

Abstract
The observation that mast cells accumulate at the periphery of growing tumours is now well documented, and the loss of mast cells correlates with reduced tumour growth. The role of mast cells as innate regulators of both inflammatory and immunosuppressive responses slowly becomes clear as novel tools become available. This review will address the role of mast cells in tumours and how they can interact with the local immune environment to mediate immune suppression contributing to tumour escape.
Keywords: acquired immunity, immunosuppression, mast cells, tumour
Introduction
Mästzellen, or mast cells (MCs), discovered now more than a century ago by Paul Ehrlich, were first described in the thesis of the then 24-year-old medical student. Ehrlich describes the typical characteristic of these cells as filled with numerous granules of varying size and notes that MCs could be found around developing tissues, initiating the idea of a close relationship between MCs and tumour growth [1].
Historically, MCs are described as the main responders to allergens and as a source of protection against parasites. However, recent literature has provided an additional layer of complexity to this simplified role, and MCs are now recognized as having a spectra of both proinflammatory and tolerogenic functions [2,3].
Mast cells are associated with a number of pathologies and reactions including type I hypersensitivity reaction, chronic inflammatory processes, tissue remodelling, wound healing and angiogenesis [4–6]. Pertinent to this discussion, the latter three are regarded as essential for the establishment and progression of neoplastic growth.
The role of MCs in the generation of adaptive immune responses is a double-edged sword. While MCs can induce overt inflammation, as seen in allergies, autoimmunity and contact hypersensitivity (CHS) [7–10], they can also exert immune suppression as seen in allograft transplantation models, chronic UV-B irradiation and mosquito bites [11–14]. The role of MCs in the regulation of acquired immunity is far from being resolved, and the seemingly conflicting character of these cells requires further mechanistic analysis. In this review we will discuss the advances made on the immunosuppressive role of MCs during the development and progression of tumours.
Distribution
We now consider MCs as the critical mediators of innate immunity, as they can be found most abundantly in areas highly exposed to the external environment, such as the skin, respiratory track, lung tissue, gastrointestinal tract and the urinary system. Anatomically, MCs are found throughout the human host and are located usually at the periphery of blood and lymphatic vessels in the connective tissue; they can also be found in sites directly adjacent to blood vessels and peripheral nerves as well as in the secondary lymphoid organs and in the central nervous system. The currently accepted MC ontogeny describes the origin and development of these cells as arising from undifferentiated and multi-potential haemopoietic progenitors resident within the bone marrow as stem cells expressing CD34 and c-kit (CD117). These progenitors receive signals through their adhesion molecules and receptors and enter the circulation where they migrate and undergo chemotaxis into the tissue. Once seeded in tissue these immature progenitor cells differentiate, perhaps terminally, to functional maturity through cues from the local microenvironment [15].
Subsets
At least two subpopulations of murine MCs have been identified based on the composition of chymases and tryptases within their granules. Mucosal MCs reside in the respiratory tract and the gastrointestinal tract mucosa. Connective tissue MCs are found in skin, lung, connective tissue and the submucosa of the gastrointestinal tract. It is likely that additional subsets will be identified as further research compartmentalizes MCs by function, organ of residence, phenotype and granule composition. Notably, we have shown that MCs can mediate immunosuppression in tolerant allografts, and it is possible that the phenotypic profile of these ‘tolerizing’ MCs would be distinct from those identified previously. We would also submit that the MCs infiltrating tumours may as well harbour a distinctive profile from either the mucosal or connective tissue MCs. Transcriptional profiling of purified MCs from tolerant tissue and from tumour sites will provide us with our first insights of these immunosuppressive MC subsets.
Mast cell mediators and effector functions
To understand more clearly the versatility of MCs and the ability of these cells to partake in pathologies, ranging from type I hypersensitivity reactions to graft tolerance, it is important to consider basic MC biology. MCs produce three categories of effector molecules: those which are stored in granules such as serotonin, histamine, heparin, tryptase and chymase; those which are synthesized de novo upon cell stimulation such as lipid mediators (PAF), prostaglandins (PDG2) and leucotrienes (LTB4, LTD4); and a large variety of cytokines such as interleukin (IL)-1, IL-3, IL-5, IL-8, IL-10, granulocyte–macrophage-colony-stimulating factor, tumour necrosis factor (TNF)-α, transforming growth factor (TGF)-β and vascular endothelial growth factor (VEGF) [2].
