

Abstract
Background
Human parainfluenza virus type 3 (hPIV-3) has been reported to cause nosocomial outbreaks of respiratory infection, in particular among hematopoietic stem cell transplantation recipients.
Design and Methods
From September 2007 through January 2008 several episodes of hPIV-3 infection were observed among young patients followed at the Oncohematology Unit (OHU) or other units of the Pediatrics Department. In 32 young patients (median age 3.5 years, range 21 days–27 years), hPIV-3 infection was diagnosed by direct fluorescent antibody staining of cells from respiratory secretions, and virus quantified by real-time RT-PCR in nasopharyngeal aspirates or bronchoalveolar lavage samples. In addition, the epidemiologic relatedness of hPIV-3 strains was investigated by sequencing two variable regions of the hemagglutinin-neuraminidase gene (nt 1–569 and nt 762–1239).
Results
Of the 32 hPIV-3-positive patients, 19 were hematopoietic stem cell transplantation recipients, 8 had hematologic malignancies, and 5 were immunocompetent children. Sixteen patients had upper, and 16 lower respiratory tract infection. All patients but one had high viral load in nasopharyngeal aspirates (>1.0×106 RNA copies/mL). One patient died from respiratory failure with a high viral load in bronchoalveolar lavage. Phylogenetic analysis showed that 16/32 strains were identical. Besides this major cluster, three other clusters were identified, each one defining a smaller outbreak.
Conclusions
Phylogenetic analysis allows identification of the role of a single or multiple hPIV-3 strains in the person-to-person transmission within an outbreak occurring in clinical units.
Keywords: nosocomial outbreak, human parainfluenza virus type 3, hematopoietic stem cell transplant recipients, phylogenetic analysis
Introduction
Human parainfluenza virus type 3 (hPIV-3) is, along with human respiratory syncytial virus, a major cause of lower respiratory tract infections (RTI) in infants and young children, as well as in immunocompromised patients, particularly hematopoietic stem cell transplantation (HSCT) recipients. In this patient population, a high incidence of hPIV-3 infection with prolonged virus shedding, and a high mortality rate have been reported.1–4
Although steroid treatment for graft-versus-host disease (GvHD) has been shown to be associated with progression to hPIV-3 pneumonia, the only identified risk factor for hPIV-3 infection after HSCT was the use of an unrelated volunteer as stem cell donor.4 In addition, mortality rates increased when hPIV-3 infection occurred within 100 days after HSCT.2,3
In recent years, several outbreaks of hPIV-3 infection among adult HSCT recipients have been described,5–8 with mortality rates reported to be either high8 or low.7 In these reports, the adoption of adequate and timely preventive isolation measures was often ineffective in controlling virus transmission, due to virus presence in asymptomatic staff members or external visitors, or even on room surfaces.6 Sequencing and phylogenetic analysis of hPIV-3 strains directly in clinical specimens allowed the verification of whether a single strain or multiple hPIV-3 strains were responsible for the current outbreak. While in some outbreaks a single strain was found to circulate,8 in other outbreaks different hPIV-3 strains were circulating, thus originating different clusters within the same outbreak.5,6
In the present report, we describe an hPIV-3 infection nosocomial outbreak occurring in both the outpatient and inpatient Oncohematology sections of the Department of Pediatrics, showing that through several weeks an apparently single episode was actually supported by different hPIV-3 strains originating four clusters of infection of different magnitude, whereas strains detected post-outbreak gave origin to a totally independent cluster.
Design and Methods
Patient population
The Department of Pediatrics consists of different sections including, among others, the Oncohematology Unit (OHU). The OHU follows young patients with hematologic malignancies who either undergo HSCT or are treated with chemotherapy. Following discharge from the hospital, patients from the OHU are followed-up twice weekly for at least 100 days after HSCT and whenever clinically indicated. In addition, patients under chemotherapy have access to the hospital every time they must receive cytotoxic drugs or transfusions. The site of these periodical controls is the Outpatient Section (OPS) of the OHU. From the waiting room, patients are taken to separate visiting rooms, following potential air-borne contacts with parents of the other patients in the waiting room. It also has to be emphasized that protective measures such as hand washing, masks, and gloves may be not sufficient to prevent virus transmission. Since most patients are infants and young children, contacts with parents and accompanying persons from the community were frequent.
The study was approved by the Institutional Review Board and the Ethics Committee of our hospital. Informed consent from single patients or their parents was obtained upon first viral examination.
Viral diagnosis
On the whole, 116 nasopharyngeal aspirates (NPA), 2 tracheal aspirates and 2 bronchoalveolar lavage (BAL) samples were taken from all patients with respiratory tract symptoms upon admission and discharge from the OHU or other Units of the Department of Pediatrics, as well as, whenever possible, at periodical ambulatory visits to monitor virus persistence.
