

Abstract
Two experiments tested language switching effects with bilingual participants in a priming paradigm with masked primes (duration of 50 ms in Experiment 1 and 100 ms in Experiment 2). Participants had to monitor target words for animal names, and ERPs were recorded to critical (non-animal) words in L1 and L2 primed by unrelated words from the same or the other language. Both experiments revealed language priming (switching) effects that depended on target language. For target words in L1, most of the language switch effect appeared in the N400 ERP component, with L2 primes generating a more negative going wave than L1 primes. For L2 target words, on the other hand, the effects of a language switch appeared mainly in an earlier ERP component (N250) peaking at approximately 250 ms post-target onset, and showing greater negativity following an L1 prime than an L2 prime. This is the first evidence for fast-acting language-switching effects occurring in the absence of overt task switching.
Keywords: Word recognition, Bilingualism, ERPs, Code-switching, Masked priming
1. Introduction
Speakers familiar with more than one language often alternate languages between or even within sentences, particularly in the company of other bilinguals (Scotton & Ury, 1977). It is now widely accepted that such code-switching incurs cognitive costs, such as a slowdown in comprehension or production when stimuli involve mixed languages, particularly when the input language changes unpredictably (Dalrymple-Alford, 1985; Grainger & Beauvillain, 1987; MacNamara & Kushnir, 1972; Meuter & Allport, 1999; Soares & Grosjean, 1984; Thomas & Allport, 2000; von Studnitz & Green, 1997). The present study follows a long tradition of experimental research investigating the precise locus of these switch costs, and the nature of the underlying mechanisms.
One key question concerning the nature of language switch costs is whether these costs are caused by a general task control mechanism, or are the result of processes specific to the control of language activation in bilinguals. In one prominent model of bilingual language comprehension, the Bilingual Interactive Activation (BIA) model (Grainger & Dijkstra, 1992; van Heuven, Dijkstra, & Grainger, 1998), language switch costs are caused by a mechanism controlling the relative activation of lexical representations in each language—so-called “language nodes.” The language nodes hypothesized in the BIA model function similarly to task and attention control mechanisms postulated in general models of cognitive control (e.g., Cohen, Dunbar, & McClelland, 1990). One possibility is that in the course of learning a second language, bilinguals preempt a general mechanism of cognitive control for the specialized purpose of controlling the relative activation of representations in each language. In the original BIA model, the language node mechanism has a direct influence on the relative activation level of lexical representations in each language, contrary to later developments of this model (the BIA+ model, Dijkstra & van Heuven, 2002). According to the original BIA model, processing costs following a language switch are the result of continuing top-down inhibition from the inappropriate language node towards the lexical representations of the target language.
However, perhaps the most popular account of switch costs in the recent literature states that they are almost exclusively the result of executive control factors and thus external to the language system and related instead to how participants control their decisions and responses in a laboratory task (Dijkstra & Van Heuven, 2002; Green, 1998; Thomas & Allport, 2000). In Green’s (1998) inhibitory control (IC) model, the concept of task schemas plays a central role in accounting for switch costs in the comprehension and production of language in bilinguals. Task schemas are part of a general task control system, and are used in the bilingual situation to link the output of lexical processing to a specific behavioral response. They are mutually inhibitory such that after using task schema A, it is harder to apply task schema B than to repeat task schema A. Since producing language always requires a behavioral response, the IC model has found interesting applications in this area (see e.g., Finkbeiner, Gollan, & Caramazza, 2006). However, outside the laboratory, a switch in the language to be comprehended does not involve changing task schemas. Surprisingly, this has not been seen as a problem, and much of the focus of laboratory research on code-switching in recent years has used situations involving a different response associated with each language (e.g., respond “yes” if the word is in a specific language and “no” otherwise).
These two accounts of switch costs are not necessarily in conflict, and indeed can be seen as complementing each other (Dijkstra, 1998; Green, 1998). The BIA model (Grainger & Dijkstra, 1992) could incorporate more complex task control mechanisms (as proposed by Dijkstra & Van Heuven, 2002, in their BIA+ model), and the IC model (Green, 1998) does allow top-down inhibition from a language-specific task schema to all lemma representations in the irrelevant language—a mechanism used to control for language activation in production. The main difference in the two approaches is the relative importance assigned to modulation of lexical activation in accounting for switch costs in comprehension. The present study aims to provide further evidence for such a mechanism by using a paradigm that minimizes any possible extra-lexical influences.
Code-switching costs have consistently been found in behavioral studies of word comprehension and production in bilinguals (e.g., Costa & Santesteban, 2004; Grainger & Beauvillain, 1987; Meuter & Allport, 1999; Soares & Grosjean, 1984; Thomas & Allport, 2000; von Studnitz & Green, 1997). A key result in the comprehension literature was reported by Grainger and Beauvillain (1987). These authors found switch costs in a generalized lexical decision task (participants responded “word” if the target was a word regardless of language, and “nonword” otherwise). In this task, therefore, the language affliation of the stimulus has no consequences for the required response. Such language information could therefore, in principle, be ignored, but Grainger and Beauvillain demonstrated that this information is not ignored. Lexical decision times to words in one language were slower when the preceding trial was in a different language, compared with a preceding stimulus from the same language; switch costs were also larger for L2 targets compared with L1 targets. This basic result was later replicated by von Studnitz and Green (1997) as well as Thomas and Allport (2000). Thomas and Allport also demonstrated that the switch-cost was not influenced by the presence of language-specific orthographic cues in target words (e.g., the WH in the word WHEAT for an English-French bilingual).
This latter finding led Thomas and Allport to prefer a task-schema account of the switch costs they observed, since the BIA model predicted some modulation of switch costs by language-specific cues. However, the major cause of switch costs in the BIA model is the state of activation of language nodes when the target word appears, and this is uniquely determined by the language of the directly preceding word. Grainger and Beauvillain (1987) suggested that the presence of language-specific orthographic cues in target words might help re-adjust language node activation and modulate observed switch costs. The results of Thomas and Allport (2000) show that this is not the case, although more recent research has shown that switch-costs are diminished in the presence of language-specific letters in Greek-English bilinguals (Orfanidou & Sumner, 2005). In any case, the observation of switch costs in generalized lexical decision is strong evidence against an exclusively taskschema account of the phenomenon.
