

Abstract
The complexes of the respiratory chain represent mosaics of nuclear and mitochondrially encoded components. The processes by which synthesis and assembly of the various subunits are coordinated remain largely elusive. During evolution, many proteins of the mitochondrial ribosome acquired additional domains pointing at specific properties or functions of the translation machinery in mitochondria. Here, we analyzed the function of Mrpl36, a protein associated with the large subunit of the mitochondrial ribosome. This protein, homologous to the ribosomal protein L31 from bacteria, contains a mitochondria-specific C-terminal domain that is not required for protein synthesis per se; however, its absence decreases stability of Mrpl36. Cells lacking this C-terminal domain can still synthesize proteins, but these translation products fail to be properly assembled into respiratory chain complexes and are rapidly degraded. Surprisingly, overexpression of Mrpl36 seems to even increase the efficiency of mitochondrial translation. Our data suggest that Mrpl36 plays a critical role during translation that determines the rate of respiratory chain assembly. This important function seems to be carried out by a stabilizing activity of Mrpl36 on the interaction between large and small ribosomal subunits, which could influence accuracy of protein synthesis.
INTRODUCTION
The respiratory chain of mitochondria allows eukaryotes to use oxidative phosphorylation (OXPHOS) as a highly efficient way to generate ATP. The complexes driving oxidative phosphorylation are a mosaic of proteins encoded by the nuclear and the mitochondrial DNA. Therefore, assembly of the respiratory chain and the ATPase requires not only expression and import of many nuclear-encoded proteins but also translation of mitochondrially encoded proteins. The mitochondrial genome encodes only a handful of proteins. The effort to express this small number of components in the mitochondrial matrix is immense, and even in simple eukaryotes such as Saccharomyces cerevisiae >250 nuclear encoded proteins are required for mitochondrial gene expression (Sickmann et al., 2003). Dysfunction of any of these components can lead to respiratory deficiency, which is often causative for human diseases (Shoubridge, 2001; DiMauro and Schon, 2008).
The small mitochondrial genome of bakers yeast encodes rRNAs, tRNAs, and eight major polypeptides from which seven are core subunits of the OXPHOS complexes: Cox1, Cox2, Cox3 of cytochrome c oxidase (COX); cytochrome b of the bc1 complex; and Atp6, Atp8, and Atp9 of the F1Fo-ATPase. To avoid misfolding and aggregation of these very hydrophobic proteins, membrane insertion occurs in a cotranslational manner. In contrast to ribosomes of the bacterial and eukaryotic cytosol, mitochondrial ribosomes in bakers yeast are tightly and permanently coupled to the membrane into which the translation products are inserted (Fiori et al., 2003; Jia et al., 2003; Ott et al., 2006). The insertion machinery of the inner membrane is physically attached to the large subunit of the ribosome to facilitate cotranslational insertion. Oxa1 is a central component of this insertion machinery. This multispanning membrane protein contains a C-terminal domain that binds to ribosomes in close proximity to the polypeptide exit tunnel (Jia et al., 2003; Szyrach et al., 2003). Mba1, a peripheral membrane protein, functions as a ribosome receptor that coordinates the positioning of the ribosomal exit tunnel to the insertion site of the inner membrane (Ott et al., 2006). Once the mitochondrially encoded core subunits are inserted into the inner membrane, they associate with nuclear-encoded subunits in a stepwise assembly process that is coordinated by numerous assembly factors (Schulte, 2001; Carr and Winge, 2003; Ackerman and Tzagoloff, 2005; Herrmann and Funes, 2005; Fontanesi et al., 2008).
Presumably due to its specialization on the synthesis of very few proteins, the mitochondrial translation machinery strongly differs from those of the bacterial and eukaryotic cytosol. Also, translational control is carried out by a very distinct system in mitochondria. At least in baker's yeast, a specific membrane-associated translational activator is required for translation of certain mRNAs (Costanzo and Fox, 1990; Naithani et al., 2003; Towpik, 2005). These translational activators might in addition play a role in the spatial and temporal organization of mitochondrial protein synthesis, but their precise function remains unclear. Similarly, the mitochondrial ribosome is very different from its ancestral bacterial type ribosome. Generally, mitochondrial ribosomes underwent major remodeling in the course of evolution and almost doubled their protein mass, in part by addition of several novel ribosomal proteins (Sharma et al., 2003). Conversely, many conserved ribosomal proteins were extended by new domains, implying new functional features to meet the specific requirements of the organelle (Schneider and Ebert, 2004; Smits et al., 2007). However, the relevance of these additional domains is not known.
One protein that acquired an extra domain during evolution is Mrpl36 (previously also called MrpL36), a component of the large ribosomal subunit (Williams et al., 2004). This protein consists of three parts: 1) an N-terminal mitochondrial targeting sequence that is proteolytically removed in the matrix; 2) a conserved domain that is similar to L31, a bacterial protein of unknown function that is loosely associated with the ribosome (Eistetter et al., 1999); and 3) a C-terminal extension domain (CE domain) that is only present in mitochondria but not in bacterial L31 proteins (Williams et al., 2004). Deletion of the entire protein leads to a respiration deficient phenotype. Interestingly, Mrpl36 was initially identified as a high copy suppressor of specific cox2 mutants (Bonnefoy et al., 2001). Genetic analyses suggested two distinct functions of the mature Mrpl36 protein: the L31 domain is necessary for respiratory and translational activity of yeast mitochondria. The CE domain, however, is dispensable for respiratory activity but might play some unknown function in translational regulation. Even when expressed on its own, this domain was found to be sufficient to suppress the defects of specific cox2 mutants (Williams et al., 2004). However, it remains still unclear how overexpression of a single ribosomal protein can help to overcome respiratory deficiency in these suppressors.
In this study, we analyzed the role of Mrpl36 for protein synthesis in mitochondria. We find that deletion of the CE domain does not affect protein synthesis per se. However, the absence of this domain leads to decreased amounts of Mrpl36 and inhibits productive folding and assembly of translation products. This results in rapid degradation of translation products and severe defects in the biogenesis of respiratory chain complexes, in particular of cytochrome c oxidase. In contrast, overexpression of Mrpl36 seems to even increase efficiency of translation. Interestingly, Mrpl36 associates with mitochondrial ribosomes in a dynamic manner and contributes to the interaction between both ribosomal subunits. Our observations are consistent with Mrpl36 being important for productive protein synthesis in mitochondria because it determines whether translation products are integrated into functional complexes or turned over by proteolysis.
MATERIALS AND METHODS
Yeast Strains and Growth Media
All strains used in this study were isogenic to wild-type strain W303-1A, except for the strain Δarg8/cox2::ARG8m (Bonnefoy and Fox, 2000), which was isogenic to D273-10B. For generation of the mrpl36ΔC mutants, the codons for amino acids 116-177 of the MRPL36 gene were replaced by a stop codon followed by either a HIS3 or a kanamycin resistance cassette. Yeast cultures were grown at 30°C in lactate medium, YP (1% yeast extract and 2% peptone) medium supplemented with 2% of either galactose (Gal) or glucose (D), or minimal medium supplemented with 20 μg/ml adenine, histidine, uracil, and tryptophan, and 30 μg/ml leucine and lysine. Mitochondria were isolated as described previously (Daum et al., 1982). Mitochondrial DNA stability was estimated by plating identical amounts of YPD-grown cells on plates containing either 2% glucose or 2% glycerol and comparing the numbers of colonies.
Construction of Plasmids
For overexpression of Mrpl36, Mrpl36-1xHA, Mrpl361-115, and Mrpl361-115-1xHA, the according fragments were cloned into the EcoRI and SalI sites of the 2μ plasmid pYX242 (Novagen, Madison, WI) containing the triosephosphate isomerase (TPI) promoter. To express the CE domain of Mrpl36, we fused the mitochondrial targeting sequence of Oxa11-48 to the N terminus of Mrpl3685-177. For in vitro transcription/translation, the open reading frames of MRPL36 and MRPL3 were cloned into EcoRI and SalI sites of pGEM4 (Promega, Madison, WI).
