ± 1.55 fmol/μg. DNA, R = 0.93; P < 0.01), whereas receptor affinity had no significant difference (Kd of 4.66 ± 0.33 nmol/L for the cirrhosis and 3.51 ± 0.26 nmol/L for the control; P > 0.05). Consistent with the receptor binding data, the mRNA expression of PAF receptor was increased significantly in cirrhotic stellate cells. PAF in a concentration-dependent manner induced PGE2 synthesis in cirrhotic hepatic stellate cells, but the effects were blocked significantly by BN52021.
CONCLUSION: Cirrhosis sensitizes hepatic stellate cells to PAF by elevating its receptor level and hepatic stellate cells maybe potential effectors of PAF induced portal hypertension.
Keywords: Platelet activating factor, Hepatic stellate cells, Kupffer cells, Cirrhosis, Receptor
INTRODUCTION
Platelet activating factor, a phospholipid, exhibits diverse biological activities[1–4]. Portal venous administration of platelet activating factor (PAF) in isolated perfused rat liver increases portal venous pressure and glycogenolysis[5,6]. Systemic (intravenous) administration of PAF to rats, in vivo, also induces these effects and, in addition, causes immediate decrease in arterial blood pressure[7]. As a potent hepatic vasoconstrictor and systemic vasodilator, PAF is implicated in portal hypertension and hyperdynamic circulation associated with liver cirrhosis. In a parallel study, we observed increased hepatic and circulating PAF concentrations in cirrhotic rats[8–11]. The increased hepatic synthesis of PAF is a major source of elevated circulating PAF and upregulates hepatic hemodynamic abnormalities, which contributes to portal hypertension[8–11]. The increased hepatic PAF in the cirrhotic liver due to its enhanced synthesis by Kupffer cells, Kupffer cells are a major source of increased platelet activating factor[12,13]. The increased portal pressure by endogenous PAF in cirrhosis can be decreased by BN52021 in some extent. Reduction in portal pressure by BN52021 measures the contribution of endogenous PAF[9,11]. In control rats, this contribution to basal pressure is zero but in cirrhotic rats endogenous PAF is responsible for 23% of the cirrhosis dependent increase in portal pressure[9]. The potential effectors of PAF induced increase in portal pressure remains undetermined. The smooth muscle component of the hepatic vasculature may contribute to PAF induced increase in portal pressure[14]. Hepatic stellate cells (HSCs), located in the space of Disse, play a crucial role in the development of liver fibrosis[15,16]. As PAF is one of only six mediators currently known to cause coupled Ca2+ increase and contraction in HSCs[17], we hypothesized the HSCs may be potential effectors of PAF induced increases in portal pressure. Therefore, PAF receptors and PAF-induced prostaglandin E2 release derived from cirrhotic hepatic stellate cells were investigated.
MATERIALS AND METHODS
Induction of cirrhosis
The experimental protocols were approved by the Institutional Animal Care and Use Committee of the Institute Infection Diseases of PLA in accordance with the guidelines of the National Institutes of Medicine. Cirrhosis was induced in male Sprague-Dawley rats (230-250 g) as described previously[13,18,19]. Briefly, intraperitoneal injections of CCl4 (0.15 mL/kg WT twice a week for 8 wk) were combined with drinking water containing phenobarbital (0.4 g/L). Control rats received injections of the vehicle (peanut oil) and phenobarbital water.
Morphometric analysis
An established histological grading system was employed for determination of pathological scores of liver injury[18–23]. Paraffin-embedded liver sections of 4 micron thickness were stained with hematoxylin-eosin and Masson’s trichrome stain. Steatosis, inflammation, necrosis and fibrosis were scored in at least 3 random fields of view in each tissue section and a score for each specific parameter was estimated. Steatosis was assessed by estimating the percentage of cells with micro- and macrovesicular fat as follows: 0 (absent); 1 (1%-25%); 2 (26%-50%); 3 (51%-75%); 4 (76%-100%). Necrosis was scored as follows: 0 (absent); 1 (1-10 necrotic cells per view); 2 (11-20 necrotic cells per view); 3 (21-30 necrotic cells per view); 4 (31-40 necrotic cells per view); 5 (41-50 necrotic cells per view). Inflammation was assessed as follows: 0 (absent); 1 (rare); 2 (scattered); 3 (scatted with localized foci); 4 (abundant with foci); 5 (extensive). Architectural change, fibrosis and cirrhosis were estimated as follows: 0 (absent); 1 (rare); 2 (scattered deposition); 3 (scatted with localized deposition); 4 (abundant with minor bridging fibrosis); 5 (bridging fibrosis); 6 (cirrhosis). A total pathology score was calculated by combining and summing the score for the above pathological parameters, and the average score per animal/treatment group was calculated.