All respiratory samples collected were tested for all known respiratory viruses (see below) by both: (i) monoclonal antibodies (MAbs) and direct or indirect fluorescent antibody (DFA or IFA) staining of respiratory cells from NPA or BAL, and from 48 h cultured cells from inoculated shell-vials of mixed mink lung (Mv1Lu) and A549 epithelial cells;9 and (ii) molecular assays, i.e. real-time reverse transcriptase (RT)-PCR or RT-PCR, as previously reported.10,11 MAbs used for routine detection of influenza viruses A and B, parainfluenza viruses 1–4, human respiratory syncytial viruses A and B, human adenoviruses were purchased from Chemicon (Millipore, Temecula, CA, USA), whereas MAbs for identification of human metapneumoviruses A and B, human coronaviruses 229E and HKU-1 were developed in the laboratory. In addition, all four human coronaviruses (229E, NL63, OC43 and HKU-1) were detected by RT-PCR,11,12 while influenza virus A, as well as human respiratory syncytial viruses A and B, human metapneumoviruses A and B, human rhinoviruses, human enteroviruses were quantified by real-time RT-PCR, and human bocavirus by PCR.11 In particular, hPIV-3 was first detected in NPA and/or shell-vial cultures by MAbs, and then quantified by real-time RT-PCR and phylogenetically analyzed by sequencing two fragments of the hemagglutinin-neuraminidase (HN) gene (see below).
Real-time RT-PCR and hPIV-3 strain sequencing
Following nucleic acid extraction from 200 μL of NPA or BAL using the automatic extractor Nuclisens® easy MAGTM (BioMérieux, Lyon, France) and subsequent elution in 50 μL, the real-time RT-PCR for hPIV-3 quantification was performed by an RT reaction (final volume 15μL from 5 μL extract) using the forward primer PIV3-5613 and the SuperScriptTM II RNase H-Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). Thermal profile was 30 min at 42°C and 15 min at 70°C. The RT product was then amplified, according to a previously published protocol,13 in a final volume of 50 μL of reaction mixture using primers PIV3-56 and PIV3-129, and probe PIV3-86 and the following thermal profile: 2 min at 50°C, 10 min at 95°C, followed by 40 cycles of 15 sec at 95°C and 60 sec at 60°C using the ABI PRISM® 7000 Sequence Detection System (Applied BioSystems, Foster City, CA, USA). A standard DNA curve was constructed by testing serial tenfold dilutions (106 through 101) of a linearized plasmid obtained by cloning a reference hPIV-3 strain with the TA-Cloning kit (Invitrogen). From this curve, the amount of virus present in clinical samples was extrapolated.
Sequencing of the HN gene of hPIV-3 strains was done by single-step RT-PCR using the enzyme SuperScriptTM One-Step RT-PCR and Platinum Taq (Invitrogen). Two distinct fragments of the HN gene were amplified. The first was the 5’ portion of the gene (nt 1–569) which was amplified using the following in-house designed primers: hPIV-3-APfor1, 5’-ATGGAATACTGGAAGCAYACC-3’; and hPIV-3-APrev1, 5’-AGCCATCAWCAGTYGTTGGCA-3’. Thermal profile was: 30 min at 50°C, 10 min at 95°C followed by 45 cycles of 45 sec at 95°C, 45 sec at 58°C, and 60 sec at 72°C. The second fragment was in the 3’ region of the gene (nt 762–1239), and primers and thermal profile were as already reported.14 Following purification of the two PCR amplification products, the sequence reaction was performed by using the ABI PRISMTM Big Dye Terminator kit (Applied Biosystems). Sequences were then analyzed using the Genetic Analyzer 3100 System (Applied Biosystems). In addition, sequences were cut and checked by using the 4.7 version of the Sequencher Software (gene Codes Corp., Ann Arbor, MI, USA), and then aligned with the MEGA 4.0 software using the Clustal W method. Phylogenetic analysis was carried out using the MEGA 4.0 software (genetic distance method), while the kimura 2-parameter was selected as an evolution model, and the bootstrap value was 1,000 repetitions.
Statistical analysis
The curves of the percentages of patients shedding hPIV-3 in the weeks following onset of infection in the two groups of immunocompromised and immunocompetent patients were calculated and expressed as cumulative incidences, and compared by the log-rank test; p values lower than 0.05 were considered to be statistically significant.
Results
hPIV-3 outbreak
From the end of September 2007 through the beginning of January 2008, 32 patients (median age 3.5 years, range 21 days–27 years) admitted to the Pediatrics Department (either OHU or other Units) of the Fondazione Istituto di Ricovero e Cura a Carattere Scientifico Policlinico San Matteo, Pavia, Italy, were found to be positive for hPIV-3 in cells from respiratory secretions by DFA. Viral load in the respiratory tract was then quantified by real-time RT-PCR.