Labeling of Mitochondrial Translation Products In Vivo
Cells were grown on minimal medium containing either 2% galactose or raffinose. A cell amount corresponding to an OD595 nm of 0.5 was collected, washed twice with growth medium, and incubated at 30°C for 10 min in the same buffer supplemented with 0.15 mg/ml of all amino acids except methionine. After inhibiting cytosolic protein synthesis with 0.15 mg/ml cycloheximide, labeling of mitochondrial proteins was started by addition of 10 μCi of[35S]methionine. Labeling was stopped after the indicated time points by addition of lysis buffer (300 mM NaOH, and 180 mM β-mercaptoethanol). After incubation for 15 min on ice, proteins were precipitated with 12% trichloroacetic acid. Pellets were resolved in sample buffer, subjected to SDS-polyacrylamide gel electrophoresis (PAGE) on 16:0.2% acrylamide/bisacrylamide gels (Preuss et al., 2005) and analyzed by autoradiography and Western blotting.
Fractionation of Mitochondrial Ribosomes on Linear Sucrose Gradients
Isolated mitochondria (1 mg) were lysed for 30 min at 4°C in 1% dodecyl-maltoside, 25 mM KCl, 1 mM phenylmethylsulfonyl fluoride (PMSF), and 20 mM HEPES/KOH, pH 7.4. In some experiments, either 5 mM EDTA and 80 mM puromycin or 5 mM MgCl2 and 1 mg/ml chloramphenicol were included. After a clarifying spin for 15 min at 25,000 × g at 4°C, the extract was layered onto a continuous 4 ml of sucrose gradient (10–30% sucrose in 0.1% dodecyl-maltoside, 25 mM KCl, 1 mM PMSF, and 20 mM HEPES/KOH, pH 7.4) and centrifuged for 1 h at 257,000 × g in an SW60 Ti rotor (Beckman Coulter, Fullerton, CA) at 4°C. Sixteen fractions were collected, and proteins were precipitated with 12% trichloroacetic acid. The resulting pellets were dissolved in sample buffer and separated by SDS-PAGE. Alternatively, a crude ribosomal fraction was obtained by centrifugation through a 1.2 M sucrose cushion. After centrifugation for 1 h at 190,000 × g at 4°C in a TLA100 rotor (Beckman Coulter), the pellets were dissolved in sample buffer, and the supernatant was precipitated with 12% trichloroacetic acid.
Separation of Assembled Respiratory Complexes by Blue Native (BN)-PAGE
Isolated mitochondria were resuspended in solubilization buffer (50 mM NaCl, 50 mM imidazole, 2 mM 6-aminohexanonic acid, and 1 mM EDTA) and lysed for 15 min at 4°C by addition of 1% dodecyl-maltoside or 2.6% digitonin, respectively (Wittig et al., 2006). After a clarifying spin for 15 min at 25,000 × g, the lysate was supplemented with 5% glycerol and subjected to a 4–13% polyacrylamide gradient gel at 4°C. The gel was either stained with Coomassie Brilliant Blue or blotted onto polyvinylidene difluoride (PVDF) membrane and analyzed by Western blotting.
Miscellaneous
Determination of enzyme activities of the respiratory chain, labeling of mitochondrial translation products, and import of precursor proteins in isolated mitochondria has been described previously (Preuss et al., 2005). Immunoelectron microscopy of chemically fixed cells was performed as described previously (Vogel et al., 2006). The antibody against the hemagglutinin (HA)-tag was purchased from Covance Research Products (Princeton, NJ). The antibody against Mrpl36 was obtained by immunizing a rabbit with purified full-length Mrpl36. Western blotting signals were quantified using AIDA software (Raytest, Straubenhardt, Germany).
RESULTS
Both Domains of Mrpl36 Are Critical for Efficient Respiratory Growth
The mature protein Mrpl36 consists of two domains (Figure 1A). The N-terminal region is similar to bacterial L31 proteins (Williams et al., 2004). The C-terminal region is not present in bacteria and represents a C-terminal extension domain (CE domain) with high probability to form a coiled-coil structure. To assess the relevance of this mitochondria-specific domain, yeast mutants were constructed that did not express Mrpl36 or only a truncated version lacking the C-terminal 62-amino acid residues (mrpl36ΔC). Mrpl36-deficient cells were not able to grow on nonfermentable carbon sources such as glycerol (Figure 1B, right). In contrast, the mrpl36ΔC mutant could respire but exhibited a pronounced growth defect. This defect could not be explained by an increased instability of the mitochondrial DNA, as the fraction of cells unable to grow on a nonfermentable carbon source was <5% (data not shown).
Figure 1.

Both domains of Mrpl36 are critical for respiratory growth. (A) Schematic representation of Mrpl36 and mrpl36ΔC. The predicted probabilities to form coiled-coil structures (Lupas, 1997) are expressed in percentage. MTS, mitochondrial targeting signal. (B) Deletion of the CE domain of Mrpl36 reduces growth on nonfermentable carbon sources. The indicated cells were grown in full medium containing galactose to log phase. Serial 10-fold dilutions were spotted on YP plates containing 2% glucose or 2% glycerol, and plates were incubated at 30°C for 2 and 4 d, respectively. (C) The L31 domain is sufficient to support respiratory growth. The indicated strains were grown on synthetic medium containing galactose to log phase. Serial 10-fold dilutions were spotted on synthetic media containing 2% glucose or 2% glycerol, and plates were incubated at 30°C for 3 and 6 d, respectively. (D) Steady-state levels of the different Mrpl36 forms. Lysates of the indicated cells grown on galactose-containing media were prepared and analyzed by Western blotting using antibodies against Tom70 (loading control), the HA-tag, and Mrpl36. Black arrow indicates Mrpl36ΔC, and white arrow indicates Mrpl36ΔC-HA. Asterisk (*), possible degradation product.
The growth defect of mrpl36ΔC cells could either be the consequence of a missing function of the CE domain in the mutant or, more indirectly, a negative effect of the truncation on the L31 domain, or both. To test which domain is required for efficient respiratory growth, we expressed C-terminally HA-tagged versions of Mrpl36ΔC and Mrpl36 from a high copy plasmid by using the constitutive promoter of the TPI in mrpl36ΔC cells. The growth defect of mrpl36ΔC cells on glycerol was corrected when either full-length Mrpl36-HA or Mrpl36ΔC-HA were expressed from the plasmids. In contrast, overexpression of a mitochondrially targeted CE domain did not improve respiratory growth (Figure 1C). From this we conclude that the L31 domain is required, and if overexpressed also sufficient, to support respiratory growth, in line with previous results (Williams et al., 2004). Cells expressing the plasmid-borne Mrpl36 showed in comparison to wild-type cells clearly increased levels of Mrpl36. The deletion of the CE domain leads to a profound destabilization of Mrpl36, so that even upon overexpression from a plasmid, Mrpl36ΔC did not reach the level of the endogenous protein (Figure 1D). In summary, our data indicate that the L31 domain is required for respiratory growth, whereas the CE domain is required for stability of Mrpl36 and possibly other functions.
mrpl36ΔC Cells Exhibit Reduced Translation Efficiency and Stability of Mitochondrially Encoded Proteins
Mrpl36 is a ribosomal protein, suggesting a function of both domains in mitochondrial protein synthesis. We therefore tested whether deletion of the CE domain of Mrpl36 has a direct effect on the synthesis of mitochondrially encoded proteins. Labeling of mitochondrial translation products with [35S]methionine in vivo revealed that both wild-type and mrpl36ΔC cells synthesize all eight mitochondrially encoded proteins (Figure 2A). However, the amount of radiolabeled proteins in mrpl36ΔC was lower than in wild-type cells, in particular for COX subunits. The most severe reduction was found for Cox1, which has been reported not to be efficiently synthesized in mutants defective in COX assembly (Perez-Martinez et al., 2003; Barrientos et al., 2004; Mick et al., 2007; Pierrel et al., 2007). However, overexpression of the translational activator of Cox1, Mss51, did not mitigate the growth defect (data not shown). Cox2, which is synthesized as a precursor protein with an N-terminal leader peptide, was processed to its mature form, indicating proper topogenesis in the mutant. Insertion of mitochondrial translation products into the inner membrane and binding of Oxa1 and Mba1 to the ribosome were not changed in mrpl36ΔC mitochondria (Supplemental Figure 1). Apparently, mitochondrial translation of the mrpl36ΔC cells can produce all the mitochondrially encoded polypeptides. Similarly, all the membrane proteins are inserted efficiently into the inner membrane.
