

Abstract
Brucella ceti causes disease in Odontoceti. The absence of control serum collections and the diversity of cetaceans have hampered the standardization of serological tests for the diagnosis of cetacean brucellosis. Without a “gold” standard for sensitivity and specificity determination, an alternative approach was followed. We designed an indirect enzyme-linked immunosorbent assay (iELISA) that recognizes immunoglobulins G (IgGs) from 17 odontocete species as a single group. For the standardization, we used Brucella melitensis and Brucella abortus lipopolysaccharides, serum samples from seven resident odontocetes with no history of infectious disease displaying negative rose bengal test (RBT) reactions, and serum samples from seven dolphins infected with B. ceti. We compared the performance of the iELISA with those of the protein G ELISA (gELISA), the competitive ELISA (cELISA), and the immunofluorescence (IF) and dot blot (DB) tests, using 179 odontocete serum samples and RBT as the reference. The diagnostic potential based on sensitivity and specificity of the iELISA was superior to that of gELISA and cELISA. The correlation and agreement between the iELISA and the gELISA were relatively good (Ri/g2 = 0.65 and κi/g = 0.66, respectively), while the correlation and agreement of these two ELISAs with cELISA were low (Ri/c2 = 0.46, Rg/c2 = 0.37 and κi/c = 0.62, κg/c = 0.42). In spite of using the same anti-odontocete IgG antibody, the iELISA was more specific than were the IF and DB tests. An association between high antibody titers and the presence of neurological symptoms in dolphins was observed. The prediction is that iELISA based on broadly cross-reacting anti-dolphin IgG antibody would be a reliable test for the diagnosis of brucellosis in odontocetes, including families not covered in this study.
Brucellosis is a disease of terrestrial and marine mammals and a relevant zoonosis caused by intracellular bacteria of the genus Brucella (34). In the last 2 decades, several strains of Brucella have been isolated from marine mammals, primarily from dolphins, porpoises, and seals (6, 11, 13-16, 18, 28, 42, 43, 49). For these marine strains, species names were proposed, first as a single cluster called “Brucella maris” (27) and, thereafter, divided into “Brucella cetaceae” and “Brucella pinnipediae” (7). Recently, the proposal of incorporating these marine strains as two new species was accepted; however, their names were corrected to Brucella ceti and Brucella pinnipedialis (15). The former Brucella species is predominant in dolphins and whales, while the latter is mainly isolated from seals and sea lions. These two new marine Brucella species can be further divided into several strains according to molecular biotyping (20), indicating that the species group is heterogeneous. The marine species display phenotypic resemblance with smooth Brucella abortus and Brucella melitensis, possessing the same relevant surface antigens that are commonly used for the serological diagnosis of brucellosis in bovines, caprines, and ovines (4).
Most of the confirmed cases of brucellosis in cetaceans have been in odontocetes stranded on the shoreline, mainly animals displaying swimming and orientation problems (25). Commonly, the arrival of dolphins and whales on the shore attracts attention and brings people into close contact with these animals, potentially serving as a source of Brucella infections to humans (5, 32, 45). Moreover, some of these stranded marine mammals, after treatment and recovering from various illnesses, are maintained in aquariums for exhibition, research purposes, or facilitated therapies (2). The status of Brucella infection and shedding of the bacteria in these retained animals are not regularly monitored, either because the disease is ignored as a cause of stranding or, more likely, due to the absence of screening tests validated for odontocetes.
The difficulty in validating serological tests for cetacean brucellosis lies in the fact that a significant number of adequate serum samples from animals with proven infections are required, as well as negative controls from Brucella-free animals. As expected, this is very difficult to accomplish with wildlife species, especially those living in open populations, such as dolphins and whales. In addition, there are two different cetacean orders with many families and their corresponding species. For instance, the order Odontoceti, which includes dolphins, toothed whales, and porpoises, contains nine families, not all of them monophyletic, and approximately 73 different species (31). In spite of these difficulties, significant efforts to test cetaceans for brucellosis have been carried out using competitive enzyme-linked immunosorbent assay (cELISA), indirect protein G ELISA (gELISA), and other serological tests originally developed for bovine or caprine brucellosis (28, 36, 46, 48, 49). However, none of the serological tests assayed with cetaceans have been validated. In this study, the performances of six serological tests have been standardized and compared for the presumptive diagnosis of brucellosis in 179 serum samples from 17 different species of odontocetes, mainly from the Atlantic coast of the United States and the Pacific coast of Costa Rica.
MATERIALS AND METHODS
Odontocete serum collection.