Mast cell degranulation and ‘piecemeal degranulation’ are two of probably a number of ways in which MCs can influence immunity through the release of the above-listed mediators. Unique and central to the development of immediate hypersensitivity is the cross-linking of FcεRI, the high-affinity IgE receptor, by allergen-specific immunoglobulin (Ig)E on MCs. Upon activation of MCs there is rapid release of the content of their granules by exocytosis, enzymatic synthesis of lipid mediators derived from precursors stored in cell membranes, and in some cases in the lipid bodies, and initiation of transcription, translation and secretion of cytokines [16]. Less is known on the mechanisms of how piecemeal degranulation occurs. Instead of utilizing high-affinity FcεRI, IgE-independent activation takes advantage of such receptors on MCs as low-affinity IgG receptors FcγRII and FcγRIII, complement receptors (C3aR, C5aR, CR2, CR4) and pattern recognition receptors. There is a growing literature that has documented the differential release of MC mediators upon selective stimulation of some of these receptors [17]. The first report on this process demonstrated that serotonin but not histamine can be secreted by MCs in response to an exogenous stimuli such as amitriptyline [18]. Another example shows the differential release of IL-6 without histamine in response to bacterial lipopolysaccharide [19]. Thus, in considering the vast roles that MCs may be playing in regulating acquired immunity, their ability to selectively release mediators capable of modulating immunity in a manner tailored to the stimulus may be critical to acquired immunity. In our studies on the role of MCs in the maintenance of tolerant allografts, it is clear that the infiltrating MCs still contained granules. As such, we have concluded that the maintenance of tolerance by MCs is probably a secretory process by which MCs release immunosuppressive mediators such as TGF-β or IL-10. Definitive proof of the MC mediators of suppression is being sought currently.
Models of MC deficiency
The role of MCs in the development and progression of human immunity and disease is implicated by their presence or the presence of their products in defined clinical conditions. In contrast, the use of mice in which MCs are selectively or specifically absent has allowed us to definitively implicate MCs in defined immunological and pathological situations.
There are currently two widely used models of MC deficiency: the (WBXC57BL/6)F1-KitW/KitWv (the W/WV) and the KitWsh/KitWsh (Wsh) mice. Both are left severely MC-deficient by a mutation in the c-kit receptor. The two strains are otherwise different in the kinds of additional abnormalities that they carry, with the Wsh mice being least affected [20,21]. Confirming MCs as the sole mediators of the phenotypes observed in these strains is performed by MC ‘knock-in’ experiments where MC-deficient mice are reconstituted with MCs derived in vitro from bone marrow cultures (BMMC), as reviewed elsewhere by Brown et al. 2008 [2]. The current gold-standard of studies in MC deficiency, however, may be advancing, as models utilizing transgenic mice harbouring MC-specific expression of Cre recombinase are being generated and tested. The newly developed Cre recombinase-expressing mouse line that allows MC-specific inactivation of genes in vivo expresses Cre under the control of the mast cell protease (Mcpt) 5 promoter [22]. When Mcpt5-Cre transgenic mice are crossed to the ROSA26-EYFP mouse efficient Cre-mediated recombination is observed in MCs from the peritoneal cavity and the skin, as indicated by YFP expression exclusively by MCs. Novel MC-deficient mouse systems are now being achieved by crossing the Mcpt5-Cre transgenic mice with the inducible DTR transgenic mice (iDTR), or with the mice in which the anti-apoptotic protein MCL-1 can be excised by Cre-mediated deletion [23,24]. The former generates a MC-deficient animal upon administration of diphtheria toxin and the later profoundly reduces MC numbers in a manner similar to that seen in the case of B and T lymphocytes ([23] and unpublished observation). An additional in vivo tool in which MC-specific genes may be ablated or in which MCs can be deleted was generated recently by constructing a mouse in which the expression of Cre recombinase is under the control of the baboon α-chymase promoter (Chm : Cre). In this mouse, Cre expression occurs specifically in mouse MCs: however, this model appears to have a more restricted MC Cre expression pattern, as only mature tissue resident MCs were observed to have undergone Cre-mediated recombination when these mice were crossed with the ROSA26R reporter mice [25]. These newly developing models will be invaluable in helping us to understand more clearly the multiple ways by which MCs exert their effects on their environment.
Mast cells and tumour
The accumulation of MCs at the periphery of tumours was first reported by Westphal in 1891 [26]. Since that time, many studies in both experimental models in rodents and in a diverse array of tumours in humans have confirmed this important observation. Interestingly, enhanced MC accumulation in tumours is associated with a poor prognosis, indicating a biological role for MCs in tumour progression [27,28].
Some of the most aggressive human tumours, breast carcinoma, malignant melanoma and colorectal adenocarcinoma are noted as tumours that are particularly populated with MCs at their periphery [29–31]. As such, it has been proposed that the infiltration of MCs at the tumour site can greatly enhance tumorigenesis [32].
Figure 1 depicts the presence of MCs in the skin and periphery of a tumour model of murine bladder carcinoma. It is worth noting the distribution of MCs mainly at the junction of skin and tumour tissue (Fig. 1).
Fig. 1.