One ERP study has provided preliminary evidence that word-level activation is directly affected by a language switch (Alvarez, Holcomb, & Grainger, 2003). These authors measured ERPs to single words while bilingual participants silently read for meaning (generating a semantic categorization response on non-critical trials). The results showed that the amplitude of the N400 ERP component was significantly modulated by the language of the word on the immediately preceding trial, and particularly when switching from L1 to L2. Thus, N400 amplitude was significantly greater to the same L2 word when that word was preceded by an unrelated L1 word compared with an unrelated L2 word. If the N400 component is sensitive to lexical-semantic processing as proposed by Grainger and Holcomb (in press), then this result suggests that language switches have a direct consequence at the level of lexical-semantic processing of word stimuli.
Functional brain imaging techniques have also been applied to the study of language-switching in bilinguals. A number of such studies have examined the role of executive control function in language-switching during language production. Hernandez, Martinez, and Kohnert (2000) asked bilinguals to name pictures in Spanish in one block and English in another block and compared these conditions to a mixed block condition. The language to be used was indicated by a cue presented immediately prior to the to-be-named picture. The mixed-language condition provoked increased activation in the dorsolateral prefrontal cortex relative to single-language conditions. Hernandez, Dapretto, Mazziotta, and Bookheimer (2001) confirmed this finding and suggested that this increased activation in the DLPFC attenuates interference from the alternation of suppressing and enhancing two languages. Furthermore, using event-related fMRI, Wang, Xue, Chen, and Dong (2006) found that Chinese-English bilinguals performing a cued picture-naming task produced increased activation in forward switching (L1-L2), but not backward switching (L2-L1) in several brain regions typically associated with executive control (i.e., bilateral frontal cortices and left ACC). Wang et al. suggest that this indicates that neural correlates of language switching may differ as a function of switch direction. However this result could also suggest that executive function plays a larger role in forward than backward language-switching.
This evidence for a primary role of executive control processes in language switching in production is perhaps not that surprising given the nature of the task (cue-induced language selection). However, evidence from one recent fMRI study examining language-switching during comprehension points to a critical role played by the left caudate, specifically for language control (Crinion et al., 2006). Therefore, although general executive control mechanisms might play a role in selecting the appropriate language for output in a cued picture naming task, mechanisms specific to the control of language activation (such as the language nodes of the BIA-model) are also likely to play a role. The present study seeks further evidence for modulation of non-target language activation in a situation where no direct executive control mechanisms are necessary and in conditions where there use is likely to be highly limited (masked priming).
The present experiments were therefore designed to provide further evidence that switch costs in comprehension are driven by modulation of the activation level of lexical representations. To do so we combined ERP recordings with the masked priming technique (Holcomb & Grainger, 2006). Although ERPs are of limited use in providing the exact location of neural sources of specific language and cognitive processes (i.e., they have poor spatial resolution), one strength of ERPs is their high temporal resolution which is in the range of milliseconds. In Holcomb and Grainger’s study, they put this strength to use by examining ERPs to target words that consisted either of full repetitions of the prime, partial repetitions of the prime, or words unrelated to the prime. Priming effects were found in four ERP components effects peaking at various points in time between 125 and 550 ms post-target onset. Of relevance here are their observations on two ERP negativities. The first negativity peaked near 250 ms and showed a progression in both amplitude and latency, with the largest and earliest effect for targets that shared no letters with their primes and the smallest and latest peak latency for full repetition trials, and an intermediate effect in both amplitude and latency for targets that differed by one letter from their primes. This was tentatively identified as an N250 effect and hypothesized to represent the activity of a system processing letters into ordered letter combinations, which are then used to generate sub-lexical phonological representations (Grainger, Kiyonaga, & Holcomb, 2006). The N250 could reflect the process of integrating information across such prelexical codes and whole-word orthographic and phonological representations (Grainger and Holcomb, in press). The second negativity peaked near 450 ms and was greatest to unrelated targets, intermediate to partial repetitions and smallest for full repetitions. This effect was argued to be on the classic N400 and its sensitivity to repetition priming was explained as reflecting a form-meaning interface in which activity between the lexical form and its semantic representation (initiated by the prime) can facilitate or inhibit later activations of these representations (activated by the target). This differs some-what from the usual explanation of the N400 semantic priming effect, in which interactions between semantic representations mediate the effect, rather than those between lexical form and semantic representations.
The BIA model predicts that switch costs should be observable from briefly presented prime stimuli, assuming that these stimuli automatically activate the appropriate language node. Furthermore, these language switching effects should be observable in relatively early phases of processing. Since these early phases can be obscured by later ones in behavioral data, ERPs are a particularly apt method for investigating when during word processing can one first observe the influence of a language switch. Additionally, since previous work on code-switching has involved overt changes in language, it is possible that extra-linguistic strategic processes may have contaminated the effects. The masked priming technique and brief prime duration used in Experiment 1 of the present study circumvents this problem, since participants are not consciously aware of any change in language across prime and target words.
In Experiments 1 and 2, target words were preceded by totally unrelated prime words that were either from the same language (within-language/non-switch trials) or the other language (between-language/switch trials). All primes were pattern masked; in Experiment 1 primes were presented for 50 ms, and in Experiment 2 the duration was extended to 100 ms. If language switching costs in comprehension are exclusively due to executive control processes then we do not expect to observe such costs in the testing conditions of Experiment 1 with masked primes, but such costs may appear in Experiment 2 with an extended prime duration. Any observation of language switching costs in Experiment 1 can be taken as strong evidence for automatic modulation of lexical (and perhaps prelexical) representations by knowledge of which language a stimulus word belongs to.
