

Abstract
Previous research suggests that action video game play improves attentional resources, allowing gamers to better allocate their attention across both space and time. In order to further characterize the plastic changes resulting from playing these video games, we administered the Attentional Network Test (ANT) to action game players and non-playing controls aged between 7 and 22 years. By employing a mixture of cues and flankers, the ANT provides measures of how well attention is allocated to targets as a function of alerting and orienting cues, and to what extent observers are able to filter out the influence of task irrelevant information flanking those targets. The data suggest that action video game players of all ages have enhanced attentional skills that allow them to make faster correct responses to targets, and leaves additional processing resources that spill over to process distractors flanking the targets.
Attentional networks and their development in action video game players
Recently, we and others have shown that playing action video games alters some of the fundamental aspects of visual attention (Bialystok, 2006; Castel et al., 2005; Green & Bavelier, 2003, 2006a, b; Greenfield et al., 1994; Trick et al., 2005). Expert action video game players (VGPs) were found to outperform non-gamer controls (NVGPs) on tasks measuring the spatial distribution and resolution of visual attention, the efficiency of visual attention over time and the number of objects that can be attended simultaneously (Green and Bavelier, 2003). Training studies demonstrating the causal effect of game playing on visual attention measures have led to the proposal that action video game playing enhances attentional resources (Green & Bavelier, 2003; Green & Bavelier, 2006a, b) and allows players of such games to better allocate their attentional resources over a visual scene.
In the present study, we propose to further document the effect of video game playing on the efficiency of attention allocation by comparing VGPs and NVGPs on the Attentional Network Test (ANT). It has been suggested that the ANT provides a reliable measure of three fundamental component processes of visual attention within one procedure: alerting, orienting and executive control (Fan et al., 2002). Alerting is the ability to make use of a cue which provides information about the onset time of a target stimulus, and thus trigger the allocation of attention at a given point in time; this process appears mediated by right frontal and parietal areas and to be linked to the release of noradrenalin (Coull et al., 1996; Witte & Marrocco, 1997). Orienting is the ability to utilize an spatial cue to direct attention towards the location of an imminent stimulus; a fronto-parietal network associated with the release of acetylcholine has been associated with orienting (Corbetta & Shulman, 1998; Wilson et al., 2005). Finally, the executive control network, which serves to direct attention towards task-relevant stimuli and inhibit the processing of distractor items, has been proposed to engage areas in the prefrontal cortex and involve the release of dopamine (Badre & Wagner, 2004; Casey et al., 2000; Diamond & Goldman-Rakic, 1989; Egner & Hirsch, 2005; Nelson et al., 2003). Thus, the ANT provides a measure of how well attention can be both allocated to a visual scene and used to filter irrelevant information within that scene. The task has also been shown to be sensitive to developmental changes in all three networks (Rueda et al., 2004).
Procedurally, the ANT requires subjects to discriminate the orientation of a target (pointing left or pointing right) that is presented either directly above or below a central fixation point. The efficiency of the alerting network is measured by contrasting trials where the target is uncued (no cue: location unknown and onset unknown) with trials where both possible locations are cued simultaneously (double cue: location unknown and onset known). Orienting efficiency is measured by comparing trials where a cue appears at the fixation point (center cue: location unknown and onset known) with those where the location of the following target is cued (spatial cue: location known and onset known). Finally, distractors also flank the target on some trials. The flankers can be either congruent with target (point in the same direction) or incongruent (point in the opposite direction). By contrasting trials with congruent and incongruent flankers, Fan et al. (2002) suggest that a measure of the efficiency of ‘executive control’ can be indexed. More specifically, this contrast appears to measure the efficiency of filtering by computing a flanker interference effect, or how successfully flanker information can be ignored and attention focused upon the task-relevant stimulus target. As discussed in Callejas et al. (2004), Dye et al. (2007) and Fernandez-Duque & Posner (1997), the components of the ANT can also be understood in terms of how it manipulates the allocation of attention upon task-relevant stimuli in a visual scene. The cueing effects that the ANT measures are henceforth given an alternative meaning. The alerting cue serves to shift the focus of attention from a diffuse distribution across the display prior to the alerting cue, to a focus upon the display area where targets are known to occur following the alerting cue (see Figure 1). The orienting cue is a valid spatial cue that focuses attention even more sharply upon the actual location of the upcoming target. This hypothesis explains the nature of the interactions between these types of cue and the influence of distractors flanking the targets, as well as accounting for the changes in those interactions as flanker eccentricity is manipulated (see Dye et al., 2007 for a more detailed discussion). We therefore use the ANT here as a measure of how efficiently an observer can use the cues to allocate their attention appropriately across a visual display and then succesfully filter out stimuli that are task irrelevant. In accordance with this proposal, we will refer to alerting, orienting and flanker compatibility effects, rather than to alerting, orienting and executive control network efficiency.
Figure 1.
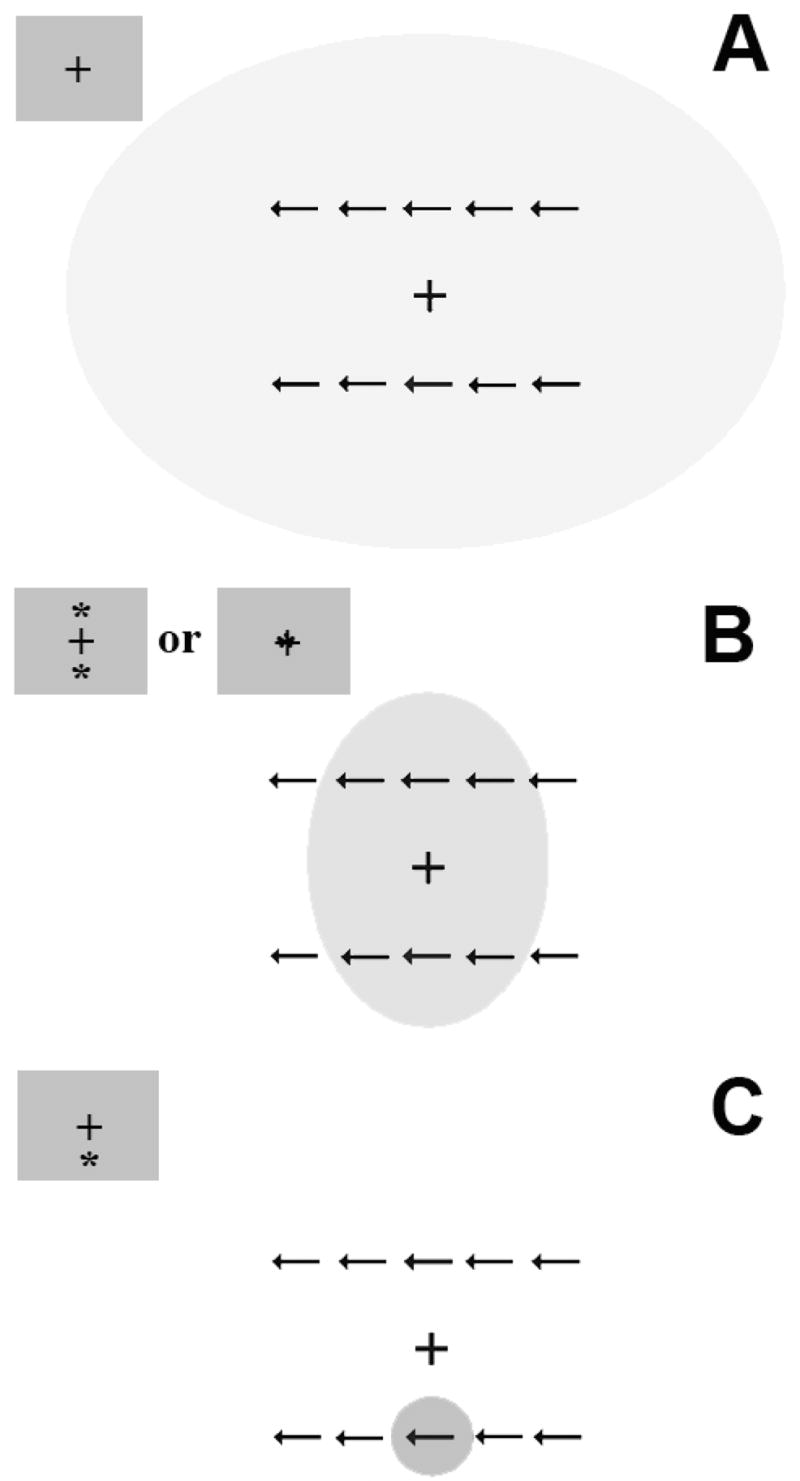
A. By default, visual attention is spread over the entire visual display. B. An alerting (double) cue serves to focus attention in on the center of the display, but the attentional spotlight still encompasses flankers that are proximal to the potential target locations. C. A valid orienting (spatial) cue further restricts the spotlight to the impending target’s spatial location. Thus, an alerting cue does not provide as much assistance to the observer when flankers are incongruent – conflict resolution between the competing responses elicited by target and flanker arrows is still required. However, an orienting cue provides a large benefit in such conditions, by focusing upon the target at the expense of the competing flanker information.