Preparation of hepatic stellate cells
Hepatic stellate cells were prepared essentially as described previously[24,25]. Briefly, cirrhotic rats (350-380 g) were digested with protease (0.05%) and collagenase (0.025%). After removal of hepatocytes and cell debris by low speed centrifugation, hepatic stellate cells were purified by density gradient centrifugation on a 28.7% (W/V) Nycodenz gradient. The cells were suspended in DEME containing 10% fetal bovine serum and 10% horse serum, antibiotics, and plated in at a density of 0.5 × 106 cells/cm2. The cells were used for experiments after an overnight incubation. The purity of the cells was determined by light microscopy and vitamin A autofluorescence. Further characterization was conducted immunohistochemically with specific markers for stellate cells (anti-desmin and -α-smooth muscle actin antibodies; DAKO) as described previously[25].
Determination of PAF receptors mRNA in hepatic stellate cells
The levels of the low abundance mRNA transcripts of PAF receptor were determined by semiquantitative reverse transcriptase polymerase chain reaction (RT-PCR) essentially as described previously[8–13,26] β-actin mRNA expression was determined using the same amount of cDNA. RNA was isolated from the livers using an RNA isolation kit (ToTALLY RNATM, Ambion, Austin, TX) as per the instructions of the manufacturer. The concentration of RNA was determined by measuring A260, and the purity was checked by the A260/A280 ratio (greater than 1.8). 2 μg total RNA obtained from hepatic stellate cells was used for the preparation of cDNA by reverse transcriptase-PCR. The PCR Primers for PAF cDNA: 5’-GCCACAACACAGAGGCTTGA-3’(F) and 5’-TCCATTGCTCTGGGCAGGAA-3’(R) (121 bp); β-actin cDNA: 5’-TTCTACAATGAGCTGCGTGTG-3’(F) and 5’-TTCATGGATGCCACAGGATTC-3’(R) (561 bp) were used. cDNA equivalent of 5 ng of original RNA was used in PCR. The reaction mixture (50 μL) contained 10 mmol/L Tris-HCl (pH 8.3), 50 mmol/L KCl, 1.5 mmol/L MgCl2, 0.2 mmol/L dNTPs, 20 pmol of PCR primer and 2 units of Platinum Taq DNA polymerase (GIBCO-Invitrogen, Carlsbad, CA). The reaction for PAF receptor was run for 35 cycles as follow: denaturation at 94°C for 1 min, annealing at 60°C for 30 s, extension at 72°C for 30 s. The PCR products were resolved in a 2.5% agarose gel and stained with 1X SYBR GreenI(FMC Biproduct, Rockland, ME). The gels were scanned under blue fluorescence light using a phosphorimager and the band intensity was quantified using ImageQuant software (Molecular Dynamics, Sunnyvale, CA, USA).
Determination of PAF receptor in hepatic stellate cells
The assay was performed as previously described[8–13,27,28]. After hepatic stellate cells incubated overnight were washed twice time with HBSS containing 10 mmol/L HEPES, pH7.4, and 0.1% bovine serum albumin (HBSS/BSA), and then incubated in 50 mmol/L Tris-HCl (pH 7.2 containing 5 mmol/L MgCl2, 125 mmol/L choline chloride, 0.25% BSA) and 0.0125-3.2 nmol/L 1-O-[3H]octa-decyl-2-O-acetyl-sn-glycero-3-phosphocholine (151 Ci/mmol, 9.96 GBq/mg; Amersham) ± 10 μmol/L unlabelled PAF (1-O-hexadecyl-2-O-acetyl-sn-glycero-3-phosphocholine; Bachem Americas, King of Prussia, PA) at 22°C for 3 h, the reaction was terminated with the addition of 5 mL ice-cold assay buffer. The cells were washed twice with assay buffer and then digested with 5% sodium dodecyl sulfate kept at 60°C for 30 min. Radioactivity was determined in β-cintillation. The results were expressed as CPM of 3H -PAF bound per μg DNA.
Determination of prostaglandin E2 (PGE2)
PGE2 was extracted from the culture medium with or without BN52021, and analyzed by ELISA with a kit from Amersham Pharmacia biotech, Inc. NJ, USA., following protocol 1 of the manufacturer’s instructions.
