

Abstract
Telomerase reverse transcribes telomere DNA onto the ends of linear chromosomes and retards cellular aging. In contrast to most normal somatic cells, which show little or no telomerase activity, immune cells up-regulate telomerase in concert with activation. Nevertheless, during aging and chronic HIV-1 infection, there are high proportions of dysfunctional CD8+ CTL with short telomeres, suggesting that telomerase is limiting. The present study shows that exposure of CD8+ T lymphocytes from HIV-infected human donors to a small molecule telomerase activator (TAT2) modestly retards telomere shortening, increases proliferative potential, and, importantly, enhances cytokine/chemokine production and antiviral activity. The enhanced antiviral effects were abrogated in the presence of a potent and specific telomerase inhibitor, suggesting that TAT2 acts primarily through telomerase activation. Our study is the first to use a pharmacological telomerase-based approach to enhance immune function, thus directly addressing the telomere loss immunopathologic facet of chronic viral infection.
Telomeres, the TTAGGG tandem repeats at chromosome ends, become progressively shorter in dividing somatic cells (1). Numerous studies have shown associations between leukocyte telomere shortening and risk of disease, suggesting that these cells may serve as biomarkers for the cumulative burden of inflammation, chronic immune activation, or oxidative stress. Indeed, hypertension and increased insulin resistance are associated with shorter leukocyte telomere length in the participants of the Framingham heart study (2), and telomere shortening in peripheral blood leukocytes chronicles ischemic heart disease risk in older people (3). Leukocyte telomere length also correlates with bone mineral density, and shorter telomeres are seen in women with osteoporosis (4). Finally, in both aging and chronic HIV-1 infection, there are increased proportions of CD8+ T lymphocytes with shortened telomeres, reduced proliferative capacity and altered effector function (5). Thus, strategies to retard or prevent telomere loss may lead to novel treatments for a variety of human pathologies.
T and B lymphocytes transiently up-regulate telomerase, the cellular reverse transcriptase that adds telomeric DNA to the ends of chromosomes (6-8). Such telomerase activity is believed to mitigate the losses of replicative capacity and function caused by chronic antigenic stimulation, oxidative stress, and cellular aging (9, 10). However, the ability of CD8+ T lymphocytes to up-regulate telomerase is lost after repeated encounters with Ag, and continued chronic stimulation ultimately leads to critically short telomeres and other changes associated with replicative senescence (10, 11).
We hypothesized that sustained telomerase activity in chronically activated CD8+ T lymphocytes might prevent or delay immune dysfunction associated with aging and/or chronic disease. In previous studies, we showed that gene transduction of CD8+ T lymphocytes from HIV-1-infected persons with the human telomerase catalytic component (hTERT)4 (12) leads to preservation of antiviral functions, telomere length stabilization, and augmented proliferative potential (13). Whereas gene therapy has a number of drawbacks, such as inability to control gene expression and the potential side-effects of gene transduction, these proof-of-principle studies suggest that telomerase activation by nongenetic strategies may be an effective approach for enhancing immune function in certain chronic diseases.
In an empirical screen of traditional Chinese medicine plant extracts and compounds with reported properties of health maintenance and enhancement of immune function, we identified TAT2 (cycloastragenol) based on its ability to up-regulate the low, basal level of telomerase in neonatal human keratinocytes (unpublished data). We sought to determine whether TAT2 could increase telomerase activity in cells of the immune system, which exhibit high levels of active telomerase after stimulation with mitogens, activatory Abs (10), or Ag (14). In this study, we demonstrate that TAT2 can transiently activate telomerase, slow telomere loss, increase replicative capacity, and, importantly, enhance immune function in CD8+ T lymphocytes from HIV-1-infected persons. These data suggest a possible novel immune-based strategy to complement current treatments, which are primarily directed at the virus. The potential benefit of telomerase enhancement is underscored by the recent clinical study showing that HIV-1-infected persons who are able to control the infection for long periods of time (nonprogressors) have significantly higher constitutive telomerase activity in HIV-1-specific CD8+ T cells compared with “fast progressors” (15).
Materials and Methods
Formulation of TAT2
TAT2 (cycloastragenol, CAS Registry no. 84605-18-5) was prepared by purification of acid hydrolyzed Astragaloside IV (16) and is available from TA Therapeutics (email charley@geron.com). TAT2 was dissolved in tissue culture grade DMSO (ATCC) at 10 mM stock concentration. All titrations of TAT2 were prepared in DMSO at 1000 times final concentration, such that the DMSO concentration in all tissue cultures experiments was constant at 0.1%. Telomerase activation by TAT2 was observed in the 0.01–10 μM range. In some experiments, activity declined above 1 μM, so in most cases TAT2 was not used above 1 μM.
Telomerase inhibitor (TI)
The TI used in our experiments is GRN163L, a synthetic lipid-conjugated, 13-mer oligonucleotide N3′ P5′-thio-phosphoramidate that is complementary to the template region of telomerase RNA (hTR) (17). GRN163L is a direct competitive enzyme inhibitor that binds and blocks the active site of the enzyme (a “telomerase template antagonist”). The IC50 of GRN163L for telomerase in human PBMCs is 1 μM (data not shown).
Telomerase activity measurements
Telomerase activity for all experiments was determined by the telomeric repeat amplification protocol (TRAP), using the reagents, protocol, and calculation details provided in the TRAPeze kit (Millipore; Cat. no. S7710). The amplified TRAP reaction products were separated on an 8% polyacrylamide gel, and the resulting bands were probed and analyzed using Packard InstantImager software. Telomerase activity for all samples was calculated for 10,000 cell-equivalents, according to the TRAPeze kit formula for “Total Product Generated.”