2. Experiment 1 (50 ms prime duration)
2.1. Methods
2.1.1. Participants
Twenty participants (12 women, 19 to 23 years of age, mean age 21) were recruited from the University of Provence and compensated for their time. All were right-handed native speakers of French who had completed at least their high school diploma in English, and some had studied English at University. All participants had normal or corrected-to-normal visual acuity with no history of neurological insult or language disability. No participant had lived in an immersive English environment for more than two months, and participants who had learned a second language outside of school before puberty were excluded.
Participants’ knowledge of and skill in English was measured by a questionnaire about their language abilities and history enumerating their experience (academic or other) with the French and English languages, and in which they evaluated their reading and speaking skills in both languages (see Table 1 for results). These questionnaires were used to screen participants for the desired level of fluency. Participants indicated that they used English for an average of 5.4% of their day.
Table 1.
Participants’ self-ratings (mean, standard deviation, and range) of their reading, speaking, and oral comprehension skills in L1 and L2, on a seven-point scale in which seven represents perfect competence, and 0 represents no competence
| Reading | Speaking | Comprehension | |
|---|---|---|---|
| French (L1) | 6.8 SD 0.55 Range 5-7 | 6.9 SD 0.31 Range 6-7 | 6.8 SD 0.41 Range 6-7 |
| English (L2) | 3.7 SD 0.99 Range 2-6 | 3.1 SD 1.07 Range 2-5 | 3.1 SD 1.00 Range 2-4 |
2.1.2. Apparatus
Electroencephalograms were collected using 32-channel caps (Electro-cap International), in which tin electrodes were arranged using the standard International 10-20 system. Twelve electrodes were arranged in mirror images on each hemisphere: FP1/2, F3/4, F7/8, FC1/2, FC5/6, C3/4, T3/4, CP1/2, CP5/6, T5/6, P3/4, and O1/2 (see Fig. 1). Five electrodes were located at midline: FPz, Fz, Cz, Pz, and Oz. One electrode was placed beside the left eye and one below the right eye to monitor for blinks and saccades. Two electrodes were placed behind the ears on the mastoid bone; the left mastoid site (A1) was used as an online reference for all electrodes, and the right mastoid site (A2) was also recorded to evaluate differential mastoid activity. An electrolyte gel was used in all electrodes in order to improve contact between the skin and the electrode. Impedance was measured at less than 2 kΩ for scalp and mastoid electrode sites and less than 10 kΩ for both eye channels.
Fig. 1.

Thrity-two-channel electrode montage.
The EEG was amplified using an SA Bioamplifier (SA Instruments, San Diego Ca.) operating on a bandpass of .01 and 40 Hz. The digitizing computer continuously sampled the EEG at a rate of 200 Hz, while the stimulus computer simultaneously issued stimuli to the participant’s monitor and transmitted codes indicated types of stimuli or behavior to the digitizing computer.
2.1.3. Design and stimuli
The stimuli for the study consisted of prime-target pairs formed of completely unrelated words that were either in the same language or from different languages. All words were between four and six letters, and were selected to be as language-specific as possible, avoiding close cognates in order to minimize confusion over which language was being presented within a given trial. Words with accents were also excluded, to prevent the use of non-lexical cues. Translations of words used in one language were specifically excluded from the set of possible stimuli in the other language. Target language (English or French) was crossed with trial type (within-language, between-language) to create four experimental conditions. These four trial types were organized into two blocks—one in which the participant saw only English target words (including French prime-English target trials and English prime-English target trials), and one in which the participant saw only French target words (including English prime-French target trials and French prime-French target trials).
2.1.4. Procedure
In Experiment 1, each trial consisted of a 200 ms forward pattern mask, a 50 ms presentation of the prime followed by a 10 ms backward mask, a 300 ms presentation of the target, and a 2900 ms inter-trial interval (see Fig. 2 for a schema of a typical trial). The inter-trial interval consisted of 900 ms of black space and 2000 ms of a schema of a blinking face, during which participants had been instructed to blink. Primes were displayed in lower case and targets in upper case to avoid perceptual repetition; additionally, the backward mask differed from the forward mask because letter post-masks seem to provide superior masking to hash marks or other non-letter masks.
Fig. 2.
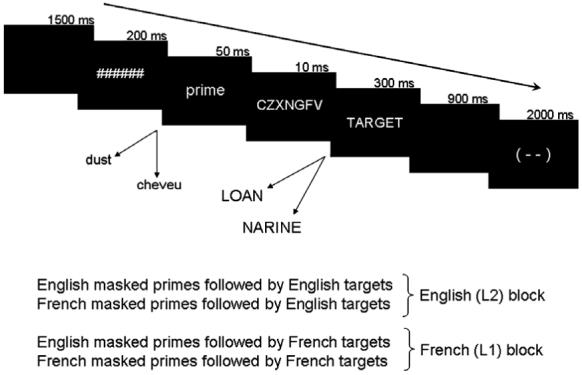
Schema of trial types and timing. Words in the prime position were not repeated in the target position.
To ensure participant attention as well as lexical processing, a go/no-go semantic categorization task was imposed. Participants were asked to press a button when they saw an animal word. In each language block 15% of trials contained an animal word in the appropriate language, half in the prime position and half in the target position. The existence of probes in the prime position provided an estimate of level of visibility of prime stimuli—but since the prime duration was so short, only probes in the target position (7.5%) were responded to.
After completing informed consent, language history, and handedness forms, participants were fitted with an electrode cap and seated in a comfortable armchair in a sound-attenuated room with dimmed lighting. Before each experimental block, participants were presented with a practice block to familiarize them with the trial sequence and the probe task. During the course of the experiment, the stimulus monitor was placed approximately 60 inches in front of the participant’s chair. EEG traces were recorded during experimental block, and the block orders were counterbalanced between participants. There were two pauses within each experimental block, the length of which was determined by the participant. Words were presented centered horizontally and vertically on the participant’s monitor, and each experimental block began when participants pressed a button to indicate their readiness. The experimental blocks typically required 30 to 35 min, depending on the length of the pauses taken by the participant.