Statistical analysis
The values are presented as averages of triplicate determinations ± SEM. Each experiment was repeated at least three times on cells from different animals. Student’s t-test was employed for statistical comparison of the paired samples. A P value of < 0.05 was considered statistically significant.
RESULTS
General characteristics of cirrhosis
There were no notable changes in the liver histology through 8 wk in control rats (Figure 1A1). In contrast CCl4-treated rats demonstrated extensive changes in liver morphology, including steatosis, inflammation, hepatocyte ballooning and necrosis. There was extensively distorted architecture due to excessive deposition of extracellular matrix that caused bridging fibrosis, and infiltration of inflammatory cells in the sinusoids and their accumulation around the islands of hepatocytes (Figure 1A3). Hepatic fibrosis was further validated histologically using Masson's trichrome stain (Figure 1A2 and 1A4).
Figure 1.

Morphometric analysis of the cirrhotic liver. A: After 8 wk of CCl4 or vehicle treatment, liver tissue was fixed and stained with hematoxylin/eosin (A1 and A3) as well as Masson's trichrome stain (A2 and A4); B: Necroinflammatory score (NFS) and scores for architectural change, fibrosis and cirrhosis (AFCS) were determined as described in the Methods section. bP < 0.01 vs control.
Characterization of HSCs isolated from normal and cirrhotic livers
HSCS viability was more than 95%. The desmin positive cell rate was (94.3% ± 3.8%). HSCs isolated from cirrhotic livers showed morphological characteristics of myofibroblasts with a spindle-like shape and low vitamin A droplets content. These cells highly expressed aSMA (Figure 2). In contrast, HSCs freshly isolated from normal rat livers showed a quiescent phenotype consisting of an oval shape and numerous vitamin A-rich droplets. These cells barely expressed aSMA.
Figure 2.

Expression of desmin and aSMA in HSCs freshly isolated from cirrhotic livers. HSCs from cirrhotic livers show immunostaining for desmin (A) and aSMA (B) (× 400).
Characters of PAF receptor expression in cirrhotic hepatic stellate cells
Scatchard plot analysis of the saturation binding data (Figure 3) revealed an about four-fold increase in 3H-PAF binding capacity in hepatic stellate cells from cirrhotic rats as compared to that of the control (Bmax: 24.65 ± 1.96 fmol/μg. DNA, R = 0.982 vs 5.74 ± 1.55 fmol/μg. DNA, R = 0.93; P < 0.01). There was no significant difference in the PAF receptors affinity between the cells from control and those from cirrhotic rats (Kd of 4.66 ± 0.33 nmol/L for the cirrhosis and 3.51 ± 0.26 nmol/L for the control; P > 0.05). The data showed that the number of PAF receptor in cirrhotic hepatic stellate cells had a dramatic increase of about 3.3 folds compared to that of the control (0.40 ± 0.09 vs 0.12 ± 0.03, P < 0.05) (Figure 3). Consistent with hepatic PAF receptor saturation binding data, the mRNA expression of PAF receptor was also increased in the cirrhotic hepatic stellate cells (Figure 4).
Figure 3.
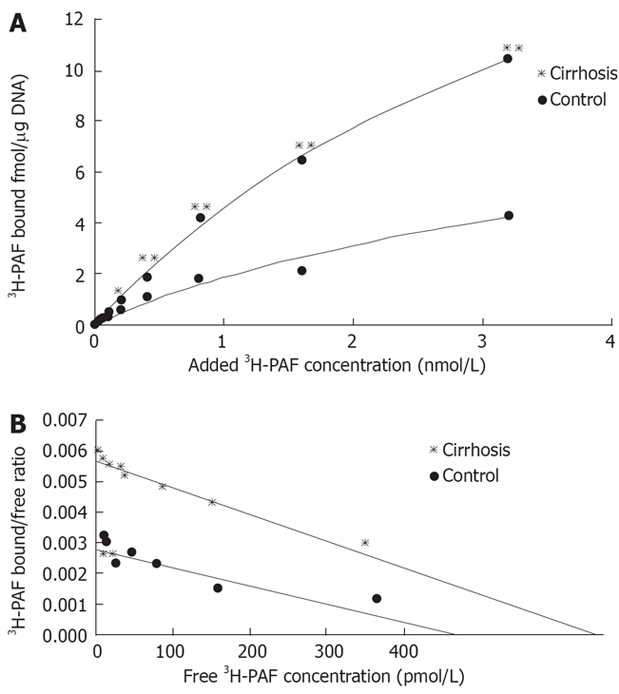
A: Saturation curve of 3H-PAF binding to cultured cirrhotic hepatic stellate cells. 3H-PAF in the concentration between 0.125 and 3.2 nmol/L,in presence or the absence of 5 μmol/L unlabeled incubated at 25°C for 3 h; B: Scatchard plot analysis of binding of 3H-PAF to cirrhotic hepatic stellate cells. Cirrhosis: Kd: 4.66 nmol/L, Bax: 24.65 ± 1.96 fmol/μg DNA, R = 0.982; Control: Kd: 3.51 nmol/L, Bax: 5.74 ± 1.55 fmol/μg DNA, R = 0.93.