Importantly, for the ANT to measure the influence of cues and distractors on performance, the difference between RTs across conditions is typically computed. This fact is particularly critical because one of the best-documented changes induced by video game playing is that VGPs have faster RTs than NVGPs (c.f. Bialystok, 2006; Castel et al., 2005; Greenfield et al., 1994; Orosy-Fildes & Allan, 1989; Yuji, 1996; Dye, Green & Bavelier, submitted). Such differences in between-groups baselines may produce interactions with within-subjects measures such as those collected by the ANT that reflect the magnitude of the baseline RTs rather than differences in processing per se (c.f. Faust et al., 1999; Madden et al., 1992, 1996). Consider two subjects, subject A who responds very rapidly across all task conditions and subject B who makes much slower responses. In a hypothetical Posner cueing task (Posner, 1980), subject A responds to neutral cue trials in 300 ms and to valid cue trials in 200 ms, while subject B responds to neutral cue trials in 600 ms and to valid cue trials in 400 ms. A standard analysis would subtract the mean valid cue RT from the mean neutral cue RT and reach the conclusion that subject B benefited twice as much from the valid cue as did subject A (200 ms vs. 100 ms benefit). However, to state that subject A benefits less from the cue would be misleading – it is the difference in baseline RTs that has given rise to the apparent difference in benefit. This problem is well known in the gerontology literature on generalized slowing (Cerella, 1991, 1994; Salthouse, 2000, 2002), and would likely surface in any comparison of VGPs and NVGPs as ‘generalized speeding’ – VGPs have significantly faster response times compared to NVGPs (with equivalent accuracy) and thus VGPs will tend to have smaller RT differences between task conditions irrespective of the particular task at hand (Dye et al., submitted). One aim of the current study was to carefully control for any such baseline RT differences where they occurred.
As well as documenting possible additional effects of action video game play on visual attention, a final aim of the study was to assess how those effects were modulated by the age of the subjects who played the games. Based upon data from the ANT, Rueda et al. (2004) report that attentional ‘alerting’ continues to develop until the age of 10 years, whereas ‘orienting’ is stable by age 7 years. The view that orienting networks are adult-like by the early childhood is widely supported (Colombo, 2001; but see Schul et al., 2003); in contrast it has been suggested that alerting networks may continue to develop past 10 years of age, well into adolescence (Lin et al., 1999). The Rueda et al. (2004) study also reported stable ‘executive control’ by 7 years of age as measured by the flanker compatibility effect in the ANT procedure. Other authors, however, have suggested that there is an increasing ability to filter out irrelevant information between 7 and 10 years of age (Enns and Akhtar, 1989; Enns and Cameron, 1997; Goldberg et al., 2001; Ridderinkhof et al., 1997). There is some disagreement over the mechanism by which filtering improves with age. The fact that the size of the filtering effect (henceforth, flanker compatibility effect) is affected by the difficulty of the target task, the location and saliency of the flankers and the amount of attentional resources available to the subject (Green & Bavelier, 2006b; Miller, 1991; Lavie & Cox, 1997) may explain some of these discrepancies, as may baseline differences in RTs at different ages (Kail, 1991; Kail & Park, 1994).
In this study we administered a child-friendly version of the ANT (Rueda et al., 2004) to subjects ranging in age from 7 to 22 years to evaluate the relative role of age and video game expertise on attentional allocation. While using the same paradigm allows for easier comparison with data from previous studies, it is important to note that in the ANT spatial cues are always valid, the target remains visible until a response is made, and the cue-target SOA is 500 milliseconds. Such a paradigm makes it unlikely that reflexive attentional processes are being indexed by the ANT. Rather the paradigm allows one to measure how attention is focused upon a target, possible through both attentional processes and eye movements. Our main aim was to determine whether video game playing altered this allocation of attention indexed by the ANT, whether it influenced the filtering of task irrelevant stimuli, and how any such effects varied as a function of age.
No studies to date have examined the impact of video game playing upon attentional development in children. In young adults, several studies have now shown that video game playing enhances attentional resources leading to better performance on a number of attentionally demanding visual tasks (Castel et al., 2005; Feng et al., 2007; Green and Bavelier, 2003, 2006a, b, 2007). One effect of this enhancement is increased processing of task-irrelevant flankers – the argument is that on tasks where the decision to be made about a target is straightforward and leaves attentional resources to spare, those resources ‘spill over’ to other stimuli in the display (see Lavie, 1995). Here we ask whether juvenile gamers will show similar increases in attentional resources, and importantly whether they will be able to harness this extra capacity to benefit visual performance, as seen in adults. In the sample of school-aged children and adults tested (7–22 years of age), we hypothesize that increased attentional resources as a result of action video game experience will result in larger flanker compatibility effects due to a ‘spill over’ of processing, as observed in our previous work in adults.