In addition, for comparison, 8 hPIV-3 strains were recovered from sporadic cases of respiratory infection observed in different wards of the hospital (4 in the Pediatrics Department, 3 in the Hematology Unit and one in the Transplantation Center of the Respiratory Disease Unit) between February and July 2008. Thus, on the whole, 40 patients with hPIV-3 infection were examined: 32 during or around the outbreak, and 8 in the following six months.
Figure 1A shows the monthly distribution of the 32 sequential cases of hPIV-3 infection in the Pediatrics Department between the end of September 2007 and the beginning of January 2008. The peak number of cases of hPIV-3 infection was reached between October and November 2007, when 24 cases were observed.
Figure 1.
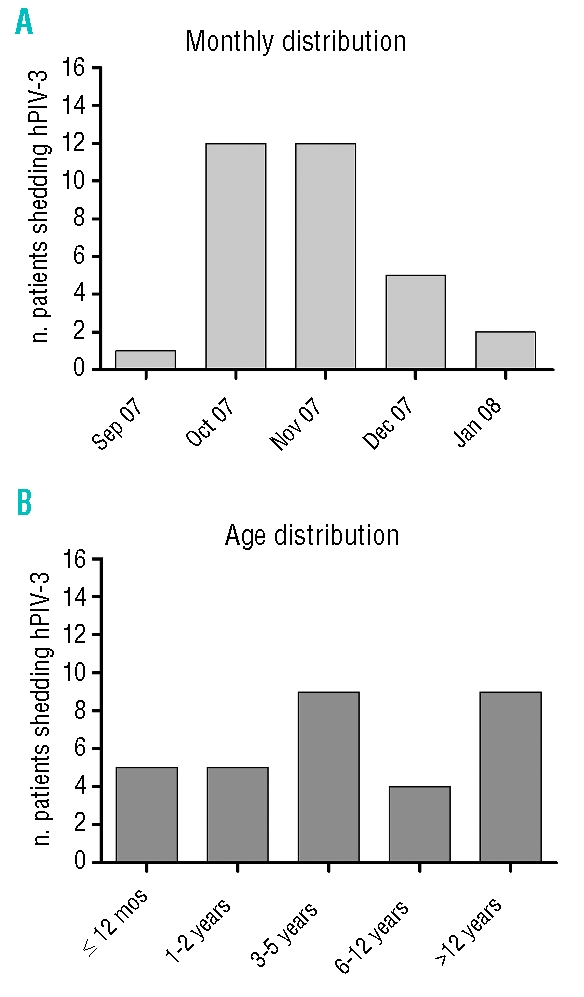
(A) Monthly and (B) age distribution of the 32 young patients involved in the outbreak of hPIV-3 infection.
The age distribution is reported in Figure 1B, where a comparable number of cases of hPIV-3 infection among different age groups was observed within the outbreak. Of the 32 patients involved in the outbreak, 19 underwent HSCT and suffered from hPIV-3 infection in the months preceding or following transplantation, while 8 patients were affected by hematologic malignancies and repeatedly attended the OHU, both in the ward and the OPS. Finally, 5 immunocompetent children admitted to other Units of the Pediatric Department were affected by hPIV-3 infection during the outbreak period.
In all cases but one, a high hPIV-3 load (>1.0×106 RNA copies/mL) was detected (median 4.9×107 RNA copies/mL; range 7.2×104–1.4×109) in respiratory secretions, suggesting that hPIV-3 was responsible for clinical symptoms. In 16/32 patients (50.0%), mild symptoms related to upper respiratory tract infection were observed (rhinitis, cough, and sore throat), whereas in 16 patients symptoms or syndromes related to lower respiratory tract infection were observed (bronchitis, bronchiolitis and pneumonia). The only patient with a hPIV-3 load <1.0×105 RNA copies/mL was an HSCT recipient (from a matched unrelated donor) with bilateral pneumonia and hPIV-3 respiratory infection ongoing from an undefined number of days (pt # 28; see below). Thus, a level of 1.0×105 RNA copies/mL NPA was selected as a cut-off for diagnosis of acute hPIV-3 infection in respiratory secretions.
The percentage of hPIV-3 shedding patients over time in the two groups of immunocompromised and immunocompetent pediatric patients is reported in Figure 2. The median duration of viral shedding was 72 days for immunocompromised patients, and 18 days for immunocompetent patients, with a highly significant difference (log-rank test, p<0.0001). The only patient with hPIV-3 infection within the outbreak group who died was a 4-month-old infant affected by severe combined congenital immunodeficiency (Omenn syndrome) with a very high hPIV-3 load (108–109 hPIV-3 RNA copies/mL) in both NPA and tracheal aspirate, in association with signs and symptoms suggestive of bronchiolitis and pneumonia. It must be noted that this child also had a disseminated infection from BCG vaccination. Notwithstanding the persisting (after 30 days) hPIV-3 infection with high viral load (in association with a low viral load human coronavirus NL63 infection) an unrelated cord blood transplantation was performed, due to severe progressively deteriorating clinical conditions of the patient, but after two weeks the patient deceased due to repeated episodes of pulmonary hemorrhage (Figure 3).
