

Abstract
This study was undertaken to determine the modulation of uterine function by chorionic gonadotrophin (CG) in a nonhuman primate. Infusion of recombinant human CG (hCG) between days 6 and 10 post ovulation initiated the endoreplication of the uterine surface epithelium to form distinct epithelial plaques. These plaque cells stained intensely for cytokeratin and the proliferating cell nuclear antigen. The stromal fibroblasts below the epithelial plaques stained positively for α-smooth muscle actin (αSMA). Expression of αSMA is associated with the initiation of decidualization in the baboon endometrium. Synthesis of the glandular secretory protein glycodelin, as assessed by Western blot analysis, was markedly up-regulated by hCG, and this increase was confirmed by immunocytochemistry, Northern blot analysis, and reverse transcriptase-PCR. To determine whether hCG directly modulated these uterine responses, we treated ovariectomized baboons sequentially with estradiol and progesterone to mimic the hormonal profile of the normal menstrual cycle. Infusion of hCG into the oviduct of steroid-hormone-treated ovariectomized baboons induced the expression of αSMA in the stromal cells and glycodelin in the glandular epithelium. The epithelial plaque reaction, however, was not readily evident. These studies demonstrate a physiological effect of CG on the uterine endometrium in vivo and suggest that the primate blastocyst signal, like the blastocyst signals of other species, modulates the uterine environment prior to implantation.
The establishment of pregnancy in pigs and ruminants requires an interaction between the embryo, uterus, and corpus luteum (CL) (1). These interactions prevent luteal regression by extending the functional life span of the CL. The antiluteolytic response results from trophoblast signals acting in a paracrine manner on the uterine endometrium to inhibit the pulsatile release of prostaglandin F2α. In contrast, the primate embryonic signal is chorionic gonadotropin (CG), which has a direct luteotrophic effect on the lifespan of the CL.
Successful implantation requires a genetically normal embryo and a receptive uterine endometrium (2, 3). Uterine receptivity has been defined as the limited period of time when the uterine luminal epithelium is favorable to blastocyst implantation (4). Estrogen and progesterone play a critical role in establishing the receptive phase. However, it is becoming increasingly evident from rodent studies that the embryo induces functional receptivity, which is required for successful nidation (5).
The presence of luteinizing hormone (LH)/CG receptors and associated G proteins has been documented in the human endometrium (6, 7). In addition, human CG (hCG) and the α-subunit of hCG have been shown to induce decidualization of human stromal fibroblasts in vitro (6, 8). Although no evidence is available for the direct effects of CG on the endometrium, the possibility exists that CG can affect the estrogen- and progesterone-primed uterine endometrium in vivo and enhance embryo–maternal interactions during implantation.
In this study we show that infusion of hCG, in a manner that mimics blastocyst transit in the baboon, has physiological effects on the three major cell types present in the uterine endometrium. We suggest that bidirectional signals between the primate embryo and hormonally primed uterine endometrium are a prerequisite for successful implantation.
MATERIALS AND METHODS
Tissue Collection.
Uterine tissue was obtained from adult female baboons (Papio anubis) either at hysterectomy or after endometriectomy. All experimental procedures were approved by the Animal Care Committee of the University of Illinois at Chicago.
Uterine tissue was obtained from four different groups: Cycling baboons treated with bioactive hCG (n = 4) or heat-inactivated hCG (n = 4) and ovariectomized baboons (n = 2) treated with steroid implants and hCG or heat-inactivated hCG. Ovulation was detected in normally cycling female baboons by measuring serum estradiol levels, beginning 7 days after the first day of menses (Fig. 1A). The day of the estradiol surge is day −1, day 0 is the day of the ovulatory LH surge, and day 1 is the day of ovulation. On day 6 post ovulation (PO), a V4 polyvinyl cannula (Bolab Products, Lake Havasu City, AZ) was inserted into the oviduct ipsilateral to the CL. The cannula was secured by suturing the mesosalpinx to a bead of medical adhesive. The open end of the cannula was exteriorized through a subcutaneous flank incision and attached to an Alzet osmotic minipump (2 ML1; Alza, Palo Alto, CA). The minipump containing bioactive or heat-inactivated (30 min of boiling) recombinant hCG (250 international units in 2 ml of distilled water) was primed overnight at 37°C according to the manufacturer’s specifications. The flow rate of the pump was 10 μl/h and the hourly infusate was 1.25 international units. The 24-h infusion of 30 international units is equivalent to the amount of CG secreted by dispersed baboon trophoblast cells cultured in vitro in a 24-h period (9). Endometrial tissue was harvested on day 10 PO for analysis. The time points for the initiation of CG infusion and endometrial tissue isolation correspond to the window of CL rescue (days 5–7 PO) and implantation (days 9–10 PO) in the baboon.
