

Abstract
In a previous study we used event-related potentials to dissociate semantic (associative) and repetition priming when prime words were masked below (40 ms primes) and above (80 and 120 ms primes) levels of awareness (Holcomb et al., 2005). While robust priming effects were present on the N400 component regardless of the prime duration in the repetition priming experiment, in the semantic priming experiment significant priming effects were only obtained when primes were consciously detected. In the current study we again compared associative and repetition priming, but this time within a single experiment and using a short interval priming paradigm. As in several recent studies we again found robust repetition priming effects on both the N400 and an earlier index of priming, the N250. However, there was no evidence of any priming effect in the semantic/associative condition. The findings are discussed in the context of the functional significance of masked priming effects.
Keywords: ERPs, N250, N400, Masked Priming, Semantic Priming, Repetition Priming
1. Introduction
Numerous studies have shown that the processing of a target word can be influenced by the nature of its relationship with a preceding prime word. For example, when the prime and target are semantically related (e.g., doctor-NURSE) or are the same word (e.g., nurse-NURSE) versus when they are unrelated words (e.g., knife-NURSE), subjects are typically faster and more accurate at making a variety of judgments about the targets (e.g., Meyer & Schvaneveldt, 1971; Norris, 1984). Such effects have played an important role in constraining theories of word processing and lexical/semantic organization.
While priming effects are most readily obtained when subjects can easily perceive the relationship between the prime and target stimuli, it is also possible to observe priming effects even when subjects are unaware of the relationship, or even the presence of the prime itself (e.g., Forster & Davies, 1984). For example, presenting a prime word very briefly and sandwiching it between a forward and backward masking stimulus, will make the prime difficult to perceive. However, if the prime parameters are set correctly, the influence of the prime on target processing can nevertheless be demonstrated and usually takes the form of facilitation of target processing if the prime and target are related. Such masked priming results are thought to reflect primarily the early automatic components of word processing (e.g., Forster & Davies, 1984).
Many studies have also shown that repetition and semantic priming both produce characteristic differences in ERPs. The most frequently reported effect is that primed words attenuate the N400 component compared to non-primed words (e.g., Bentin et al., 1985; Holcomb, 1988). Such N400 “effects” are believed by many to be sensitive to the lexical and/or semantic properties of the stimulus and its context (e.g., Holcomb, 1993). According to this view words that are easily integrated with their contextual framework produce an attenuated N400, while those that are impossible or difficult to integrate with the surrounding context generate larger N400s.
There is some controversy, however, as to whether N400 effects might also require conscious processing. Brown and Hagoort (1991) were the first who sought to determine if there was an unconscious/automatic contribution to the N400 in semantic priming. Using both masked and unmasked semantic primes they found evidence for modulation of target N400s only when primes were not masked. They concluded that the N400 is sensitive to higher order semantic integration and not to automatic lexical access. Subsequent studies (e.g., Deacon et al., 2000; Grossi, 2006; Kiefer & Spitzer, 2000; Kiefer, 2002) have challenged this view based on their having found significant masked semantic priming effects on the N400. However, recently Holcomb et al. (2005) demonstrated that while N400 masked repetition priming effects were unrelated to the detectability of primes, masked semantic priming effects were predicted by prime detectability. This suggests that previous demonstrations of masked semantic priming might be due to what Holcomb et al referred to as “conscious leakage” of semantic information from the masked primes.
One potential problem with the Holcomb et al. (2005) conclusion is that their study used a relatively long prime-target stimulus-onset-asynchrony (SOA) of about 500 ms. Grossi (2006) has argued that some of the semantic information activated by the prime may decay over this interval, and her study, which used a brief 50 ms prime-target SOA, found evidence of robust masked semantic priming on the N400 component. In support of this decay hypothesis Holcomb & Grainger (2007) have recently shown that prime-target SOA has a dramatic effect on the size of ERP masked repetition priming effects, with shorter intervals (< 300 ms) producing larger effects in some cases.