Cell isolation and stimulation
Human peripheral blood samples were acquired after informed consent and in accordance with the University of California-Los Angeles Institutional Review Board. After centrifugation, the PBMC layer was carefully removed and washed twice in “complete RPMI 1640” [10% FBS, 10 mM HEPES, 2 mM glutamine, and 50 IU/ml penicillin/streptomycin]. Purified T cells were obtained from PBMC using the Pan T isolation kit (Miltenyi Biotec), and, for some experiments, CD8+ T cells were further purified by negative selection using a CD4+ T cell isolation kit (Miltenyi Biotec). For isolation of CD28high and CD28low cells, the CD8+ T cell population was incubated for 10 min at 4°C with biotinylated anti-CD28 Ab (BD Pharmingen), followed by 15 min incubation at 4°C with anti-biotin magnetic microbeads (Miltenyi Biotec), and passaged through a MACS magnetic separation column (LS). T cells were stimulated with CD3/CD28 or CD2/CD3/CD28 Ab-coated beads at a bead:cell ratio of 0.5:1, and PBMC were stimulated with culture-tested PHA (5–12 μg/ml; Sigma-Aldrich Cat. no.L1668). For short-term stimulation, in addition to PHA or Ab-coated beads, DMSO (0.1%) or TAT2 (1 nM to 10 μM) was added to wells. Every 48–72 h, half of the media in the wells was removed and replaced with fresh media containing either DMSO or TAT2. Long-term T cell cultures were established by stimulation of PBMC with CD2/CD3/CD28 Ab-coated beads (Miltenyi Biotec) and IL-2 (50 U/ml). At the time of stimulation, TAT2 (1 μM) or DMSO (0.1%) was added to cells. Fresh media was added to cells every 48–96 h along with TAT2 or DMSO. Every 13–17 days, cells were re-stimulated with CD2/CD3/CD28 beads and IL-2. In some experiments, CD4+ and CD8+ T cells were isolated from purified T cells by positive and negative selection, respectively, and stimulated as described above.
Telomere length real-time PCR
The general protocol for telomere length real-time PCR was followed as previously described (18). CD8+ T cells were isolated by negative selection and cultured in a 24-well plate (1 × 106/ml) with CD2/CD3/CD28 Ab-coated beads (Miltenyi Biotec), IL-2 (50 U/ml), and either TAT2 (1 μM) or DMSO (0.1%). Fresh media was added every 48–96 h along with TAT2 or DMSO. Every 13–17 days, cells were re-stimulated with CD2/CD3/CD28 beads and IL-2. Immediately before each stimulation, 1 × 106 cells were removed and cryopreserved for subsequent telomere length analysis, which was performed on all samples in parallel. DNA was isolated from cells using DNeasy kit (Qiagen) and diluted to 50 ng/100 μl in dilution buffer. Each ml of buffer consists of: 40 μl of Escherichia coli DNA (100 ng/μl), 100 μl of 10× Taq polymerase buffer, and 860 μl of H20. DNA samples were boiled at 95°C for 30 min. Samples were run in triplicate in a 96-well plate in the I-CycleriQ Multicolor Real-Time Detection System (Bio-Rad). Separate plates were used for the telomere PCR and the control Human β Globin (HBG) PCR. In the telomere PCR plate, each well contained 10 μl of DNA, 10 μl of iQ SYBR Green Super Mix, 0.2 μl of Tel primer 1 (20 μM stock), and 0.8 μl of Tel primer 2 (20 μM stock), for a total of 21 μl. In the HBG PCR plate, each well contained 10 μl of DNA, 10 μl of iQ SYBR Green Super Mix, 0.3 μl of HBG primer 1 (20 μM stock), and 0.7 μl of HBG primer 2 (20 μM stock), for a total of 21 μl. The sequences of the primers used were: Tel primer 1 (5′ CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTT 3′), Tel primer 2 (5′ GGCTTGCCTTACCCTTACCCTTACCCTTACCCTTACCCT 3′), HBG primer 1 (5′ GCTTCTGACACAACTGTGTTCACTAGC 3′), and HBG primer 2 (5′ CACCAACTTCATCCACGTTCACC 3′). The IQcycler program for the telomere PCR consisted of initial denaturation at 94°C for 1 min, followed by 25 PCR cycles at 95°C for 15 s and 56°C for 1 min (single fluorescence measurement). The IQcycler program for the HBG PCR consisted of initial denaturation at 94°C for 1 min, followed by 36 PCR cycles at 95°C for 15 s, 58°C for 20 s, and 72°C for 20 s (single fluorescence measurement). Standard curves were created for both telomere DNA and HBG DNA. Values were calculated as a ratio of telomere DNA to HBG DNA for each sample, and data was expressed as % of cell line 1301 telomeric DNA. This acute lymphoblastic T cell line is routinely used in real-time PCR telomere length studies (19, 20). In a separate set of experiments (data not shown), we verified that the relative telomere length measurements determined by real-time PCR correlated well with absolute telomere length measurements using Southern blots of terminal restriction fragments, as reported by others (21, 22).
Pathway inhibitors
T cells (1 × 106/ml), isolated as described above, were cultured for 7 days with CD2/CD3/CD28 Ab-coated beads (Miltenyi Biotec) and 20 U/ml of IL-2. Cells were then washed and incubated for 2 h in complete RPMI 1640 containing one of the following inhibitors (Calbiochem): AKT inhibitor 1 (phosphatidylinositol ether analog that potently blocks binding of PIP3 to AKT, Cat. no. 124009), AKT inhibitor 2 (phosphatidylinositol analog that prevents generation of PIP3 by PI3K, Cat. no. 124005), ERK1/2 inhibitor (13-amino acid peptide corresponding to the N terminus of MEK1 (MAPKK), Cat. no. 328000), and MAPK inhibitor PD98059 (blocks the activation of MEK, thus preventing its phosphorylation by cRaf or MEKK, Cat. no. 513000). Cells were then washed twice with media and re-plated in 24-well plates with complete RPMI 1640. TAT2 (1 μM) or DMSO (0.1%) was added to cells, which were harvested 24 h later for the TRAP assay.
p-ERK1/2 activation ELISA
T cells were isolated as described and cultured in a T100 flask in complete RPMI 1640 at a concentration of 1 × 106/ml. Cells were stimulated and grown with or without TAT2, as described. The amount of p-ERK1/2 was determined at the indicated time points, using an ERK1/2 Phospho-Specific (Thr185/Tyr187) Elisa kit (Calbiochem) per the manufacturer’s instructions.
IFN-γ ELISPOT
CD8+ T cells were isolated from PBMC derived from HIV+ donors using the RosetteSep human CD8+ T cell enrichment mixture (StemCell Technologies), and stimulated with Ab-coated beads and IL-2, as described above. Every 48 –72 h, half the media was replaced with fresh complete RPMI 1640 supplemented with either TAT2 (1 μM) or DMSO (0.1%). After 14 days, cells were harvested and tested in an ELISPOT assay for IFN-γ (23). HIV peptides used (based on previous mapping studies for these donors) were Gag 5026, Vif 6036, Nef 5172, Nef 5158, and Env 6398, all at 5 μg/ml.