Figure 1.

Diagrammatic illustration of the experimental design for hCG infusion into normally cycling baboons (A) and ovariectomized baboons (B). The steroid treatment regimen for ovariectomized baboons is based on an idealized 28-day menstrual cycle (10). Day 0 in this group is 30 days after ovariectomy. E2, estradiol; P, progesterone.
To determine whether hCG acts directly on the endometrium, we developed an ovariectomized, simulated pregnant model. Ovariectomized baboons were treated sequentially with silastic implants containing estradiol and progesterone to mimic the hormonal changes during the menstrual cycle (ref. 10; Fig. 1B). Six days after the insertion of the first progesterone implant, hCG treatment was initiated and continued for 4 days. Endometrial tissue was harvested at the end of the treatment period. This hormonal regimen results in serum hormone levels that are comparable to those measured in intact animals (10, 11).
Estradiol and progesterone levels in peripheral serum were determined with a validated radioimmunoassay (11). Uterine flushings were obtained from all animals at the time of surgery by washing the uterine lumen with 10 ml of sterile Ca2+- and Mg2+-free Hanks buffered salt solution.
Histology and Immunocytochemistry.
Uterine tissues were immersion-fixed in Bouin’s solution for 24 h at room temperature, dehydrated in graded ethanol, cleared in xylene, and embedded in paraffin (12). For histological analyses the tissue sections (6 μm) were stained with Gomori’s trichrome stain (13). For immunocytochemistry, a monoclonal antibody to α-smooth muscle actin (αSMA) (ref. 14; Dako) at a 1:1000 dilution and a polyclonal antibody to human glycodelin (15) at 1:750 dilution were used. The immunoreactive product was visualized by using an ABC Vectastain kit (Vector Laboratories) and diaminobenzidine (14). Controls consisted of preimmune serum at the same dilution or the omission of the primary antibody.
Explant Culture and Immunoblotting.
Endometrial tissues (75 or 150 mg) from each of the four groups were cultured under serum-free conditions (16). The explant culture media were harvested after 24 h of culture (16). Secretory proteins in explant culture media (200 μl) or uterine flushings (25 μg) were resolved by one-dimensional SDS/PAGE and transferred to nitrocellulose membranes (17). The membranes were incubated overnight with glycodelin antibody (1:5000 dilution), and the immunoreactive product was visualized by using an enhanced chemiluminescence kit (Amersham).
Northern Blotting and Reverse Transcriptase (RT)-PCR.
Northern blots. Total endometrial RNA (20 μg) from the hCG-treated samples and heat-inactivated controls was subjected to electrophoresis on 1% formaldehyde/agarose gels and transferred to nitrocellulose membranes (12). The membranes were hybridized with a 32P random primer-labeled 822-bp cDNA insert complementary to human glycodelin DNA (18). The hybridization signal was visualized by exposing the membrane to Kodak X-Omat film with a DuPont Cronex intensifying screen at −80°C. The autoradiographs were densitometrically scanned with a Molecular Dynamics PhosphorImager.
RT-PCR.
RNA from all experimental groups was reverse transcribed and subjected to PCR. As a control, RNA preparations that were not reverse transcribed were also subjected to PCR to ensure that no genomic contamination was being amplified. Specificity of the PCR products was further confirmed by Southern hybridization with the human glycodelin cDNA (18). The primers were synthesized on the basis of previously described sequences (19). The sense primer sequence was 5′-CCC CCA GAC CAA GCA GGA CCT GGA GCT CCC 3-′, and the antisense primer was 5′-ATC GTC CTG CAC CAG GAC TCT GGC CAG GTA-3′. The histone 3.3 PCR product was coamplified with glycodelin and served as an internal standard (20).