Figure 2.

mrpl36ΔC cells exhibit reduced translation efficiency and stability of mitochondrially encoded proteins. (A) Cells of the indicated strains were grown in synthetic media containing 2% galactose. Cytosolic translation was inhibited by cycloheximide, and mitochondrial translation products were radiolabeled for the indicated times. Labeling was stopped by alkaline lysis and samples were separated by SDS-PAGE. Western blotting against Tom70 and Tim23 served as loading controls. (B) Mitochondrial translation products were labeled in vivo in the indicated strains for 15 min. Next, [35S]methionine and cycloheximide were removed by extensive washing. The cells were further incubated in complete synthetic medium with 2% galactose. After the indicated times, proteins were extracted by alkaline lysis and applied to SDS-PAGE and autoradiography. (C) Signals of the experiment presented in B were densitometrically analyzed. The start value (0 min) was set to 1. Solid line, wild type; dashed line, mrpl36ΔC. (D) Stability of Cox3/Atp6 in wild-type and mrpl36ΔC cells as quantified from four independent experiments. The initial signal (0 min) was set to 1 and compared with the remaining signal after 150 min. A Student's t test was used to reveal significance.
Next, we set out to follow the fate of the newly made polypeptides during prolonged chase periods. To this end, mitochondrially encoded proteins were radiolabeled for 15 min. Then, cycloheximide and [35S]methionine were removed by extensive washing, and protein synthesis was allowed to occur in the presence of unlabeled amino acids. Under those conditions, cells can synthesize all the nuclear-encoded subunits to induce assembly of the labeled mitochondrial translation products into functional OXPHOS complexes. The newly synthesized translation products of wild-type cells were stable for at least 150 min (Figure 2B). In contrast, the labeled polypeptides were rapidly and continuously degraded in the mrpl36ΔC cells (Figure 2B, quantification in C and D). This instability was surprising because translation in the mutant produces less proteins, which should be assembled into functional OXPHOS complexes at least as efficiently as in wild-type cells.
Steady-State Levels of Mitochondrially Encoded Proteins Are Reduced in mrpl36ΔC Cells
The increased degradation of the newly synthesized proteins suggests that respiratory chain complexes are present in lower amounts in mrpl36ΔC mitochondria. To test this, we prepared mitochondria from strains grown under respiratory (lactate) or fermentative (galactose) growth conditions. Under respiratory growth conditions, OXPHOS complexes have to accumulate to allow growth, whereas they are not essential on a fermentable carbon source. Western blotting with antibodies against various mitochondrial proteins revealed unchanged steady-state levels of proteins involved in protein import, export, and mitochondrial translation, among them translocase of the outer membrane (Tom) 70, translocase of the inner membrane (Tim) 23, Oxa1, Mba1, Mrp20 (large ribosomal subunit), and Mrp51 (small ribosomal subunit) (Figure 3A). In contrast, the amounts of Cox2 were severely reduced in mitochondria from mrpl36ΔC cells (Figure 3A, left). Similarly, subunits of the bc1 complex (Cor2, cytochrome b) or the ATPase (Atp4) were significantly reduced, at least under fermentative growth conditions (Figure 3A, right). Apparently, OXPHOS subunits are diminished in mrpl36ΔC cells. If growth absolutely requires respiratory activity, these proteins can accumulate to some extent.
Figure 3.

Steady-state levels of mitochondrially encoded proteins are reduced in mrpl36ΔC cells. (A) Mitochondrially encoded proteins fail to accumulate on media not requiring respiration. Indicated amounts of mitochondria prepared from cultures grown on lactate or galactose were separated by SDS-PAGE and analyzed by Western blotting. (B) Mitochondria prepared from the two strains grown on lactate medium were solubilized in 2.6% digitonin (left) or 1% dodecyl-maltoside (right) and separated by BN-PAGE. The gel was stained with Coomassie (left) or blotted onto PVDF membrane (right) and analyzed by Western blotting. The positions of the supercomplexes are indicated. V2, ATPase dimer; III2/IV2, dimeric bc1 complexes associated with a dimer of COX complexes; III2/IV, dimer of bc1 complexes associated with a monomeric COX complex; V, ATPase monomer. Arrow (left) indicates a band containing nonassembled nuclear encoded subunits of the respiratory chain. (C) The enzyme activities depicted were measured with the mitochondria isolated from lactate grown cultures. Data were normalized to malate dehydrogenase and activities of the wild-type mitochondria were set to 1. (D) Overexpression of Mrpl36ΔC-HA allows the accumulation of mitochondrially encoded proteins under growth conditions not requiring respiration. Lysates of cells grown with galactose as carbon source were prepared and analyzed with Western blotting against Tom70 (loading control), Cox2, and aconitase (Aco1).
The observed decrease in abundance of the OXPHOS complexes in mrpl36ΔC cells lead us to investigate a possible assembly defect in this mutant. Mitochondrial OXPHOS complexes form high molecular assemblies (Arnold et al., 1998; Schagger and Pfeiffer, 2000). In yeast, dimers of bc1 complex associate with either the monomer or the dimer of COX complex giving rise to trimeric (III2/IV) or tetrameric (III2/IV2) supercomplexes (Cruciat et al., 2000). To analyze the organization of the respiratory chain in mrpl36ΔC cells, we lysed mitochondria isolated from cultures grown on lactate media with the mild detergent digitonin and separated the complexes by BN-PAGE. In mitochondria from wild-type cells, mitochondrial supercomplexes migrated with an apparent molecular mass of 800 kDa and were easily detectable in Coomassie-stained gels (Figure 3B, left lane). In contrast, no supercomplexes were observed in mitochondria from the mrpl36ΔC strain. Interestingly, a new complex occurred (Figure 3B, arrow) in the mutant, with an apparent mass of 280 kDa. Mass spectrometric analysis revealed that this complex contains several nuclear encoded OXPHOS subunits: Cor1, Qcr7, and Cor2 from the bc1 complex and Cox4 and Cox6 from the COX complex (Supplemental Figure 2). This complex might represent a nonnative association of nuclear-encoded subunits, which seems to be due to the reduction of mitochondrially encoded subunits in the mutant.
Next, we asked whether the defect in supercomplex formation is due to defects in the assembly of the bc1 complex, the COX complex, or both. Therefore, we assessed abundance of respiratory chain complexes by BN-PAGE by using dodecyl-maltoside as detergent. Mitochondrial supercomplexes dissociate in dodecyl-maltoside and the separated OXPHOS enzymes can be analyzed by Western blotting. Mitochondria from the mrpl36ΔC strain showed a prominent reduction of the COX complex, whereas levels of the bc1 complex remained unchanged (Figure 3B, right). These findings were also reflected in the activities of the OXPHOS complexes (Figure 3C), which revealed severely reduced activities of the COX complex in the mutants. Diminished activities were also observed for the oligomycin-sensitive ATPase and the bc1 complex, but the defects were less pronounced.