Figure 2.
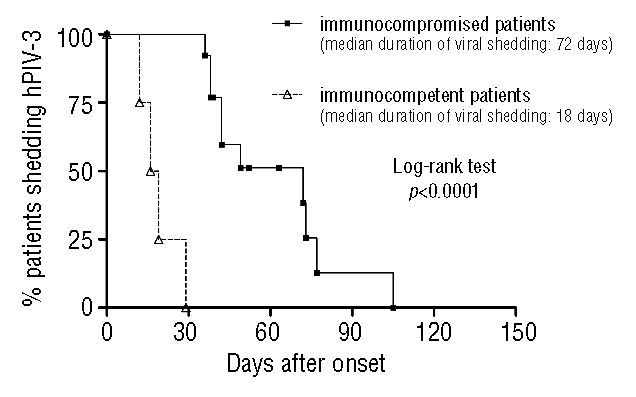
Percentage of immunocompromised and immunocompetent young patients shedding hPIV-3. The duration of viral shedding was significantly higher in immunocompromised patients.
Figure 3.
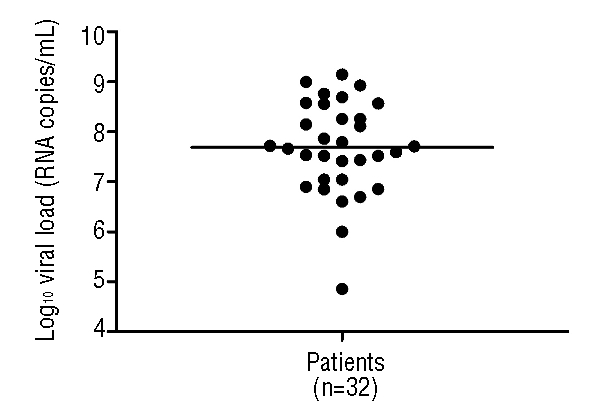
Peak levels of hPIV-3 RNA in respiratory secretions from the 32 young patients followed at the Oncohematology Unit or other units of the Pediatrics Department. The horizontal line indicates the median level of viral load in respiratory secretions.
In 5/32 patients involved in the outbreak, hPIV-3 infection was associated with a co-infection by another virus. In 3 cases, the co-infecting virus was a human rhinovirus, in one case the human coronavirus OC43, and in one case the human coronavirus NL63. In addition, in 1/8 patients who experienced hPIV-3 infection outside the outbreak, the viral infection was associated with a co-infection by two other viruses (human rhinovirus and human bocavirus).
Molecular analysis
Phylogenetic analysis of the HN gene of the 40 hPIV-3 strains relevant for the 40 patients examined (32 within the outbreak period, and 8 following the outbreak) allowed the construction of the phylogenetic tree shown in Figure 4. Two major clusters were identified: cluster #1, including 16 identical hPIV-3 strains (relevant to patients #9, 10, 13, 14, 15, 16, 17, 18, 19, 20, 23, 24, 25, 26, 28 and 32), and cluster #2, including six identical hPIV-3 strains (relevant to patients #1, 2, 4, 5, 6 and 11). In addition, besides single genetically distant strains, cluster #3 included three identical strains (patients #3, 7, and 8), and cluster #4 included two identical strains (patients #29 and 30) and one strain (patient #31) differing by a single nucleotide. Finally, 6/8 strains identified after the outbreak (relevant in patients #34, 35, 36, 37, 38, and 39), although clearly genetically distant from one another, showed a trend towards the formation of a cluster (cluster #5), in which the three closest strains (patients #37, 38, and 39) were detected in 3 adult patients followed at the Hematology Unit (adult patients) in the same period.
Figure 4.