2.1.5. Analysis
Multi-factor repeated measures designs were used including language of the target word (L1 vs. L2) and trial type (within-language vs. between-language) as the primary independent variables. In the midline analysis there was a factor of electrode site, and in lateral analyses both electrode site and hemisphere were included as factors; non-midline electrode sites were separated into three columns to assess relatively lateralization of effects within hemispheres. The dependent measure was the EEG signal, which was averaged by condition to create ERPs. ERPs were time-locked to the prime in order to acquire a suffcient pre-target baseline and then adjusted to the presentation of the target (60 ms after the prime in Experiment 1).
Analysis involved both visual inspection and statistical testing. Statistical analyses were conducted by repeated measures analysis of variance (ANOVA) with the Geisser-Greenhouse correction on mean amplitude and peak latency measurements of designated epochs from selected electrode sites. Based on visual analysis of the grand average waveforms and conventional windows for identifying ERP components in masked priming experiments (e.g., Holcomb & Grainger, 2006), two windows were selected: 175-300 ms to detect reactions in the N250 time window and 375-550 ms for reactions in the N400 time window.
2.2. Behavioral results
No participant pressed to probe items (animal names) in the prime position, suggesting that primes were largely invisible in Experiment 1. When probe items were in the target position, the accuracy rate in L1 (French) was 94.1%, and in L2 (English) was 55.89%.
2.3. ERP results
2.3.1. 175-300 ms post-target
ERPs in this epoch were associated with a larger negativity to French compared with English targets (main effect of target language, midline: F(1, 19) = 6.571, p = .019; Column 1: F(1, 19) = 7.907, p = .011; Column 2: F(1,19) = 8.356, p = .0009; Column 3: F(1,19) = 8.387, p = .009). Additionally, target ERPs in between-language trials were more negative-going than target ERPs in within-language trials (main effects of trial-type, midline: F(1, 19) = 9.386, p = .006; Column 1: F(1,19) = 8.489, p = .009; Column 2: F(1,19) = 9.818, p = .005; Column 3: F(1,19) = 11.825, p = .003). Finally, there was an interaction between trial-type, target language and electrode site at the midline, central, and peripheral sites (midline: F(4, 76) = 6.878, p = .003; Column1: F(2,38) = 3.589, p = .037; Column 3: F(4,76) = 4.404, p = .032).
To break down this interaction, follow-up analyses were run separately for English-target trials and French-target trials. There was a main effect of trial type in English-target (L2) trials (midline: F(1,19) = 6.010, p = .024; Column 1: F(1,19) = 7.514, p = .013; Column 2: F(1,19) = 6.422, p = .02; Column 3: F(1,19) = 4.573, p = .046, indicating that target ERPs in between-language trials (i.e., French prime and English target, L1-L2) were more negative-going than within-language trials (i.e., English prime and English target, L2-L2—see Figs. 3 and 5). For the French-target trials, there was a trial type by electrode interaction on midline sites (F(4,76) = 5.628, p = .009), as well as a trial type by hemisphere by electrode site interaction in central sites (Column 1: F(2,38) = 4.075, p = .030). This interaction (see Figs. 4 and 5) was due to a greater negativity on between-language trials (i.e., English primes and French targets, L2-L1) as compared to within-language trials (i.e., French primes and French targets, L1-L1) at anterior sites, yet a greater positivity of between-language trials at posterior sites. These effects were also stronger in the left hemisphere than the right.
Fig. 3.

Results of Experiment 1 (50 ms prime duration) showing within-language trials L2-L2 (full) and between-language trials L1-L2 (dashed) in the English block. (a) Thrity-two-channel ERPs. (b) Blowup of CP1 site.
Fig. 5.

Results of Experiment 1 shown as voltage maps of language switching effect sizes (between-language trials minus within-language trials) in two epochs centered on the N250 and N400 ERP components. The upper panel shows the effects of language switching when targets are in L2 (English), and the lower panel shows the effects for targets in L1 (French).
Fig. 4.

Results of Experiment 1 (50 ms prime duration) showing within-language trials L1-L1 (full) and between-language trials L2-L1 (dashed) in the French block. (a) Thirty-two-channel ERPs. (b) Blowup of CP1 site.
2.3.2. 375-550 ms post-target
Between 375 and 550 ms post-target, there was a significant language by electrode site interaction (midline: F(4,76) = 5.820, p = .006; Column 1: F(2,38) = 8.237, p = .003; Column 2: F(3,57) = 9.546, p = .002; Column 3: F(4,76) = 5.773, p = .006), indicating that ERPs to French targets continued to be more negative-going than English-target trials, particularly at posterior sites. Within the French-target block, language-switching trials (that is, L2 prime followed by an L1 target) were marginally more negative than non-switch trials at central sites; this effect was strongest at the right fronto-central sites (Column 1: F(2,38) = 3.160, p = .059).
2.4. Discussion
The results of Experiment 1 show clear effects of code-switching in a masked priming paradigm. These code-switching effects are mainly present in the epoch of the N250 component (Holcomb & Grainger, 2006), especially for L2 (English) targets, preceded by L1 (French) primes. As can be seen in Fig. 5, this early code-switching effect for L2 targets had a fairly widespread distribution. On the other hand, the code-switching effect found in this same epoch for L1 targets had a more anterior distribution (see Fig. 5). The fact that the N250 code-switching effect was overall in the same direction in L1 and L2 (see Fig. 5) allows us to rule out a prime word frequency account of these effects. That is, the effect of an L1 prime on L2 targets compared with L2 primes could be due to the higher subjective frequency of L1 primes compared with L2 primes. If this were the case, however, we would expect to see exactly the opposite effect of code-switching with L1 targets—i.e., greater negativities with L1 primes than L2 primes. In line with a code-switching interpretation of this effect, in the majority of electrode sites we found greater negativities with L2 primes compared with L1 primes during the processing of L1 targets.