Method
Subjects
A total of 131 subjects participated – of these, 75 were classified as NVGPs and 56 as VGPs. The frequency of video game play and the type of games played were assessed using a background questionnaire that required children to list the ten games they had played the most in the preceding 12 months, and to estimate how long they played each game in a typical session along with how many sessions they played per month. A subject was classified post-hoc as a VGP if they reported playing any action-based video game for any length of time in the 12 months prior to testing. As a result of parental constraints on video game playing, using the more strict criterion employed in previous studies would have resulted in too few juvenile VGPs for statistical analysis. All other subjects were classified as NVGPs (including those who played other types of video game in the preceding 12 months).
Socio-economical status was not collected, but all subjects were recruited from the Brighton Central School District and the University of Rochester, both in Rochester, NY. This school district has an affluent and relatively homogeneous catchment area, with 63.2% of adults having at least a bachelors degree (national average = 24.4%) according to the 2000 US Census and low number of students claiming free school lunches (9.1%; http://www.newsweek.com/id/39380). As a consequence, socioeconomic status is likely to be relatively high in our sample.
Informed consent was obtained prior to participation, including permission from parents where juvenile subjects were being tested. Children were given a $15 gift card for their participation, and adults received $8 per hour of participation. The study was approved by the IRB at the University of Rochester and by the Board of Governors at Brighton Central School District in Rochester, NY. A breakdown of the sample by age group and gender is given in Table 1, along with mean ages.
Table 1.
Subject characteristics of NVGPs and VGPs included in data analyses.
| 7–10 years |
11–13 years |
14–17 years |
18–22 years |
|||||
|---|---|---|---|---|---|---|---|---|
| NVGP | VGP | NVGP | VGP | NVGP | VGP | NVGP | VGP | |
| N | 30 | 13 | 13 | 15 | 12 | 15 | 14 | 13 |
| Mean (SD) | 8;11 | 9;5 | 12;1 | 12;9 | 15;10 | 14;11 | 20;4 | 19;9 |
| Age | (1;2) | (1;1) | (0;11) | (0;7) | (1;1) | (0;8) | (1;4) | (1;3) |
| # Males | 11 | 9 | 4 | 12 | 0 | 14 | 13 | 7 |
Design
All subjects were administered the child-friendly version of the ANT (see Rueda et al., 2004), in which subjects are required to make a speeded decision to indicate the direction of a central target (a fish) with a key press The experiment included two between-subjects factors (age group – 7–10 yrs, 11–13 yrs, 14–17 yrs, 18–22 yrs; and video game experience – NVGP, VGP) and two within-subjects factors (flanker type – incongruent, congruent, absent; and cue type – absent, center, double, spatial).
Flankers were two fish presented horizontally aligned on either side of a central target fish (see Figure 2). Flankers could either be incongruent with the target (fish pointing in the opposite direction), congruent with the target (fish pointing in the same direction) or absent (target presented in isolation). Each fish subtended 5.4 degree of visual angle, with the edges of adjacent fish separated by 0.4 degrees of visual angle. The fish were presented 1.7 degrees of visual angle above or below a central fixation point. The fish appeared above or below the fixation point with equal probability.
Figure 2.

Children’s version of the Attentional Network Test.
The cue consisted of one or two asterisks presently briefly prior to the onset of the arrow(s). The cue was either absent, central (presented at the fixation point), double (two cues presented simultaneously above and below the fixation point at both possible target locations) or spatial (a single cue presented above or below the fixation point and indicating the location of the subsequent target).
There were a total of 48 experimental trials for each subject in each block, determined by the combinations of flanker (3), cue type (4), target location (2) and target direction (2). Each subject participated in three blocks of experimental trials, with the first block preceded by 24 practice trials, resulting in a total of 168 trials overall.
The total duration of each trial was set to 4000 ms. A pre-stimulus fixation point appeared for a variable duration of 400–1600 ms. This was then accompanied by a cue presented for 100 ms. After the offset of the cue, there was a 400 ms interval prior to the onset of the fish. The fixation point was present at all times. Following the subject’s response, the fish were removed from the display, leaving only the post-stimulus fixation point. The next trial was initiated after 3500 ms minus the duration of the pre-stimulus fixation point and minus the reaction time of the subject (total duration = pre-stimulus fixation point + 100 + 400 + reaction time + 3500 − pre-stimulus fixation point − reaction time).
Apparatus
Stimuli were presented on a 23-inch LCD display (Apple Computer, Inc.) with a 1024×5768 pixel resolution and a 60 Hz frame rate. A Java script was used to run the experiment (available from http://www.sacklerinstitute.org/users/jin.fan/) under Mac OS X on a PowerBook G4 laptop computer (Apple Computer, Inc.). As a result the experimental details closely mirror those reported in Rueda et al. (2004).
Procedure
Children were tested in a dimly-lighted room in their homes in the Rochester, NY area and adults were tested in a laboratory at the University of Rochester. The experimental environment and setup was the same for both VGPs and NVGPs. They were seated 40 cm from the centre of the LCD display, and instructed to maintain fixation on the central fixation point (a crosshair) at all times. The subjects were instructed to respond to the target fish by pressing a key congruent with the direction of the fish as quickly and as accurately as possible. The practice block took approximately 3 minutes, and each experimental block approximately 4 minutes, for a total duration of 15 minutes.
Results
Reaction times from incorrect trials were excluded from analyses. Following this, on a subject-by-subject basis, a mean RT was calculated for each of the twelve conditions (3 flanker types by 4 cue types). Data were collapsed across target direction (left/right) and target location (above/below). On the basis of unusually slow and error-prone responses (more than two standard deviations beyond the mean for their age group and gaming status), data from six NVGPs were excluded from analysis completely. For the remaining subjects, if the response time for a trial was greater then two standard deviations from the mean for its condition, then it was excluded as an outlier; neither RT nor accuracy data were analyzed for these outlier trials. Median RTs were then calculated for each condition for each subject and submitted for further analysis.
Gender Differences in RT
Males are more likely than girls to play action video games, and this is reflected in an asymmetric distribution of males and females across the NVGP and VGP categories (Table 1). Mezzacappa (2004) reported a small gender effect on RTs in the ANT in a study of 118 young girls and 131 young boys. An initial analysis of the overall RTs for males and females in our NVGP group revealed no significant difference in RTs (F(1, 68)=1.45, p > .05, eta2p=.02), suggesting that gender had no measurable impact on RT performance in the age range tested.
Controlling for Baseline Differences in RT
A four-way mixed ANOVA was performed on the median RT data with flanker type (incongruent, congruent) and cue type (absent, center, double, spatial) as within subjects factors, and age group (7–10 yrs, 11–13 yrs, 14–17 yrs, 18–22 yrs) and video game playing (NVGP, VGP) as between subjects factors. This analysis revealed significant main effects of age group (F (3, 117)=50.07, p<.001, eta2p=.56) and video game playing (F (1, 117)=8.68, p=.004, eta2p=.07), suggesting baseline differences in RT as a function of both age and gaming experience.
Before proceeding with any further analyses, these main effects of age group and video game playing were addressed. The median RTs for older subjects were faster than those for younger subjects (M7–10YRS = 678msec, M11–13YRS = 554msec, M14–17YRS = 496msec, M18–22YRS = 467msec). In addition, VGPs (MVGP = 525 msec) had faster median RTs than NVGPs (MNVGP = 597 msec). As outlined previously, these baseline differences are of concern when interpreting interactions (see Faust et al., 1999; Madden et al., 1992, 1996 for more discussion). The next stage of the analysis sought to address these baseline differences before reanalyzing the data.