Figure 4.
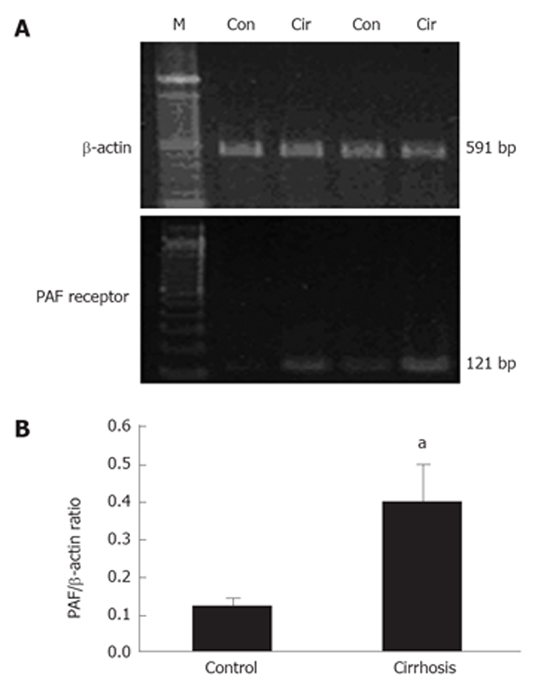
PAF receptor expression in cirrhotic hepatic stellate cells. RT-PCR of PAF receptor mRNA was performed with cDNA prepared from RNA samples of control and cirrhotic HSCs. Expression of β-actin mRNA was assessed using the same amout of cDNA. A: PCR products of PAF and β-actin from control (Con) and cirrhotic (Cir) rat livers are shown; B: Ratio of the PAF receptor and β-actin mRNA. aP < 0.05 vs control.
Effect of PAF and its antagonist BN52021 on synthesis of prostaglandin E2 in cirrhotic hepatic stellate cells
PAF stimulated PGE2 synthesis in a concentration-dependent manner both in control and cirrhotic hepatic stellate cells; the effect being more manifest in the latter. The date revealed an about 1.8-fold increase in hepatic stellate cells from cirrhotic rats compared with that from the control (3.89 ± 0.21 pg/μg. DNA vs 2.08 ± 0.13 pg/μg. DNA, P < 0.01) in 100 Nm (Figure 5).
Figure 5.
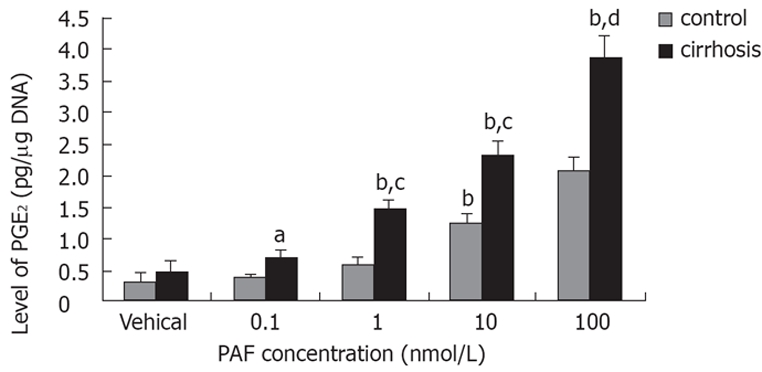
Effect of PAF concentration on PGE2 synthesis in cirrhotic hepatic stellate cells. Cell were placed in serum-free medium after overnight culture, and were challenged with PAF 24 h later. PGE2 in the medium was measured by ELISA after 15 min incubation. aP < 0.05, bP < 0.01 vs vehical; cP < 0.05, dP < 0.01 vs control.