Code-switching effects were weaker overall in the later time window centered on the N400 ERP component. Here the effects were most evident for L1 targets, with greater negativities following an L2 prime than an L1 prime. This again counters a prime word frequency account of the effects we observed.
The early influence of prime word language found in Experiment 1 strongly implies that at least part of the language switching effects found in comprehension are the result of fast-acting and automatic modulation of lexical and possibly prelexical representations. Such modulation of the activation of whole-word orthographic representations (as envisioned in the original BIA model) would be sufficient to affect the processing reflected in the N250 ERP component as well as later components. Grainger and Holcomb (in press) have argued that the N250 reflects integration of information across sublexical representations (i.e., letters and letter clusters) and whole-word representations. Accordingly, any inhibition operating on whole-word representations would therefore influence this process. Thus, the prime word rapidly activates the corresponding language node (this will occur faster for L1 compared with L2 primes) and this language node activation sends inhibition to all lexical representations in the other language, including the upcoming target word representation (van Heuven et al., 1998). This inhibition operating on whole-word representations then affects the integration process across prelexical and lexical representations (N250) and later form-meaning integration processes (N400). Since L1 primes are processed faster than L2 primes, they have an earlier influence on target word processing, hence the stronger code-switching effect on L2 targets in the N250.
3. Experiment 2 (100 ms prime duration)
According to the BIA model, increasing prime duration should increase the size of code-switching effects without producing a qualitative change. The increased prime duration should lead to increased activation of language nodes and, therefore, a stronger influence of the language of prime stimuli. We therefore expect to see stronger code-switching effects in the N400 ERP component, which has been shown to be sensitive to overt language switching in prior research (Alvarez et al., 2003). Increasing prime duration will also allow us to evaluate the influence of conscious processing of language information (i.e., knowing that the prime stimulus is a word in a given language) on code-switching effects.
3.1. Methods
3.1.1. Participants
The 20 participants who participated in Experiment 1 were asked to return for Experiment 2 at least two weeks after their participation in Experiment 1.
3.1.2. Design, stimuli, and apparatus
These were the same as in Experiment 1.
3.1.3. Procedure
This was the same as Experiment 1 except for the extended duration of the prime. Each trial consisted of a 200 ms forward patterned mask, a 100 ms presentation of the prime followed by a 10 ms backward mask, a 300 ms presentation of the target, and a 2900 ms inter-trial interval. Since participants had already participated in Experiment 1, they were not required to complete language history and handedness forms again.
3.1.4. Analysis
As in Experiment 1, multi-factor repeated measures designs were used including language of the target word (L1 vs. L2) and trial type (within-language vs. between-language) as the primary independent variables. The same electrode montage was used, and again ERPs were time-locked to the prime in order to acquire a suffcient pre-target baseline and then adjusted to the presentation of the target (110 ms after the prime in Experiment 2). The data were analyzed applying the same time windows as in Experiment 1.
3.2. Behavioral results
Accuracy rates to probe items (animal names) in the prime position in L1 (French) was 78.67% and in L2 (English) 45.33%. When probe items were in the target position, the accuracy rate in L1 (French) was 94.33%, and in L2 (English) was 62.0%. Performance to probe items in the prime position demonstrates that primes were visible in Experiment 2, compared with Experiment 1.
3.3. ERP results
3.3.1. 175-300 ms post-target
Significant main effects of trial type in the medial three analysis columns (midline: F(1,19) = 10.345, p = .005; Column 1: F(1,19) = 14.721, p = .001; Column 2: F(1,19) = 15.541, p = .001) indicated that between-language trials tended to produce more negative ERPs than within-language trials, in both language blocks. However, examination of Figs. 6 and 7 and the significant language by trial type interaction (midline: F(1,19) = 5.615, p = .029; Column 1: F(1,19) 7.377, p = .014; Column 2: F(1,19) = 8.534, p = .009) suggests this difference was greater for the English-target (L2) trials. This observation was confirmed in follow-up analyses run separately for the English and French blocks. While between-language targets were significantly more negative-going than within language trials for the English-targets (main effect of trial-type English, midline: F(1,19) = 10.937, p = .004; Column 1: F(1,19) = 16.501, p = .001; Column 2: F(1,19) = 18.022, p < .001; Column 3: F(1,19) = 15.423, p = .001; trial type by hemisphere interaction, C2: F(1,19) = 7.990, p = .011; C3: F(1,19) = 5.232, p = .032; trial type by hemisphere by electrode site interaction, C3: F(4,76) = 4.772, p = .002—see Fig. 6a), there were no significant differences for between and within language trials for French targets (all ps > .4 - Fig. 7a).
Fig. 6.

Results of Experiment 2 (100 ms prime duration) showing within-language trials L2-L2 (full) and between-language trials L1-L2 (dashed) in the English block. (a) Thirty-two-channel ERPs. (b) Blowup of CP1 site.
Fig. 7.

Results of Experiment 2 (100 ms prime duration) showing within-language trials L1-L1 (full) and between-language trials L2-L1 (dashed) in the French block. (a) Thirty-two-channel ERPs. (b) Blowup of CP1 site.