The RT-PCR was performed as previously described for baboon endometrial tissues (20). The RT-PCR procedure for glycodelin was optimized with RNA isolated from the endometrium of an early pregnant baboon. A linear curve was plotted with number of cycles of amplification versus densitometric values of PCR products, and 24 cycles was found to be in the linear range for both PCR products. For these studies, 1 μg of RNA was subjected to reverse transcription and PCR amplification consisting of an initial denaturation at 92°C for 90 sec, followed by 24 cycles of 92°C (1 min), 55°C (30 sec), 72°C (1 min), and a final extension at 72°C for 15 min. The PCR products were electrophoresed in 1.5% agarose gels and subjected to autoradiography. Data were quantitated by image analysis using the PhosphorImager.
RESULTS
Morphology and αSMA Induction.
In intact baboons, infusion of hCG results in the formation of distinct epithelial plaques in the luminal epithelium (arrow; Fig. 2A). In ovariectomized baboons, infusion of hCG increases the cell height of the luminal epithelium but does not induce epithelial plaque formation (Fig. 3A). In contrast, infusion of heat-inactivated hCG has no effect on the luminal epithelium (Figs. 2B and 3B) and the morphological appearance is comparable to the luminal epithelium at day 10 PO during the normal menstrual cycle (data not shown).
Figure 2.

Morphology (A and B) and immunolocalization of αSMA (C and D) in cycling baboons treated with hCG. Note the distinct epithelial plaques that are associated with the luminal epithelium (arrow; A) and the localization of αSMA in stromal fibroblasts below the plaques (St; C) in hCG-treated animals. The controls treated with heat-inactivated CG showed no response in either of the two cell types (B and D). [Final magnification ×77 (A and B) and ×45 (C and D).]
Figure 3.

Morphology (A and B) and immunolocalization of αSMA (C and D) in ovariectomized baboons. Note the absence of distinct epithelial plaques in the hCG-treated baboon (A). In contrast, the induction of αSMA in stromal cells (St; C) was comparable to that seen in cycling baboons treated with hCG. Ovariectomized baboons treated with heat-inactivated CG showed no response in either cell type (B and D). (Final magnification ×45.)
Our previous studies showed that long-term treatment with intramuscular injections of hCG (11, 14) induces the synthesis of αSMA in stromal fibroblasts. Immunostaining of serial sections obtained in this study with a monoclonal antibody to αSMA (14, 20) showed that the stromal fibroblasts directly below the epithelial plaques and luminal epithelium of hCG-treated baboons stained intensely for αSMA (Figs. 2C and 3C). In contrast, staining for αSMA was evident only in the smooth muscle cells of the spiral arteries in the heat-inactivated controls (Figs. 2D and 3D).
Glycodelin Expression.
Glycodelin is markedly up-regulated in the glandular epithelium of pregnant and long-term hCG-treated baboons (15). Short-term infusion of hCG into intact baboons also markedly increases the staining for this protein in the glandular epithelium (Fig. 4A). Immunostaining of tissues from heat-inactivated controls showed limited staining within the glands (Fig. 4B).
Figure 4.

Immunocytochemical localization of glycodelin in cycling baboons treated with hCG. Treatment with hCG markedly up-regulates the staining for glycodelin in the glandular epithelial cells (A). A limited amount of immunoreactivity is evident in the perinuclear region of the epithelial cells in the heat-inactivated controls (B). (Final magnification ×74.)
Western blot analysis of explant culture media from four hCG-treated intact baboons shows that the major form of glycodelin synthesized is a 30-kDa protein (Fig. 5, lanes 3–6). In contrast, explant culture media of endometrial tissues obtained from the four heat-inactivated controls shows the presence of the lower molecular weight form of this protein (Fig. 5, lanes 7–10). This form of glycodelin is also secreted by the baboon endometrium on day 10 PO of the normal menstrual cycle (Fig. 5, lane 2). Lane 1 is the purified form of human glycodelin isolated from cytosolic extracts of first-trimester decidua (21).
Figure 5.

Western blot of explant culture medium. Protein from conditioned media after 24 h of culture was immunoreacted with a polyclonal antibody to human glycodelin. Treatment with hCG markedly increased the synthesis and the molecular size of glycodelin (lanes 3–6). Endometrial explants from the heat-inactivated controls secreted a low molecular weight form of glycodelin (lanes 7–10) comparable to that seen normally at day 10 PO (lane 2). Lane 1 is the human protein (0.25 μg) purified from first-trimester cytosolic extracts.