In order to address baseline differences as a function of age group, the average median RT (collapsed across all target-only conditions, i.e. using only those trials where no flankers accompanied the target) was computed for each NVGP subject, and this was plotted against their age in months. Following Cerella and Hale (1994), these data were fitted using an exponential decay function (see Equation 1 and Figure 3A):
| Equation 1 |
Figure 3.

(A) Using the function outlined by Cerella and Hale (1994), RT was fitted as a function of NVGP subject age using an exponential decay function. (B) After controlling for age differences in RT, normalized RTs from the flanker absent conditions were used to fit NVGP group RTs against RTs obtained from the VGP group. The linear fit from the resulting linear regression was used to control for baseline differences in RT between gamers and non-gamers.
A goodness-of-fit metric for the non-linear function, analogous to R2, was computed using the method provided by Haessel (1978). This revealed a good fit to the data: Cos2φ= 0.646. On the basis of this function, a predicted RT score was computed for all NVGP and VGP subjects and used to normalize their median RTs for each condition. For example, if a subject had an age-predicted RT of 450msec and their performance within a condition was 400msec, then their transformed RT would be 400/450 or 0.89. This age-normalized RT (RTage) was used for all further analyses.
To control for RT differences resulting from video game experience – a categorical variable – another procedure was employed; the RTage for each of the four target-only conditions were computed for the NVGP and VGP groups. These were plotted against each other, and a linear fit obtained (see Equation 2 and Figure 3B):
| Equation 2 |
This linear function was used to transform the RTage for NVGPs in each of the other eight experimental conditions formed by crossing flanker type (incongruent, congruent) with cue type (absent, center, double, spatial). The resulting gamer-transformed age-normalized RTs (henceforth, transformed normalized RTs – RTnormed) represent the extent to which RTs deviate from what is expected given the age and video gaming experience of individual subjects and thus provide a measure of the effects of flanker congruency and cue type that is not biased by baseline differences in speed of response.
Alerting, Orienting and Flanker Compatibility Effects
In line with previous studies using the ANT, we calculated ‘attentional network’ scores to reflect the effects of alerting, orienting and flanker compatibility. These were computed using these RTnormed values, and entered into two-way ANOVAs with age group (7–10 yrs, 11–13 yrs, 14–17 yrs, 18–22 yrs) and video game playing (NVGP, VGP) as between subjects factors.
Alerting effects – measuring the efficiency with which a temporal cue enhances processing of the target – were computed by subtracting RTnormed values for the double cue conditions from those for the no cue conditions for each subject. The main effect of age group was statistically significant (F (3, 117)=2.68, p=.05, eta2p=.06) with younger children exhibiting larger alerting effects than older children and adults (M7–10YRS=0.076, M11–13YRS=0.068, M14–17YRS=0.044, M18–22YRS=0.053). A priori contrasts revealed significant differences between the alerting scores of 7–10 year olds and 11–22 year olds (p=.027). The main effect of video gaming playing (eta2p=.02) and the interaction between age group and video game playing (eta2p=.02) did not approach statistical significance (see Figure 4A).
Figure 4.

A. Alerting effects were computed by contrasting the no cue and double cue conditions. A significant effect of age was observed, with 7–10 year olds having larger alerting effects than older children and adults. These larger alerting effects possibly reflect higher levels of inattention in young children that are alleviated by presenting a temporally informative cue. The effect of video game playing was not statistically significant. B. In contrast, orienting effects – computed by comparing center cue and spatial cue conditions – did not vary as a function of age. However, there was a main effect of video game playing with VGPs having larger orienting effects than NVGPs, suggesting that action video game players may be better able to use a spatial cue to orient their attention to a target.
Orienting scores – measuring the efficiency with which a valid spatial cue enhances processing of the target – were computed by subtracting RTnormed values for the spatial cue conditions from those for the center cue conditions for each subject. The ANOVA revealed no significant main effect of age group (F (3, 177)=0.17, p=.914, eta2p<.01) nor a significant interaction between age group and video game playing (eta2p=.03). However, the analysis revealed a significant main effect of video game playing on orienting effects (F (1, 117)=10.20, p=.002, eta2p=.08), with VGPs (MVGP=0.060) exhibiting larger orienting effects than NVGPs (MNVGP=0.038; see Figure 4B). This effect will be returned to in the ANOVA analysis reported below.
Finally, flanker compatibility effects – measuring the extent to which flankers interfere with processing of the target – were computed by subtracting RTnormed values for the congruent flanker conditions from those for the incongruent flanker conditions for each subject. The ANOVA revealed no significant age group effect (F (3, 117)=1.08, p=.361, eta2p=.03) and no interaction between age group and video game playing (eta2p=.02). There was, however, a significant main effect of video game playing (F(1, 117)=19.71, p<.001, eta2p=.14), with VGPs (MVGP=0.103) having larger flanker compatibility effects, or in other words experiencing more interference from flankers, than NVGPs (MNVGP=0.070).
The data failed to reveal a significant interaction between age group and the size of flanker compatibility effects. Although the data reported in Figure 5 suggest that such an interaction may be present – with 7–13 year old gamers having disproportionately larger flanker compatibility effects than their non-gaming peers – the effect appears to be driven by large flanker compatibility effects for 7–10 year old gamers and small flanker compatibility effects for 11–13 year old non-gamers. Therefore the data are inconclusive with respect to action video gaming having greater effects for younger gamers.
Figure 5.

Incongruent flankers slowed down the responses of video game players more than it did those of non-video game players, given the RTs expected as a function of both age and video game experience. Higher flanker compatibility effects provide an index of the extent to which task-irrelevant flankers were processed.
The omnibus ANOVA below further addresses how changes in orienting and flanker compatibility effects may be best understood in terms of attentional allocation, by looking at changes to performance in which of the experimental conditions lead to the observed differences.
Omnibus RTnormed ANOVA
The omnibus ANOVA was repeated using the RTnormed that were used to calculate the attention effects. Importantly, the main effects of age group (F (3, 117) = 1.27, p = .289, eta2p = .03) and videogame playing (F (1, 117) = 1.19, p = .277, eta2p = .01) did not approach significance, nor did they interact significantly (F (3, 117) = 0.34, p = .771, eta2p = .01). With the applied corrections achieving their aims – there were no statistically significant baseline differences in RT between groups – the analysis also revealed, as expected, significant main effects of flanker type, due to slower RTnormed values in the presence of incongruent flankers (F (1, 117) = 484.88, p < .001, eta2p = .81) and of cue type (F (3, 351) = 164.71, p < .001, eta2p = .59). Two-way interactions between flanker type and cue type (F (3, 351) = 12.69, p < .001, eta2p = .10; Figure 6), flanker type and video game playing (F (1, 117) = 19.71, p < .001, eta2p = .14; Figure 7A) and cue type and video game playing (F (3, 351) = 4.96, p = .002, eta2p = .04; Figure 7B) were statistically significant. There was also a statistically significant three-way interaction between flanker type, cue type and video game playing (F (3, 351)=2.76, p=.042, eta2p=.02).