2. The Current Study
The current study further investigates masked repetition and semantic priming. Two modifications were made to the Holcomb et al. (2005) paradigm. First, semantic/associated and repetition trials were included within a single session. Holcomb et al (2005) used two separate experiments with two different groups of subjects for their semantic and repetition priming effects. It could be argued that different strategies due to the mix of repeated versus semantic pairs altered the pattern of priming across experiments. Second, the priming paradigm used by Holcomb et al (2005) had a long prime-target SOA of 500 ms. In the current study the interval between primes and targets was a short 70 ms. Using a short prime-target SOA, Holcomb and Grainger (2006; 2007) also found that the N400 component was larger to target words following unrelated primes than primes that were identical to (table-TABLE) or nearly identical to (teble-TABLE) the target. However, they also found repetition effects on earlier ERP components as well. The most interesting for the current study was a widely distributed ERP negativity that started around 170ms and peaked near 250 ms. Holcomb and Grainger referred to this as the N250 component, and argued that it was sensitive to the degree of orthographic overlap between the prime and target stimuli. They found that the amplitude of the N250 was proportional to the number of letters that the prime and target words had in common, being largest when primes and targets shared no letters, intermediate when prime and target shared several letters (partial repetition) and smallest when they shared all letters (full repetition).
The current experiment sought to determine if there are different target ERP effects produced by masked repetition and associative priming when the prime-target SOA was short. Participants were presented with pairs of items consisting of a brief pattern masked prime word and a subsequent clearly visible target word. Masked primes could be followed by a clearly visible target word that was either a repetition of the prime word (repeated targets), a semantic associate of the target (associated targets) or another unrelated word (unrelated targets). Primes were sandwiched between two pattern masks (forward and backward) and had a duration of 50 ms. Participants performed a semantic categorization task in which they were instructed to monitor all stimuli for occasional exemplars (probes) from the category of animal names, and to press a button when such items were detected. No response was required to the critical prime or target stimuli. One feature of this task is that it provides a measure of the effectiveness of the masking procedure by allowing subjects to respond on trials with animal names in the prime position.
The following predictions were made. First, following Holcomb and Grainger (2006) it was predicted that there would be an attenuation of both the N250 and N400 components to target words immediately preceded by identity masked primes as compared to target words preceded by an unrelated masked prime words. This is because the N250 is thought to be sensitive to processing at the interface between sub-lexical orthographic and lexical representations and the N400 either because of processing at the lexical form and semantic interface (Grainger & Holcomb, in press) or within the semantic system itself. Second, following Grossi (2006) it was predicted that there would be an attenuation of the N400 (but not the N250) to target words immediately preceded by semantically associated primes as compared to target words preceded by an unrelated masked prime words. N400 effects are predicted because of the semantic overlap between prime and target words for associated but not unrelated pairs. No N250 effects were predicted for the associatively related pairs because these items have no more overlap at the orthographic level of representation than unrelated items.
3. Methods
3.1. Participants
Twenty-one right-handed, native English speaking Tufts University students (8 male, mean age = 20.11) received $20 compensation for participation in this experiment.
3.2. Stimuli and Procedure
All words used in the current experiment were 4 to 6 letters in length. The critical stimuli for this experiment were formed from 480 nouns, with frequency less than 30 per million. An additional set of non-critical stimulus pairs were formed by combining 90 different animal names (frequency < 30/million) with 90 unrelated non-animal words (nouns; frequency < 50/million). For both the critical and non-critical items the first member of each pair was referred to as the prime and the second member as the target. Stimulus lists consisted of 330 trials each containing a prime-target pair of words. There were three critical stimulus conditions in each list: repetitions, which contained 80 pairs of repeated words (e.g. chair-CHAIR), associated words, which contained 80 pairs of semantically associated words (e.g., table-CHAIR, mean forward associative strength = .31 on the South Florida University Word Association norms, http://w3.usf.edu/FreeAssociation) and unrelated words, which contained 80 pairs of words with semantically unrelated primes and targets (e.g. uncle-CHAIR). For the 90 non-critical pairs 45 contained animal names in the prime position and an unrelated word in the target position, while the other 45 trials contained an unrelated word in the prime position and an animal name in the target position. The animal names served as probe items in a go/no-go semantic categorization task in which participants were instructed to rapidly press a single button (response hand counterbalanced) whenever they detected an animal name in either the prime or target position. Participants were told to read all other words passively (i.e., critical stimuli did not require an overt response).