Chemokine assays
For detection of chemokine secretion by TAT2-treated CD8+ T cells, the following Quantikine ELISA kits were used (R&D Systems), according to the manufacturer’s instructions; CCL3/MIP1α kit (Cat no. DMA00, CCL4/MIP1β kit (Cat. no. DMB00) and CCL5/RANTES kit (Cat. no. DRN00B)) was used.
Viral inhibition assay
Coculture viral inhibition assays were conducted as previously described (24). In brief, PBMC from HIV+ donors were isolated as described above. To expand separate populations of CD8+ and CD4+ T cells, PBMC were plated in 24-well plates with 1 μg/ml of either CD3/CD4 or CD3/CD8 bi-specific Ab (a gift from Dr. Johnson T. Wong, Harvard Medical School, Boston, MA). During the expansion period, cells were maintained in 50 units IL-2/ml;TAT2 (1 μM) or DMSO (0.1%) was added every 48–72 h to the CD8+ cells. After infection of the CD4+ cells, cocultures with the autologous CD8+ cells were established in round-bottom 96-well plates and incubated for 10 days with IL-2 (20–50 U/ml) supplemented complete RPMI 1640 containing either TAT2 (1 μM) or DMSO (0.1%), (in some cases along with a TI). On days 4, 7, and 10, half the media in each well was carefully removed for analysis and replaced with fresh media. Viral production was measured using a commercial p24 ELISA kit (PerkinElmer, Cat. No. NEK050). Since viral production varied dramatically between donors, viral inhibition in the experimental cultures are reported as a percent of that seen in the DMSO control cocultures for each donor: (1-(mean p24 levels for experimental wells)/(mean p24 for control wells)) × 100.
Statistical analysis
Quantitative, matched pairs data were analyzed using a two-sided Student’s paired t test to account for the correlation in the matched pairs. p values <0.05 were considered significant. From the data seen in Fig. 5d, longitudinal measurements were assessed using a mixed effects model to account for the correlation across time. Each subject’s intercept was allowed to deviate randomly from the population mean. Results from the analysis were used to project time until half and full log decrease in viral production.
FIGURE 5.

Inhibition of HIV viral production from infected CD4+ T cells by TAT2-treated CD8+ T cells from HIV-1-infected donors. a, Relative inhibition of viral production from infected CD4+ T cells by TAT2-treated CD8+ T cells from nine separate HIV+ donors after 10-day coculture. Values are given as percent reduction in p24 levels in coculture assay with TAT2-treated CD8+ T cells relative to DMSO-treated CD8+ T cells. Mean and SD are given. b, Longitudinal analysis of viral inhibition for the same nine donors as in a during coculture (gray lines show individual donors, black line shown average for the nine donors). Values are given as log reduction in p24 levels in coculture assay with TAT2-treated CD8+ T cells relative to DMSO-treated CD8+ T cells. c, Reduction in p24 values in coculture assay (n = 3) for TAT2-treated CD8+ T cells with and without TI, GRN163L (3 μM). All values given are relative to p24 levels in DMSO-treated coculture. See Materials and Methods for formulas used for calculations. Using a mixed effects analysis, the slope of the line from the estimated equation for the decrease in viral production was significantly different from 0 (β = -0.034, p < 0.0001). Error bars indicate SD. * indicates p-value <0.05 for TAT2 vs DMSO treated cells. ** indicates p value ≤0.05 for TAT2 vs TAT2 plus TI.
Results
Telomerase activity in PBMC and T lymphocytes is increased by TAT2 treatment
We evaluated the effect of short-term exposure to TAT2 on PHA-activated PBMC from 21 individual adult donors, including healthy persons and HIV-infected individuals. In all cases, short-term exposure to TAT2 resulted in enhancement of telomerase activity. A representative TRAP gel for PBMC from an HIV-infected donor (Fig. 1a) illustrates the telomerase enhancement effect of TAT2, and a dose-response experiment shows that this enhancement effect is seen over a wide range of TAT2 concentrations (Fig. 1b). The TAT2 telomerase enhancement effect on PHA-activated PBMC from healthy adults was relatively modest, ranging from 1.5- to 2.5-fold (Fig. 1c), possibly due to the fact that PBMC from younger, healthy individuals already have high endogenous activation-induced telomerase levels. By contrast, the increased telomerase activity induced by TAT2 in PBMC from persons who were chronically infected with HIV-1 but asymptomatic, or from persons who had progressed to AIDS, was more dramatic, ranging from 2.5- to 7-fold (Fig. 1c). This observation was consistent with the previous demonstration of lower telomerase activity in PBMC from HIV-1-infected donors compared with age-matched seronegative controls (25). Indeed, the most dramatic telomerase enhancement by TAT2 was observed in the PBMC that had the lowest levels of endogenous telomerase activity, i.e., from chronically HIV-1-infected and AIDS donors. Similar telomerase enhancement was observed in purified T lymphocytes stimulated with CD2/CD3/CD28-coated beads, where TAT2 treatment not only increased telomerase activity, but prolonged the period poststimulation during which telomerase was active (Fig. 1d). We also observed that exposure to TAT2 enhanced telomerase activity in both the CD28high and CD28low subsets (Fig. 1e). Loss of CD28 expression is associated with chronic antigenic stimulation, and cells that lack CD28 have shorter telomeres and reduced antiviral activity compared with those that still express the CD28 molecule (13, 26).
FIGURE 1.

Telomerase activity in PBMC and T lymphocytes treated with TAT2. a, An 8% acrylamide gel showing TRAP products from PHA (12 μg/ml)-stimulated PBMC of an HIV+ donor. Cells were treated with TAT2 (1 μM) or DMSO (0.1%) every 48 h for 12 days, at which point cells were harvested for TRAP assay. Each lane shows and equivalent of 1 × 104 cells. HI, Heat inactivation; IC, internal control. b, Dose-response curve of PHA-stimulated PBMC from HIV-infected individuals (n = 6), showing mean (and SD) telomerase activity from cultures treated with TAT2 vs DMSO every 48 h for 12 days. c, Fold-increase over DMSO control of telomerase activity from PHA-stimulated PBMC treated with TAT2 (1 μM) for 7–12 days. Error bars indicate SD. d, Kinetics of telomerase activation of T lymphocytes stimulated with CD2/CD3/CD28 Ab-coated beads. Cells were treated with TAT2 (1 μM) or DMSO (0.1%) at multiple time points after stimulation (days 1, 3, 7, 11, 15, and 19) and harvested for TRAP assay 24 h after treatment. Each symbol represents a time point at which telomerase activity was evaluated (n = 3). e, Telomerase activity of separated CD28high and CD28low CD8+ T cells after treatment with TAT2 or DMSO (n = 6). CD8+ T cells were isolated, stimulated with CD2/CD3/CD28 Ab-coated beads, and treated with TAT2 or DMSO every 48 h. After 12 days, CD28high and CD28low cells were isolated from the treated CD8+ population by incubation with anti-CD28-magnetic microbeads and passage through a magnetic separation column, then tested for telomerase activity. Error bars indicate SD. * indicates p value <0.05.