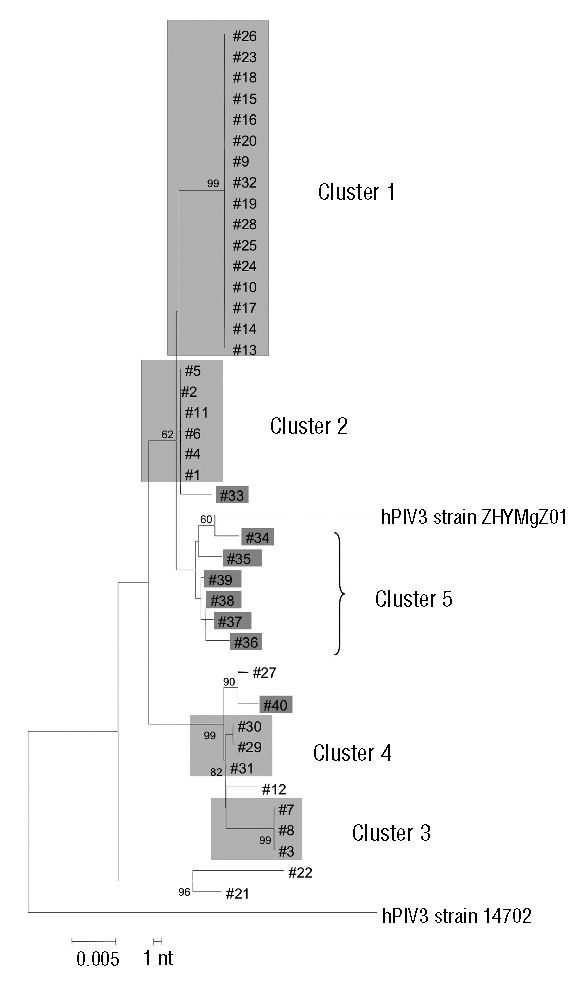
Phylogenetic analysis of the 32 hPIV-3 strains (each detected in a different patient) of the nosocomial outbreak (clusters #1, #2, #3, and #4), and the 8 post-outbreak strains (cluster #5). The strains belonging to the same cluster are grouped within the same light grey area, while the 8 strains post-outbreak are individually marked with a darker grey area. The 4 strains not highlighted are individual strains that appeared during the outbreak. GenBank accession numbers are EU424062 for hPIV3 strain 14702, and EU326526 for hPIV3 strain ZHYMgZ01.
Epidemiological analysis
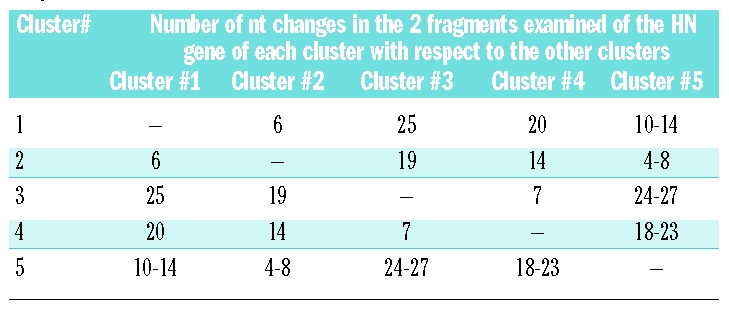
The chronological sequence for the detection of different hPIV-3 strains belonging to different clusters identified during the outbreak is reported in Figure 5, where patients infected by an identical hPIV-3 strain are reported within the same grey area, and genetically single strains are reported separately. Chronologically, the first cluster to emerge between the end of September and October 2007, was cluster #2, which also included an immunocompetent young child (#4). At the same time, cluster #3 was detected, while the major cluster #1 started appearing at the end of October 2007, lasting through the beginning of January 2008, and again included an immunocompetent child (#26). At the end of December 2007, a new small cluster #4 appeared within patients followed in OHU, the strains having a very close similarity to the strain detected in an immunocompetent child (patient #31). The number of nucleotide changes detected among the four hPIV-3 clusters identified during the outbreak and in comparison with cluster #5 is reported in Table 1.
Figure 5.
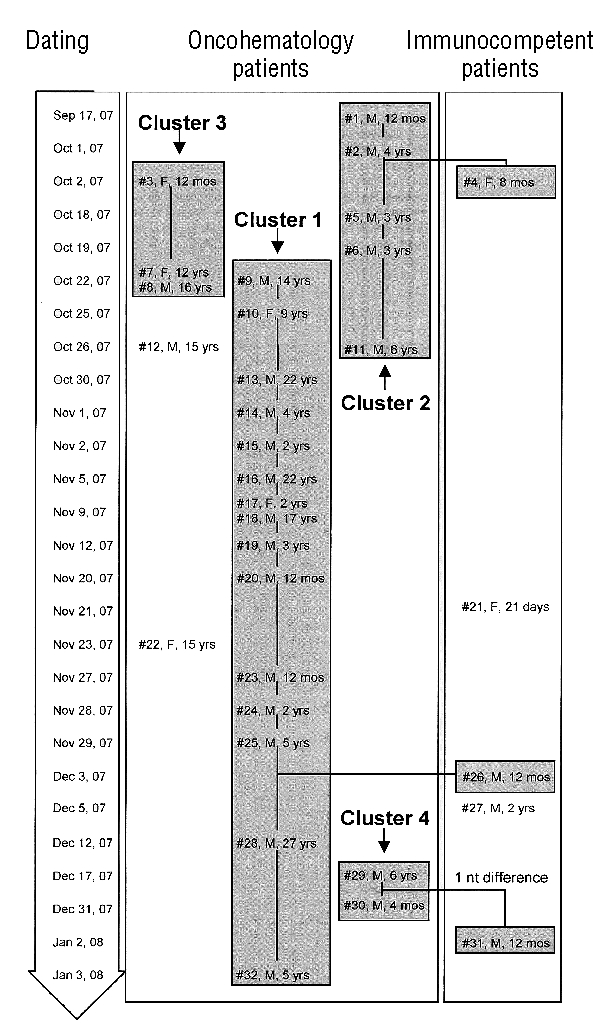
Chronological appearance of the 32 hPIV-3 strains of the nosocomial outbreak. Each strain is reported as patient #, sex and age. Patients are grouped within the single clusters (grey areas).