Seven different lists were formed so that each critical item appeared once per list, but across lists each critical item appeared in each of the critical trial positions (repeated, associated and unrelated). This assures that when comparing ERPs between related and unrelated conditions and across the Repetition and Associated conditions that waves are formed from the same stimulus items.
All stimuli were displayed in the center of a 19 inch monitor as white letters on a black background in the Arial font. The schematic of a trial is displayed in Figure 1. As can be seen, each trial began with the termination of a fixation stimulus and 500 ms later a forward pattern mask was presented for 500 ms (#########). The mask was immediately replaced at the same location on the screen by the prime word in lower case letters (e.g., table) and was displayed for 50 ms. The prime was then replaced by a 20 ms backward mask (#########), which was in turn replaced by a 500 ms target word (e.g., TABLE, CHAIR or UNCLE). Target words were presented in upper case letters and was followed by a blank screen for 1000 ms. Finally the end of the trial was signaled by the fixation stimulus which was presented for 2000 ms. Subjects were asked to blink only when the fixation/blink stimulus was on the screen.
Figure 1.
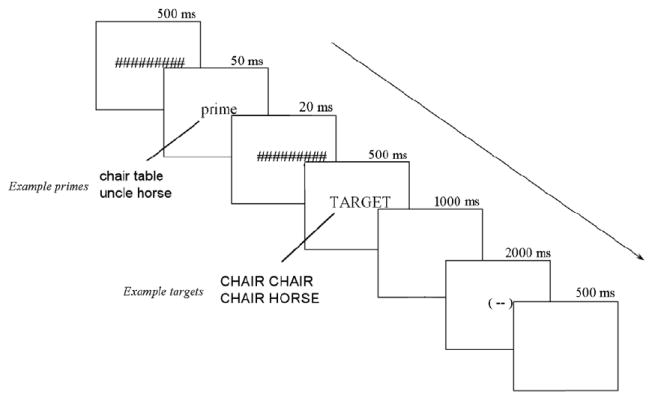
Schematic of different trial types.
3.3. EEG Recording Procedure
Participants were seated in a comfortable chair in sound attenuated darkened room. An electro-cap with tin electrodes was used to record continuous EEG from 29 sites on the scalp including sites over left and right fronto-polar (FP1/FP2), frontal (F3/F4, F7/F8), frontal-central (FC1/FC2, FC5/FC6), central (C3/C4), temporal (T5/T6, T3/T4), central-parietal (CP1/CP2, CP5/CP6), parietal (P3/P4), and occipital (O1/O2) areas and five midline sites over the frontal pole (FPz), frontal (Fz), central (Cz), parietal (Pz) and occipital (Oz) areas (see Figure 2). In addition, four electrodes were attached to the face and neck area: one below the left eye (to monitor for vertical eye movement/blinks), one to the right or the right eye (to monitor for horizontal eye movements), one over the left mastoid (reference) and one over the right mastoid (recorded actively to monitor for differential mastoid activity). All EEG electrode impedances were maintained below 5 kΩ (impedance for eye electrodes was less than 10 kΩ). The EEG was amplified by an SA Bioamplifier with a bandpass of 0.01 and 40 Hz and the EEG was continuously sampled at a rate of 250 Hz.
Figure 2.
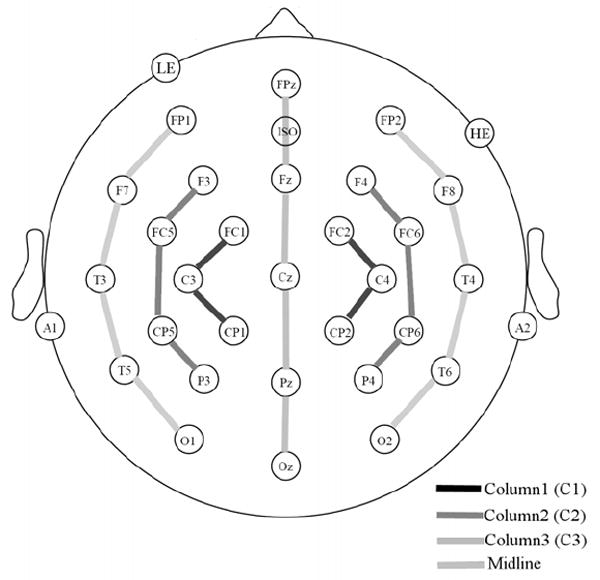
The electrode montage and columns used in analyses.