Figure 6.

The effects of incongruent flankers were more pronounced – relative to those of congruent flankers – following center or double cues that provided only temporal information about target onset.
Figure 7.

(A) Incongruent flankers slowed down the responses of VGPs more than it did those of NVGPs, given the RTs expected as a function of both age and video game experience; (B) there were differential effects of cue as a function of video game experience, with VGPs responding comparatively more slowly than NVGPs in the absence of cues providing valid spatial information (again, given the RTs expected as a function of both age and video game experience).
Omnibus Error Analysis
A four-way mixed ANOVA was performed on the error data with flanker type (incongruent, congruent) and cue type (absent, center, double, spatial) as within subjects factors, and age group (7–10 yrs, 11–13 yrs, 14–17 yrs, 18–22 yrs) and video game playing (NVGP, VGP) as between subjects factors. This analysis revealed significant main effects of flanker type (F (1, 117)=103.67, p<.001, eta2p=.47) and age group (F (3, 117)=8.46, p<.001, eta2p=.178). These were qualified by a significant two-way interaction between flanker type and age group (F (3, 117)=5.06, p=.002, eta2p=.115). For incongruent flanker trials, younger subjects made more errors than older subjects (M7–10YRS=7.7%, M11–13YRS=6.0%, M14–17YRS=4.6%, M18–22YRS=2.4%). Error rates were equivalent for congruent flanker trials across the ages tested (M7–10YRS=1.8%, M11–13YRS=1.1%, M14–17YRS=1.3%, M18–22YRS=0.8%), reflecting the small impact of congruent flankers observed in RT measures. Importantly, the main effect of video game playing on error rate was not statistically significant (F (1, 117)=2.74, p=.102, eta2p=.02; MNVGP=2.84%, MVGP=2.91%), nor did it interact with any other factor (all Fs < 1).
Discussion
Effects of action video game experience on attention skills
Analysis of raw RT data revealed that VGPs responded more quickly than NVGPs, but did not make more errors. Speeded processing of visual information without a concomitant decrease in accuracy has now been reported by several groups in studies with adult subjects (e.g. Bialystok, 2006; Castel et al., 2005; Clark et al., 1987; Dye, Green & Bavelier, submitted), and here that finding is extended to children as young as 7 years of age. It suggests that across a wide range of development VGPs are not more likely to make speed-accuracy trade-offs, but are faster to respond accurately. It has been suggested that group baseline differences in RT in mixed designs can lead to between and within subjects measures interacting, with the interaction reflecting the baseline difference and not a group difference in processing on the within subjects measure per se (Faust et al., 1999; Madden et al., 1992, 1996). Following Madden et al., we transformed RT data to remove the difference in baseline across groups. After doing so, and controlling for the effect of age on RT (see above), we found a significant interaction between whether or not subjects played action video games and both flanker congruency and the type of cue that preceded the target-flanker display.
The effect of video gaming on utilizing an alerting cue was not statistically significant, offering no evidence for differences in alerting efficiency between NVGPs and VGPs. Effects of video game playing were found on the orienting cue measure and the flanker compatibility measure, with VGPs exhibiting greater benefit from an orienting cue and greater interference from flankers. Although it is tempting to conclude that VGPs (i) lack the ability to spatially focus their attention and (ii) have deficiencies in selecting task-relevant information, we argue below that these effects are best understood in terms of enhanced attentional resources in VGPs.
The effect of videogame playing on the orienting effect is reflected in the interaction between cue type and game playing experience. VGPs respond comparatively more slowly than NVGPs unless a spatial cue is provided, in which case the two groups do not differ (Figure 5A). It should first be definitively stressed that this does not mean that the VGPs took more time to make their responses. In fact, in the raw (un-normed) RT data it is the case the VGPs responded more quickly across all experimental conditions. Instead, what this means is that the VGPs took more time than expected considering the RT advantages conferred by video game playing. Second, the comparatively greater time taken by VGPs applies to the baseline condition used to compute the orienting effect, but not to the spatial cue condition. Thus, a spatially informative cue focuses attention equally well in VGPs and NVGPs, supporting the view that there is little difference between NVGPs and VGPs in terms of how they use a valid spatial cue to allocate their attention. This highlights the importance of examining RTs by condition in the ANT paradigm, and not simply relying upon difference scores, in order to accurately interpret effects.
The effect of video game playing on flanker compatibility effect sizes is reflected in the interaction between flanker type and gaming experience. Again, it is not the case that VGPs were slower than NVGPs when incongruent flankers were presented; rather, they responded comparatively more slowly to incongruent as compared to congruent flankers given what would be expected considering the reaction time advantages conferred by video game playing. This result is consistent with the proposal of greater attentional resources in VGPs, allowing them to (unavoidably) devote more processing resources to flankers and thus to exhibit comparatively greater flanker effects.
Thus we argue that the orienting and flanker compatibility effect differences noted between VGPs and NVGPs are best understood in terms of changes in the spread of attention over the visual scene. The notion of “spread of attention” has been used in previous studies to explain the interaction between cue type and flanker type. The spatial cue, by focusing attention tightly over the target area, diminishes the extent to which flankers are processed. This limits the impact of flankers on target processing, and thus the size of the flanker compatibility effect. On the other hand, by enhancing attention but only loosely restricting it in space, the center and double cues result in efficient processing of both target and flankers, leading to greater flanker compatibility effects and a cue by flanker interaction (see Callejas et al., 2004; Dye et al., 2007; Fernandez-Duque & Posner, 1997). A similar account can also explain the double and triple interactions of gaming experience with flanker type and cue type. Greater spread of attentional resources in gamers will result in greater processing of the flankers and thus greater interference from incongruent flankers (accounting for the gaming experience by flanker type interaction), except when a spatial cue focuses attention over the target area (accounting for both the gaming experience by cue type interaction and the triple interaction). The spatial cue focuses attention tightly on the location of target, resulting in easier flanker exclusion, reducing response conflict relative to conditions in which center and double cues are present (see Dye et al., 2007). Thus, the presence of the spatial cue works against the spill-over of attentional resources to the flankers and diminishes the gaming experience by flanker type interaction (see Figure 8).
Figure 8.

The cues serve to focus attention more narrowly upon the target arrow. The ovals represent the attentional window resulting from the preceding cue (c.f. Figure 2). Enhanced attentional resources in VGPs result in a larger spill over of attention to distractors flanking the target, and thus greater flanker compatibility effects. We propose that the spatial cue serves to focus attention so tightly on the target arrow that the spill over of attention is attenuated, resulting in comparable flanker compatibility effects for NVGPs and VGPs following spatial cues, despite larger flanker compatibility yeffects following non-spatial (alerting) cues that do not convey information about target location.