Table 1.
Number of nt changes observed among the 4 clusters of the hPIV-3 outbreak and in comparison with cluster #5 relevant to hPIV-3 strains detected post-outbreak.
Persistence over time of the same hPIV-3 strain in mmunocompromised patients
In a group of 6 patients (#5, #19, #21, #23, #25, #27), monitoring of hPIV-3 shedding was continued for a median time of 57 (range 22–72) days with a median number of 3.5 (range 2–6) viral tests/patient. The virus strain was consistently found to be identical for each patient throughout the entire virologic monitoring period.
Discussion
Results of the present study indicate that a nosocomial outbreak of hPIV-3 infection occurred among patients followed at the OHU of the Pediatrics Department of our University Hospital over a period of about 15 weeks during the fall season. The chronological analysis of the outbreak with respect to sporadic cases of hPIV-3 infection observed in the post-outbreak period suggested that a single strain could have been responsible for the outbreak. However, the phylogenetic analysis of the outbreak and post-outbreak strains revealed that multiple hPIV-3 strains were responsible for the outbreak. In detail, a single hPIV-3 strain contributed to cluster #1 including 16 patients (15 immunocompromised, and one immunocompetent patient), while another strain formed cluster #2 consisting of 6 patients (5 immunocompromised, and one immunocompetent), and two additional strains formed the two small clusters #3 and #4, including 3 patients each. One of the 3 patients of cluster #4 differed by a single nucleotide (patient #31) from the other 2 patients. Besides the four strains forming the four clusters, four single strains were found during the outbreak (patients #12, 21, 22, and 27), which presumably were introduced into the wards or the OPS from the community (2 HSCT recipients, and 2 immunocompetent patients) in the absence of nosocomial transmission. These findings allow the hypothesis that both routes of transmission (community-acquired and nosocomial) may have contributed to the outbreak.
In the past, several outbreaks of hPIV-3 infection among adult HSCT recipients have been reported, notwithstanding the preventive isolation measures taken to limit virus transmission within the interested units. Phylogenetic analysis has been of great utility in tracing virus transmission within clinical wards. The first report appeared in 1993 and showed (based on sequencing of a fragment of the F gene) that within a pediatric intermediate care unit 6 children were infected by genetically very similar hPIV-3 strains different from community isolates.15 In 1998, in an adult HSCT unit, it was shown (again by sequencing a fragment of the F gene) that multiple genetically distinct hPIV-3 strains were introduced from the community into the unit, with subsequent nosocomial transmission of a single strain.16 In addition, the same report emphasized the sustained shedding of the same hPIV-3 strain by transplanted patients for up to four months. These findings fit with the results of the present study dealing with pediatric HSCT recipients and oncohematologic patients and show that: (i) multiple genetically distinct hPIV-3 strains were introduced from the community into the unit during the outbreak; (ii) while a single strain caused the major outbreak lasting more than three months, three other distinct strains caused minor levels of transmission, and some additional strains detected during the outbreak did not give any sign of nosocomial transmission; (iii) shedding over time of hPIV-3 strains by young hematologic patients was significantly longer than that of immunocompetent children.
More recently, several additional outbreaks of hPIV-3 infection among adult HSCT recipients have been reported based on molecular analysis of the HN gene5,6 or the F gene8 or even in the absence of phylogenetic analysis.7 The epidemiologic, in the absence of the molecular, approach to the study of an hPIV-3 outbreak has been repeatedly shown to be unable to define the role of a single or multiple hPIV-3 strains7 in the person-to-person transmission within an outbreak. It must be noted that often during the molecular analysis of hPIV-3 outbreaks, differences in a small number of nt (1–3) within the sequence analyzed of a given gene, have been considered to represent a virus strain very similar to the major strain responsible for the outbreak, and, thus, have been grouped together with the major virus strain not displaying nt changes among several patients.5,6 However, as previously reported by other8 and confirmed in our study, hPIV-3 strains do not undergo any nt change even over an extended period of time (at least five months).