Prior studies on transduction of Ag-specific CD8+ T lymphocytes with hTERT, the catalytic component of human telomerase, suggested that the maintenance of telomerase activity is significantly correlated with both increased proliferative potential and telomere length stabilization (13). We therefore tested the effect of chronic exposure to TAT2 on the proliferation and telomere length of T lymphocytes. Preliminary short-term experiments showed that TAT2 treatment of Ab-stimulated purified CD4+ and CD8+ T lymphocytes from HIV-1-infected individuals resulted in a modest, but statistically significant, increase in the number of population doublings compared with control-treated cells even over a period of 7 days (data not shown). We therefore evaluated the effect of chronic TAT2 exposure in long-term (∼5 mo) cultures of CD8+ T lymphocyte cultures from both uninfected (n = 3) and HIV-1-infected (n = 2) individuals. We observed that repeated exposure to TAT2 resulted in apparent retardation of telomere loss, especially during the first few weeks of culture, and an increased proliferative capacity (Fig. 2). Both control and TAT2 cultures were dependent upon IL-2 and stimulation for growth, with no evidence of immortalization. This is an important observation, as cell immortalization, as seen in some cases of hTERT transduction (27), could present a transformation risk.
FIGURE 2.
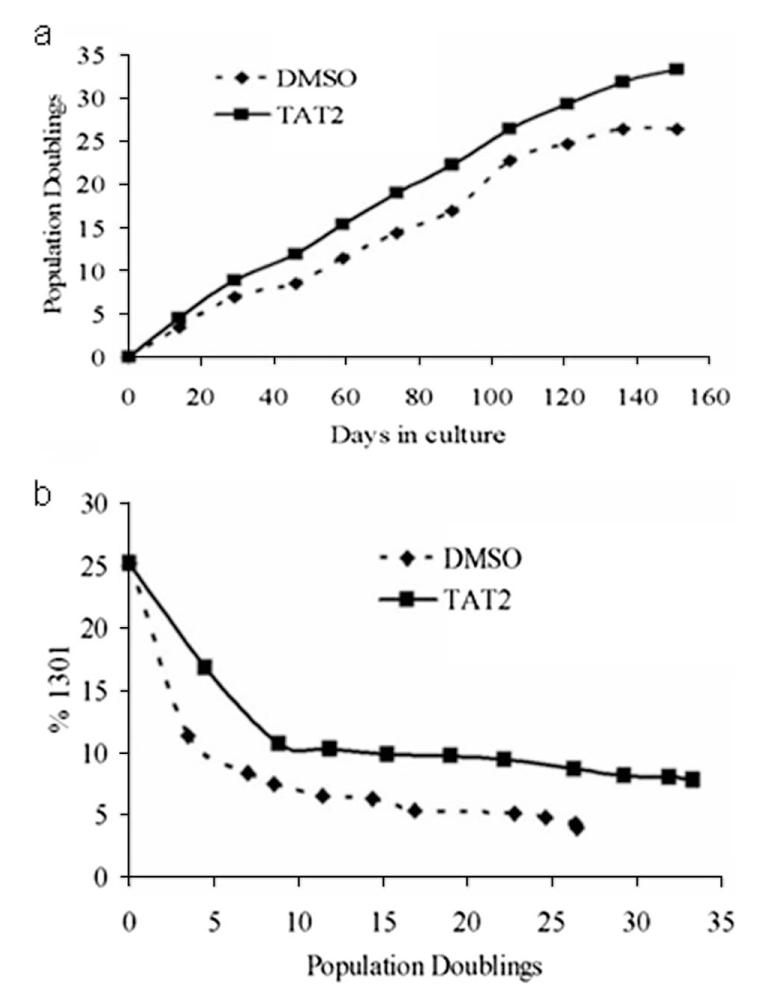
Effects of TAT2 on proliferative capacity and telomere length in T cells. a, Population doublings of CD8+ T cells from a representative HIV+ donor treated with either TAT2 (1 μM) or DMSO (0.1%) every 48–96 h. Symbols indicate stimulation of cells with CD2/3/28 Ab-coated beads, done every 13–17 days. Immediately before addition of beads, cells were counted and total population doublings were calculated. b, Relative telomere length of CD8+ T cells from HIV+ donor treated with TAT2 or DMSO. Symbols indicate time points at which cells were harvested for telomere length assessment. Telomere length data are reported as % telomere length of the acute lymphoblastic T cell line, 1301.
TAT2 treatment activates the MAPK/ERK pathway in CD8+ T lymphocytes
To help elucidate the mechanism involved in the TAT2 effect, we focused on the MAPK/ERK and AKT pathways, based on published studies on telomerase regulation (28, 29) and our own preliminary findings showing that exposure of human neonatal keratinocytes to 10 nM TAT2 for 16 h resulted in a 2.5-fold increase in RSK1, a downstream target of the MAPK pathway (data not shown). Pathway inhibitor experiments on T lymphocytes suggest that activation of telomerase by TAT2 may be mediated through the MAPK/ERK pathway and not the AKT pathway. AKT inhibitors had little effect on the ability of TAT2 to increase telomerase activity, whereas the MAPK and ERK inhibitors abolished the telomerase enhancement by TAT2 (Fig. 3a). These results were confirmed in an ELISA, where both forms of phosphorylated-ERK (p-ERK1/2) were increased in the TAT2-treated cells within 30 to 60 min (Fig. 3b). Consistent with findings in epidermal growth factor-treated fibroblasts, where ERK activation also results in increased hTERT transcription (30), our data suggest that TAT2 increased hTERT transcript levels in “resting” CD8+ T lymphocytes after 12–24 h of treatment, as measured by quantitative real-time PCR (Supplemental Fig. 1).5 We observed that the greatest TAT2-mediated enhancement of both telomerase activity and hTERT transcript occurs in cells that were no longer rapidly proliferating in response to antigenic or receptor-mediated stimulation.