We note that the ANT flanker compatibility score cannot unambiguously resolve whether a greater score is in fact due to enhanced attentional resources or due to poor attentional selection. This score therefore needs to be considered in terms of RTs in each of the conditions used to compute that measure of flanker compaitibility. In this respect, the findings that VGPs respond faster in incompatible flanker conditions than NVGPs, and are no less accurate, support the view that VGPs perform better even in the incompatible condition. In addition to providing the most parsimonious account for the data, this explanation is in line with previous work indicating greater attentional resources in action game players (Dye & Bavelier, 2004; Green & Bavelier, 2003, 2006a, b).
Effects of age on visual attention skills
Children made accurate responses more rapidly as they got older, in accord with many studies that have demonstrated increases in speed of information processing through childhood (Kail, 1991; Kail and Park, 1994). We were concerned about these baseline differences in RT across age groups when examining age group by within subjects factor interactions. As has been discussed extensively elsewhere (Faust et al., 1999; Madden et al., 1992, 1996), these baseline differences can produce ‘spurious’ interactions, where group differences on the within subjects factor can be driven by a global speed of processing function rather than the experimental factor of interest. Following Cerella and Hale (1994), we modeled the effect of age on RTs using an exponential decay function. We showed that this correction adequately removed baseline RT differences across groups.
Computing attentional effects, our study revealed a small effect of age group on the alerting effect – 7–10 year olds obtained more benefit from a temporal cue than did older children. This confirms the results of Rueda et al. (2004) who found improvements in alerting up until 10 years of age. Rueda et al. suggested possible greater inattention in younger children during inter-trial intervals, a conclusion consistent with data from the current study. An additional contribution of our study concerns changes in flanker compatibility effects over the course of development. Although flanker compatibility effects as measured by RT did not vary with age, error analyses revealed that as subjects got older they made fewer incorrect responses to targets flanked by incongruent distractors, suggesting an increasing ability with age to filter out distractors. Thus there is evidence that the presence of incongruent flankers did not slow down the responses of the youngest children – in accordance with Rueda et al. – but rather made them more prone to committing errors. This latter finding indicates that the filtering network keeps maturing at least until 10 years of age, in agreement with the conclusions of Enns and colleagues (Enns & Akhtar, 1989;Enns & Cameron, 1997). Flanker compatibility effects as a function of action gaming experience were statistically equivalent across the age ranges tested. While differences between VGPs and NVGPs appeared larger for 7–13 year old children than for 14–22 year olds, the three-way interaction was not statistically significant.
This work further documents the enhanced attentional resources of action video gamers and establishes faster reaction times in that population without a notable loss in accuracy. These effects were seen throughout the age range studied suggesting similar effects of action game playing from the early school years through to adulthood. While causality can only be inferred with a training study, the findings are in accord with attentional changes that have been previously trained in NVGPs using action video games (Feng et al., 2007; Green & Bavelier, 2003, 2006b).
Finally, this work calls for caution when interpreting ANT scores related to alerting, orienting and executive control scores. First, differences in baselines across the groups compared need to be taken into consideration. Otherwise, one may attribute processing differences to the populations compared, when in fact they reflect a generalized baseline performance difference rather than a specific processing difference. Second, greater or lower scores on the ANT may not always readily associated with better or worse attentional control. Rather, the pattern of interactions appear crucial in determining how cues alter attention allocation and thus the efficiency with which targets and distractors will be processed.
Acknowledgments
We are grateful to the parents and children in Rochester NY who gave their time to facilitate the developmental aspects of the work reported here. The research was made possible by a grant from the John F. Merck Foundation to DB.
Appendix 1: Omnibus ANOVAs Using Raw RTs
Flanker Type F (1, 117) = 363.30, p < .001, pe2 = .76
Flanker Type * Age Group F (3, 117) = 5.75, p = .001, pe2 = .13
Flanker Type * Gaming F (1, 117) = 1.01, p = .316, pe2 = .01
Flanker Type * Age Group * Gaming F (3, 117) = 0.69, p = .559, pe2 = .02
Cue Type F (3, 351) = 124.69, p < .001, pe2 = .52
Cue Type * Age Group F (9, 351) = 3.95, p < .001, pe2 = .09
Cue Type * Gaming F (3, 351) = 0.52, p = .668, pe2 < .01
Cue Type * Age Group * Gaming F (9, 351) = 0.81, p = .611, p = .02
Flanker Type * Cue Type F (3, 351) = 9.16, p < .001, pe2 = .07
Flanker Type * Cue Type * Age Group F (9, 351) = 0.41, p = .931, pe2 = .01
Flanker Type * Cue Type * Gaming F (3, 351) = 1.40, p = .242, pe2 = .01
Flanker Type * Cue Type * Age Group * Gaming F (9, 351) = 0.31, p = .973, pe2 = .01
Age Group F (3, 117) = 50.07, p < .001, pe2 = .56
Gaming F (1, 117) = 8.68, p = .004, pe 2 = .07
Age Group * Gaming F (3, 117) = 1.09. p = .355, pe2 = .03
Appendix 2: Video Game Questionnaire
We are interested in how often you play video games, and what type of games you play. We will use this information to examine the effects of video game playing on the development of visual attention skills.
We want you to think of the 6 video games you played the most in the last year. For each game, please write in the name of the game, the number of hours you play the game in a typical session, and the number of times you play the game in a typical month. Also please indicate the console used. We are interested only in the games you have played in the last year.
Example 1. If you played Solitaire on your PC twice a week for about 30 minutes at a time, then you would write in Solitaire (name of game), 1/2 (hours per session), 8 (2 ×4 = 8 sessions per month) and PC (console).
| Name of game | Hours per session | Sessions per month | Console | |
|---|---|---|---|---|
| Ex. 1 | Solitaire | 1/2 | 8 | PC |
| Ex. 2 | Final Fantasy XI | 3 | 1 | PS2 |
| 1 | ||||
| 2 | ||||
| 3 | ||||
| 4 | ||||
| 5 | ||||
| 6 |
Example 2. If you played Final Fantasy XI on your Play Station 2 once a month, typically for 3 hours at a time, then you would write in Final Fantasy (name of game), 3 (hours per session), 1 (sessions per month) and PS2 (console).
Appendix 3: Video Game Classifications
Games Categorized As Action Video Games
007 – Everything or Nothing; 007 – Golden Eye; 007 – Goldfinger; 007 – Nightfire; Battlefield 1942; Bionicle; Counterstrike; Devil May Cry; Ghost Recon; Grand Theft Auto; Grand Theft Auto – San Andreas; Grand Theft Auto – Vice City; Half Life 2; Halo; Halo 2; Hitman 2; Medal of Honor; Metal Gear Solid; Metal Gear Solid 2; Quake III; Rainbow Six-Three; Splinter Cell; Star Wars – Jedi Starfighter; Unreal Tournament; Viet Cong.