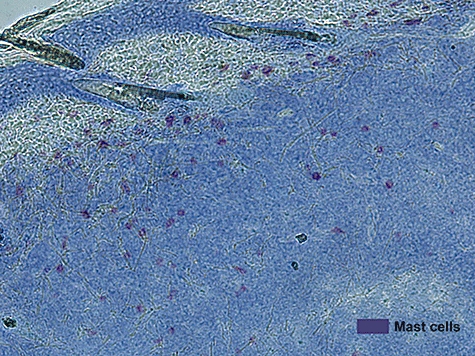
Mast cells accumulate at the periphery of murine bladder carcinoma. Toludine blue staining of tumour, and skin surrounding tumour, show mast cells in purple against the blue tissue.
It is believed that MCs may impact upon the growth of tumours by multiple mechanisms, including angiogenesis. Multiple recent studies have demonstrated that early angiogenic activity is dependent upon MCs and is an essential part of neoplastic development, with MCs mediating this activity by releasing heparin, VEGF and IL-8 [33–36].
So far, reports addressing MCs specifically as actors in tumour immunosuppression come from Feng et al., where the authors mobilize MCs to infiltrate tumour by means of stem cell factor (SCF). It is observed that tumour-infiltrating MCs express multiple proinflammatory factors and increase IL-17 expression in tumour. These SCF-activated MCs also exacerbate tumour immunosuppression by releasing adenosine and increasing the number of T regulatory cells (Tregs) present in the tumour microenvironment, thereby augmenting the suppression of T cells and natural killer cells in the tumour. This is perhaps one of the first reports to implicate MCs specifically both as regulators of inflammation as well as of immunosuppression in the tumour microenvironment [37].
Other mechanisms by which MCs may impact the growth and metastasis of tumours include enhancing tissue remodelling through the degradation of the extracellular matrix, by releasing growth factors essential for tumour growth and by suppressing the hosts’ acquired immune response against the tumour.
Mast cells as mediators of tumour immunosuppression
It is appreciated that tumours dynamically create immune tolerance against themselves by creating sites of immune privilege and by inducing anti-tumour T cell tolerance [38]. The induction of immunological ignorance, induction of anergy in effector T cells and active suppression of effector cells have all been reported to contribute to the lack of immune responses to tumours [39–43]. Moreover, cross-presentation of tumour antigens by tolerogenic host antigen-presenting cells (APCs) is thought currently to probably be a major mechanism by which effector T cells are rendered anergic to a developing tumour. In addition, suppression is thought to be created locally within the tumour itself by a number of suppressive mechanisms, including the induction and recruitment of Treg cells. Tolerogenic APCs and Tregs are prevailing as the major players in immunosuppression and are seen as important contributors to the tolerogenic tumour microenvironment [38]. The question that emerges is whether and how MCs contribute to the development of immune privilege within the tumour microenvironment.
Mast cells influence dendritic cell migration and functionality
As mentioned, critical to the initiation of tumour-specific immune responses is the presentation of tumour antigens by host APCs to cognate T cells at the tumour site and within the regional draining lymph nodes (dLN). In this context, MCs exert a profound role in regulating the early functions of dendritic cells (DCs) whose activities are essential in generating an anti-tumour immune response. As such, MCs inevitably become important players in the initiation of acquired immunity.
Mast cells and DCs are found in similar locations in the periphery of the host and are considered to be one of the first lines of immunological defence. Both these cell types mediate the communication of the innate arm of the immune system with the adaptive. DCs, like MCs, are found preferentially in the peripheral areas of the tumour [44]. As such, it is plausible that MC-induced migration of DCs is one way in which MCs exert control over the initial DC priming of tumour-specific T cells. Over recent years there is emerging evidence of the impact of MCs on migration and functionality of DCs. Among the numerous mediators produced by MCs able to impact DCs are prostaglandin E2 (PGE2) and prostaglandin D2 (PGD2). These factors lead to DC maturation and concomitant production of the proinflammatory cytokines, IL-12 and IL-6 [45–47]. Furthermore, it has been shown that PGE2 induces metalloproteinase-9 and increases the expression of the CCR7 ligands CCL17/21 on the surface of DCs, promoting DCs migration from the skin to the dLN under inflammatory conditions [45,48]. PGD2, on the other hand, has the opposite effect, and inhibits migration of DCs as seen in both skin and lung [47,49]. Both PGE2 and PGD2 matured DCs exhibit an enhanced capacity to activate T cells. Another MC-derived mediator for DC and Langerhans cell (LC) migration is TNF-α[10,50]. TNF-α levels are seen to be elevated in ultraviolet-exposed skin, and is thought to be the main contributor to altered LC traffic and function. Studies involving intracutaneous injections of TNF-α have shown to elicit LC damage similar to that observed upon ultraviolet irradiation. Additionally, antibodies to TNF-α protect LCs from the morphological changes seen because of ultraviolet radiation [51,52]. MCs accumulate and upon activation release large amounts of TNF-α. As both MCs and LCs reside proximally in the skin, MCs are a significant source of TNF-α, indicating that in a tumour setting, MCs may also drive TNF-α-mediated DC migration [53]. Because DCs from the tumour-dLN present antigen in a tolerizing manner, MC-derived TNF-α could be contributing in maintaining the immunosuppressive environment by increasing antigen presentation in the dLN in a immunosuppressive way [38].