3.4. Data and Analysis
Separate waveforms were calculated by averaging compound ERPs to prime and target words based on the three prime-target conditions (Repeated, Associated/Semantic, Unrelated). Only trials without muscle artifact or eye movement/blink activity were included in the averaging process. Mean target amplitudes for the same three latency windows used by Holcomb & Grainger (2007) were used to quantify the target ERPs in this experiment: 0-175 ms, 175-300 ms and 300-550 ms.
Separate analyses of variance (ANOVAs) were performed using the within-subject factors of prime duration (short, medium, long) and repetition (repeated, unrepeated) for each of four sub-montages of electrode site columns: midline (FPz, Fz, Cz, Pz, Oz), Column1 (FC1, C3, CP1, FC2, C4, CP2), Column2 (F3, FC5, CP5, P3, F4, FC6, CP6, P4) and Column3 (FP1, F7, T3, T5, O1, FP2, F8, T4, T6, O2 – see Figure 2 for columns). Column1, Column2 and Column3 also included a factor of hemisphere (left vs. right).
4. Results
4.1. Event-related Potentials
Plotted in Figure 3 are the ERPs (prime and target) contrasting the conditions with repeated, associated and unrelated targets. Plotted in Figure 4 is a blow up of target ERPs at the CP1 site. As can be seen in these figures, the ERPs in this experiment produced a set of positive and negative deflections. It is important to keep in mind that the early deflections (P1, N1 and P2) to both the prime and target words are an amalgamation of overlapping components produced by the rapid succession of mask-prime-mask-target stimuli.
Figure 3.
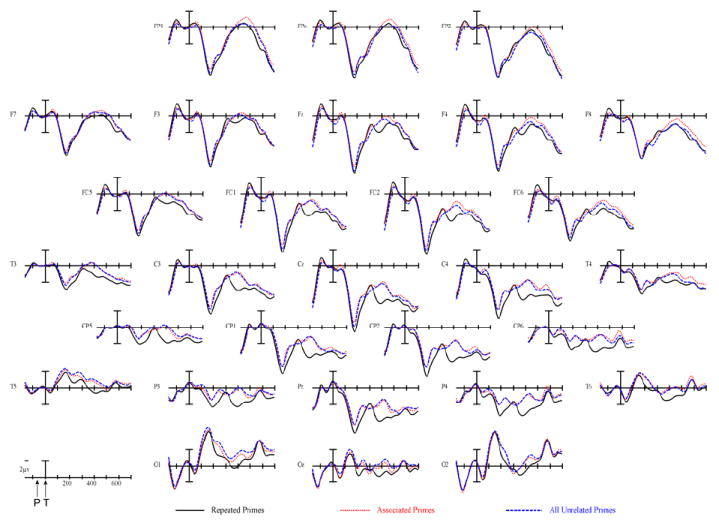
Compound ERPs time locked to prime onset in the Repeated, Associated and Unrelated conditions. Note that target onset is at the large vertical calibration bar (indicated by the “T” below the time-axis in the lower left of the figure) and prime onset is 70 ms earlier (indicated by the “P”). Negative voltages are plotted in the upward direction.
Figure 4.
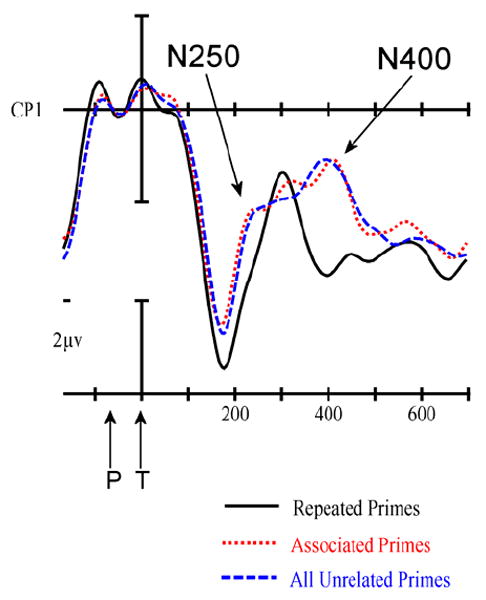
A blow-up for the CP1 site from Figure 3.