This defect to accumulate OXPHOS complexes could either be due to the low level of Mrpl36ΔC in the mrpl36ΔC strain or due to the missing CE domain. Therefore, we asked which domain of Mrpl36 is required to support stable expression of mitochondrially encoded proteins. To test this, we prepared extracts of cells grown on galactose and asked whether mrpl36ΔC cells transformed with plasmids expressing different versions of Mrpl36 can regain the capacity to accumulate mitochondrially encoded proteins under fermentative growth conditions. Wild-type cells accumulated robust amounts of Cox2 (Figure 3D). In contrast, mrpl36ΔC cells with an empty vector or a vector encoding a mitochondrially targeted CE domain of Mrpl36 had strongly reduced steady-state levels of Cox2. Importantly, mrpl36ΔC cells expressing Mrpl36-HA or Mrpl36ΔC-HA from a plasmid were able to accumulate Cox2 at almost wild-type levels. This indicates that the relative amounts of the L31 domain determine the ability of the cells to accumulate mitochondrially encoded proteins.
In summary, we conclude that OXPHOS complexes fail to accumulate in the mrpl36ΔC cells despite the relatively normal production of mitochondrially encoded proteins and the presence of imported nuclear-encoded subunits.
Mitochondrial Translation Generates Unproductive Proteins in mrpl36ΔC Cells
Our results could be interpreted in two ways: First, either the absence of the CE domain or the reduced amounts of the L31 domain might directly influence the assembly of respiratory chain complexes. Alternatively, the efficiency of the translation reaction in mitochondria could be reduced in the mutant, i.e., the primary or secondary structure of the translation products might be incorrect preventing their productive assembly. To differentiate between both possibilities, we analyzed biogenesis and stability of a soluble matrix protein that is normally not synthesized in mitochondria. We reasoned that if the assembly machinery for membrane complexes is defective, folding and activity of this protein should not be impaired. In contrast, if this soluble protein is synthesized but unstable, mitochondrial protein synthesis apparently produces unproductive polypeptides in mrpl36ΔC cells.
To this end, we analyzed the biogenesis of a mitochondrially encoded version of the matrix protein Arg8. Arg8 is an enzyme essential for the biosynthesis of arginine. We made use of an elegant approach that was instrumental for the genetic dissection of mitochondrial gene expression (Steele et al., 1996). In the system we used, arginine prototrophy requires expression of Arg8 from a mitochondrial genome where the recoded open reading frame of ARG8 was inserted in place of the open reading frame of COX2 (cox2::ARG8m) (Bonnefoy and Fox, 2000). In vivo labeling revealed that cox2::ARG8m cells do not synthesize Cox2 but Arg8, with an apparent molecular mass of 46 kDa (Figure 4A). The synthesis of Arg8 was not abolished in mrpl36ΔC with a mitochondrial genome containing cox2::ARG8m: When we labeled mitochondrial translation products in vivo, Arg8 was synthesized efficiently in both cell types (Figure 4B).
Figure 4.

Mitochondrial translation generates unproductive proteins in mrpl36ΔC cells. (A) Labeling profiles of mitochondrial translation in cells with wild-type and cox2::ARG8m mitochondrial genomes. (B) Cells of the indicated strains were grown in synthetic media containing 2% galactose. Cytosolic translation was inhibited by cycloheximide. Mitochondrial translation products were radiolabeled for the indicated times, after which labeling was stopped by alkaline lysis. (C) Steady-state levels of Arg8. Proteins of isolated mitochondria were separated by SDS-PAGE and probed with antibodies against Arg8. Western blotting against Tim23 and Mrp20 was used as loading control. (D) Cells of the indicated strains were spotted in 10-fold dilutions onto media with or without arginine and incubated for 3 d at 30°C.
Next, we assessed the steady-state levels of Arg8 in mitochondria by Western blotting. Surprisingly, Arg8 was hardly detectable in mrpl36ΔC mitochondria and could only be visualized upon very long exposure times (Figure 4C). Consistently, this strain showed a severe growth defect on arginine-deficient media, although no difference in growth was observed in the presence of arginine (Figure 4D). In summary, Arg8 can be efficiently synthesized by mrpl36ΔC ribosomes but this protein, like endogenous translation products, does not accumulate to wild-type levels but rather is rapidly removed by degradation. This led us to the conclusion that not only respiratory growth but also, more generally, growth depending on mitochondrial translation requires an intact Mrpl36.
Ribosomes Are Destabilized in Mitochondria from mrpl36ΔC Cells
The defect in stability of newly synthesized proteins seems to be the direct consequence of either the deletion of the CE domain of Mrpl36 or the reduced amounts of the L31 domain, resulting in unstable polypeptides. Little is known about how mitochondrial protein synthesis is kept accurate, but the observed defect should be linked to the function of Mrpl36 within the context of the mitochondrial ribosome. Therefore, we asked whether the association of mitochondrial ribosomal subunits is similar in mrpl36ΔC and wild-type cells. To this end, mitochondria were allowed to synthesize proteins in the presence of [35S]methionine before they were lysed. The lysates were subjected to density gradient centrifugation. Subsequently, the gradients were fractionated and analyzed using autoradiography and Western blotting. Mitochondrial ribosomes from wild-type cells migrated deep into the gradient and could be separated into two different fractions: a fraction containing mainly the small subunit (SSU, Figure 5A quantification), and a fraction containing assembled ribosomes (ASR, Figure 5A quantification). In contrast, in mitochondria from mrpl36ΔC cells, two different populations were observed. In one fraction, only the small subunit was recovered (SSU, Figure 5B, quantification), whereas in the other fraction almost exclusively the large subunit was detected (LSU, Figure 5B, quantification). However, assembled ribosomes were hardly present in lysates of the mrpl36ΔC mitochondria. From this we conclude that under conditions in which wild-type mitochondrial ribosomes are stable, ribosomes from the mrpl36ΔC mitochondria are destabilized.
Figure 5.

Interaction of ribosomal subunits is destabilized in mrpl36ΔC mitochondria. Translation products of wild-type (A) or mrpl36ΔC (B) mitochondria were labeled with [35S]methionine. (C) Labeling of translation products in wild-type mitochondria was stopped after 20 min by the addition of puromycin. Next, mitochondria were reisolated, lysed with 1% dodecyl-maltoside, and the cleared lysate subjected to velocity centrifugation on a linear sucrose gradient. The gradient was fractionated and analyzed by autoradiography and Western blotting. Quantifications of the signals for Mrp20 and Mrp51 are presented.
Next, we asked whether the migration behavior of mrpl36ΔC ribosomes reflected the migration behavior of inactive ribosomes. Puromycin releases nascent chains from the ribosome, which is known to result in an efficient dissociation of cytosolic ribosomes. Therefore, we incubated wild-type mitochondria with puromycin. However, these puromycin-treated wild-type ribosomes did not disassemble efficiently and migrated in two fractions, one composed of mainly SSU (Figure 5C, quantification) and another containing ASR (Figure 5C, quantification). Only upon treatment with both puromycin and Mg2+-chelating agents were ribosomes efficiently dissociated (Supplemental Figure 3A). In contrast, pretreatment with chloramphenicol did not stabilize mrpl36ΔC ribosomes (Supplemental Figure 3B). Together, we conclude that mitochondrial ribosomes are destabilized in mrpl36ΔC cells.
Overexpression of Mrpl36 Can Suppress a Mutant with a Defect in Accuracy of Mitochondrial Translation
The increased degradation of translation products in mrpl36ΔC mitochondria could be due to defects in their structure. We reasoned that combination of this mutation with mutations that reduce accuracy of translation might further impair respiratory competence of mrpl36ΔC cells. To this end, we analyzed whether the CE domain of Mrpl36 interacts genetically with Guf1, the mitochondrial homologue of bacterial LepA (Bauerschmitt et al., 2008). LepA is a factor that mediates the back translocation of the ribosome to prevent amino acid misincorporation (Qin et al., 2006). Conversely, presence of LepA in an in vitro translation assay increases accuracy (Qin et al., 2006). Cells deficient in either Guf1 or the CE domain of Mrpl36 could grow on a nonfermentable carbon source. In contrast, the combination of both deletions inhibited respiratory growth completely (Figure 6A).