Games Categorized As Non-Action Video Games
Age of Empires; Age of Mythology; Angelica; ATV Crossroad Fury; Backyard Baseball; Backyard Soccer; Barbie; Brute Force; Bubble Trouble; Burning Monkey Mahjong; Civilization III; Crash Bandicoot; Cross Country USA; Dance Dance Revolution; Dave Mirra Freestyle BMX; DBZ; Deimos Rising; Donkey Kong; Downhill Domination; Dr. Mario; Dr. Muto; Dracula; Duck Hunt; Emperor’s New Groove; Empires – Dawn of the Modern World; ESPN NBA Basketball 2004; Extreme Ghostbusters; FIFA 2001; Final Fantasy VII; Formula One 2001; Free Cell; Frogger; Gauntlet – Dark Legacy; Harry Potter and the Chamber of Secrets; Harry Potter and the Sorceror’s Stone; High Heat 2003; Home Run King; Illusion of Gaia; Jedi Knights; Karaoke Revolution; King’s Quest VII; Kingdom Hearts; Lego Island 2; Lego Racers; Lord of the Rings – Return of the King; Lord of the Rings – The Two Towers; Mad Dash; Mario Kart; Mario Kart Double Dash; Mario Party 3; Mario Super Party; Master Matt’s Kung Fu Rama; Masters of Orion III; Mickey Mouse Kitchen; Mine Sweeper; MLB Slugfest; MVP Baseball 2004; MX Unleashed; Myst; Mystery of the Monkey Kingdom; NBA Live 2004; Need 4 Speed – Porsche Unleashed; Need for Speed – Underground; Neo Pets; NFL Blitz; NFL Fever 2002; NFL Madden 2004; NHL 1999; NHL 2004l; Pokemon; Postopia; Quad – Desert Fury; Railroad Tycoon; Scooby Doo – Night of 100 Frights; Shrek; Sim City; Skateboarder Tycoon; Sly Cooper; Snood; Solitaire; Soul Caliber II; Spiderman; Spy Fox – Operation Ozone; SSX 3; SSX Tricky 2; Star Craft; Star Fox; Star Wars – Knights of the Old Republic; Super Mario Land; Super Mario RPG; Supersmash Brothers Melee; Tetris; Text Twist; The Sims; The Sims –Bustin’ Out; Tiger Woods Golf; Tony Hawke Pro Skater 4; Tony Hawke’s Underground; Treasure Mountain; Vieautiful Joe; Virtual Pinball; Where In The World Is Carmen Sandiego?; World of Warcraft; World Tour Soccer 3; Yoshi’s Island; Zelda – Ocarina of Time; Zelda –Wind Waker; Zoo Tycoon; Zoombinis.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Badre D, Wagner AD. Selection, integration, and conflict monitoring; assessing the nature and generality of prefrontal cognitive control mechanisms. Neuron. 2004;41(3):473–487. doi: 10.1016/s0896-6273(03)00851-1. [DOI] [PubMed] [Google Scholar]
- Bialystok E. Effect of bilingualism and computer video game experience on the Simon task. Canadian Journal of Experimental Psychology. 2006;60(1):68–79. doi: 10.1037/cjep2006008. [DOI] [PubMed] [Google Scholar]
- Callejas A, Lupianez J, Tudela P. The three attentional networks: On their independence and interactions. Brain & Cognition. 2004;54(3):225–227. doi: 10.1016/j.bandc.2004.02.012. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Thomas KM, Welsh TF, Badgaiyan RD, Eccard CH, Jennings JR, et al. Dissociation of response conflict, attentional selection, and expectancy with functional magnetic resonance imaging. Proceedings of the National Academy of Sciences USA. 2000;97(15):8728–8733. doi: 10.1073/pnas.97.15.8728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castel AD, Pratt J, Drummond E. The effects of action video game experience on the time course of inhibition of return and the efficiency of visual search. Acta Psychologica. 2005;119(2):217–230. doi: 10.1016/j.actpsy.2005.02.004. [DOI] [PubMed] [Google Scholar]
- Cerella J. Age effects may be global, not local: Comment on Fisk and Rogers (1991) Journal of Experimental Psychology: General. 1991;120(2):215–223. doi: 10.1037/0096-3445.120.2.215. [DOI] [PubMed] [Google Scholar]
- Cerella J. Generalized slowing in Brinley plots. Journal of Gerontology. 1994;49(2):65–71. doi: 10.1093/geronj/49.2.p65. [DOI] [PubMed] [Google Scholar]
- Cerella J, Hale S. The rise and fall in information-processing rates over the life span. Acta Psychologica. 1994;86(2–3):109–197. doi: 10.1016/0001-6918(94)90002-7. [DOI] [PubMed] [Google Scholar]
- Clark JE, Lanphear AK, Riddick CC. The effects of video game playing on the response selection processing of elderly adults. Journal of Gerontology. 1987;42(1):82–85. doi: 10.1093/geronj/42.1.82. [DOI] [PubMed] [Google Scholar]
- Colombo J. The development of visual attention in infancy. Annual Review of Psychology. 2001;52:337–367. doi: 10.1146/annurev.psych.52.1.337. [DOI] [PubMed] [Google Scholar]
- Corbetta M, Shulman GL. Human cortical mechanisms of visual attention during orienting and search. Philosophical Transactions of the Royal Society of London B: Biological Sciences. 1998;353(1373):1353–1362. doi: 10.1098/rstb.1998.0289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coull JT, Frith CD, Frackowiak RS, Grasby PM. A fronto-parietal network for rapid visual information processing: A PET study of sustained attention and working memory. Neuropsychologia. 1996;34(11):1085–1095. doi: 10.1016/0028-3932(96)00029-2. [DOI] [PubMed] [Google Scholar]
- Diamond A, Goldman-Rakic PS. Comparison of human infants and rhesus monkeys on Piaget’ AB task: Evidence for dependence on dorsolateral prefrontal cortex. Experimental Brain Research. 1989;74(1):24–40. doi: 10.1007/BF00248277. [DOI] [PubMed] [Google Scholar]
- Dye MWG, Bavelier D. Playing video games enhances visual attention in children [Abstract] Journal of Vision. 2004;4(11):40a. [Google Scholar]
- Dye MWG, Baril DE, Bavelier D. Which aspects of visual attention are changed by deafness? The case of the Attentional Network Test. Neuropsychologia. 2007;45:1801–1811. doi: 10.1016/j.neuropsychologia.2006.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dye MWG, Green CS, Bavelier D. Faster reaction times across domains: Transfer of learning with action videogames. Current Directions in Psychological Science (submitted) [Google Scholar]
- Egner T, Hirsch J. Cognitive control mechanisms resolve conflict through cortical amplification of task-relevant information. Nature Neuroscience. 2005;8(12):1784–1790. doi: 10.1038/nn1594. [DOI] [PubMed] [Google Scholar]
- Enns JT, Akhtar N. A developmental study of filtering in visual attention. Child Development. 1989;60(5):1188–1199. [PubMed] [Google Scholar]
- Enns JT, Cameron S. Selective attention in young children: The relations between visual search, filtering, and priming. Journal of Experimental Child Psychology. 1987;44(1):38–63. doi: 10.1016/0022-0965(87)90021-x. [DOI] [PubMed] [Google Scholar]
- Fan J, McCandliss BD, Sommer T, Raz A, Posner MI. Testing the efficiency and independence of attentional networks. Journal of Cognitive Neuroscience. 2002;14(3):340–347. doi: 10.1162/089892902317361886. [DOI] [PubMed] [Google Scholar]