In response to hCG treatment, two distinct glycodelin transcripts (1.7 and 1.0 kb) were evident on Northern blots, whereas message levels were very low after infusion of heat-inactivated hCG (data not shown). These two transcripts are similar to those that are expressed in the endometrium during pregnancy and after long-term treatment with exogenous CG (15).
Immunocytochemical analysis of endometrial tissues from ovariectomized baboons treated with hCG showed very limited immunoreactivity for glycodelin compared with the intact animals (data not shown). However, Western blot analysis of explant culture media (Fig. 6, lanes 2 and 3) and uterine flushings (lane 6) from these animals revealed that the higher molecular weight form of glycodelin was induced in response to hCG treatment. Lane 1 is culture medium from uterine endometrium at early pregnancy (day 14). Heat inactivation of hCG failed to induce the high molecular weight form (lanes 4, 5, and 7).
Figure 6.
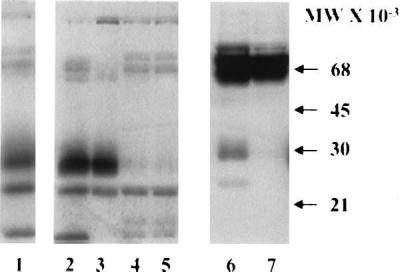
Western blot of explant culture medium (lanes 2–5) and uterine flushings (lanes 6 and 7) from ovariectomized baboons. Note that treatment with bioactive hCG (lanes 2, 3, and 6) resulted in the secretion of the higher molecular weight form of glycodelin both in vivo (lane 6) and in vitro (lanes 2 and 3). Only the lower molecular weight form was evident in explant culture media (lanes 4 and 5) but not in uterine flushings (lane 7) obtained from the heat-inactivated controls (lanes 4 and 5). Endometrial explant culture medium obtained at early pregnancy (day 14) is shown for comparison (lane 1).
Regulation of glycodelin mRNA by hCG was further confirmed by RT-PCR (Fig. 7 A and B). Two major amplicons (383 and 297 bp) are strongly expressed during early pregnancy (lanes 1 and 2) and after hCG infusion in cycling baboons (lanes 3 and 4). Although little or no mRNA was evident in the endometrium of heat-inactivated controls by Northern blot analysis, RT-PCR demonstrated that a lower level of glycodelin expression is readily evident (lanes 5 and 6). This corresponds to the limited immunostaining that is shown in Fig. 4B. Infusion of hCG into ovariectomized, steroid-treated baboons induced amplicons comparable to those in cycling controls (Fig. 7, lane 7). In contrast, no amplicons were evident after infusion of heat-inactivated hCG into ovariectomized baboons (lane 8). The histone 3.3 internal control was amplified equally in all lanes and the ratio of each of the amplicons to the internal control is shown graphically in Fig. 7B.
Figure 7.

(A) RT-PCR for glycodelin transcripts in early pregnant (lanes 1 and 2), hCG-treated (lanes 3 and 4), and heat-inactivated hCG-treated (lanes 5 and 6) cycling baboons (A). Lanes 7 and 8 represent ovariectomized baboons treated with hCG or heat- inactivated hCG, respectively. Note that two major transcripts (383 and 279 bp) are amplified during pregnancy and after hCG infusion. The 213-bp transcript is the histone 3.3 internal standard that was coamplified with the glycodelin primers. (B) The bar graphs represent the mean values of the densitometric scans of each transcript shown in A and expressed as a ratio to histone 3.3. Note that in response to CG either in pregnancy (bars 1) or with hCG infusion (bars 2) the higher molecular weight transcript (383 bp; black bars) predominates. The response after ovariectomy is attenuated, but the higher molecular weight form is evident in animals treated with bioactive hCG (lane 7 in A and 4 in B). The gray bars represent the low molecular weight (279 bp) transcript.
DISCUSSION
In all mammalian species, an ovulatory surge of LH initiates a series of events that culminates in ovulation. The resulting CL begins to secrete progesterone, which is essential for developing a receptive endometrium that is favorable to embryo implantation. In pigs and ruminants, embryonic signals secreted during the period of maternal recognition of pregnancy exert paracrine antiluteolytic effects on the endometrium to inhibit the luteolytic pulses of prostaglandin F2α (1). In contrast, CG, the embryonic signal in primates, exerts a direct luteotrophic effect on the CL and extends its lifespan (22).