FIGURE 3.
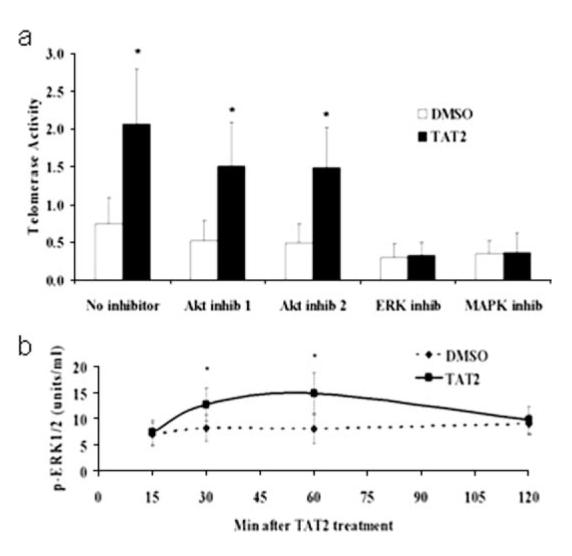
Mechanism of action of TAT2 in T cells. a, Effect of inhibiting cell signaling proteins AKT, ERK1/2, and MEK on telomerase activity in cells treated with TAT2 or DMSO (n = 3). T cells from HIV-infected individuals (n = 3) were stimulated with CD2/CD3/CD28 Ab-coated beads for 7 days, followed by a 2-h exposure to one of the pathway inhibitors, then a 24-h treatment with either TAT2 or DMSO, as described in detail in the Materials and Methods section. Telomerase activity was then determined by TRAP. b, Amount of phospho-ERK (p-ERK) in T cells treated with TAT2 or DMSO, as detected by ELISA. Twelve days after stimulation of T cells from HIV-infected donors (n = 3) with CD3/CD28 Ab-coated beads, TAT2 (1 μM) or DMSO (0.1%) was added and cells were harvested at various times after this treatment (15, 30, 60, and 120 min.); the amount of p-ERK1/2 was determined using the ERK1/2 Phospho-Specific Elisa kit a-c, error bars indicate SD. * indicates p value <0.05.
TAT2 increases the antiviral functions of CD8+ T cells from HIV-1-infected donors
CD8+ T lymphocytes play a crucial role in controlling HIV-1 infection, both during the acute and chronic phases (31, 32), by killing infected cells and secreting antiviral cytokines that can suppress HIV-1 replication (33, 34). Transduction of hTERT into HIV-1-specific CD8+ T lymphocytes from infected persons has been shown to enhance T cell antiviral effector functions (13). To determine whether the telomerase enhancement effect of TAT2 on T cells from HIV-infected persons was also associated with increased functional activities, we first evaluated its effect on HIV-1-specific release of IFN-γ. CD8+ T lymphocytes from HIV-infected persons who had been previously mapped for peptide reactivity were cultured with TAT2, and IFN-γ secretion in response to HIV-1 peptides was evaluated by ELISPOT assays. For the two representative donors shown in Fig. 4a, TAT2-treatement caused a 1.5- to 17.5-fold increase in the number of IFN-γ-producing cells (Fig. 4b). Ag-specific release of the β-chemokines MIP-1α, MIP-1β, and RANTES, which have been shown to exert anti-HIV-1 activity against some strains in vitro (35), was also evaluated. Exposure to TAT2 significantly enhanced HIV-1-specific secretion of all three chemokines (Fig. 4c). Importantly, this increase was telomerase-dependent; addition of a telomerase-specific inhibitor (GRN163L, see Materials and Methods) at a concentration sufficient to inhibit over 90% of the telomerase activity in PBMCs abrogated the effect of TAT2 (Fig. 4c). Finally, treatment of CD8+ T lymphocytes with TAT2 resulted in a significant increase in their ability to suppress HIV-1 replication in autologous CD4+ T cells. Across multiple assays on cells from nine individual HIV-1-infected donors, TAT2 significantly enhanced the CD8+ T lymphocyte suppression of HIV-1 replication (Fig. 5, a and b). Macaque CD8+ T lymphocyte depletion studies demonstrate that CTL are perhaps the major determinant of viremia setpoint (36). Thus, even a 50% increase in virus suppression by TAT2-treated CD8+ T lymphocytes in a short-term 10-day assay would suggest compounded longer term effects leading to about a half log reduction in viral replication in 14.51 days, and a full log decrease in 29.23 days (See Statistical Analysis). Significantly, the TAT2-mediated enhancement of antiviral activity was nearly completely abrogated in the presence of the specific TI (Fig. 5c). Importantly, exposure of purified CD4+ T cells isolated from HIV-1-infected persons to TAT2 did not cause a significant change in the amount of HIV-1 produced by these cells (Supplemental Fig. 2).
FIGURE 4.

Production of IFN-γ and chemokines by TAT2-treated CD8+ T cells from HIV-1-infected donors. a, ELISpot plate showing IFN-γ producing CD8+ T cells from two representative HIV+ donors. Purified CD8+ T cells, stimulated with Ab coated beads and IL-2 and treated with TAT2 or DMSO during a 14-day culture period, were added to ELISPOT plate that contained an various HIV-derived peptides, based on previous mapping (- cont: irrelevant HIV peptide, + cont: CD2/CD28). b, Graphical representation of the average number of IFN-γ-producing cells/106 total CD8+ T cells in each well of the ELISPOT for the two donors shown in a. Error bars (SD) indicate variation in the triplicate wells in each plate. c, Amount of MIP1α, MIP1β, and RANTES produced by stimulated CD8+ T cells that were treated with TAT2 or DMSO (n = 3). CD8+ T cells were isolated and stimulated with CD2/CD3/CE28 Ab-coated beads and treated with TAT2 (1 μM) or DMSO (0.1%). For each condition, one-half of the cells were also treated with the TI, GRN163L (3 μM). After 48 h, supernatants were collected for ELISA. Error bars indicate SD. * indicates p value <0.05, ** indicates p value <0.01 for the comparison of responses in TAT2 vs DMSO treated cells.