As for the mortality rate of HSCT recipients infected by hPIV-3 during an outbreak, disparate results have been reported. During some outbreaks a high mortality rate was reported, reaching levels as high as 47% (7/15 HSCT patients with lower RTI associated with hPIV-3 infection).8 In other reports, the mortality rate ranged from 25% (2/8 patients)17 or 26% (16/61 patients)2 to 8% (1/12 patients).5 Infection and mortality rates appeared to vary according to type of transplant, conditioning and immunosuppression regimens, and post-transplantation complications. However, in most cases, it appears difficult to attribute the cause of death to hPIV infection, since virus in lungs or BAL secretions is often not detected, and, thus, only an association of hPIV-3 infection and death can be documented. In our study, only one out of the 32 patients (3.1%) with hPIV-3 infection died. We cannot exclude that the original disease (Omenn syndrome) causing profound defects in both arms (cellular and humoral) of the immune response, as well as the disseminated BCG infection, contributed to the death of this patient, although the lack of necroscopy findings did not permit more information to be gained about the role of hPIV-3 infection in the final outcome of this patient.
The selection of a hPIV-3 load cut-off of 1.0×106 RNA copies/mL respiratory secretions was determined by the detection of equal or greater amounts of virus in all patients with an acute hPIV-3 infection, with the exception of a single patient in whom the virus was likely to have infected the patient since an undefined number of days. On this basis, it was possible, in case of double infection, to identify the virus responsible for the current episode of acute respiratory tract infection.
In our study, the waiting room could be identified as the major site of virus transmission between the community and hospital patients, as already reported in a previous study.6 Virus strains could have been introduced into the waiting room by either symptomatic or asymptomatic children, accompanying family members or health-care workers, and then transmitted either directly through personal contact or indirectly through contact with contaminated surfaces. It is known that hPIV may be shed with respiratory secretions by healthy asymptomatic children and adults18,19 and may persist on surfaces for weeks.6 Once introduced, viruses could spread from patient to patient, within the ward and the OPS of the OHU, despite the use of conventional protection devices, such as masks, disposable gowns, gloves, as well as hand disinfection. Findings of the present study indicate and confirm that preventive measures should be applied more intensively also at the level of the OPS of OHU. They should consist in the isolation of infected patients and routine hand cleaning with ethanolic disinfectants by staff members, patients and parents/visitors. As performed in this study, results of viral diagnosis should be given to clinicians the same day as NPA collection, in order to guide preventive strategies during the outbreak.
Acknowledgments
we thank the entire technical staff of the Virology Service for performing the assays, the nurses of the Pediatrics Department and other clinical wards participating in the study for taking the samples from the patients, and Daniela Sartori for preparing the manuscript. We also thank Laurene Kelly for revising the English of this manuscript.
Footnotes
Authorship and Disclosures
AP: analysis and interpretation of data; EP: performance and interpretation of immunofluorescence assays; ADC: patient follow-up; FL: critical revision of the manuscript; GG: drafting the manuscript.
The authors reported no potential conflicts of interest.
Funding: this work was partially supported by grants from the Ministero della Salute, Fondazione IRCCS Policlinico San Matteo Ricerca corrente (grants 80541 e 80622) and Ricerca Finalizzata (grant 89302).
References
- 1.Wendt CH, Weisdorf DJ, Jordan MC, Balfour HH, jr, Hertz MI. Parainfluenza virus respiratory infection after bone marrow transplantation. N Engl J Med. 1992;326:921–6. doi: 10.1056/NEJM199204023261404. [DOI] [PubMed] [Google Scholar]
- 2.Lewis VA, Champlin R, Englund J, Couch R, Goodrich JM, Rolston K, et al. Respiratory disease due to parainfluenza virus in adult bone marrow transplant recipients. Clin Infect Dis. 1996;23:1033–7. doi: 10.1093/clinids/23.5.1033. [DOI] [PubMed] [Google Scholar]
- 3.Elizaga J, Olavarria E, Apperley J, Goldman J, Ward K. Parainfluenza virus 3 infection after stem cell transplant: relevance to outcome of rapid diagnosis and ribavirin treatment. Clin Infect Dis. 2001;32:413–8. doi: 10.1086/318498. [DOI] [PubMed] [Google Scholar]