A third major effector produced by MCs and known to impact DCs is histamine. DCs express all four known histamine receptors, H1R, H2R, H3R and H4R [54,55]. H4R on DCs has been shown to improve antigen uptake and cross-presentation [56]. H4R stimulation of DCs leads to the suppression of IL-12 production but does not alter IL-10 secretion, thereby biasing the DCs to contain an immunosuppressive character [57]. This could potentially be an additional mechanism by which MCs drive intratumoral-DCs to present antigen in a tolerogenic fashion. These effects, combined with the fact that MCs can recruit DCs and LCs actively to the site of inflammation by the release of CCL1, could contribute significantly to the establishment and maintenance of an immunosuppressive environment [58].
Mast cells directly modulate T cell responses
In addition to the regulation of DC functionality and migration, MC products can also impact directly on T cell activation. Not only can the mediators secreted by MCs modulate the response of the T cell directly, but they also contribute to enhanced migration and better accessibility by changing the permeability of the vasculature [59]. Given the breadth of mediators potentially produced by MCs, it is impossible to predict how MCs will skew a helper T cell response without careful consideration of how the MC is activated and the microenvironmental context at hand. It is quite possible that within the tumour microenvironment, MC products can mediate the development of T cell tolerance through the release of soluble mediators.
Interleukin-10 is known commonly as one of the immunomodulatory cytokines produced by a subset of Tregs[60]. However, MCs are also able to produce IL-10 and play an important role in the suppression of secondary responses to a hapten in combination with ultraviolet B (UV-B) [12]. Studies show that in both the CHS model, as during chronic UV-B irradiation, Fc receptor-mediated activation of MCs leads to production of IL-10. These observations stem from studies showing that MC-deficient mice exhibit stronger CHS responses to urushiol, the allergen-bearing sap derived from poison oak and poison ivy, than their wild-type counterparts. Early (day 5) and late (day 10) post-challenge MC-deficient mice and those reconstituted with IL-10−/− BMMCs showed increased epidermal ulceration and necrosis and dermal leucocytic infiltration compared with their control littermates, indicating that MC production of IL-10 is required for limiting inflammation and decreasing the swelling associated with urushiol. Dermal MCs and MC-produced IL-10 play a similar suppressive role in reducing inflammation in response to chronic UV-B irradiation [13]. It has been shown that MC-derived IL-10 also dampens immune responses after mosquito bites [14]. The production of IL-10 by MCs at the tumour site could therefore contribute towards generating an immunosuppressive microenvironment favorable for tumour growth.
Recently, serotonin, thought of generally as a neurotransmitter, has been recognized as an important player in immunological responses. It has been shown that serotonin can promote T cell proliferation and is considered an accessory signal for T cell activation [61–63]. MCs express the serotonin-specific transporter, enabling them to take up and store serotonin. During inflammation, the stored serotonin is released during either complement binding or FcεRI cross-linking. The presence of MCs around the tumour could, therefore, lead to depletion of serotonin in the tumour microenvironment helping to create an environment unfavourable for T cell activation and proliferation.
In addition to the secretion of immunosuppressive mediators such as IL-10 or the removal of proinflammatory mediators such as serotonin, MCs might also interact directly with T cells via the expression of surface receptors. In addition to having modest antigen-presenting capacities and the ability to express co-stimulatory molecules, MCs express PD-L1 and PD-L2, the ligands for the inhibitory receptor PD1 expressed by T cells [64]. It has been found that tumours and tumour-infiltrating DCs express high levels of these inhibitory ligands, as such contributing to immune system evasion by the tumour [65]. Signalling via PD-1 can lead to a variety of responses in T cells, of which apoptosis, maintenance of anergy and exhaustion and IL-10 production are the most relevant for tumour escape [66].
As mentioned earlier, the release of histamine by MCs results in a skewed cytokine balance towards that of the immunosuppressive cytokine IL-10. Moreover, histamine in tumour tissue has been shown to suppress local tumour immunity, where results suggest an increase in Treg function by binding of histamine to the H2 receptors. Treg, in turn, release higher levels of IL-10 and may induce apoptosis of APCs. Treatment of mice with cimetidine, an H2 receptor antagonist, has been reported to slow the growth of melanoma [67,68].
Whether MCs are actually able to functionally suppress T cell responses via any one of these pathways in a tumour setting, however, still remains unresolved.