Discussion
We have characterized the effects of a small molecule (TAT2), discovered in an empirical screen for telomerase activators, on the function of CD8+ T cells from HIV/AIDS donors. We have shown that TAT2 increases telomerase activity in PBMCs (and, specifically, T lymphocytes) from both healthy and HIV/AIDS donors (n = 21 total), with the relative increase in telomerase activity most prominent in cells from HIV-1-infected individuals (especially those with AIDS). Telomerase activation in CD8+ T cells from HIV/AIDS donors is associated with improvement in proliferative capacity and critical immune effector functions, including Ag-specific increase in the production of cytokines and chemokines, and significant reduction in viral replication in CD8/CD4 coculture experiments. Moreover, these antiviral activities of TAT2-treated cells appear dependent upon telomerase activation, since the effects are abrogated by cotreatment with a potent and specific TI.
Our initial investigations into the mechanism of action of TAT2 suggest that telomerase activation, which typically peaks 24 – 48 h after TAT2 treatment, is preceded by early activation of the MAPK/ERK pathway (within minutes of TAT2 exposure), followed by increased production (or reduced turnover) of hTERT transcripts (peaking around 12 h). Interestingly, the main nuclear effectors of the MAPK pathway include the ETS transcription factors (37), which are known to play a major role in the transcriptional regulation of telomerase activity (38). The MAPK pathway has also been implicated in posttranslational up-regulation of telomerase activity through phosphorylation of hTERT (14), but the effects of TAT2 on this pathway have not been investigated. Thus, although the direct binding partner of TAT2 is not known, our data suggest that TAT2 up-regulates telomerase activity via activation of the MAPK pathway and subsequent increase in hTERT mRNA and/or active phosphorylated forms of hTERT protein.
This investigation expands upon previous hTERT transfection experiments with CD8+ T lymphocytes. Maintenance of telomerase activity in these cells extended their proliferative life span without any discernible alterations in growth characteristics or phenotype (39). Parallel findings were also reported for pathogen-specific CD8+ T lymphocytes (40). Specifically, transfecting HIV-1-specific CD8+ T lymphocytes with hTERT increased a number of antiviral effects (13). Despite these in vitro observations, gene therapy may not be practical in translational studies due to significant safety concerns and the inability to control the level and extent of telomerase activation throughout the body. By contrast, pharmacologic telomerase activators allow greater regulation of dosing and duration of effects. Moreover, in our ex vivo studies, TAT2-treated CD8+ T lymphocytes showed functional enhancement that was comparable to that using hTERT gene transduction. Ongoing work is characterizing the safety and pharmacological activities of TAT2 in animal models.
The ability to enhance telomerase activity and antiviral functions of CD8+ T lymphocytes suggests that this strategy could be useful in treating HIV disease as well as immunodeficiency and increased susceptibility to other viral infections associated with chronic diseases or aging (41- 43). Indeed, short telomeres in white blood cells from elderly persons is correlated with a 7- to 8-fold increased risk of subsequent death from infection (20). Moreover, in HIV-1-infected individuals, shorter telomeres and reduced telomerase activity in virus-specific CD8+ T cells were features associated with more rapid disease progression (15). Arguably, any telomerase activator, despite its beneficial immune effects, might pose a risk of increased proliferation of tumor cells. However, in all ex vivo studies conducted to date, there was no evidence that TAT2 promoted loss of growth control or transformation. For example, TAT2 did not lead to any significant increase in the constitutive telomerase activity in the Jurkat T cell tumor line (Supplemental Fig. 3). Moreover, chronic exposure to TAT2 did not alter the rate of EBV transformation of normal B lymphocytes in cell culture (Supplemental Fig. 4), nor did it increase HIV-1 production in isolated infected CD4+ T lymphocytes from HIV-1-infected donors (Supplemental Fig. 2). In primary T cell cultures, TAT2 enhanced proliferative potential and retarded telomere loss, but the cells were nevertheless dependent on IL-2 and restimulation for growth. Moreover, after ∼30 population doublings, a sharp decline in growth rate in the TAT2-treated cultures was noted, suggesting that the cells were approaching replicative senescence. We speculate that TAT2 might moderately extend the lifespan of “presenescent cells” with low telomerase activity through modest up-regulation of telomerase and extension of the shortest telomeres in individual cells. Importantly, we have observed that the telomerase up-regulation effects are short term and reversible; removal of TAT2 from cells returns telomerase levels to baseline within a few days without any effects on cell viability (Supplemental Fig. 5).
In sum, our studies indicate that telomerase activators, such as TAT2, may constitute a novel class of therapeutic agents which improve immune function at a fundamental, cellular aging level, thus complementing existing drugs for the treatment of HIV/AIDS and a variety of age-related diseases associated with immune deficiency. Indeed, the potential utility of TAT2 in treating HIV-1 infection is underscored by clinical studies documenting the association of high constitutive telomerase activity and longer telomeres in HIV-specific CD8+ T lymphocytes in individuals with greater control over the infection and slower disease progression (15).
Supplementary Material
Footnotes
This work was supported by the following National Institutes of Health Grants: AG023720 and AI060362 (to R.B.E.), AI52031 (to S.R.F.), AI028697 (University of California-Los Angeles Center for AIDS Research), and by Geron, TA Therapeutics Ltd., and the Frank H. Jernigan Foundation.
- hTERT
- human telomerase catalytic component
- TI
- telomerase inhibitor
- TRAP
- telomeric repeat amplification protocol
- HBG
- human β globin
The online version of this article contains supplemental material.
Disclosures: The research conducted at University of California-Los Angeles by R. Effros and colleagues was funded in part by Geron and TA Therapeutics. C. B. Harley is an employee and shareholder of Geron and is working on the therapeutic development of telomerase activators. A. C. Chin was an employee and shareholder of Geron during the time this research was conducted.