The presence of specific binding sites for CG has been reported in the human endometrium and myometrium (23–25). In addition, in vitro studies have suggested that hCG can influence biochemical changes in isolated stromal cells (6, 8). This study demonstrates that CG has physiological effects in vivo on the primate uterine endometrium during the period of uterine receptivity, and it supports the concept that the blastocyst plays an active role in transforming the epithelial barrier into a localized gateway to the stromal cells (26). It is evident from our studies that CG induces specific changes in the luminal epithelium, glandular epithelium, and stromal fibroblasts in the progestationally primed uterine endometrium. Further support for this response is evidenced by the fact that the morphological, biochemical, and molecular changes induced by CG are reproducible in an ovariectomized baboon treated with exogenous steroids and hCG.
Morphological transformation of the endometrium into a secretory phenotype is a prerequisite for implantation. In pregnant baboons, an early maternal response to pregnancy is the formation of the epithelial plaque (27). This response is characterized by the hypertrophy of the surface epithelium and cells in the neck glands that round up and form acinar clusters (28, 29). In pregnant baboons the plaque reaction is restricted to the implantation site (27). However, in the-CG treated animals, the plaque reaction was evident over the entire luminal surface. This difference may reflect a local response to CG secreted by the implanting blastocyst during pregnancy as opposed to a more diffused response to the infusion of CG in this simulated model. Thus, although the morphological response of the luminal epithelium to hCG infusion or blastocyst implantation is similar to a certain extent, the other factor(s) required for the transformation remain to be elucidated. Implantation has been characterized as an inflammatory-type response (26). In coronary disease, plaque growth within the arterial wall is thought to be fostered by cytokines and other growth-regulatory molecules produced by inflammatory cells and the smooth muscle (30). Since a primary function of the uterine epithelium is thought to be the activation of the preimplantation blastocyst by means of the local action of cytokines or eicosanoids (31–33), it is conceivable that the induction of the plaque reaction may be activated by the infiltration of inflammatory cells into the uterus and may be a prerequisite for the initial phase of blastocyst–endometrial interactions.
Studies with transformed Leydig cells (MA10 cells) suggest that hCG induces cytoskeletal-dependent receptor clustering, which appears to be a prerequisite for steroidogenesis (34, 35). In addition, rapid reorganization of the cytoskeleton is necessary for cellular motility and cell division (36, 37). Our studies indicate that the primary effect of hCG on stromal fibroblasts is the induction of αSMA. We propose that the induction of αSMA in stromal fibroblasts occurs in response to integrins on the stromal cell membranes binding to secreted extracellular matrix (ECM) proteins. We have previously demonstrated that hCG induces specific changes in integrin expression and ECM secretion in the baboon endometrium (38). The interaction between integrins and the ECM induces changes in the actin cytoskeleton that are thought to be critical for signal transduction (39, 40). It is also becoming increasingly evident that the actin cytoskeleton plays a critical role in mediating the cellular response to stimulation by growth factors (41, 42). Thus, it is conceivable that the induction of αSMA by hCG may be essential for stromal cell proliferation and differentiation during the initial stages of pregnancy. Indeed, αSMA transcriptionally inhibits the induction of insulin-like growth factor binding protein 1 (IGFBP-1; ref. 43). Since IGFBP-1 is thought to inhibit trophoblast migration (44), the induction of αSMA prior to trophoblast attachment may also play a role to facilitate the rapid migration of trophoblasts into the maternal endometrium.
Glycodelin is the major secretory protein of the human endometrium during the late luteal phase and early pregnancy (21). We have previously demonstrated that the synthesis of glycodelin by the baboon uterine endometrium parallels the rise and decline of CG in the peripheral circulation (15). In this study we demonstrate that infusion of hCG during the period of uterine receptivity appears to up-regulate glycodelin transcription and translation. Immunocytochemical and RT-PCR analyses suggest that glycodelin expression was higher in the intact baboons compared with the ovariectomized animals. A close temporal relationship between relaxin and glycodelin serum profiles has been reported in human females (45). Since we have previously demonstrated that CG increases relaxin production by the CL,§ it is possible that the difference we observed between the two groups may be related to the absence of relaxin synthesis from the CL in the ovariectomized animals. The post-translational modification includes glycosylation, and it has been suggested that the carbohydrate moieties on glycodelin are associated with its immunomodulatory properties (46, 47) and its ability to inhibit sperm binding to the zona pellucida (48). More recently, transfection studies with human glycodelin cDNA have suggested that this protein may play an important role in epithelial cell differentiation (49). Successful implantation of a normal blastocyst requires a receptive uterine endometrium that is both morphologically and functionally mature (2). During the normal menstrual cycle, estrogen and progesterone suitably prime the endometrium to be receptive, but it is becoming increasingly evident that a functionally receptive uterus is induced only by a suitable embryonic stimulus. Thus, we propose that the induction of glycodelin by CG permits the glandular epithelial cells to be transformed into a fully differentiated secretory phenotype.