Conclusion
Accumulating evidence from a variety of murine models and from correlative studies in humans suggest strongly that MCs can mediate peripheral T cell tolerance. As such, the accumulation of MCs in tumours represent a barrier to the emergence of protective immunity to cancer. Our laboratory showed clearly the necessity of MCs during the initial phase of tolerance induction in a skin graft model using the Wsh MC-deficient mouse model. Our results were confirmed in rats using a heterotopic allogeneic heart transplant model [69]. It was shown that MCs are essential for the initiation phase of tolerance; however, it should be noted that for the maintenance phase MC accumulation has been detrimental, as reported by several groups [69–71]. If translated directly into oncology models, one might suggest that MCs will interfere with the development of protective anti-tumour immunity and, coupled with their substantive contribution to tumour-induced angiogenesis and tissue remodelling, MCs represent an attractive target for elimination in tumour immunotherapy regimens.
References
- 1.Krishnaswamy G. Mast cells: methods and protocols. New York: Humana Press; 2005. [Google Scholar]
- 2.Sayed BA, Christy A, Quirion MR, Brown MA. The master switch: the role of mast cells in autoimmunity and tolerance. Annu Rev Immunol. 2008;26:705–39. doi: 10.1146/annurev.immunol.26.021607.090320. [DOI] [PubMed] [Google Scholar]
- 3.Christy AL, Brown MA. The multitasking mast cell: positive and negative roles in the progression of autoimmunity. J Immunol. 2007;179:2673–9. doi: 10.4049/jimmunol.179.5.2673. [DOI] [PubMed] [Google Scholar]
- 4.Metz M, Grimbaldeston MA, Nakae S, et al. Mast cells in the promotion and limitation of chronic inflammation. Immunol Rev. 2007;217:304–28. doi: 10.1111/j.1600-065X.2007.00520.x. [DOI] [PubMed] [Google Scholar]
- 5.Murdoch C, Muthana M, Coffelt SB, et al. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer. 2008;8:618–31. doi: 10.1038/nrc2444. [DOI] [PubMed] [Google Scholar]
- 6.Weller K, Foitzik K, Paus R, et al. Mast cells are required for normal healing of skin wounds in mice. FASEB J. 2006;20:2366–8. doi: 10.1096/fj.06-5837fje. [DOI] [PubMed] [Google Scholar]
- 7.Busse W. Mechanisms and advances in allergic diseases. J Allergy Clin Immunol. 2000;105:593–8. doi: 10.1067/mai.2000.106149. [DOI] [PubMed] [Google Scholar]
- 8.Nelson KC, Zhao M, Schroeder PR, et al. Role of different pathways of the complement cascade in experimental bullous pemphigoid. J Clin Invest. 2006;116:2892–900. doi: 10.1172/JCI17891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nigrovic PA, Lee DM. Synovial mast cells: role in acute and chronic arthritis. Immunol Rev. 2007;217:19–37. doi: 10.1111/j.1600-065X.2007.00506.x. [DOI] [PubMed] [Google Scholar]
- 10.Suto H, Nakae S, Kakurai M, et al. Mast cell-associated TNF promotes dendritic cell migration. J Immunol. 2006;176:4102–12. doi: 10.4049/jimmunol.176.7.4102. [DOI] [PubMed] [Google Scholar]
- 11.Lu LF, Lind EF, Gondek DC, et al. Mast cells are essential intermediaries in regulatory T-cell tolerance. Nature. 2006;442:997–1002. doi: 10.1038/nature05010. [DOI] [PubMed] [Google Scholar]
- 12.Hart PH, Grimbaldeston MA, Swift GJ, et al. Dermal mast cells determine susceptibility to ultraviolet B-induced systemic suppression of contact hypersensitivity responses in mice. J Exp Med. 1998;187:2045–53. doi: 10.1084/jem.187.12.2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grimbaldeston MA, Nakae S, Kalesnikoff J, et al. Mast cell-derived interleukin 10 limits skin pathology in contact dermatitis and chronic irradiation with ultraviolet B. Nat Immunol. 2007;8:1095–104. doi: 10.1038/ni1503. [DOI] [PubMed] [Google Scholar]
- 14.Depinay N, Hacini F, Beghdadi W, et al. Mast cell-dependent down-regulation of antigen-specific immune responses by mosquito bites. J Immunol. 2006;176:4141–6. doi: 10.4049/jimmunol.176.7.4141. [DOI] [PubMed] [Google Scholar]
- 15.Gurish MF, Boyce JA. Mast cells: ontogeny, homing, and recruitment of a unique innate effector cell. J Allergy Clin Immunol. 2006;117:1285–91. doi: 10.1016/j.jaci.2006.04.017. [DOI] [PubMed] [Google Scholar]