- 4.Nichols WG, Corey L, Gooley T, Davis C, Boeckh M. Parainfluenza virus infections after hematopoietic stem cell transplantation: risk factors, response to antiviral therapy, and effect on transplant outcome. Blood. 2001;98:573–8. doi: 10.1182/blood.v98.3.573. [DOI] [PubMed] [Google Scholar]
- 5.Cortez KJ, Erdman DD, Perez TCT, Gill VJ, Childs R, Barrett AJ, et al. Outbreak of human parainfluenza virus 3 infections in a hematopoietic stem cell transplant population. J Infect Dis. 2001;184:1093–7. doi: 10.1086/322041. [DOI] [PubMed] [Google Scholar]
- 6.Nichols WG, Erdman DD, Han A, Zukerman C, Corey L, Boeckh M. Prolonged outbreak of human parainfluenza virus 3 infection in a stem cell transplant outpatient department: insights from molecular epidemiologic analysis. Biol Blood Marrow Transplant. 2004;10:58–64. doi: 10.1016/j.bbmt.2003.09.010. [DOI] [PubMed] [Google Scholar]
- 7.Dignan F, Alvares C, Riley U, Ethell M, Cunningham D, Treleaven J, et al. Parainfluenza type 3 infection post stem cell transplant: high prevalence but low mortality. J Hosp Infect. 2006;63:452–8. doi: 10.1016/j.jhin.2006.03.010. [DOI] [PubMed] [Google Scholar]
- 8.Jalal H, Bibby DF, Bennett J, Sampson RE, Brink NS, MacKinnon S, et al. Molecular investigations of an outbreak of parainfluenza virus type 3 and respiratory syncytial virus infections in a hematology unit. J Clin Microbiol. 2007;45:1690–6. doi: 10.1128/JCM.01912-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Huang Y-T, Turchek BM. Mink lung cells and mixed mink lung and A549 cells for rapid detection of influenza virus and other respiratory viruses. J Clin Microbiol. 2000;38:422–3. doi: 10.1128/jcm.38.1.422-423.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sarasini A, Percivalle E, Rovida F, Campanini G, Genini E, Torsellini M, et al. Detection and pathogenicity of human metapneumovirus respiratory infection in pediatric Italian patients during a winter-spring season. J Clin Virol. 2006;35:59–68. doi: 10.1016/j.jcv.2005.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gerna G, Piralla A, Campanini G, Marchi A, Stronati M, Rovida F. The human bocavirus role in acute respiratory tract infections of pediatric patients as defined by viral load quantification. New Microbiol. 2007;30:383–92. [PubMed] [Google Scholar]
- 12.Gerna G, Campanini G, Rovida F, Percivalle E, Sarasini A, Marchi A, et al. Genetic variability of human coronavirus OC43-, 229E-, and NL63-like strains and their association with lower respiratory tract infections of hospitalized infants and immunocompromised patients. J Clin Virol. 2006;78:938–49. doi: 10.1002/jmv.20645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hu A, Colella M, Zhao P, Li F, Tam JS, Rappaport R, et al. Development of a real-time RT-PCR assay for detection and quantitation of parainfluenza virus 3. J Virol Methods. 2005;130:145–8. doi: 10.1016/j.jviromet.2005.06.014. [DOI] [PubMed] [Google Scholar]
- 14.Echevarrìa JE, Erdman DD, Swicrkosz EM, Holloway BP, Anderson LJ. Simultaneous detection and identification of parainfluenza viruses 1, 2, and 3 from clinical samples by multiplex PCR. J Clin Microbiol. 1998;36:1388–91. doi: 10.1128/jcm.36.5.1388-1391.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Karron RA, O’Brien KL, Froehlich JL, Brown VA. Molecular epidemiology of a parainfluenza type 3 virus outbreak on a pediatric ward. J Infect Dis. 1993;167:1441–5. doi: 10.1093/infdis/167.6.1441. [DOI] [PubMed] [Google Scholar]
- 16.Zambon M, Bull T, Sadler CJ, Goldman JM, Ward KN. Molecular epidemiology of two consecutive outbreaks of parainfluenza 3 in a bone marrow transplant unit. J Clin Microbiol. 1998;36:2289–93. doi: 10.1128/jcm.36.8.2289-2293.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Whimbey E, Vartivarian SE, Champlin RE, Elting LS, Luna M, Bodey GP. Parainfluenza virus infection in adult bone marrow transplant recipients. Eur J Clin Microbiol Infect Dis. 1993;12:699–701. doi: 10.1007/BF02009383. [DOI] [PubMed] [Google Scholar]
- 18.Muchmore HG, Parkinson AJ, Humphries JE, Scott EN, McIntosh DA, Scott LV, et al. Persistent parainfluenza virus shedding during isolation at the South Pole. Nature. 1981;289:187–9. doi: 10.1038/289187a0. [DOI] [PubMed] [Google Scholar]
- 19.Frank AL, Taber LH, Wells CR, Wells JM, Glezen WP, Paredes A. Patterns of shedding of myxoviruses and paramyxoviruses in children. J Infect Dis. 1981;144:433–41. doi: 10.1093/infdis/144.5.433. [DOI] [PubMed] [Google Scholar]