4.1.1. 0-175 ms Target Epoch
Although Figure 3 and 4 suggest that the first effects of priming begin towards the end of this epoch, there were no significant effects in this epoch.
4.1.2. 175-300 ms Target Epoch
As can be seen in Figure 3-4, the effects of priming are clearly visible in this epoch (main effect of prime-target type, midline: F(2,40) = 4.95, p = .019; Column1: F(2,40) = 4.89, p = .019; Column2: F(2,40) = 4.52, p = .023; and Column3: F(2,40) = 5.23, p = .019). Planned follow-up analyses contrasting the two types of primed targets (Repeated and Associated) separately to the Unrelated targets revealed that Repeated targets were significantly less negative going than Unrelated targets (main effect of prime-target type, midline: F(1,20) = 5.32, p = .032; Column1: F(1,20) = 5.00, p = .037; Column2: F(1,20) = 5.03, p = .036; and Column3: F(1,20) = 5.18, p = .032), but there was no evidence of a difference between Associated and Unrelated (all Fs < 1.0).
4.1.3. 300-550 ms Target Epoch
As can be seen in Figures 3 and 4 there were clear effects of priming in this epoch across the scalp (main effect of repetition, F(2,40) = 8.05, p = .002; Column1: F(2,40) = 9.70, p = .001; Column2: F(2,40) = 9.27, p = .001; and Column3: F(2,40) = 6.64, p = .005). Planned follow-up analyses contrasting the two types of primed targets (Repeated and Associated) separately to the Unrelated targets revealed that Repeated targets were significantly less negative going than Unrelated targets (main effect of prime-target type, midline: F(1,20) = 9.62, p = .006; Column1: F(1,20) = 9.98, p = .002; Column2: F(1,20) = 11.66, p = .003; and Column3: F(1,20) = 6.25, p = .021), but there was no evidence of a difference between Associated and Unrelated (all Fs < 1.1).
4.2. Behavioral Data
Participants detected an average of 87% of animal probes in the target position within a window of 200 to 1500 ms post-probe onset. In the prime position participants detected 4% of animal probes.
5. Discussion
Like several previous studies, robust effects of repetition in the region of the N400 component were found for target words that were preceded by very briefly displayed masked identity prime words (Holcomb et al., 2005; Holcomb & Grainger, 2006; 2007; Kiyonaga et al., 2007). There were also clear effects of repetition on an earlier component peaking near 250 ms. Holcomb and Grainger referred to this as the N250 and argued that it reflects the influence of orthographic overlap between the prime and target words. Of most interest was the complete failure to find any evidence of priming on either the N250 or N400 components for targets that were semantically associated with the prior prime stimulus. Remarkably, the ERPs to associated targets appeared to be in near perfect registration with the ERPs to the semantically unrelated control words throughout the entire post-target epoch.
Because the semantic priming result is null, it is important that it occurred in the presence of robust repetition priming and that these repetition effects occurred on the same target items and in the same participants where we failed to find any evidence for priming of associated word pairs. This is compelling evidence that semantic priming does not occur when primes are effectively masked below normal awareness levels. This conclusion is consistent with our previous study in which masked semantic priming effects were found to correlate with detectability of the prime stimulus (Holcomb et al., 2005). In that study, which used a long prime-target SOA, there was a range of performance by participants in their detection of masked primes. In the current study with a short prime-target interval, virtually no primes were detected on the trials designed to tap subject awareness of the masked primes (only one subject detected more than 2 primes).