In summary, our in vivo studies have clearly demonstrated that infusion of hCG, in a manner that mimics the period of blastocyst transit and initial attachment, markedly alters the morphology and function of the luminal epithelium, glandular epithelium, and stromal fibroblasts. In addition, we have provided direct evidence to suggest that besides its luteotrophic function, CG has additional physiological effects on the uterine endometrium during the period of uterine receptivity. What is not readily evident are the signaling mechanisms by which CG affects each of the different cell types within the endometrium. In vitro studies with isolated cells from the human endometrium suggest that the expression of LH/CG receptors is higher in the glandular epithelium compared with stromal cells (23). However, no direct response to hCG in glandular epithelial cells cultured in vitro has been reported. In contrast, stromal cells have been shown to respond to exogenous hCG by increasing the expression of cyclooxygenase 2 and prolactin (8, 50). Whether this response is induced by the intact hCG molecule by means of the well established G-protein-coupled LH/CG receptor or by means of a novel mechanism induced locally by the α-subunit of hCG continues to be a matter of debate (6, 8). On the basis of our preliminary data in the baboon, we suggest that stromal cell transformation may be induced by a novel receptor mechanism. The signal transduction pathways and the resulting gene transcription of αSMA and glycodelin require investigation.
Acknowledgments
We thank Serono Laboratories for their generous donation of recombinant hCG and Dr. Stephen Bell for the glycodelin antibody and cDNA. The technical assistance of Ms. Heidi Hausermann and Ms. Stephanie Banaszak, and valuable discussions with Dr. Harold Verhage are gratefully acknowledged. This work was supported by the National Cooperative Program for Markers of Uterine Receptivity for Blastocyst Implantation funded by the National Institutes of Health Cooperative Agreement HD 29964.
ABBREVIATIONS
- CL
corpus luteum
- CG
chorionic gonadotrophin
- hCG human CG
LH, luteinizing hormone
- PO
post ovulation
- αSMA
α-smooth muscle actin
- RT-PCR
reverse transcriptase–PCR
Footnotes
Hild-Petito, S. & Fazleabas, A. T., Society for Gynecologic Investigation Annual Meeting, March 20–24, 1993, Chicago, abstr. S188.
References
- 1.Bazer F W, Ott T L, Spencer T E. In: Endocrinology of Pregnancy. Bazer F W, editor. Totowa, NJ: Humana; 1998. pp. 1–34. [Google Scholar]
- 2.Yoshinaga K. In: Maternal and Fetal Endocrinology. Tulchinsky D, Little A B, editors. Philadelphia: Saunders; 1994. pp. 336–349. [Google Scholar]
- 3.Heap R B, Flint A P, Gadsby J E. Br Med Bull. 1979;35:129–135. doi: 10.1093/oxfordjournals.bmb.a071559. [DOI] [PubMed] [Google Scholar]
- 4.Pschoyos A. Ann N Y Acad Sci. 1986;476:36–42. doi: 10.1111/j.1749-6632.1986.tb20920.x. [DOI] [PubMed] [Google Scholar]
- 5.Dey S K, Paria B C, Huet-Hudson Y M. In: Molecular and Cellular Aspects of the Periimplantation Process. Dey S K, editor. New York: Springer; 1995. pp. 113–124. [Google Scholar]