References
- 1.Allsopp RC, Vaziri H, Patterson C, Goldstein S, Younglai EV, Futcher AB, Greider CW, Harley CB. Telomere length predicts replicative capacity of human fibroblasts. Proc. Natl. Acad. Sci. USA. 1992;89:10114–10118. doi: 10.1073/pnas.89.21.10114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Demissie S, Levy D, Benjamin EJ, Cupples LA, Gardner JP, Herbert A, Kimura M, Larson MG, Meigs JB, Keaney JF, Aviv A. Insulin resistance, oxidative stress, hypertension, and leukocyte telomere length in men from the Framingham Heart Study. Aging Cell. 2006;5:325–330. doi: 10.1111/j.1474-9726.2006.00224.x. [DOI] [PubMed] [Google Scholar]
- 3.Starr JM, McGurn B, Harris SE, Whalley LJ, Deary IJ, Shiels PG. Association between telomere length and heart disease in a narrow age cohort of older people. Exp. Gerontol. 2007;42:571–573. doi: 10.1016/j.exger.2006.12.002. [DOI] [PubMed] [Google Scholar]
- 4.Valdes AM, Richards JB, Gardner JP, Swaminathan R, Kimura M, Xiaobin L, Aviv A, Spector TD. Telomere length in leukocytes correlates with bone mineral density and is shorter in women with osteoporosis. Osteoporos. Int. 2007;18:1203–1210. doi: 10.1007/s00198-007-0357-5. [DOI] [PubMed] [Google Scholar]
- 5.Effros RB, Cai Z, Linton PJ. CD8 T cells and aging. Crit. Rev. Immunol. 2003;23:45–64. doi: 10.1615/critrevimmunol.v23.i12.30. [DOI] [PubMed] [Google Scholar]
- 6.Blackburn EH. Telomerases. Annu. Rev. Biochem. 1992;61:113–129. doi: 10.1146/annurev.bi.61.070192.000553. [DOI] [PubMed] [Google Scholar]
- 7.Greider CW. Telomeres, telomerase and senescence. BioEssays. 1990;12:363–369. doi: 10.1002/bies.950120803. [DOI] [PubMed] [Google Scholar]
- 8.Hiyama K, Hirai Y, Kyoizumi S, Akiyama M, Hiyama E, Piatyszek MA, Shay JW, Ishioka S, Yamakido M. Activation of telomerase in human lymphocytes and hematopoietic progenitor cells. J. Immunol. 1995;155:3711–3715. [PubMed] [Google Scholar]
- 9.Roth A, Yssel H, Pene J, Chavez EA, Schertzer M, Lansdorp PM, Spits H, Luiten RM. Telomerase levels control the lifespan of human T lymphocytes. Blood. 2003;102:849–857. doi: 10.1182/blood-2002-07-2015. [DOI] [PubMed] [Google Scholar]
- 10.Bodnar AG, Kim NW, Effros RB, Chiu CP. Mechanism of telomerase induction during T cell activation. Exp. Cell Res. 1996;228:58–64. doi: 10.1006/excr.1996.0299. [DOI] [PubMed] [Google Scholar]
- 11.Valenzuela HF, Effros RB. Divergent telomerase and CD28 expression patterns in human CD4 and CD8 T cells following repeated encounters with the same antigenic stimulus. Clin. Immunol. 2002;105:117–125. doi: 10.1006/clim.2002.5271. [DOI] [PubMed] [Google Scholar]
- 12.Nakamura TM, Morin GB, Chapman KB, Weinrich SL, Andrews WH, Lingner J, Harley CB, Cech TR. Telomerase catalytic subunit homologs from fission yeast and human. Science. 1997;277:955–959. doi: 10.1126/science.277.5328.955. [DOI] [PubMed] [Google Scholar]
- 13.Dagarag M, Evazyan T, Rao N, Effros RB. Genetic manipulation of telomerase in HIV-specific CD8+ T cells: enhanced antiviral functions accompany the increased proliferative potential and telomere length stabilization. J. Immunol. 2004;173:6303–6311. doi: 10.4049/jimmunol.173.10.6303. [DOI] [PubMed] [Google Scholar]
- 14.Weng NP, Palmer LD, Levine BL, Lane HC, June CH, Hodes RJ. Tales of tails: regulation of telomere length and telomerase activity during lymphocyte development, differentiation, activation, and aging. Immunol. Rev. 1997;160:43–54. doi: 10.1111/j.1600-065x.1997.tb01026.x. [DOI] [PubMed] [Google Scholar]
- 15.Lichterfeld M, Mou D, Cung TD, Williams KL, Waring MT, Huang J, Pereyra F, Trocha A, Freeman GJ, Rosenberg ES, et al. Telomerase activity of HIV-1-specific CD8+ T cells: constitutive upregulation in controllers and selective increase by blockade of PD ligand 1 in progressors. Blood. 2008 doi: 10.1182/blood-2008-01-135442. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wang P, Jou S, Chen W, Lee S. An improved oxidative cleavage method for large scale preparation of some acid-labile aglycones form glycosides. J. Chin. Chem. Soc. 2002;49:103–106. [Google Scholar]
- 17.Herbert BS, Gellert GC, Hochreiter A, Pongracz K, Wright WE, Zielinska D, Chin AC, Harley CB, Shay JW, Gryaznov SM. Lipid modification of GRN163, an N3′–>P5′ thio-phosphoramidate oligonucleotide, enhances the potency of telomerase inhibition. Oncogene. 2005;24:5262–5268. doi: 10.1038/sj.onc.1208760. [DOI] [PubMed] [Google Scholar]
- 18.Kao HT, Cawthon RM, Delisi LE, Bertisch HC, Ji F, Gordon D, Li P, Benedict MM, Greenberg WM, Porton B. Rapid telomere erosion in schizophrenia. Mol. Psychiatry. 2008;13:118–119. doi: 10.1038/sj.mp.4002105. [DOI] [PubMed] [Google Scholar]
- 19.Kilpatrick RD, Rickabaugh T, Hultin LE, Hultin P, Hausner MA, Detels R, Phair J, Jamieson BD. Homeostasis of the naive CD4+ T cell compartment during aging. J. Immunol. 2008;180:1499–1507. doi: 10.4049/jimmunol.180.3.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cawthon RM, Smith KR, O’Brien E, Sivatchenko A, Kerber RA. Association between telomere length in blood and mortality in people aged 60 years or older. Lancet. 2003;361:393–395. doi: 10.1016/S0140-6736(03)12384-7. [DOI] [PubMed] [Google Scholar]
- 21.Cawthon RM. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002;30:e47. doi: 10.1093/nar/30.10.e47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Njajou OT, Cawthon RM, Damcott CM, Wu SH, Ott S, Garant MJ, Blackburn EH, Mitchell BD, Shuldiner AR, Hsueh WC. Telomere length is paternally inherited and is associated with parental lifespan. Proc. Natl. Acad. Sci. USA. 2007;104:12135–12139. doi: 10.1073/pnas.0702703104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tary-Lehmann M, Hricik DE, Justice AC, Potter NS, Heeger PS. Enzyme-linked immunosorbent assay spot detection of interferon-γ and interleukin 5-producing cells as a predictive marker for renal allograft failure. Transplantation. 1998;66:219–224. doi: 10.1097/00007890-199807270-00014. [DOI] [PubMed] [Google Scholar]