Figure 6.
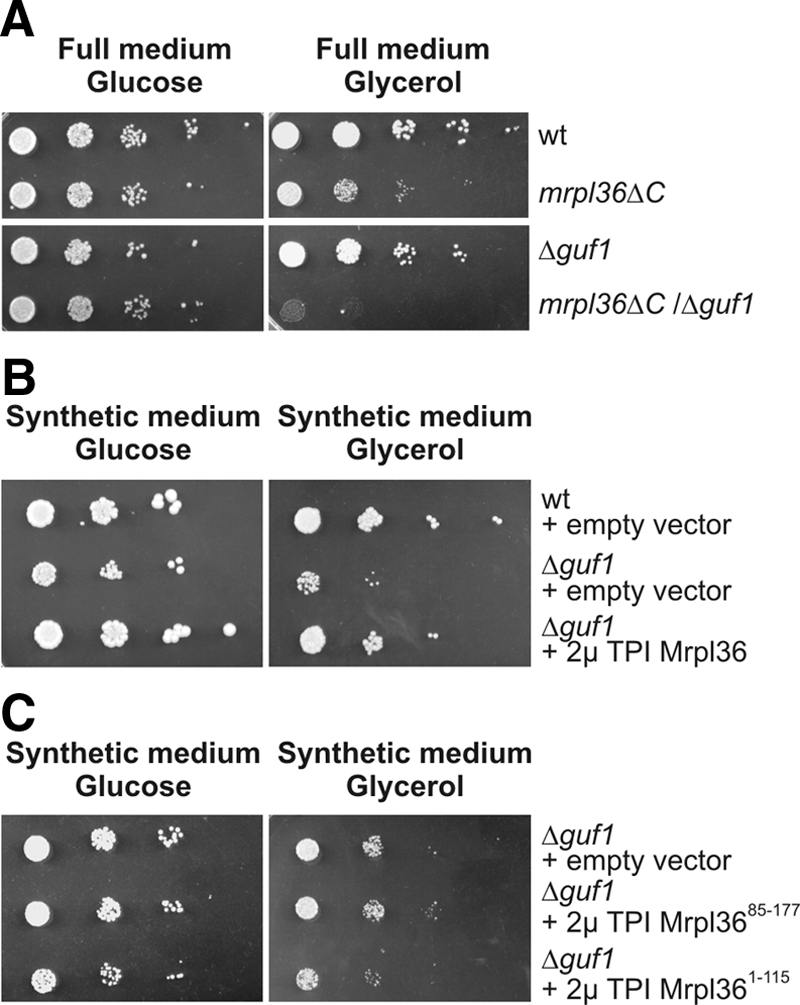
MRPL36 interacts genetically with GUF1. (A) The indicated strains were grown on galactose-containing medium to log phase. Serial 10-fold dilutions were spotted on YP plates containing 2% glucose or 2% glycerol, and plates were incubated at 30°C for 2 and 4 d, respectively. (B) The indicated cells were transformed with 2μ plasmids either containing or not containing MRPL36 under the control of the TPI promoter. The cells were grown on galactose-containing synthetic medium to log phase. Serial 10-fold dilutions were spotted on synthetic media without leucine containing either 2% glucose or 2% glycerol and incubated at 30°C for 3 and 6 d, respectively. (C) Mrpl36ΔC or the CE domain of Mrpl36 cannot suppress the Δguf1 phenotype. Δguf1 cells containing the indicated plasmids were analyzed as described in B.
If Mrpl36 determines more directly the accuracy of the mitochondrial translation system, overexpression of Mrpl36 might overcome the growth defects of Δguf1 cells. To test this, we expressed Mrpl36 from a plasmid, resulting in 3 times more Mrpl36 in the mitochondrial matrix as found in wild-type mitochondria. Δguf1 cells showed a pronounced growth defect on synthetic media with glycerol as carbon source (Figure 6B). Overexpression of Mrpl36 in these cells suppressed this defect and resulted in a respiratory growth similar to wild type.
Next, we asked which domain of Mrpl36 is responsible for the observed suppression of the Δguf1 phenotype. To test this, we overexpressed either Mrpl36ΔC or a mitochondrially targeted CE domain in these cells and tested the growth on media containing either glucose or glycerol (Figure 6C). Neither the L31 domain nor the CE domain, when expressed separately, could revert the growth phenotype of Δguf1 cells. From this, we conclude that only the full-length Mrpl36 can suppress a GUF1 deletion.
Overexpressed Mrpl36 Accumulates in a Soluble Pool in the Matrix, from Which the Protein Can Dynamically Integrate into the Ribosome
How could this positive effect of overexpression of Mrpl36 be explained? We found that Mrpl36 is a factor required to stabilize the interaction between large and small ribosomal subunits. We estimated that Mrpl36 is present in more or less stoichiometric amounts compared with other ribosomal proteins such as Mrp20 (data not shown). Therefore, all translating ribosomes should contain one copy of Mrpl36. Overexpression should only have an impact, if Mrpl36 could dynamically associate with the ribosome and change between a ribosome-bound and a soluble state. Therefore, we overexpressed Mrpl36 in wild-type cells and investigated whether the overexpressed protein was bound quantitatively to the ribosome or accumulated in the matrix. In mitochondria from cells overexpressing Mrpl36 and from wild-type cells, comparable quantities of Mrpl36 were recovered with ribosomes (Figure 7A, quantification). In contrast to wild-type mitochondria, however, a large portion of Mrpl36 was recovered in the soluble fraction when Mrpl36 was overexpressed, in line with previous observations (Williams et al., 2004). Mrpl36 was exclusively recovered in the soluble fraction of mitochondria from a rho0 strain that does not contain assembled mitochondrial ribosomes. This indicates that Mrpl36 can be present in the mitochondrial matrix as a stable soluble protein, in contrast to other ribosomal proteins such as Mrp51.
Figure 7.

Overexpressed Mrpl36 accumulates in a soluble pool in the matrix, from where the protein can dynamically integrate into the ribosome. (A) Mrpl36 is stable as a soluble matrix protein. Indicated mitochondria were lysed with Triton X-100. One-half of the obtained lysate was kept and served as a 100%-total (T); the other half was fractionated by centrifugation through a sucrose cushion. The resulting fractions, supernatant (S) and the pellet (P), together with the totals, were separated by SDS-PAGE and analyzed by Western blotting. (B) Imported Mrpl36 can dynamically integrate into preexisting ribosomes. Radiolabeled precursor of Mrpl36 was incubated with energized mitochondria of the indicated strains. After 30 min, mitochondria were reisolated and either exposed to hypotonic buffers to rupture the outer membrane (Swelling) or to iso-osmotic buffers. Mitochondria were treated with proteinase K (PK) to remove nonimported material; Western blotting against Tom70 was used to assess successful removal of exposed domains. A portion of the import reaction was lysed and separated by centrifugation through a sucrose cushion into a soluble and a ribosomal fraction. (C) Imported Mrpl3 cannot integrate into the ribosome. The experiment was conducted as described in B by using radiolabeled precursor of Mrpl3.