Cells were placed in serum-free medium after overnight culture, and were challenged with PAF(100 nmol/L) ± BN52021 (5 nmol/L) 24 h later. PGE2 synthesis in the medium was decreased with BN52021 both in control (2.08 ± 0.13 pg/μg DNA vs 1.02 ± 0.10 pg/μg. DNA, P < 0.01) and cirrhotic hepatic stellate cells (3.89 ± 0.21 pg/μg. DNA vs 1.28 ± 0.14 pg/μg. DNA, P < 0.01).
DISCUSSION
In the present study, stellate cells from cirrhotic liver had a 3.3-fold increase in PAF receptor density and a 1.8-fold increase in PAF induced PGE2 synthesis relative to control cells, suggesting cirrhosis sensitizes hepatic stellate cells to PAF and HSCs may be potential effectors of PAF induced portal hypertension. One of the major characteristics of liver cirrhosis is proliferation and activation of stellate cells, which are responsible for excessive deposition of fibrous tissue and the contractile component of portal hypertension. After acute or chronic liver injury, HSCs transdifferentiate to a transforming myofibroblastic phenotype, increasing contractility, proliferating, accumulating, and expressing a number of fibrogenic and proliferative cytokines and their cognate receptors[29–31]. Previous work showed PAF cause contraction in HSCs[17], but as there is on reliable assay system for normal stellate cell contractility, we measured PAF dependent PGE2 synthesis in HSCs, which were increased in cirrhotic HSCs including PAF stimulated and basal synthesis. The existence of increased PAF receptor density in cirrhotic HSCs and PAF induced PGE2 synthesis was blocked by its antagonist BN52021 suggest PAF act its biological effects by its specific receptors. It has been shown that PGE2, a hepatoprotective eicosanoid, can inhibit growth factor induced proliferation of activated stellate cells by causing inhibition of MAP kinase activity[32]. So it is reasonable to assume that enhanced synthesis of PGE2 (basal as well as PAF stimulated) in cirrhosis may be a homeostatic mechanism limiting the contractile response.
Previous study showed the increased PAF in the cirrhotic liver due to its enhanced synthesis by the Kupffer cells, with ET-1 playing a major role. Endothelin-1 (ET-1) stimulates PAF synthesis by Kupffer cells[13,33]. Cirrhotic Kupffer cells also had elevated densities of functional receptors for both PAF and ET-1 (exclusively ETB)[12,13,33]. By respective receptor, both ET-1 and PAF were shown to stimulate PGE2 synthesis in Kupffer cells[34,35]. Since both PAF and ET-1 individually cause portal hypertension, an increase in ET-1-stimulated PAF synthesis in Kupffer cells was to exacerbate the hepatic and extrahepatic complications of cirrhosis. As activated stellate cells also synthesize greater amounts of ET-1[36,37], the implication of platelate-activating factor, ET-1, Kupffer Cells and hepatic stellate cells in the pathology of liver cirrhosis is summarized in a schematics (Figure 6).
Figure 6.

Implication of platelate-activating factor, ET-1, Kupffer cells and hepatic stellates cells in the pathology of liver cirrhosis. Increased ET-1 released by stellates and endothelial cells in the cirrhosis liver acts on upregulated ETB receptors in Kupffer cells and stimulates the synthesis of PAF. Likewise, autocrine actions of proinflammatory mediator such as IL-1 and TNF-α may also stimulate PAF synthesis by Kupffer cells. PAF then acts on hepatic vascular smooth muscle cells and stellate cells and contributes to portal hypertension by causing their contraction. Its spillover into systemic circulation is likely a cause of hypertension associated with cirrhosis. Autocrine PAF and paracrine ET-1 can act on their respective upregulated receptors in Kupffer cells and stellate cells to cause the synthesis and release of prostaglandin E2, which in turn may act to limit the liver injury.
In conclusion, the findings of this paper shows upregulated PAF receptors in cirrhotic HSCs, by which PAF induce PGE2 synthesis, suggesting HSCs may be potential effectors of PAF induced portal hypertension. PAF is an integral part of portal hypertension.
COMMENTS
Background
Platelet activating factor (PAF) is a potent hepatic vasoconstrictor and systemic vasodilator. In a parallel study, the increased hepatic and circulating PAF concentrations in cirrhotic rats was observed, which contributes to portal hypertension. The potential effectors of PAF induced increase in portal pressure remains undetermined. Hepatic stellate cells play a crucial role in the development of liver fibrosis. So we hypothesized the HSCs may be potential effectors of PAF induced increases in portal pressure. Therefore, PAF receptors and PAF-induced prostaglandin E2 release derived from cirrhotic hepatic stellate cells were investigated.