- 16.Abbas AK, Lichtman AH. Cellular and molecular immunology. 5th edn. Philadelphia, PA: Saunders; 2003. [Google Scholar]
- 17.Theoharides TC, Kempuraj D, Tagen M, et al. Differential release of mast cell mediators and the pathogenesis of inflammation. Immunol Rev. 2007;217:65–78. doi: 10.1111/j.1600-065X.2007.00519.x. [DOI] [PubMed] [Google Scholar]
- 18.Theoharides TC, Bondy PK, Tsakalos ND, et al. Differential release of serotonin and histamine from mast cells. Nature. 1982;297:229–31. doi: 10.1038/297229a0. [DOI] [PubMed] [Google Scholar]
- 19.Leal-Berumen I, Conlon P, Marshall JS. IL-6 production by rat peritoneal mast cells is not necessarily preceded by histamine release and can be induced by bacterial lipopolysaccharide. J Immunol. 1994;152:5468–76. [PubMed] [Google Scholar]
- 20.Grimbaldeston MA, Chen CC, Piliponsky AM, et al. Mast cell-deficient W-sash c-kit mutant KitW-sh/W-sh mice as a model for investigating mast cell biology in vivo. Am J Pathol. 2005;167:835–48. doi: 10.1016/S0002-9440(10)62055-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kitamura Y, Go S, Hatanaka K. Decrease of mast cells in W/Wv mice and their increase by bone marrow transplantation. Blood. 1978;52:447–52. [PubMed] [Google Scholar]
- 22.Scholten J, Hartmann K, Gerbaulet A, et al. Mast cell-specific Cre/loxP-mediated recombination in vivo. Transgenic Res. 2008;17:307–15. doi: 10.1007/s11248-007-9153-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Opferman JT, Letai A, Beard C, et al. Development and maintenance of B and T lymphocytes requires antiapoptotic MCL-1. Nature. 2003;426:671–6. doi: 10.1038/nature02067. [DOI] [PubMed] [Google Scholar]
- 24.Buch T, Heppner FL, Tertilt C, et al. A Cre-inducible diphtheria toxin receptor mediates cell lineage ablation after toxin administration. Nat Methods. 2005;2:419–26. doi: 10.1038/nmeth762. [DOI] [PubMed] [Google Scholar]
- 25.Musch W, Wege AK, Mannel DN, et al. Generation and characterization of alpha-chymase-Cre transgenic mice. Genesis. 2008;46:163–6. doi: 10.1002/dvg.20378. [DOI] [PubMed] [Google Scholar]
- 26.Westphal E. Uber mastzellen. Berlin: Hirschwald Press; 1891. [Google Scholar]
- 27.Imada A, Shijubo N, Kojima H, et al. Mast cells correlate with angiogenesis and poor outcome in stage I lung adenocarcinoma. Eur Respir J. 2000;15:1087–93. doi: 10.1034/j.1399-3003.2000.01517.x. [DOI] [PubMed] [Google Scholar]
- 28.Takanami I, Takeuchi K, Naruke M. Mast cell density is associated with angiogenesis and poor prognosis in pulmonary adenocarcinoma. Cancer. 2000;88:2686–92. [PubMed] [Google Scholar]
- 29.Ch'ng S, Wallis RA, Yuan L, et al. Mast cells and cutaneous malignancies. Mod Pathol. 2006;19:149–59. doi: 10.1038/modpathol.3800474. [DOI] [PubMed] [Google Scholar]
- 30.Amini RM, Aaltonen K, Nevanlinna H, et al. Mast cells and eosinophils in invasive breast carcinoma. BMC Cancer. 2007;7:165. doi: 10.1186/1471-2407-7-165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lachter J, Stein M, Lichtig C, et al. Mast cells in colorectal neoplasias and premalignant disorders. Dis Colon Rectum. 1995;38:290–3. doi: 10.1007/BF02055605. [DOI] [PubMed] [Google Scholar]
- 32.Theoharides TC, Conti P. Mast cells: the JEKYLL and HYDE of tumor growth. Trends Immunol. 2004;25:235–41. doi: 10.1016/j.it.2004.02.013. [DOI] [PubMed] [Google Scholar]
- 33.Coussens LM, Raymond WW, Bergers G, et al. Inflammatory cells up-regualate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 1999;13:1382–97. doi: 10.1101/gad.13.11.1382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gounaris E, Erdman S, Restaino C, et al. Mast cells are an essential hematopoietic component for polyp development. Proc Natl Acad Sci USA. 2007;104:19977–82. doi: 10.1073/pnas.0704620104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nakayama T, Yao L, Tosato G. Mast cell-derived angiopoietin-1 plays a critical role in the growth of plasma cell tumors. J Clin Invest. 2004;114:1317–25. doi: 10.1172/JCI22089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Starkey JR, Crowle PK, Taubenberger S. Mast-cell-deficient W/Wv mice exhibit a decreased rate of tumor angiogenesis. Int J Cancer. 1988;42:48–52. doi: 10.1002/ijc.2910420110. [DOI] [PubMed] [Google Scholar]