How could an increased pool of soluble Mrpl36 help to improve efficiency of translation? Dynamic exchange of Mrpl36 during the translation reaction could enable soluble Mrpl36 from the matrix to assemble into preexisting ribosomes. To test this, we imported radiolabeled precursor of Mrpl36 into isolated mitochondria. On import, the mitochondrial targeting signal was proteolytically removed by mitochondrial processing peptidase and the mature Mrpl36 was present in a protease-resistant localization. Next, mitochondria were lysed and fractionated by centrifugation through a sucrose cushion. In wild-type mitochondria, Mrpl36 was imported efficiently and was recovered quantitatively with the preexisting ribosome (Figure 7B). In contrast, after import into rho0 mitochondria, Mrpl36 was largely recovered with the soluble fraction. Other mitochondrial ribosomal proteins such as Mrpl3 (Figure 7C), Ppe1, and Mrp20 (data not shown) did not assemble into ribosomes upon import into wild-type mitochondria. From this we conclude that Mrpl36 can dynamically integrate from a soluble pool into the ribosome.
Mrpl36 Dynamically Interacts with the Ribosome In Vivo
The ability of Mrpl36 to assemble into preexisting ribosomes in organello inspired us to test whether changes of interactions of Mrpl36 with mitochondrial ribosomes can be observed more directly. To test this, we determined the distribution of Mrpl36 and Mrp20 relative to the inner membrane using immunoelectron microscopy of chemically fixed cells. In yeast mitochondria, ribosomes tightly interact with the inner membrane and the insertion component Oxa1. Mrp20, a tightly bound ribosomal subunit of the polypeptide exit tunnel, was shown to interact with Oxa1 even under conditions in which translation was inhibited with puromycin (Jia et al., 2003). To investigate the intramitochondrial distribution of Mrp20 or Mrpl36 in vivo, we followed the localization of both proteins. First, antibody concentrations were adjusted to concentrations yielding very low levels of decoration to ensure highly specific detection (Vogel et al., 2006). Next, we measured the distance of the center of each 10-nm gold particle relative to the inner membrane (Figure 8A). In cells from cultures fixed under normal growth conditions, both Mrpl36 and Mrp20 were mainly associated with the inner membrane (Figure 8B). In contrast, when cells were pretreated with puromycin for 30 min before fixation, Mrpl36 shifted partially to a matrix localization (Figure 8A, right and B, top), whereas Mrp20 did not change localization and was found as a membrane-associated protein (Figure 8B, bottom). This indicated that under normal growth conditions, Mrpl36 is bound to ribosomes. When ribosomes are inactive, Mrpl36 at least partially dissociates from the ribosome. This implies that Mrpl36 interacts mainly with actively translating ribosomes. We conclude that Mrpl36 indeed dynamically associates with and dissociates from mitochondrial ribosomes.
Figure 8.

Mrpl36 dynamically interacts with the ribosome in vivo. (A) Wild-type cells were grown in lactate containing media in the absence (left) or presence (right) of puromycin and fixed with glutaraldehyde and formaldehyde. Next, mitochondrial ribosomal proteins were localized by postembedding immunogold labeling of cryosectioned cells. The distance of each gold particle relative to the inner membrane was measured. (B) Statistical analysis of the distribution of gold particles in mitochondria from cells grown in the absence (left) or presence of puromycin (right). Particles were grouped according to their distance to the inner membrane. n indicates the number of gold particles analyzed.
DISCUSSION
In this study, we analyzed the relevance of the CE domain of Mrpl36 for the biogenesis of mitochondrially encoded proteins. Although not required for protein synthesis per se, the presence of this domain is critical for the generation of respiratory chain complexes, in particular for the COX complex. This observation can be explained in two ways: either the CE domain of Mrpl36 plays a dedicated role in membrane insertion, folding, or assembly of translation products; or the CE domain affects assembly more indirectly, for example, by being critical for the production of assembly-competent products. According to the latter hypothesis, the translation products that are synthesized in the absence of the CE domain of Mrpl36 are unproductive and rapidly removed by the mitochondrial quality control system. The second explanation seems much more likely and is consistent with the defects observed in the mrpl36ΔC mutant. The rather general destabilization of endogenous translation products as well as of mitochondrially expressed Arg8 clearly argues against a specific function of Mrpl36 in the insertion or assembly of specific subunits of the respiratory chain.
We find that in the absence of the CE domain, the remaining L31 domain is destabilized. Importantly, overexpression of Mrpl36ΔC can correct the defect in accumulation of mitochondrially encoded proteins in mrpl36ΔC cells. This indicates that the relative amounts of the L31 domain determine the ability of the cells to synthesize productive polypeptides in mitochondria. However, the role of the CE domain is presumably not restricted to its ability to stabilize Mrpl36: overexpression of full-length Mrpl36, but not of either the L31 or the CE domain, can suppress the growth phenotype of Δguf1 cells. Moreover, overexpression of the CE domain was shown to suppress certain COX2 mutations (Williams et al., 2004), which indicates a function different from the stabilization of the L31 domain.
Why is an intact Mrpl36 critical for the synthesis of functional products? Our data strongly suggest that Mrpl36 is required to maintain the proper architecture and dynamics of the assembled ribosome. Despite some exciting insight into the composition and overall structure of mitochondrial ribosomes (Sharma et al., 2003), no high-resolution images of mitochondrial ribosomes exist that could show the position of Mrpl36. However, its bacterial homologue L31 was localized to the interface of large and small ribosomal subunits, close to L5, S13, and the tRNA that is positioned in the E site (Laurberg et al., 2008). The positioning of L31 at such a critical connection points to an important function of L31 for the alignment of the two subunits of the bacterial ribosome and for controlling loading/unloading of the E site. Large conformational changes in both subunits accompany each elongation round (Frank and Agrawal, 2000; Stark et al., 2000; Valle et al., 2003). Interestingly, L31 is located in proximity to L5 and S13, subunits that undergo large structural reconfigurations during translocation (Agirrezabala et al., 2008). It seems likely that Mrpl36 directly influences these dynamics. This is consistent with the observed dissociation of mrpl36ΔC ribosomes. At present, it cannot be discriminated whether the dissociation is caused by the reduced levels of the L31 domain or, more directly, by the absence of the CE domain (Figure 9). It seems conceivable that defects in the coordination of the translation process lead to a diminished accuracy of the products and hence to their increased degradation. This hypothesis is supported by the observed genetic interaction of MRPL36 with GUF1, a fidelity factor of mitochondrial translation (Qin et al., 2006; Bauerschmitt et al., 2008).
Figure 9.
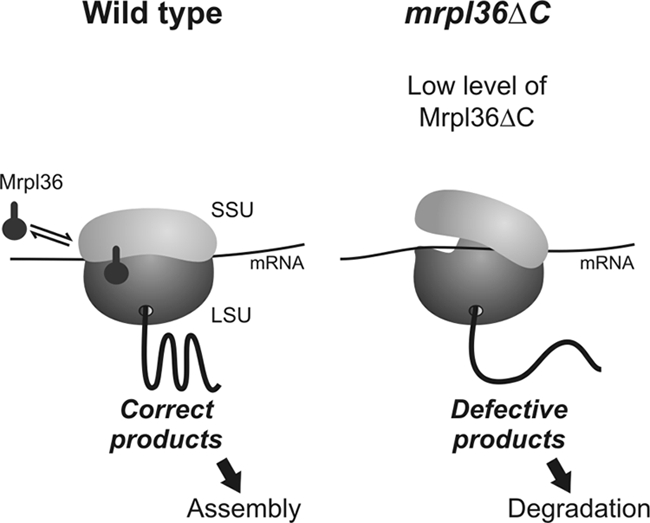
Hypothetical model for the role of Mrpl36 in mitochondrial protein synthesis. Under normal conditions (left), Mrpl36 binds close to the interface of large and small ribosomal subunit. This contact leads to a tight interaction of both subunits and hence allows efficient protein synthesis. In ribosomes from the mrpl36ΔC strain (right), this contact is destabilized, resulting in the production of defective polypeptides. During translation, Mrpl36 dynamically interacts with the ribosome at an as-yet unidentified step.