Research frontiers
This study demonstrated upregulated PAF receptors in cirrhotic HSCs, suggesting HSCs may be potential effectors of PAF induced portal hypertension.
Innovations and breakthroughs
This study explained one of the possible mechanisms of development of portal hypertension in CCl4-induced cirrhotic rats, showing that hepatic stellate cells may be potential effectors of PAF induced portal hypertension.
Applications
In this study, the implication of Platelate-activating Factor and Hepatic Stellate Cells in the pathology of liver cirrhosis was explored. Upregulated PAF receptors in cirrhotic HSCs were determined. So it is a strategy to block the interaction of PAF and HSCs for treatment of portal hypertension.
Terminology
The existence of PAF receptor in HSCs suggests HSCs may be potential effectors of PAF induced portal hypertension. PAF acts its biological activities by its receptor.
Peer review
This study determined upregulated PAF receptors in cirrhotic HSCs and explained the implication of PAF and HSCs in the pathology of liver cirrhosis. It is a strategy to block the interaction of PAF and HSCs for treatment of portal hypertension. It was thought to be very innovative and deserved to be published.
Supported by The Key Scientific and Technological Research Foundation of the National 863 Program, No. 2003AA208106; Medical Outstandard Foundation of Army, No. 04J020
S- Editor Liu Y L- Editor Alpini GD E- Editor Wang HF
References
- 1.Chao W, Olson MS. Platelet-activating factor: receptors and signal transduction. Biochem J. 1993;292(Pt 3):617–629. doi: 10.1042/bj2920617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Montrucchio G, Alloatti G, Camussi G. Role of platelet-activating factor in cardiovascular pathophysiology. Physiol Rev. 2000;80:1669–1699. doi: 10.1152/physrev.2000.80.4.1669. [DOI] [PubMed] [Google Scholar]
- 3.Prescott SM, Zimmerman GA, Stafforini DM, McIntyre TM. Platelet-activating factor and related lipid mediators. Annu Rev Biochem. 2000;69:419–445. doi: 10.1146/annurev.biochem.69.1.419. [DOI] [PubMed] [Google Scholar]
- 4.Snyder F. Platelet-activating factor and related acetylated lipids as potent biologically active cellular mediators. Am J Physiol. 1990;259:C697–C708. doi: 10.1152/ajpcell.1990.259.5.C697. [DOI] [PubMed] [Google Scholar]
- 5.Buxton DB, Shukla SD, Hanahan DJ, Olson MS. Stimulation of hepatic glycogenolysis by acetylglyceryl ether phosphoryl-choline. J Biol Chem. 1984;259:1468–1471. [PubMed] [Google Scholar]
- 6.Hines KL, Braillon A, Fisher RA. PAF increases hepatic vascular resistance and glycogenolysis in vivo. Am J Physiol. 1991;260:G471–G480. doi: 10.1152/ajpgi.1991.260.3.G471. [DOI] [PubMed] [Google Scholar]
- 7.Kleber G, Braillon A, Gaudin C, Champigneulle B, Cailmail S, Lebrec D. Hemodynamic effects of endotoxin and platelet activating factor in cirrhotic rats. Gastroenterology. 1992;103:282–288. doi: 10.1016/0016-5085(92)91124-m. [DOI] [PubMed] [Google Scholar]
- 8.Yang YP, Ma XM, Wang CP, Han J, Lu YY, Xiang Y, Su SH, Feng YY. Effect of increased hepatic platelet activating factor and its receptor portal hypertension in CCl4-induced liver cirrhosis. World J Gastroenterol. 2006;12:709–715. doi: 10.3748/wjg.v12.i5.709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yang Y, Nemoto EM, Harvey SA, Subbotin VM, Gandhi CR. Increased hepatic platelet activating factor (PAF) and PAF receptors in carbon tetrachloride induced liver cirrhosis. Gut. 2004;53:877–883. doi: 10.1136/gut.2003.024893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ma XM, Wang CP, Han J, Xiang Y, Su SH, Feng YY, Yang YP. Increased production of platelet activating factor (PAF) and PAF receptors in CL4-induced cirrhosis: its implications in chronic liver injury. Med J Chin PLA. 2004;29:1062–1064. [Google Scholar]