- 37.Huang B, Lei Z, Zhang GM, et al. SCF-mediated mast cell infiltration and activation exacerbate the inflammation and immunosuppression in tumor microenvironment. Blood. 2008;112:1269–79. doi: 10.1182/blood-2008-03-147033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Munn DH, Mellor AL. The tumor-draining lymph node as an immune-privileged site. Immunol Rev. 2006;213:146–58. doi: 10.1111/j.1600-065X.2006.00444.x. [DOI] [PubMed] [Google Scholar]
- 39.Cuenca A, Cheng F, Wang H. Extra-lymphatic solid tumor growth is not immunologically ignored and results in early induction of antigen-specific T-cell anergy: dominant role of cross-tolerance to tumor antigens. Cancer Res. 2003;63:9007–15. [PubMed] [Google Scholar]
- 40.Gajewski TF, Meng Y, Blank C, et al. Immune resistance orchestrated by the tumor microenvironment. Immunol Rev. 2006;213:131–45. doi: 10.1111/j.1600-065X.2006.00442.x. [DOI] [PubMed] [Google Scholar]
- 41.Ochsenbein AF, Klenerman P, Karrer U, et al. Immune surveillance against a solid tumor fails because of immunological ignorance. Proc Natl Acad Sci USA. 1999;96:2233–8. doi: 10.1073/pnas.96.5.2233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sotomayor EM, Borrello I, Rattis DM, et al. Cross-presentation of tumor antigens by bone marrow-derived antigen-presenting cells is the dominant mechanism in the induction of T-cell tolerance during B-cell lymphoma progression. Blood. 2001;98:1070–7. doi: 10.1182/blood.v98.4.1070. [DOI] [PubMed] [Google Scholar]
- 43.Zou W. Regulatory T cells, tumour immunity and immunotherapy. Nat Rev Immunol. 2006;6:295–307. doi: 10.1038/nri1806. [DOI] [PubMed] [Google Scholar]
- 44.Chaux P, Favre N, Martin M, et al. Tumor-infiltrating dendritic cells are defective in their antigen-presenting function and inducible B7 expression in rats. Int J Cancer. 1997;72:619–24. doi: 10.1002/(sici)1097-0215(19970807)72:4<619::aid-ijc12>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 45.Scandella E, Men Y, Gillessen S, et al. Prostaglandin E2 is a key factor for CCR7 surface expression and migration of monocyte-derived dendritic cells. Blood. 2002;100:1354–61. doi: 10.1182/blood-2001-11-0017. [DOI] [PubMed] [Google Scholar]
- 46.Gosset P, Bureau F, Angeli V, et al. Prostaglandin D2 affects the maturation of human monocyte-derived dendritic cells: consequence on the polarization of naive Th cells. J Immunol. 2003;170:4943–52. doi: 10.4049/jimmunol.170.10.4943. [DOI] [PubMed] [Google Scholar]
- 47.Hammad H, de Heer HJ, Soullie T, et al. Prostaglandin D2 inhibits airway dendritic cell migration and function in steady state conditions by selective activation of the D prostanoid receptor 1. J Immunol. 2003;171:3936–40. doi: 10.4049/jimmunol.171.8.3936. [DOI] [PubMed] [Google Scholar]
- 48.Yen JH, Khayrullina T, Ganea D. PGE2-induced metalloproteinase-9 is essential for dendritic cell migration. Blood. 2007;111:260–70. doi: 10.1182/blood-2007-05-090613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Angeli V, Faveeuw C, Roye O, et al. Role of the parasite-derived prostaglandin D2 in the inhibition of epidermal Langerhans cell migration during schistosomiasis infection. J Exp Med. 2001;193:1135–47. doi: 10.1084/jem.193.10.1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Heib V, Becker M, Warger T, et al. Mast cells are crucial for early inflammation, migration of Langerhans cells, and CTL responses following topical application of TLR7 ligand in mice. Blood. 2007;110:946–53. doi: 10.1182/blood-2006-07-036889. [DOI] [PubMed] [Google Scholar]
- 51.Walsh LJ. Ultraviolet B irradiation of skin induces mast cell degranulation and release of tumor necrosis factor-alpha. Immunol Cell Biol. 1995;73:226–33. doi: 10.1038/icb.1995.37. [DOI] [PubMed] [Google Scholar]
- 52.Hart PH, Grimbaldeston MA, FInlay-Jones JJ. Sunlight, immunosuppression and skin cancer: role of histamine and mast cells. Clin Exp Pharmacol Physiol. 2001;28:1–8. doi: 10.1046/j.1440-1681.2001.03392.x. [DOI] [PubMed] [Google Scholar]