- Faust ME, Balota DA, Spieler DH, Ferraro FR. Individual differences in information-processing rate and amount: Implications for group differences in response latency. Psychological Bulletin. 1999;125(6):777–799. doi: 10.1037/0033-2909.125.6.777. [DOI] [PubMed] [Google Scholar]
- Feng J, Spence I, Pratt J. Playing an action video game reduces gender differences in spatial cognition. Psychological Science. 2007;18(10):850–855. doi: 10.1111/j.1467-9280.2007.01990.x. [DOI] [PubMed] [Google Scholar]
- Fernandes-Duque D, Posner M. Relating the mechanisms of orienting and alerting. Neuropsychologia. 1997;35(4):477–486. doi: 10.1016/s0028-3932(96)00103-0. [DOI] [PubMed] [Google Scholar]
- Goldberg MC, Maurer D, Lewis TL. Developmental changes in attention: The effects of endogenous cueing and of distractors. Developmental Science. 2001;4(2):209–219. [Google Scholar]
- Green CS, Bavelier D. Action-video-game experience alters the spatial resolution of vision. Psychological Science. 2007;18(1):88–94. doi: 10.1111/j.1467-9280.2007.01853.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Enumeration versus multiple object tracking: The case of action video game players. Cognition. 2006a;101(1):217–245. doi: 10.1016/j.cognition.2005.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Effect of action video games on the spatial distribution of visuospatial attention. Journal of Experimental Psychology: Human Perception and Performance. 2006b;32(6):1465–1468. doi: 10.1037/0096-1523.32.6.1465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Action video game modifies visual selective attention. Nature. 2003;423(6939):534–537. doi: 10.1038/nature01647. [DOI] [PubMed] [Google Scholar]
- Greenfield PM, deWinstanley P, Kilpatrick H, Kaye D. Action video games and informal education: Effects on strategies for dividing visual attention. Journal of Applied Developmental Psychology. 1994;15:105–123. [Google Scholar]
- Haessel W. Measuring goodness of fit in linear and nonlinear models. Southern Economic Journal. 1978;44(3):648–652. [Google Scholar]
- Kail R. Developmental change in speed of processing during childhood and adolescence. Psychological Bulletin. 1991;109:490–501. doi: 10.1037/0033-2909.109.3.490. [DOI] [PubMed] [Google Scholar]
- Kail R, Park Y. Processing time, articulation time, and memory span. Journal of Experimental Child Psychology. 1994;57:281–291. doi: 10.1006/jecp.1994.1013. [DOI] [PubMed] [Google Scholar]
- Lavie N. Perceptual load as a necessary condition for selective attention. Journal of Experimental Psychology: Human Perception & Performance. 1995;21(3):451–468. doi: 10.1037//0096-1523.21.3.451. [DOI] [PubMed] [Google Scholar]
- Lavie N, Cox S. On the efficiency of visual selective attention: Efficient visual search leads to inefficient distractor rejection. Psychological Science. 1997;8:395–398. [Google Scholar]
- Lin CC, Hsiao CK, Chen WJ. Development of sustained attention assessed using the continuous performance test among children 6–15 years of age. Journal of Abnormal Child Psychology. 1999;27(5):403–412. doi: 10.1023/a:1021932119311. [DOI] [PubMed] [Google Scholar]
- Madden DJ, Pierce TW, Allen PA. Adult age differences in attentional allocation during memory search. Psychology & Aging. 1992;7(4):594–601. doi: 10.1037//0882-7974.7.4.594. [DOI] [PubMed] [Google Scholar]
- Madden DJ, Pierce TW, Allen PA. Adult age differences in the use of distractor homogeneity during visual search. Psychology & Aging. 1996;11(3):454–474. doi: 10.1037//0882-7974.11.3.454. [DOI] [PubMed] [Google Scholar]
- Mezzacappa E. Alerting, orienting, and executive attention: developmental properties and sociodemographic correlates in an epidemiological sample of young, urban children. Child Development. 2004;75(5):1373–1386. doi: 10.1111/j.1467-8624.2004.00746.x. [DOI] [PubMed] [Google Scholar]
- Miller J. The flanker compatibility effect as a function of visual angle, attentional focus, visual transients and perceptual load: A search for boundary conditions. Perception & Psychophysics. 1991;49:270–288. doi: 10.3758/bf03214311. [DOI] [PubMed] [Google Scholar]
- Nelson JK, Reuter-Lorenz PA, Sylvester CY, Jonides J, Smith EE. Dissociable neural mechanisms underlying response-based and familiarity-based conflict in working memory. Proceedings of the National Academy of Sciences USA. 2003;100(19):11171–11175. doi: 10.1073/pnas.1334125100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orosy-Fildes C, Allan RW. Psychology of computer use: XII. Video game play: Human reaction time to visual stimuli. Perceptual & Motor Skills. 1989;69:243–247. [Google Scholar]
- Posner M. Orienting of attention. The Quarterly Journal of Experimental Psychology. 1980;32:3–25. doi: 10.1080/00335558008248231. [DOI] [PubMed] [Google Scholar]
- Ridderinkhof KR, van der Molen MW, Band GPH. Sources of interference from irrelevant information: A developmental study. Journal of Experimental Child Psychology. 1997;65:315–341. doi: 10.1006/jecp.1997.2367. [DOI] [PubMed] [Google Scholar]
- Rueda MR, Fan J, McCandliss BD, Halparin JD, Gruber DB, Lercari LP, et al. Development of attentional networks in childhood. Neuropsychologia. 2004;42(8):1029–1040. doi: 10.1016/j.neuropsychologia.2003.12.012. [DOI] [PubMed] [Google Scholar]
- Salthouse TA. Aging and measures of processing speed. Biological Psychology. 2000;54:35–54. doi: 10.1016/s0301-0511(00)00052-1. [DOI] [PubMed] [Google Scholar]
- Salthouse TA, Hedden T. Interpreting reaction time measures in between-group comparisons. Journal of Clinical & Experimental Neuropsychology. 2002;24(7):858–872. doi: 10.1076/jcen.24.7.858.8392. [DOI] [PubMed] [Google Scholar]
- Schul R, Townsend J, Stiles J. The development of attentional orienting during the school-age years. Developmental Science. 2003;6(3):262–272. [Google Scholar]
- Trick LM, Jaspers-Fayer F, Sethi N. Multiple-object tracking in children: The “atch the Spies”task. Cognitive Development. 2005;20(3):373–387. [Google Scholar]
- Wilson KD, Woldorff MG, Mangun GR. Control networks and hemispheric asymmetries in parietal cortex during attentional orienting in different spatial reference frames. Neuroimage. 2005;25(3):668–683. doi: 10.1016/j.neuroimage.2004.07.075. [DOI] [PubMed] [Google Scholar]
- Witte EA, Marrocco RT. Alteration of brain noradrenergic activity in rhesus monkeys affects the alerting component of covert orienting. Psychopharmacology. 1997;132(4):315–323. doi: 10.1007/s002130050351. [DOI] [PubMed] [Google Scholar]
- Yuji H. Computer games and information-processing skills. Perceptual & Motor Skills. 1996;83(2):643–647. doi: 10.2466/pms.1996.83.2.643. [DOI] [PubMed] [Google Scholar]