Mrpl36 was initially found as a multicopy suppressor of mitochondrial DNA mutations (Bonnefoy et al., 2001). This suppression was very puzzling because it was unclear how overexpression of one individual ribosomal protein can have a positive effect on translation. Here, we show that Mrpl36 is not a stable component of the mitochondrial ribosome but rather associates with the ribosome in a puromycin-sensitive manner. This suggests that, in vivo, Mrpl36 is specifically associated with actively translating ribosomes. As pointed out before, increased amounts of Mrpl36 can suppress the growth defect of certain cox2 alleles (Williams et al., 2004). In these mutants, a hairpin structure in the coding region of the COX2 mRNA is stabilized by additional hydrogen bonds. We speculate that suppression of these mutants by overexpression of Mrpl36 could be explained by a more accurate interaction of ribosomal subunits throughout the elongation process. Hence, ribosomal translocation could occur with increased efficiency and thus could allow the linearization of more stably folded mRNAs. Moreover, the beneficial effect of overexpression of Mrpl36 suggests that the levels of Mrpl36 determine the rate of productive protein synthesis in mitochondria, at least in the mutants analyzed.
The mitochondrial translation machinery produces almost exclusively highly hydrophobic proteins that serve as reaction centers of the OXPHOS machinery. Especially, the assembly of the COX complex, which reduces oxygen to water through a series of highly reactive oxygen intermediates, is highly hazardous for the cell (Poyton, 1998). It therefore seems of vital interest to tightly control production and assembly of COX subunits to prevent accumulation of unassembled or defective COX subunits. Recently, an elegant feedback mechanism was identified that controls the synthesis of Cox1, the membrane-embedded reaction center of the enzyme. This regulation is achieved by the specific binding of Mss51, a translation factor required for Cox1 synthesis, to nonassembled Cox1 (Perez-Martinez et al., 2003; Barrientos et al., 2004; Mick et al., 2007; Pierrel et al., 2007). Thereby, assembly defects lead to a sequestration of Mss51 and to an efficient block of Cox1 synthesis. Our results suggest that Mrpl36 is part of an additional regulatory mechanism, because this ribosomal component is critical for the assembly competence of translation products. Especially, the improved respiratory growth in mutants that overexpress Mrpl36 points to an active and decisive role of Mrpl36 in the biogenesis of respiratory chain complexes. In this function, Mrpl36 obviously cooperates with the mitochondrial quality control system that recognizes defective translation products and mediates their proteolytic breakdown (Tatsuta and Langer, 2008).
Not much is known about how the mitochondrial ribosome functions in detail, mostly because of the lack of an in vitro translation assay. Much more work is required to elucidate the exact molecular functions of the mitochondrial ribosome and its associated proteins. A deep understanding of this fundamental machinery might provide important insight into the organization of OXPHOS assembly in which translation of the mitochondrially encoded subunits is the first critical step.
Supplementary Material
ACKNOWLEDGMENTS
We are grateful to Sabine Knaus and Andrea Trinkaus for excellent technical assistance. We thank Stephan Nussberger, Eli van der Sluis, Jan Riemer, Markus Hildenbeutel, and Steffi Gruschke for stimulating discussions and comments on the manuscript. We are grateful to Tom Fox (Cornell University, Ithaca, NY) for antiserum against Mrp51 and to Natalie Bonnefoy (CNRS, Gif-sur-Yvette, France) for the gift of strain Δarg8/cox2::ARG8m. This work was supported by grants of the Deutsche Forschungsgemeinschaft (SFB530, Teilprojekt C15) and the Stiftung für Innovation in Rheinland-Pfalz (to J.M.H.).
Abbreviations used:
- COX
cytochrome c oxidase
- HA
hemagglutinin
- OXPHOS
oxidative phosphorylation.
Footnotes
This article was published online ahead of print in MBC in Press (http://www.molbiolcell.org/cgi/doi/10.1091/mbc.E08-12-1162) on April 1, 2009.
REFERENCES
- Ackerman S. H., Tzagoloff A. Function, structure, and biogenesis of mitochondrial ATP synthase. Prog. Nucleic Acid Res. Mol. Biol. 2005;80:95–133. doi: 10.1016/S0079-6603(05)80003-0. [DOI] [PubMed] [Google Scholar]
- Agirrezabala X., Lei J., Brunelle J. L., Ortiz-Meoz R. F., Green R., Frank J. Visualization of the hybrid state of tRNA binding promoted by spontaneous ratcheting of the ribosome. Mol. Cell. 2008;32:190–197. doi: 10.1016/j.molcel.2008.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold I., Pfeiffer K., Neupert W., Stuart R. A., Schagger H. Yeast mitochondrial F1F0-ATP synthase exists as a dimer: identification of three dimer-specific subunits. EMBO J. 1998;17:7170–7178. doi: 10.1093/emboj/17.24.7170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrientos A., Zambrano A., Tzagoloff A. Mss51p and Cox14p jointly regulate mitochondrial Cox1p expression in Saccharomyces cerevisiae. EMBO J. 2004;23:3472–3482. doi: 10.1038/sj.emboj.7600358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauerschmitt H., Funes S., Herrmann J. M. The membrane-bound GTPase Guf1 promotes mitochondrial protein synthesis under suboptimal conditions. J. Biol. Chem. 2008;283:17139–17146. doi: 10.1074/jbc.M710037200. [DOI] [PubMed] [Google Scholar]
- Bonnefoy N., Bsat N., Fox T. D. Mitochondrial translation of Saccharomyces cerevisiae COX2 mRNA is controlled by the nucleotide sequence specifying the pre-Cox2p leader peptide. Mol. Cell. Biol. 2001;21:2359–2372. doi: 10.1128/MCB.21.7.2359-2372.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnefoy N., Fox T. D. In vivo analysis of mutated initiation codons in the mitochondrial COX2 gene of Saccharomyces cerevisiae fused to the reporter gene ARG8m reveals lack of downstream reinitiation. Mol. Gen. Genet. 2000;262:1036–1046. doi: 10.1007/pl00008646. [DOI] [PubMed] [Google Scholar]
- Carr H. S., Winge D. R. Assembly of cytochrome c oxidase within the mitochondrion. Acc. Chem. Res. 2003;36:309–316. doi: 10.1021/ar0200807. [DOI] [PubMed] [Google Scholar]
- Costanzo M. C., Fox T. D. Control of mitochondrial gene expression in Saccharomyces cerevisiae. Annu. Rev. Genet. 1990;24:91–113. doi: 10.1146/annurev.ge.24.120190.000515. [DOI] [PubMed] [Google Scholar]
- Cruciat C. M., Brunner S., Baumann F., Neupert W., Stuart R. A. The cytochrome bc1 and cytochrome c oxidase complexes associate to form a single supracomplex in yeast mitochondria. J. Biol. Chem. 2000;275:18093–18098. doi: 10.1074/jbc.M001901200. [DOI] [PubMed] [Google Scholar]
- Daum G., Bohni P. C., Schatz G. Import of proteins into mitochondria. Cytochrome b2 and cytochrome c peroxidase are located in the intermembrane space of yeast mitochondria. J. Biol. Chem. 1982;257:13028–13033. [PubMed] [Google Scholar]
- DiMauro S., Schon E. A. Mitochondrial disorders in the nervous system. Annu. Rev. Neurosci. 2008;31:91–123. doi: 10.1146/annurev.neuro.30.051606.094302. [DOI] [PubMed] [Google Scholar]
- Eistetter A. J., Butler P. D., Traut R. R., Fanning T. G. Characterization of Escherichia coli 50S ribosomal protein L31. FEMS Microbiol. Lett. 1999;180:345–349. doi: 10.1111/j.1574-6968.1999.tb08816.x. [DOI] [PubMed] [Google Scholar]