3.3.2. 375-550 ms post-target
Between 375 and 550 ms post-target onset, ERPs to targets in the French block were more negative-going than ERPs to targets in the English block (main effect of language, midline: F(1,19) = 4.877, p = .040; Column 2: F(1,19) = 5.743, p = .027; Column 3: F(1,19) = 6.016, p = .024), and between-language trials were more negative than within-language trials (main effect of trial-type, midline (marginal): F(1,19) = 4.265, p = .053; Column 1: F(1,19) = 5.342, p = .032; Column 2: F(1,19) = 4.601, p = .045; Column 3: F(1,19) = 5.200, p = .034). There were also language by trial-type by electrode site interactions present at all four analysis columns (Midline: F(4,76) = 9.242, p = .001; Column 1: F(2,38) = 9.680, p = .003; Column 2: F(3,57) = 12.290, p = .001; Column 3: F(4,76) = 7.677, p = .005. Separate follow-up analyses for the two target languages were again used to break down these interactions. For English targets (Fig. 6a), significant trial-type by electrode site interactions were found for all analysis columns (midline: F(4,76) = 6.547, p = .008; Column 1: F(2,38) = 10.455, p = .002; Column 2: F(3,57) = 8.944, p = .003; Column 3: F(4,76) = 4.585, p = .035), indicating that between-language trials (French primes and English targets, L1-L2) were more negative than within-language trials (English primes and English targets, L2-L2) at anterior sites. This effect either reversed or became equivalent at posterior sites. The effects obtained with French targets were larger overall, but the trial-type by electrode site interaction was significant only at the midline sites F(4,76) = 4.672, p = .019. Moreover, the pattern of the effect was different for French targets: between-language trials (L2-L1) were more negative-going than within-language trials (L1-L1) at posterior sites, while within-language trials were more negative-going than between-language trials at anterior sites (Fig. 8).
Fig. 8.
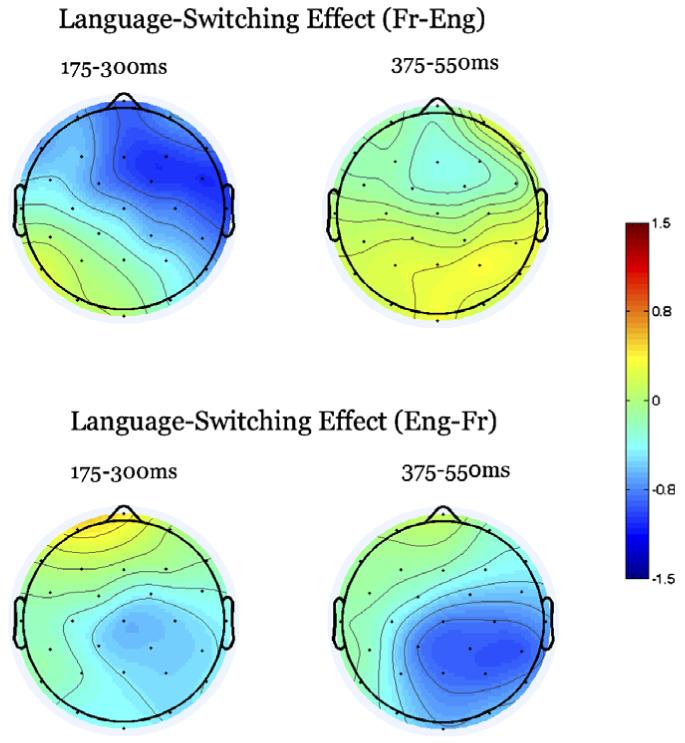
Results of Experiment 2 shown as voltage maps of language switching effect sizes (between-language trials minus within-language trials) in two epochs centered on the N250 and N400 ERP components. The upper panel shows the effects of language switching when targets are in L2 (English), and the lower panel shows the effects for targets in L1 (French).
3.4. Discussion
The results of Experiment 2 show a pattern that is qualitatively very similar to the one found in Experiment 1. Code-switching effects were once again primarily evident in the N250 component for L2 targets. However, the code-switching effects in later ERP components were more robust in Experiment 2, with clearer evidence for an influence of language switching in the N400 region for both L1 and L2 targets. Once again, the fact that switch effects were in the same direction for L1 and L2 targets (i.e., greater negativity following a language switch) argues against a prime word frequency account of these effects. It would appear that it is indeed the language of the prime that is the critical factor here. In addition, primes were clearly visible in Experiment 2 compared to the limited visibility conditions of Experiment 1. We would therefore expect to have seen a qualitative shift in priming effects across these two experiments if executive control mechanisms were playing a central role, so the fact that the pattern of effects was quite similar in Experiments 1 and 2 is further evidence against a task or decision-level source of code-switching costs.
4. General discussion
The results of the experiments reported in the present study provide considerable constraint for theoretical accounts of language-switching costs in language comprehension in bilinguals. The most important result is that a change in language across otherwise unrelated prime and target words was found to influence target processing in conditions in which primes were largely invisible and all visible words were from the same language (since target language was blocked). This influence was found primarily in the N250 ERP component, and mostly for L2 target words. The N250 ERP component was more negative going during the processing of English (L2) target words when these followed a French (L1) prime word compared to an English prime word. We interpret this result as reflecting a greater effort in integrating information across prelexical and lexical representations during the processing of L2 target words following a language switch. Prior monolingual research combining masked priming and ERPs has shown increased negativity in the region of the N250 when prime and target are different words compared with repetition primes (e.g., Holcomb & Grainger, 2006).
At a general level of theorizing, the language switching effect found in the N250 with very brief prime exposures in Experiment 1 is striking evidence for a remarkable reactivity of the bilingual brain to changes in language that occur largely unconsciously.1 Participants in this experiment were only aware of stimuli in one language, the target language, over a given block of trials, yet there was a significant influence of the language of prime stimuli. This therefore suggests that information concerning the language a word belongs to is automatically computed during word recognition in bilinguals, and constitutes a key element in the process of bilingual word comprehension. This is one central hypothesis of the BIA-model of bilingual word recognition (Grainger & Dijkstra, 1992; van Heuven et al., 1998).