- 24.Fauce SR, Yang OO, Effros RB. Autologous CD4/CD8 coculture assay: a physiologically-relevant composite measure of CD8+ T lymphocyte function in HIV-infected persons. J. Immunol. Methods. 2007;327:75–81. doi: 10.1016/j.jim.2007.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ballon G, Ometto L, Righetti E, Cattelan AM, Masiero S, Zanchetta M, Chieco-Bianchi L, De Rossi A. Human immunodeficiency virus type 1 modulates telomerase activity in peripheral blood lymphocytes. J. Infect. Dis. 2001;183:417–424. doi: 10.1086/318072. [DOI] [PubMed] [Google Scholar]
- 26.Monteiro J, Batliwalla F, Ostrer H, Gregersen PK. Shortened telomeres in clonally expanded CD28-CD8+ T cells imply a replicative history that is distinct from their CD28+CD8+ counterparts. J. Immunol. 1996;156:3587–3590. [PubMed] [Google Scholar]
- 27.Hooijberg E, Ruizendaal JJ, Snijders PJ, Kueter EW, Walboomers JM, Spits H. Immortalization of human CD8+ T cell clones by ectopic expression of telomerase reverse transcriptase. J. Immunol. 2000;165:4239–4245. doi: 10.4049/jimmunol.165.8.4239. [DOI] [PubMed] [Google Scholar]
- 28.Goueli BS, Janknecht R. Upregulation of the catalytic telomerase subunit by the transcription factor ER81 and oncogenic HER2/Neu, Ras, or Raf. Mol. Cell. Biol. 2004;24:25–35. doi: 10.1128/MCB.24.1.25-35.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kang SS, Kwon T, Kwon DY, Do SI. Akt protein kinase enhances human telomerase activity through phosphorylation of telomerase reverse transcriptase subunit. J. Biol. Chem. 1999;274:13085–13090. doi: 10.1074/jbc.274.19.13085. [DOI] [PubMed] [Google Scholar]
- 30.Maida Y, Kyo S, Kanaya T, Wang Z, Yatabe N, Tanaka M, Nakamura M, Ohmichi M, Gotoh N, Murakami S, Inoue M. Direct activation of telomerase by EGF through Ets-mediated transactivation of TERT via MAP kinase signaling pathway. Oncogene. 2002;21:4071–4079. doi: 10.1038/sj.onc.1205509. [DOI] [PubMed] [Google Scholar]
- 31.Borrow P, Lewicki H, Hahn BH, Shaw GM, Oldstone MB. Virus-specific CD8+ cytotoxic T-lymphocyte activity associated with control of viremia in primary human immunodeficiency virus type 1 infection. J. Virol. 1994;68:6103–6110. doi: 10.1128/jvi.68.9.6103-6110.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Koup RA, Safrit JT, Cao Y, Andrews CA, McLeod G, Borkowsky W, Farthing C, Ho DD. Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency virus type 1 syndrome. J. Virol. 1994;68:4650–4655. doi: 10.1128/jvi.68.7.4650-4655.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yang OO, Walker BD. CD8+ cells in human immunodeficiency virus type I pathogenesis: cytolytic and noncytolytic inhibition of viral replication. Adv. Immunol. 1997;66:273–311. doi: 10.1016/s0065-2776(08)60600-8. [DOI] [PubMed] [Google Scholar]
- 34.Garzino-Demo A, Moss RB, Margolick JB, Cleghorn F, Sill A, Blattner WA, Cocchi F, Carlo DJ, DeVico AL, Gallo RC. Spontaneous and antigen-induced production of HIV-inhibitory β-chemokines are associated with AIDS-free status. Proc. Natl. Acad. Sci. USA. 1999;96:11986–11991. doi: 10.1073/pnas.96.21.11986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cocchi F, DeVico AL, Garzino-Demo A, Arya SK, Gallo RC, Lusso P. Identification of RANTES, MIP-1 α, and MIP-1 β as the major HIV-suppressive factors produced by CD8+ T cells. Science. 1995;270:1811–1815. doi: 10.1126/science.270.5243.1811. [DOI] [PubMed] [Google Scholar]
- 36.Jin X, Bauer DE, Tuttleton SE, Lewin S, Gettie A, Blanchard J, Irwin CE, Safrit JT, Mittler J, Weinberger L, et al. Dramatic rise in plasma viremia after CD8+ T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 1999;189:991–998. doi: 10.1084/jem.189.6.991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wasylyk B, Hagman J, Gutierrez-Hartmann A. Ets transcription factors: nuclear effectors of the Ras-MAP-kinase signaling pathway. Trends Biochem. Sci. 1998;23:213–216. doi: 10.1016/s0968-0004(98)01211-0. [DOI] [PubMed] [Google Scholar]
- 38.Dwyer J, Li H, Xu D, Liu JP. Transcriptional regulation of telomerase activity: roles of the Ets transcription factor family. Ann. NY Acad. Sci. 2007;1114:36–47. doi: 10.1196/annals.1396.022. [DOI] [PubMed] [Google Scholar]
- 39.Rufer N, Migliaccio M, Antonchuk J, Humphries RK, Roosnek E, Lansdorp PM. Transfer of the human telomerase reverse transcriptase (TERT) gene into T lymphocytes results in extension of replicative potential. Blood. 2001;98:597–603. doi: 10.1182/blood.v98.3.597. [DOI] [PubMed] [Google Scholar]
- 40.Dagarag M, Ng H, Lubong R, Effros RB, Yang OO. Differential impairment of lytic and cytokine functions in senescent human immunodeficiency virus type 1-specific cytotoxic T lymphocytes. J. Virol. 2003;77:3077–3083. doi: 10.1128/JVI.77.5.3077-3083.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Targonski PV, Jacobson RM, Poland GA. Immunosenescence: role and measurement in influenza vaccine response among the elderly. Vaccine. 2007;25:3066–3069. doi: 10.1016/j.vaccine.2007.01.025. [DOI] [PubMed] [Google Scholar]
- 42.Pawelec G, Gouttefangeas C. T-cell dysregulation caused by chronic antigenic stress: the role of CMV in immunosenescence? Aging Clin. Exp. Res. 2006;18:171–173. doi: 10.1007/BF03327436. [DOI] [PubMed] [Google Scholar]
- 43.Koch S, Solana R, Dela RO, Pawelec G. Human cytomegalovirus infection and T cell immunosenescence: a mini review. Mech. Ageing Dev. 2006;127:538–543. doi: 10.1016/j.mad.2006.01.011. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.