The above pattern of effects has important implications for understanding the mechanisms underlying masked priming as well as the processing nature of ERP components sensitive to manipulations in this paradigm. But before we turn to these issues, an important question that needs to be addressed is why have several previous studies, at least one of which used a similar short interval masking paradigm (Grossi, 2006), reported significant semantic priming effects on the N400? The most likely explanation is the one we offered in Holcomb et al (2005), that is, that these previous studies obtained semantic effects because they mixed trials or subjects where some of the primes were consciously processed. If even some of the primes are consciously processed then any ERP effects obtained could be due to the influence of those items. Consistent with this possibility is the observation that masked priming effects tend to be smaller than unmasked priming. To assess subject awareness of primes it would seem to be critical to have a measure of prime awareness built in the actual task where the effects of priming are being tested. Otherwise differing demand characteristics between the assessment task and the priming task could result in an underestimation of prime awareness. None of the previous ERP semantic masked priming studies have used such a procedure. Another issue that could explain the different results of Grossi (2006) and the current study, both of which used the short interval masked priming paradigm, is that we used a full Latin Square counterbalancing of items between unrelated control and semantic priming conditions. That is, our ERPs were formed from the exact same items (primes and targets) in the related and unrelated conditions. Grossi used different lexical items for related and unrelated trials (albeit matched for length and frequency). This latter design always carries the risk that obtained differences are the result of some uncontrolled lexical or semantic factor.
5.1. Prime visibility and effects of repetition and semantic priming
The present study provides further evidence that masked repetition priming occurs in conditions where prime visibility is severely limited, whereas semantic/associative priming does not. We will discuss several possible ways (which are not mutually exclusive) of accommodating this specific pattern of priming effects. The first two proposals were put forward by Holcomb et al. (2005) in order to account for their observation that the size of semantic priming effects depended on prime exposure duration, whereas the size of repetition priming effects did not, and are directly applicable to the present findings.
The first proposal hinges on the hypothesis that form information can be extracted automatically (i.e., without attention-demanding resources), while the extraction of semantic information is either not automatic at all and depends on the allocation of capacity-limited resources or relies to a greater extent on capacity-limited resources. If it is further hypothesized that it is precisely the allocation of attention-demanding resources that determines the level of visibility of prime stimuli, then this explains why semantic priming does not occur in the absence of prime visibility. However, one also has to assume that most of the effect in repetition priming is driven by shared form representations.
Holcomb et al. (2005) suggested an alternative interpretation that would avoid having such a relatively ad-hoc assumption. According to this second proposal, the critical difference between repetition and semantic priming is that repetition priming is driven primarily by the pre-activation of representations that are shared by prime and target, while semantic priming involves, to a greater extent, establishing connections across related representations (e.g., as in theories such as compound-cue, Ratcliff & McKoon, 1988). In this account it is not the type of representation (form or meaning) that determines the automaticity of priming effects, but rather the type of mechanism utilized (pre-activation versus establishing connections). Thus repetition priming is more automatic in that it relies relatively more on pre-activation of form and meaning representations, whereas semantic priming is less automatic because it relies relatively more on connecting up the representations of related concepts after the target has been at least partially processed. The latter post-target connecting mechanism would require allocation of attention-demanding resources thus explaining why semantic priming emerges with more visible primes.
A third proposal is that a minimal amount of bottom-up activation is first necessary in order for target stimuli to benefit from semantic overlap with prime stimuli. When prime and target mismatch completely in terms of orthography and phonology, then transfer of higher-level semantic information from prime to target could be blocked. In other words, any information extracted from the prime stimulus is simply ignored during target processing when there is not enough bottom-up form overlap to get the process initiated. Another way of looking at this proposal is in terms of backward priming from target to prime stimulus that allows processing of the prime stimulus to continue when the target stimulus appears. Shared form representations across prime and target would allow processing of the prime to continue until semantic information is accessed. According to this account, extraction of semantic information from prime stimuli can proceed automatically in the absence of awareness, but this process is blocked by concurrent processing of the target. In line with this proposal are the results of one recent study (Morris, Frank, Grainger, & Holcomb, 2007) showing that the N400 is sensitive to semantic relatedness across prime and target when primes and targets also overlap orthographically, as is the case with morphologically related primes and targets (e.g., baker-bake compared with corner-corn).