In the present experiments language switching effects were also seen in later ERP components, affecting processing up to and including the N400 component. The effects in later components were generally weaker and tended to be stronger with French (L1) targets, and were also significant at a greater number of electrode sites with the longer prime duration in Experiment 22 The language switching effects seen in the N400 component are interpreted as reflecting greater effort in integrating information across lexical and semantic representations. The fact that these effects were overall weaker than the effects seen in the N250 component, particularly with the brief prime exposure of Experiment 1, is likely due to the fleeting nature of prime word activation in such extreme conditions; however, it should be noted that in Experiment 2, the trial type by electrode site interaction is significant in the N400 window, for English (L2) target trials at all sites and for French (L1) target trials at midline sites. The significant switching effects from L2 to L1 found in the present study are all the more remarkable given the relatively low L2 proficiency level of our participants. This suggests that language control mechanisms are rapidly set-up during the earliest phases of second language learning.
The present study was designed to test alternative accounts of language switching effects in bilinguals, and in particular whether they are due to extra-lexical task control processes, or to the automatic modulation of language-specific representations. According to Green’s (1998) IC model and to the BIA+ model (Dijkstra & Van Heuven, 2002), task schemas play a central role in generating language switching effects observed in laboratory tasks. Changing languages is akin to changing tasks, such that following a language change there is a cost in processing because the task control mechanism was set to operate for the incorrect language. It is not obvious how this mechanism could account for the fast-acting language switching effects found with brief prime exposures in the present study. Since masking minimizes extra-lexical processing of the masked stimulus, if these executive control procedures were the main source of language-switching costs, then we would not have expected to see language switching costs in Experiment 1. Furthermore, if extra-lexical executive control processes were playing any role in code-switching costs we expected to see a different pattern of effects with the supraliminal prime presentations of Experiment 2. However, although the effects were stronger with the longer prime duration, the overall pattern was very similar in the two experiments.
The results of the present study are clearly more in line with the predictions of the original BIA model (Grainger & Dijkstra, 1992; van Heuven et al., 1998) according to which language switching effects are the result of automatic topdown modulation of activation of lexical representations as a function of language. Prime words rapidly activate the corresponding language node, which then inhibits the representations of all words in the irrelevant language (van Heuven et al., 1998). When prime and target are from the same language there is no influence of prime language node activation on processing of the target word. However, when the prime is from a different language, then the target word representation receives inhibition from the language node activated by the prime word, hence influencing on-going target word processing. We would speculate that this type of automatic modulation of the activation levels of linguistic representations might be housed in the left caudate nucleus, as suggested by one recent fMRI study (Crinion et al., 2006).
The code-switching effects found in the present study varied as a function of the direction of the language switch. French primes preceding English targets (that is, L1-L2 switches) showed the greatest effect in the N250, while French primes preceding English targets (L2-L1 switches) showed their greatest effect in the N400 window. This pattern is likely to be the result of a combination of effects of the limited prime durations used in the present study, and the relative speed of processing L1 and L2 prime words in our participants. The faster processing of L1 words enables a faster acting influence of these primes on the processing of L2 targets. This is in line with the BIA model’s account of code-switching effects as being driven by the language-switch condition rather than the no-change condition (it is the inhibition in the switch condition that causes the effect rather than any facilitatory influence in the no-switch condition). However, this effect is fleeting since the prime word’s influence will be cancelled as soon as the target word appears and the language node corresponding to the target word is activated. Primes in L2 take longer to activate the L2 language node, hence the delayed influence of language switching in the L2-L1 direction.
The fact that switching effects tended to be stronger in the L2-L1 direction in the N400 ERP component is in line with the above account, but appears to contradict to the results of Alvarez et al. (2003). These authors found stronger code-switching effects in the N400 for L1-L2 switches compared with L2-L1 switches in a sequential word reading paradigm. Thus, it would appear that with suffcient time (as in the Alvarez et al. study), code-switching effects are also stronger from L1 to L2 in later ERP components. It should be noted that studies of language switching effects in language production typically show the opposite pattern (e.g., Costa & Santesteban, 2004; Meuter & Allport, 1999). This apparent inconsistency across the production and comprehension literature is consistent with the BIA model’s account of control of lexical activation. In comprehension, the language node mechanism operates passively as a function of stimulus input and contextual information. Words in L1 provide stronger activation input to language nodes, hence the stronger cost when switching from L1 to L2. In production, the control mechanism operates top-down, even if the decision is triggered by an external cue. This results in an active regulation of language node activation as a function of the required level of inhibition, which is greater when producing an L2 word, since the irrelevant L1 words are harder to suppress than irrelevant L2 words when producing an L1 word.
Therefore, according to this account of differences in language-switching effects in comprehension and production, it is what one has to do with the first stimulus that is critical. This account predicts that a primed language production task, with passive processing of the prime stimulus, should show stronger switching effects from L1 to L2. This prediction is currently the object of on-going experimentation. Finally, It should be noted that the vast majority of natural code-switching occurs in the auditory modality, and not the visual modality that was tested in the present study. One problem with auditory experiments, however, is that there are many more prelexical cues to language identity (language-specific phonemes) than in the visual modality (for languages that share the same alphabet), possibility resulting in smaller switch costs. Nevertheless, on-going research in our laboratory is investigating language-switching effects in the auditory modality.
5. Conclusions
The present study examined language switching effects in a masked priming paradigm with ERP recordings. Language switching effects were observed in an early ERP component when targets were in L2. Language switching effects with L1 targets were mostly visible in the N400 component, and were more robust at the longer prime duration of Experiment 2. The fact that a language switching effect was found in conditions without an overt change in language (target language was blocked, and primes were largely invisible in Experiment 1) is strong evidence that such effects are generated by highly automatized mechanisms that are part of the core processes of bilingual word comprehension.
Footnotes
This research was supported by HD043251.
It is likely that the P200 is an amalgamation of ERPs generated by all preceding stimuli—target, backward mask, and prime, in approximately 80 ms. Some of the activity in the N250 window in this case could therefore be due to the P200.
Due to the spatially underdetermined nature of ERPs, no strong distributional claims can be made; but as factors for laterality and anterior/posterior position were included in the analysis, some distributional descriptions may be noted as consistent with other research on the N250 and N400. The N250 typically shows a broad scalp distribution (Holcomb & Grainger, 2006); in this case, the N250 was indeed seen at a broad range of sites. Additionally, consistent with Johnson and Hamm (2000), the N400 effect was most prominent at frontocentral and centroparietal sites. Although no strong claims can be made from this, it does seem highly likely that the components we are calling the N250 and the N400 in this analysis are the same as those reported in other studies.