- 53.Hart PH, Townley SL, Grimbaldeston MA, et al. Mast cell, neuropeptides, histamine, and prostaglandins in UV-induced systemic immunosuppression. Methods. 2002;28:79–89. doi: 10.1016/s1046-2023(02)00201-3. [DOI] [PubMed] [Google Scholar]
- 54.Idzko M, la Sala A, Ferrari D, et al. Expression and function of histamine receptors in human monocyte-derived dendritic cells. J Allergy Clin Immunol. 2002;109:839–46. doi: 10.1067/mai.2002.124044. [DOI] [PubMed] [Google Scholar]
- 55.Ohtani T, Aiba S, Mizuashi M, et al. H1 and H2 histamine receptors are absent on Langerhans cells and present on dermal dendritic cells. J Invest Dermatol. 2003;121:1073–9. doi: 10.1046/j.1523-1747.2003.12570.x. [DOI] [PubMed] [Google Scholar]
- 56.Amaral MM, Davio C, Ceballos A, et al. Histamine improves antigen uptake and cross-presentation by dendritic cells. J Immunol. 2007;179:7891–8. doi: 10.4049/jimmunol.179.6.3425. [DOI] [PubMed] [Google Scholar]
- 57.Gutzmer R, Diestel C, Mommert S, et al. Histamine H4 receptor stimulation suppresses IL-12p70 production and mediates chemotaxis in human monocyte-derived dendritic cells. J Immunol. 2005;174:5224–32. doi: 10.4049/jimmunol.174.9.5224. [DOI] [PubMed] [Google Scholar]
- 58.Gombert M, Dieu-Nosjean MC, Winterberg F, et al. CCL1-CCR8 interactions: an axis mediating the recruitment of T cells and Langerhans-type dendritic cells to sites of atopic skin inflammation. J Immunol. 2005;174:5082–91. doi: 10.4049/jimmunol.174.8.5082. [DOI] [PubMed] [Google Scholar]
- 59.Wedemeyer J, Galli SJ. Decreased susceptibility of mast cell-deficient KitW/KitW-v mice to the development of 1, 2-dimethylhydrazine-induced intestinal tumors. Lab Invest. 2005;85:388–96. doi: 10.1038/labinvest.3700232. [DOI] [PubMed] [Google Scholar]
- 60.Askenasy N, Kaminitz A, Yarkoni S. Mechanisms of T regulatory cell function. Autoimmun Rev. 2008;7:370–5. doi: 10.1016/j.autrev.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 61.Aune TM, Golden HW, McGrath KM. Inhibitors of serotonin synthesis and antagonists of serotonin 1A receptors inhibit T lymphocyte function in vitro and cell-mediated immunity in vivo. J Immunol. 1994;153:489–98. [PubMed] [Google Scholar]
- 62.Laberge S, Cruikshank WW, Beer DJ, et al. Secretion of IL-16 (lymphocyte chemoattractant factor) from serotonin-stimulated CD8+ T cells in vitro. J Immunol. 1996;156:310–5. [PubMed] [Google Scholar]
- 63.León-Ponte M, Ahern GP, O'Connell PJ. Serotonin provides an accessory signal to enhance T-cell activation by signaling through the 5-HT7 receptor. Blood. 2007;109:3139–46. doi: 10.1182/blood-2006-10-052787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nakae S, Suto H, Iikura M, et al. Mast cells enhance T cell activation: importance of mast cell costimulatory molecules and secreted TNF. J Immunol. 2006;176:2238–48. doi: 10.4049/jimmunol.176.4.2238. [DOI] [PubMed] [Google Scholar]
- 65.Keir ME, Butte MJ, Freeman GJ, et al. PD-1 and its ligands in tolerance and immunity. Annu Rev Immunol. 2008;26 doi: 10.1146/annurev.immunol.26.021607.090331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Zou W, Chen L. Inhibitory B7-family molecules in the tumour microenvironment. Nat Rev Immunol. 2008;8:467–77. doi: 10.1038/nri2326. [DOI] [PubMed] [Google Scholar]
- 67.Aubin F. Mechanisms involved in ultraviolet light-induced immunosuppression. Eur J Dermatol. 2003;13:515–23. [PubMed] [Google Scholar]
- 68.Nordlund JJ, Askenase PW. The effect of histamine, antihistamine, and mast cell stabilizer on the growth of Cloudman melanoma cells in DBA/2 mice. J Invest Dermatol. 1983;81:28–31. doi: 10.1111/1523-1747.ep12538356. [DOI] [PubMed] [Google Scholar]
- 69.Boerma M, Fiser WP, Hoyt G, et al. Influence of mast cells on outcome after heterotopic cardiac transplantation in rats. Transpl Int. 2007;20:256–65. doi: 10.1111/j.1432-2277.2006.00420.x. [DOI] [PubMed] [Google Scholar]
- 70.Colvin RB, Dvorak HF. Basophils and mast cells in renal allograft rejection [Letter. Lancet. 1974;1:212–4. doi: 10.1016/s0140-6736(74)92512-4. [DOI] [PubMed] [Google Scholar]
- 71.Yousem SA. The potential role of mast cells in lung allograft rejection. Hum Pathol. 1997;28:179–82. doi: 10.1016/s0046-8177(97)90103-9. [DOI] [PubMed] [Google Scholar]