Finally, one could also argue that the level of semantic overlap is simply not great enough, and semantic processing not rapid enough to resist the effects of pattern masking and limited prime duration used in the present study. Repetition priming involves full semantic overlap (as well as full form overlap) between primes and targets, and this extra overlap might just tip the balance in favor of obtaining priming effects. Against this account, however, is the fact that there was no hint of an effect of semantic priming in the present study. According to this account one might have expected to observe at least a trend toward a priming effect, but this was clearly not the case. Furthermore, in prior research we did find significant modulation of N250 and N400 amplitudes with primes that only partially overlapped orthographically with targets (e.g., teble-table: Holcomb & Grainger, 2006), showing that graded effects of orthographic overlap can be observed in the same testing conditions as the present study.
Future research could test these alternative hypotheses using the same manipulation as in the present study but with translation equivalents in bilingual persons (e.g., arbre-TREE for a French-English bilingual). If the reason why we failed to find semantic/associative priming in the present study is due to a fundamental limitation in the way semantic information can be extracted from masked prime stimuli, either because these are attention-demanding processes or because target processing blocks higher-level processing of prime stimuli when primes do not share orthographic information with targets, then one would not expect to observe an effect of translation primes in bilinguals. On the other hand, if the absence of semantic priming is due to insufficient semantic overlap across prime and target, or because semantic priming operates via establishing connections between representations (as opposed to preactivation of shared representations), then one ought to be able to observe translation priming effects in the testing conditions of the present study.
5.2. Implications for the Functional Significance of the N250 and N400
The fact that N400 amplitude was significantly modulated by repetition primes in this and prior masked priming studies (e.g., Chauncey, Holcomb, & Grainger, 2008; Holcomb & Grainger, 2006; Kiyonaga et al., 2007) would seem to call into question the interpretation of the functional significance of the N400 offered by Brown and Hagoort (1993) and others who have maintained that the N400 is exclusively sensitive to post-lexical semantic processes. Our repetition priming effects would appear to fit better with Holcomb, Grainger, and O’Rourke’s (2002) proposal that the N400 might partly reflect processing at a form-meaning interface that is sensitive to the compatibility of co-activated form (orthographic and phonological) and meaning representations. Thus modulations of the N400 could reflect the influence of form and/or meaning information extracted from prime stimuli on processing of target stimuli at this form-meaning interface. According to this interpretation of the N400, the absence of semantic priming in the present study is attributed to limitations either in the amount or in the type of semantic processing (e.g., attention-demanding processes) that can be achieved with brief prime durations. The N400 component is modulated by semantically related primes with longer prime durations (e.g., Holcomb et al., 2005).
An earlier ERP component, the N250, was also found to be modulated by repetition priming in the present study, thus replicating our prior observations of this effect (Chauncey et al., 2008; Holcomb & Grainger, 2006; Kiyonaga et al., 2007). Like the N400, this component was not modulated by prime-target semantic relatedness in the present study. The absence of semantic priming effects on the N250 fits with the functional interpretation of this component proposed by Holcomb and Grainger (2006) as reflecting prelexical orthographic and phonological processing (see also Grainger, Kiyonaga, & Holcomb, 2006). However, Morris et al. (2007) have shown that the N250 (as well as the N400, as noted above) is sensitive to semantic relatedness when primes and targets are morphologically related. This particular result can be accommodated in a cascaded interactive-activation model (McClelland & Rumelhart, 1981) that allows fast access to semantic representations on the basis of partial bottom-up information. This semantic information could then feedback to influence on-going prelexical processing of the target word.
Finally, it is important to note that the N250 component is typically not found with longer prime-target intervals (e.g., Holcomb et al., 2005), and it is not found when primes and targets are in different modalities (Kiyonaga et al., 2007). This again fits with the proposal that this component reflects early prelexical processing of printed word stimuli, processing that would be modality-specific and that would decay rapidly following prime offset.