- 11.Wang CP, Han J, Ma XM, Dong K, Xiang Y, Su SH, Feng YY, Yang YP. Influence of platelet activating factor and its antagonist on portal hypertension associated with liver cirrhosis: an experiment with rats. Zhonghua Yixue Zazhi. 2005;85:3337–3341. [PubMed] [Google Scholar]
- 12.Wang CP, Su SH, Lu YY, Zhou L, Chen Y, Feng YY, Yang YP. Snthesis of platelet activating factor(PAF) and expression of PAF receptor in Kupffer cells in portal hypertension in rat. Med J Chin PLA. 2006;31:784–786. [Google Scholar]
- 13.Yang Y, Harvey SA, Gandhi CR. Kupffer cells are a major source of increased platelet activating factor in the CCl4-induced cirrhotic rat liver. J Hepatol. 2003;39:200–207. doi: 10.1016/s0168-8278(03)00229-0. [DOI] [PubMed] [Google Scholar]
- 14.Halonen M, Dunn AM, Palmer JD, McManus LM. Anatomic basis for species differences in peripheral lung strip contraction to PAF. Am J Physiol. 1990;259:L81–L86. doi: 10.1152/ajplung.1990.259.2.L81. [DOI] [PubMed] [Google Scholar]
- 15.Friedman SL. Seminars in medicine of the Beth Israel Hospital, Boston. The cellular basis of hepatic fibrosis. Mechanisms and treatment strategies. N Engl J Med. 1993;328:1828–1835. doi: 10.1056/NEJM199306243282508. [DOI] [PubMed] [Google Scholar]
- 16.Rockey D. The cellular pathogenesis of portal hypertension: stellate cell contractility, endothelin, and nitric oxide. Hepatology. 1997;25:2–5. doi: 10.1053/jhep.1997.v25.ajhep0250002. [DOI] [PubMed] [Google Scholar]
- 17.Pinzani M, Gentilini P. Biology of hepatic stellate cells and their possible relevance in the pathogenesis of portal hypertension in cirrhosis. Semin Liver Dis. 1999;19:397–410. doi: 10.1055/s-2007-1007128. [DOI] [PubMed] [Google Scholar]
- 18.Gandhi CR, Nemoto EM, Watkins SC, Subbotin VM. An endothelin receptor antagonist TAK-044 ameliorates carbon tetrachloride-induced acute liver injury and portal hypertension in rats. Liver. 1998;18:39–48. doi: 10.1111/j.1600-0676.1998.tb00125.x. [DOI] [PubMed] [Google Scholar]
- 19.Gandhi CR, Sproat LA, Subbotin VM. Increased hepatic endothelin-1 levels and endothelin receptor density in cirrhotic rats. Life Sci. 1996;58:55–62. doi: 10.1016/0024-3205(95)02255-4. [DOI] [PubMed] [Google Scholar]
- 20.Luckey SW, Petersen DR. Activation of Kupffer cells during the course of carbon tetrachloride-induced liver injury and fibrosis in rats. Exp Mol Pathol. 2001;71:226–240. doi: 10.1006/exmp.2001.2399. [DOI] [PubMed] [Google Scholar]
- 21.Shinozaki K, Kawasaki T, Kambayashi J, Sakon M, Shiba E, Uemura Y, Ou M, Iwamoto N, Mori T. A new method of purification and sensitive bioassay of platelet-activating factor (PAF) in human whole blood. Life Sci. 1994;54:429–437. doi: 10.1016/0024-3205(94)00701-2. [DOI] [PubMed] [Google Scholar]
- 22.Bito H, Honda Z, Nakamura M, Shimizu T. Cloning, expression and tissue distribution of rat platelet-activating-factor-receptor cDNA. Eur J Biochem. 1994;221:211–218. doi: 10.1111/j.1432-1033.1994.tb18731.x. [DOI] [PubMed] [Google Scholar]
- 23.Nudel U, Zakut R, Shani M, Neuman S, Levy Z, Yaffe D. The nucleotide sequence of the rat cytoplasmic beta-actin gene. Nucleic Acids Res. 1983;11:1759–1771. doi: 10.1093/nar/11.6.1759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gandhi CR, Kuddus RH, Uemura T, Rao AS. Endothelin stimulates transforming growth factor-beta1 and collagen synthesis in stellate cells from control but not cirrhotic rat liver. Eur J Pharmacol. 2000;406:311–318. doi: 10.1016/s0014-2999(00)00683-x. [DOI] [PubMed] [Google Scholar]