References
- Alvarez RP, Holcomb PJ, Grainger J. Accessing word meaning in two languages: An event-related brain potential study of beginning bilinguals. Brain and Language. 2003;87(2):290–304. doi: 10.1016/s0093-934x(03)00108-1. [DOI] [PubMed] [Google Scholar]
- Cohen JD, Dunbar K, McClelland JL. On the control of automatic processes: A parallel distributed processing account of the Stroop effect. Psychological Review. 1990;97(3):332–361. doi: 10.1037/0033-295x.97.3.332. [DOI] [PubMed] [Google Scholar]
- Costa A, Santesteban M. Lexical access in bilingual speech production: Evidence from language switching in highly proficient bilinguals and L2 learners. Journal of Memory and Language. 2004;50:491–511. [Google Scholar]
- Crinion J, Turner R, Grogan A, Hanakawa T, Noppeney U, Devlin JT, et al. Language control in the bilingual brain. Science. 2006;312:1537–1540. doi: 10.1126/science.1127761. [DOI] [PubMed] [Google Scholar]
- Dalrymple-Alford EC. Language switching during bilingual reading. British Journal of Psychology. 1985;76(1):111–122. [Google Scholar]
- Dijkstra T. From tag to task: Coming to grips with bilingual control issues. Bilingualism: Language and Cognition. 1998;1(2):88–89. [Google Scholar]
- Dijkstra T, Van Heuven WJB. The architecture of the bilingual word recognition system: From identification to decision. Bilingualism: Language and Cognition. 2002;5(3):175–197. [Google Scholar]
- Finkbeiner M, Gollan TH, Caramazza A. Lexical access in bilingual speakers: What’s the (hard) problem? Bilingualism: Language and Cognition. 2006;9(2):153–166. [Google Scholar]
- Grainger J, Beauvillain C. Language blocking and lexical access in bilinguals. Quarterly Journal of Experimental Psychology: Human Experimental Psychology. 1987;39A(2):295–319. [Google Scholar]
- Grainger J, Dijkstra T. On the representation and use of language information in bilinguals. In: Harris RJ, editor. Cognitive processing in bilinguals. North Holland; Amsterdam: 1992. [Google Scholar]
- Grainger J, Holcomb PJ. Neural constraints on a functional architecture for word recognition. In: Cornelissen P, Hansen P, Kringelbach M, Pugh K, editors. The neural basis of reading. Oxford University Press; Oxford: in press. [Google Scholar]
- Grainger J, Kiyonaga K, Holcomb PJ. The time-course of orthographic and phonological code activation. Psychological Science. 2006;17:1021–1026. doi: 10.1111/j.1467-9280.2006.01821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green DW. Mental control of the bilingual lexico-semantic system. Bilingualism: Language and Cognition. 1998;1(2):67–81. [Google Scholar]
- Hernandez AE, Martinez A, Kohnert K. In search of the language switch: An fMRI study of picture-naming in Spanish-English bilinguals. Brain and Language. 2000;73(3):421–431. doi: 10.1006/brln.1999.2278. [DOI] [PubMed] [Google Scholar]
- Hernandez A, Dapretto M, Mazziotta J, Bookheimer S. Language switching and language representation in Spanish-English bilinguals: an fMRI study. NeuroImage. 2001;14(2):510–520. doi: 10.1006/nimg.2001.0810. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. On the time course of visual word recognition: An event-related potential investigation using masked repetition priming. Journal of Cognitive Neuroscience. 2006;18(10):1631–1643. doi: 10.1162/jocn.2006.18.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson BW, Hamm JP. High-density mapping in an N400 paradigm: evidence for bilateral temporal lobe generators. Clinical Neurophysiology. 2000;111(3):532–545. doi: 10.1016/s1388-2457(99)00270-9. [DOI] [PubMed] [Google Scholar]
- Macnamara J, Kushnir SL. Linguistic independence of bilinguals: The input switch. Journal of Verbal Learning and Verbal Behavior. 1972;10(5):480–487. [Google Scholar]
- Meuter RFI, Allport A. Bilingual language switching in naming: Asymmetrical costs of language selection. Journal of Memory and Language. 1999;40(1):25–40. [Google Scholar]
- Orfanidou E, Sumner P. Language switching and the effects of orthographic specificity and response repetition. Memory & Cognition. 2005;33(2):355–369. doi: 10.3758/bf03195323. [DOI] [PubMed] [Google Scholar]
- Scotton CM, Ury W. International Journal of the Sociology of Language. Vol. 13. Mouton Publishers; 1977. Bilingual strategies: The social functions of code-switching; pp. 5–20. [Google Scholar]
- Soares C, Grosjean F. Bilinguals in a monolingual and a bilingual speech mode: The effect of lexical access. Memory & Cognition. 1984;12:380–386. doi: 10.3758/bf03198298. [DOI] [PubMed] [Google Scholar]
- Thomas MSC, Allport A. Language switching costs in bilingual visual word recognition. Journal of Memory and Language. 2000;43(1):44–66. [Google Scholar]
- van Heuven WJB, Dijkstra T, Grainger J. Orthographic neighborhood effects in bilingual word recognition. Journal of Memory and Language. 1998;39(3):458–483. [Google Scholar]
- von Studnitz RE, Green DW. Lexical decision and language switching. International Journal of Bilingualism. 1997;1:3–24. [Google Scholar]
- Wang Y, Xue G, Chen C, Dong Qi. Neural bases of asymmetric language switching in second-language learners: An ER-fMRI study. NeuroImage. 2006;35(2):862–870. doi: 10.1016/j.neuroimage.2006.09.054. [DOI] [PubMed] [Google Scholar]