- 6.Rao C V, Sanfilippo J S. Endocrinologist. 1997;7:107–111. [Google Scholar]
- 7.Bernardini L, Rojas-Moretti I, Brush M, Rojas F J, Balmaceda J P. J Soc Gynecol Invest. 1995;2:630–635. doi: 10.1016/1071-5576(95)00010-c. [DOI] [PubMed] [Google Scholar]
- 8.Nemansky M, Moy E, Lyons C D, Yu I, Blithe D L. J Clin Endocrinol Metab. 1998;83:575–581. doi: 10.1210/jcem.83.2.4593. [DOI] [PubMed] [Google Scholar]
- 9.Bambra C S, Tarara R. J Reprod Fertil. 1990;88:9–16. doi: 10.1530/jrf.0.0880009. [DOI] [PubMed] [Google Scholar]
- 10.Fazleabas A T, Miller J B, Verhage H G. Biol Reprod. 1988;39:729–736. doi: 10.1095/biolreprod39.3.729. [DOI] [PubMed] [Google Scholar]
- 11.Hild-Petito S, Donnelly K M, Miller J B, Verhage H G, Fazleabas A T. Endocrine. 1995;3:639–651. doi: 10.1007/BF02746340. [DOI] [PubMed] [Google Scholar]
- 12.Fazleabas A T, Jaffe R C, Verhage H G, Waites G, Bell S C. Endocrinology. 1989;124:2321–2329. doi: 10.1210/endo-124-5-2321. [DOI] [PubMed] [Google Scholar]
- 13.Skeehan D C, Hrapchak B B. Theory and Practice of Histotechnology. St. Louis: Mosby; 1973. pp. 111–112. [Google Scholar]
- 14.Christensen S, Verhage H G, Nowak G, de Lanerolle P, Fleming S, Bell S C, Fazleabas A T, Hild-Petito S. Biol Reprod. 1995;53:596–606. doi: 10.1095/biolreprod53.3.598. [DOI] [PubMed] [Google Scholar]
- 15.Hausermann H M, Donnelly K M, Bell S C, Verhage H G, Fazleabas A T. J Clin Endocrinol Metab. 1998;83:1226–1233. doi: 10.1210/jcem.83.4.4741. [DOI] [PubMed] [Google Scholar]
- 16.Fazleabas A T, Verhage H G. Biol Reprod. 1987;37:979–988. doi: 10.1095/biolreprod37.4.979. [DOI] [PubMed] [Google Scholar]
- 17.Towbin H, Staehelin T, Gordon J. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Garde J, Bell S C, Eperon I C. Proc Natl Acad Sci USA. 1991;88:2456–2460. doi: 10.1073/pnas.88.6.2456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ace C I, Okulicz W C. Mol Cell Endocrinol. 1995;115:95–103. doi: 10.1016/0303-7207(95)03674-v. [DOI] [PubMed] [Google Scholar]
- 20.Kim J J, Jaffe R C, Fazleabas A T. Biol Reprod. 1998;59:160–168. doi: 10.1095/biolreprod59.1.160. [DOI] [PubMed] [Google Scholar]
- 21.Bell S C, Hales M W, Patel S, Kirwan P H, Drife J O. Br J Obstet Gynecol. 1985;92:793–803. doi: 10.1111/j.1471-0528.1985.tb03048.x. [DOI] [PubMed] [Google Scholar]
- 22.Stouffer R L, Hearn J P. In: Endocrinology of Pregnancy. Bazer F W, editor. Totowa, NJ: Humana; 1998. pp. 35–58. [Google Scholar]
- 23.Reshef E, Lei Z M, Rao Ch V, Pridham D D, Chegini N, Luborsky J L. J Clin Endocrinol Metab. 1990;70:421–430. doi: 10.1210/jcem-70-2-421. [DOI] [PubMed] [Google Scholar]
- 24.Lei Z M, Reshef E, Rao C V. J Clin Endocrinol Metab. 1990;75:651–659. doi: 10.1210/jcem.75.2.1379262. [DOI] [PubMed] [Google Scholar]
- 25.Bhattacharya S, Banerjee J, Sen S, Manna P R. Acta Endocrinol. 1993;129:15–19. doi: 10.1530/acta.0.1290015. [DOI] [PubMed] [Google Scholar]
- 26.Lopata A. Mol Hum Reprod. 1996;2:519–525. doi: 10.1093/molehr/2.7.519. [DOI] [PubMed] [Google Scholar]
- 27.Enders A C, Lantz K C, Peterson P E, Hendrickx A G. Hum Reprod Update. 1997;3:561–573. doi: 10.1093/humupd/3.6.561. [DOI] [PubMed] [Google Scholar]