Footnotes
This research was supported by HD25889 and HD043251
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bentin S, McCarthy G, Wood CC. Event-related potentials, lexical decision and semantic priming. Electroencephalography and Clinical Neurophysiology. 1985;60:343–355. doi: 10.1016/0013-4694(85)90008-2. [DOI] [PubMed] [Google Scholar]
- Brown C, Hagoort P. The Processing Nature of the N400: Evidence from Masked Priming. Journal of Cognitive Neuroscience. 1993;5(1):34–44. doi: 10.1162/jocn.1993.5.1.34. [DOI] [PubMed] [Google Scholar]
- Chauncey K, Holcomb PJ, Grainger J. Effects of stimulus font and size on masked repetition priming: An ERP investigation. Language and Cognitive Processes. 2008 doi: 10.1080/01690960701579839. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chwilla DJ, Brown CM, Hagoort P. The N400 as a function of the level of processing. Psychophysiology. 1995;32(3):274–285. doi: 10.1111/j.1469-8986.1995.tb02956.x. [DOI] [PubMed] [Google Scholar]
- Deacon D, Hewitt S, Yang C-M, Nagata M. Event-related potential indices of semantic priming using masked and unmasked words: Evidence that the N400 does not reflect a post-lexical process. Cognitive Brain Research. 2000;9(2):137–146. doi: 10.1016/s0926-6410(99)00050-6. [DOI] [PubMed] [Google Scholar]
- Forster KI, Davis C. Repetition priming and frequency attenuation in lexical access. Journal of Experimental Psychology: Learning, Memory, & Cognition. 1984;10(4):680–698. [Google Scholar]
- Grossi G. Relatedness proportion effects on masked associative priming: An ERP study. Psychophysiology. 2006;43:21–30. doi: 10.1111/j.1469-8986.2006.00383.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Automatic and Attentional Processing: An Event-Related Brain Potential Analysis of Semantic Priming. Brain and Language. 1988;35:66–85. doi: 10.1016/0093-934x(88)90101-0. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Semantic priming and stimulus degradation: implications for the role of the N400 in language processing. Psychophysiology. 1993;30(1):47–61. doi: 10.1111/j.1469-8986.1993.tb03204.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J, O’Rourke T. An event-related brain potential study of orthographic similarity. Journal of Cognitive Neuroscience. 2002;14:938–950. doi: 10.1162/089892902760191153. [DOI] [PubMed] [Google Scholar]
- Kiefer M. The N400 is modulated by unconsciously perceived masked words: further evidence for an automatic spreading activation account of N400 priming effects. Cognitive Brain Research. 2002;13(1):27–39. doi: 10.1016/s0926-6410(01)00085-4. [DOI] [PubMed] [Google Scholar]
- Kiefer M, Spitzer M. Time course of conscious and unconscious semantic brain activation. Neuroreport. 2000;11(11):2401–2407. doi: 10.1097/00001756-200008030-00013. [DOI] [PubMed] [Google Scholar]
- Kiyonaga K, Grainger J, Midgley KJ, Holcomb PJ. Masked cross-modal repetition priming: An event-related potential investigation. Language and Cognitive Processes. 2007;22:337–376. doi: 10.1080/01690960600652471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer DE, Schvaneveldt RW. Facilitation in recognizing pairs of words: Evidence of a dependence between retrieval operations. Journal of Experimental Psychology. 1971;90(2):227–234. doi: 10.1037/h0031564. [DOI] [PubMed] [Google Scholar]
- Misra M, Holcomb PJ. Event-related potential indices of masked repetition priming. Psychophysiology. 2003;40:115–130. doi: 10.1111/1469-8986.00012. [DOI] [PubMed] [Google Scholar]
- Morris J, Franck T, Grainger J, Holcomb PJ. Semantic transparency and masked morphological priming: AN ERP investigation. Psychophysiology. 2007;44:506–521. doi: 10.1111/j.1469-8986.2007.00538.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norris J. The effects of frequency, repetition and stimulus quality in visual word recognition. Quarterly Journal of Experimental Psychology. 1984;36A:507–518. [Google Scholar]
- Shnyer DM, Allen JB, Forster KI. Event-Related Brain Potential Examination of Implicit Memory Processes: Masked and Unmasked Repetition Priming. Neuropsychology. 1997;11(2):243–260. doi: 10.1037//0894-4105.11.2.243. [DOI] [PubMed] [Google Scholar]