- 25.Uemura T, Gandhi CR. Inhibition of DNA synthesis in cultured hepatocytes by endotoxin-conditioned medium of activated stellate cells is transforming growth factor-beta and nitric oxide-independent. Br J Pharmacol. 2001;133:1125–1133. doi: 10.1038/sj.bjp.0704151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bito H, Honda Z, Nakamura M, Shimizu T. Cloning, expression and tissue distribution of rat platelet-activating-factor-receptor cDNA. Eur J Biochem. 1994;221:211–218. doi: 10.1111/j.1432-1033.1994.tb18731.x. [DOI] [PubMed] [Google Scholar]
- 27.Gomez J, Bloom JW, Yamamura HI, Halonen M. Characterization of receptors for platelet-activating factor in guinea pig lung membranes. Am J Respir Cell Mol Biol. 1990;3:259–264. doi: 10.1165/ajrcmb/3.3.259. [DOI] [PubMed] [Google Scholar]
- 28.Ibe BO, Sander FC, Raj JU. Platelet-activating factor receptors in lamb lungs are downregulated immediately after birth. Am J Physiol Heart Circ Physiol. 2000;278:H1168–H1176. doi: 10.1152/ajpheart.2000.278.4.H1168. [DOI] [PubMed] [Google Scholar]
- 29.Pinzani M, Milani S, De Franco R, Grappone C, Caligiuri A, Gentilini A, Tosti-Guerra C, Maggi M, Failli P, Ruocco C, et al. Endothelin 1 is overexpressed in human cirrhotic liver and exerts multiple effects on activated hepatic stellate cells. Gastroenterology. 1996;110:534–548. doi: 10.1053/gast.1996.v110.pm8566602. [DOI] [PubMed] [Google Scholar]
- 30.Hines KL, Bataller R, Nicolás JM, Gineès P, Görbig MN, Garcia-Ramallo E, Lario S, Tobías E, Pinzani M, Thomas AP, et al. Contraction of human hepatic stellate cells activated in culture: a role for voltage-operated calcium channels. J Hepatol. 1998;29:398–408. doi: 10.1016/s0168-8278(98)80057-3. [DOI] [PubMed] [Google Scholar]
- 31.Bataller R, Sancho-Bru P, Ginès P, Lora JM, Al-Garawi A, Solé M, Colmenero J, Nicolás JM, Jiménez W, Weich N, et al. Activated human hepatic stellate cells express the renin–angiotensin system and synthesize angiotensin II. Gastroenterology. 2003;125:117–125. doi: 10.1016/s0016-5085(03)00695-4. [DOI] [PubMed] [Google Scholar]
- 32.Bataller R, Ginès P, Nicolás JM, Görbig MN, Garcia-Ramallo E, Gasull X, Bosch J, Arroyo V, Rodés J. Angiotensin II induces contraction and proliferation of human hepatic stellate cells. Gastroenterology. 2000;118:1149–1156. doi: 10.1016/s0016-5085(00)70368-4. [DOI] [PubMed] [Google Scholar]
- 33.Su SH, Wang CP, Lu YY, Zhou L, Chen Y, Feng YY, Yang YP. Effects of endothelin on platelet activating factor(PAF) synthesis and release by cultured Kupffer cells from CL4-induced cirrhosis rats. Med J Chin PLA Med. 2007;32:201–204. [Google Scholar]
- 34.Gandhi CR, Hanahan DJ, Olson MS. Two distinct pathways of platelet-activating factor-induced hydrolysis of phosphoinositides in primary cultures of rat Kupffer cells. J Biol Chem. 1990;265:18234–18241. [PubMed] [Google Scholar]
- 35.Gandhi CR, Stephenson K, Olson MS. A comparative study of endothelin- and platelet-activating-factor-mediated signal transduction and prostaglandin synthesis in rat Kupffer cells. Biochem J. 1992;281(Pt 2):485–492. doi: 10.1042/bj2810485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gandhi CR, Sproat LA, Subbotin VM. Increased hepatic endothelin-1 levels and endothelin receptor density in cirrhotic rats. Life Sci. 1996;58:55–62. doi: 10.1016/0024-3205(95)02255-4. [DOI] [PubMed] [Google Scholar]
- 37.Gandhi CR, Kuddus RH, Uemura T, Rao AS. Endothelin stimulates transforming growth factor-beta1 and collagen synthesis in stellate cells from control but not cirrhotic rat liver. Eur J Pharmacol. 2000;406:311–318. doi: 10.1016/s0014-2999(00)00683-x. [DOI] [PubMed] [Google Scholar]