- Fiori A., Mason T. L., Fox T. D. Evidence that synthesis of the Saccharomyces cerevisiae mitochondrially encoded ribosomal protein Var1p may be membrane localized. Eukaryot. Cell. 2003;2:651–653. doi: 10.1128/EC.2.3.651-653.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fontanesi F., Soto I. C., Barrientos A. Cytochrome c oxidase biogenesis: new levels of regulation. IUBMB Life. 2008;60:557–568. doi: 10.1002/iub.86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank J., Agrawal R. K. A ratchet-like inter-subunit reorganization of the ribosome during translocation. Nature. 2000;406:318–322. doi: 10.1038/35018597. [DOI] [PubMed] [Google Scholar]
- Herrmann J. M., Funes S. Biogenesis of cytochrome oxidase-sophisticated assembly lines in the mitochondrial inner membrane. Gene. 2005;354:43–52. doi: 10.1016/j.gene.2005.03.017. [DOI] [PubMed] [Google Scholar]
- Jia L., Dienhart M., Schramp M., McCauley M., Hell K., Stuart R. A. Yeast Oxa1 interacts with mitochondrial ribosomes: the importance of the C-terminal region of Oxa1. EMBO J. 2003;22:6438–6447. doi: 10.1093/emboj/cdg624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laurberg M., Asahara H., Korostelev A., Zhu J., Trakhanov S., Noller H. F. Structural basis for translation termination on the 70S ribosome. Nature. 2008;454:852–857. doi: 10.1038/nature07115. [DOI] [PubMed] [Google Scholar]
- Lupas A. Predicting coiled-coil regions in proteins. Curr. Opin. Struct. Biol. 1997;7:388–393. doi: 10.1016/s0959-440x(97)80056-5. [DOI] [PubMed] [Google Scholar]
- Mick D. U., Wagner K., van der Laan M., Frazier A. E., Perschil I., Pawlas M., Meyer H. E., Warscheid B., Rehling P. Shy1 couples Cox1 translational regulation to cytochrome c oxidase assembly. EMBO J. 2007;26:4347–4358. doi: 10.1038/sj.emboj.7601862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naithani S., Saracco S. A., Butler C. A., Fox T. D. Interactions among COX1, COX2, and COX3 mRNA-specific translational activator proteins on the inner surface of the mitochondrial inner membrane of Saccharomyces cerevisiae. Mol. Biol. Cell. 2003;14:324–333. doi: 10.1091/mbc.E02-08-0490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ott M., Prestele M., Bauerschmitt H., Funes S., Bonnefoy N., Herrmann J. M. Mba1, a membrane-associated ribosome receptor in mitochondria. EMBO J. 2006;25:1603–1610. doi: 10.1038/sj.emboj.7601070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez-Martinez X., Broadley S. A., Fox T. D. Mss51p promotes mitochondrial Cox1p synthesis and interacts with newly synthesized Cox1p. EMBO J. 2003;22:5951–5961. doi: 10.1093/emboj/cdg566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierrel F., Bestwick M. L., Cobine P. A., Khalimonchuk O., Cricco J. A., Winge D. R. Coa1 links the Mss51 post-translational function to Cox1 cofactor insertion in cytochrome c oxidase assembly. EMBO J. 2007;26:4335–4346. doi: 10.1038/sj.emboj.7601861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poyton R. O. Assembling a time bomb–cytochrome c oxidase and disease. Nature genetics. 1998;20:316–317. doi: 10.1038/3778. [DOI] [PubMed] [Google Scholar]
- Preuss M., Ott M., Funes S., Luirink J., Herrmann J. M. Evolution of mitochondrial oxa proteins from bacterial YidC. Inherited and acquired functions of a conserved protein insertion machinery. J. Biol. Chem. 2005;280:13004–13011. doi: 10.1074/jbc.M414093200. [DOI] [PubMed] [Google Scholar]
- Qin Y., Polacek N., Vesper O., Staub E., Einfeldt E., Wilson D. N., Nierhaus K. H. The highly conserved LepA is a ribosomal elongation factor that back-translocates the ribosome. Cell. 2006;127:721–733. doi: 10.1016/j.cell.2006.09.037. [DOI] [PubMed] [Google Scholar]
- Schagger H., Pfeiffer K. Supercomplexes in the respiratory chains of yeast and mammalian mitochondria. EMBO J. 2000;19:1777–1783. doi: 10.1093/emboj/19.8.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider A., Ebert D. Covariation of mitochondrial genome size with gene lengths: evidence for gene length reduction during mitochondrial evolution. J. Mol. Evol. 2004;59:90–96. doi: 10.1007/s00239-004-2607-x. [DOI] [PubMed] [Google Scholar]
- Schulte U. Biogenesis of respiratory complex I. J. Bioenerg. Biomembr. 2001;33:205–212. doi: 10.1023/a:1010730919074. [DOI] [PubMed] [Google Scholar]
- Sharma M. R., Koc E. C., Datta P. P., Booth T. M., Spremulli L. L., Agrawal R. K. Structure of the mammalian mitochondrial ribosome reveals an expanded functional role for its component proteins. Cell. 2003;115:97–108. doi: 10.1016/s0092-8674(03)00762-1. [DOI] [PubMed] [Google Scholar]
- Shoubridge E. A. Nuclear genetic defects of oxidative phosphorylation. Hum. Mol. Genet. 2001;10:2277–2284. doi: 10.1093/hmg/10.20.2277. [DOI] [PubMed] [Google Scholar]
- Sickmann A. C., et al. The proteome of Saccharomyces cerevisiae mitochondria. Proc. Natl. Acad. Sci. USA. 2003;100:13207–13212. doi: 10.1073/pnas.2135385100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smits P., Smeitink J. A., van den Heuvel L. P., Huynen M. A., Ettema T. J. Reconstructing the evolution of the mitochondrial ribosomal proteome. Nucleic Acids Res. 2007;35:4686–4703. doi: 10.1093/nar/gkm441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stark H., Rodnina M. V., Wieden H. J., van Heel M., Wintermeyer W. Large-scale movement of elongation factor G and extensive conformational change of the ribosome during translocation. Cell. 2000;100:301–309. doi: 10.1016/s0092-8674(00)80666-2. [DOI] [PubMed] [Google Scholar]
- Steele D. F., Butler C. A., Fox T. D. Expression of a recoded nuclear gene inserted into yeast mitochondrial DNA is limited by mRNA-specific translational activation. Proc. Natl. Acad. Sci. USA. 1996;93:5253–5257. doi: 10.1073/pnas.93.11.5253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szyrach G., Ott M., Bonnefoy N., Neupert W., Herrmann J. M. Ribosome binding to the Oxa1 complex facilitates co-translational protein insertion in mitochondria. EMBO J. 2003;22:6448–6457. doi: 10.1093/emboj/cdg623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatsuta T., Langer T. Quality control of mitochondria: protection against neurodegeneration and ageing. EMBO J. 2008;27:306–314. doi: 10.1038/sj.emboj.7601972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Towpik J. Regulation of mitochondrial translation in yeast. Cell. Mol. Biol. Lett. 2005;10:571–594. [PubMed] [Google Scholar]
- Valle M., Zavialov A., Sengupta J., Rawat U., Ehrenberg M., Frank J. Locking and unlocking of ribosomal motions. Cell. 2003;114:123–134. doi: 10.1016/s0092-8674(03)00476-8. [DOI] [PubMed] [Google Scholar]
- Vogel F., Bornhovd C., Neupert W., Reichert A. S. Dynamic subcompartmentalization of the mitochondrial inner membrane. J. Cell Biol. 2006;175:237–247. doi: 10.1083/jcb.200605138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams E. H., Perez-Martinez X., Fox T. D. MrpL36p, a highly diverged L31 ribosomal protein homolog with additional functional domains in Saccharomyces cerevisiae mitochondria. Genetics. 2004;167:65–75. doi: 10.1534/genetics.167.1.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wittig I., Braun H. P., Schagger H. Blue native PAGE. Nat. Protoc. 2006;1:418–428. doi: 10.1038/nprot.2006.62. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.