- 28.Tarara R, Enders A C, Hendrickx A C. Anat Embryol. 1987;176:267–275. doi: 10.1007/BF00310182. [DOI] [PubMed] [Google Scholar]
- 29.Enders A C. Placenta. 1991;12:309–325. doi: 10.1016/0143-4004(91)90340-l. [DOI] [PubMed] [Google Scholar]
- 30.Kol A, Libby P. Trends Cardiol Med. 1998;8:191–193. doi: 10.1016/s1050-1738(98)00010-3. [DOI] [PubMed] [Google Scholar]
- 31.Cross J C, Werb Z, Fisher S J. Science. 1994;266:1508–1518. doi: 10.1126/science.7985020. [DOI] [PubMed] [Google Scholar]
- 32.Sharkey A. Reprod Med Rev. 1995;4:87–100. [Google Scholar]
- 33.Tabibzadeh S, Babaknia A. Hum Reprod. 1995;10:1579–1602. doi: 10.1093/humrep/10.6.1579. [DOI] [PubMed] [Google Scholar]
- 34.Ascoli M, Segaloff D L. Endocrinology. 1987;120:1161–1172. doi: 10.1210/endo-120-3-1161. [DOI] [PubMed] [Google Scholar]
- 35.Maurer D R, Wimalasena J, Indrapichate K, Puet D, Majercik M H. Cell Physiol Biochem. 1995;5:361–370. [Google Scholar]
- 36.Berfield A K, Spicer D, Abrass C K. Histochem Cytochem. 1997;45:583–593. doi: 10.1177/002215549704500410. [DOI] [PubMed] [Google Scholar]
- 37.Feldman E L, Sullivan K A, Kim B, Russell J W. Neurobiol Dis. 1997;4:201–214. doi: 10.1006/nbdi.1997.0156. [DOI] [PubMed] [Google Scholar]
- 38.Fazleabas A T, Bell S C, Fleming S, Sun J, Lessey B A. Biol Reprod. 1997;56:348–356. doi: 10.1095/biolreprod56.2.348. [DOI] [PubMed] [Google Scholar]
- 39.Clark E A, Brugge J S. Science. 1995;268:233–239. doi: 10.1126/science.7716514. [DOI] [PubMed] [Google Scholar]
- 40.Dedhar S. Cancer Metastasis Rev. 1995;14:165–172. doi: 10.1007/BF00690289. [DOI] [PubMed] [Google Scholar]
- 41.Dedhar S, Hannigan G E. Curr Opin Cell Biol. 1996;8:657–669. doi: 10.1016/s0955-0674(96)80107-4. [DOI] [PubMed] [Google Scholar]
- 42.Blakesley V A, Koval A P, Stannard B S, Scringeour A, LeRoith D. J Biol Chem. 1998;273:18411–18422. doi: 10.1074/jbc.273.29.18411. [DOI] [PubMed] [Google Scholar]
- 43.Kim J J, Jaffe R C, Fazleabas A T. Endocrinology. 1999;140:997–1004. doi: 10.1210/endo.140.2.6474. [DOI] [PubMed] [Google Scholar]
- 44.Irwin J C, Giudice L C. Growth Horm IGF Res. 1998;8:21–31. doi: 10.1016/s1096-6374(98)80318-3. [DOI] [PubMed] [Google Scholar]
- 45.Stewart D R, Erickson M S, Erickson M E, Nakajima S T, Overstreet J W, Laskey B L, Amento E P, Seppala M. J Clin Endocrinol Metab. 1997;82:839–846. doi: 10.1210/jcem.82.3.3839. [DOI] [PubMed] [Google Scholar]
- 46.Pockley A C, Bolton A E. Immunology. 1990;69:137–142. [PMC free article] [PubMed] [Google Scholar]
- 47.Okamoto N A, Uchida A, Takamura K, Kariya Y, Kanzaki H. Am J Reprod Reprod Immunol. 1991;26:137–142. doi: 10.1111/j.1600-0897.1991.tb00713.x. [DOI] [PubMed] [Google Scholar]
- 48.Oehinger S, Coddington C C, Hodgen G D, Seppala M. Fertil Steril. 1995;63:377–383. doi: 10.1016/s0015-0282(16)57372-5. [DOI] [PubMed] [Google Scholar]
- 49.Kamarainen M, Seppala M, Virtanen I, Anderson L C. Lab Invest. 1997;77:565–573. [PubMed] [Google Scholar]
- 50.Han S W, Lei Z M, Rao C V. Endocrinology. 1996;137:1791–1797. doi: 10.1210/endo.137.5.8612516. [DOI] [PubMed] [Google Scholar]