From a total of 179 odontocete serum samples corresponding to 17 species (Fig. 1 and Table 1), 145 were from stranded animals attended at the Dolphin and Whale Hospital, Mote Marine Laboratory, Sarasota, FL; 14 belonged to resident animals from Six Flags Marine World, Vallejo, CA; seven were collected from resident animals at the Gulf World, Panama City Beach, FL; 12 were obtained from stranded dolphins on the Pacific coast of Costa Rica; and one was obtained from a dolphin stranded on the Atlantic coast of Spain. The negative control serum samples were chosen from seven healthy resident odontocetes (one member of Kogia breviceps, two of Steno bredanensis, three of Tursiops truncatus, and one of Stenella coeruleoalba) with no clinical symptoms of infectious diseases, all displaying negative reactions in rose bengal agglutination tests (RBT) (1). Positive samples were obtained from seven dolphins (six members of S. coeruleoalba and one of Tursiops truncatus), from which B. ceti was isolated (25). Serum samples were maintained frozen at −80°C or in 50% glycerol at −20°C. From these, one serum sample from S. coeruleoalba with an RBT agglutination titer of 1/16 and high readings (optical density at 460 nm of >1.6) in ELISA was used as a positive serum control in the assays. Serum samples from healthy and B. abortus-infected bovines were from Costa Rica (17).
FIG. 1.

Determination of antibody and conjugate reactivity against sera from different odontocete species. (A) Rabbit antiserum anti-IgG from S. bredanensis (S) was tested by immunodiffusion against sera from Delphinus delphis (Dd), Feresa attenuata (Fa), Grampus griseus (Gg), Kogia breviceps (Kb), Kogia sima (Ks), Lagenodelphis hosei (Lh), Mesoplodon europaeus (Me), Orcinus orca (Oo), Peponocephala electra (Pe), Physeter macrocephalus (Pm), Stenella attenuata (Sa), Stenella clymene (Scl), Stenella coeruleoalba (Sc), Stenella frontalis (Sf), Stenella longirostris (Sl), Steno bredanensis (Stb), and Tursiops truncatus (Tt). Serum from Bos taurus (Bt) was used as an out-group control. (B) An anti-S. bredanensis IgG-HRP conjugate was tested by WB against sera from 17 different odontocete species and a cow, and the reactivity against the heavy-chain IgG was measured by pixel densitometry. The WB experiments were performed at least four times, and the density of the IgG heavy-chain band of each serum species normalized using the IgG-heavy chain of S. bredanensis. Notice the weaker and partial reaction of Kogia species in immunodiffusion and the significantly weaker reaction of these species in WBs. *, P < 0.05; **, P < 0.005.
TABLE 1.
Distribution of positive serum samples from Odontoceti species in different diagnostic assays
| Species | Total no. of samples | No. of positive reactions ina:
|
|||||
|---|---|---|---|---|---|---|---|
| iELISA | cELISA | gELISA | RBT | IF | DB | ||
| T. truncatus | 55 | 13 | 15 | 7 | 14 | 20 | 31 |
| L. hosei | 24 | 5 | 5 | 2 | 5 | 7 | 13 |
| K. breviceps | 22 | 2 | 2 | 6 | 6 | 4 | 4 |
| S. bredanensis | 23 | 11 | 12 | 5 | 12 | 13 | 14 |
| S. coeruleoalba | 12 | 11 | 9 | 9 | 10 | 11 | 11 |
| G. griseus | 11 | 8 | 8 | 6 | 8 | 8 | 7 |
| S. longirostris | 7 | 6 | 3 | 7 | 7 | 6 | 7 |
| K. sima | 6 | 0 | 0 | 1 | 0 | 0 | 0 |
| S. clymene | 6 | 0 | 1 | 0 | 0 | 0 | 0 |
| S. attenuata | 4 | 0 | 0 | 0 | 0 | 0 | 0 |
| F. attenuata | 3 | 1 | 1 | 1 | 1 | 1 | 1 |
| S. frontalis | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| P. electra | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| D. delphis | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| M. europaeus | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| O. orca | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| P. macrocephalus | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 179 | 58 (32.4) | 58 (32.4) | 45 (25.1) | 63 (35.2) | 71 (39.7) | 89 (49.7) |
Numbers within parentheses are the percentage of positive serum samples per assay.
Brucella antigens and generation of antibodies and conjugates.
Lipopolysaccharides (LPS) from B. abortus 2308 (AC epitope type) and B. melitensis 16 M (MC epitope type) were extracted and purified as previously described (3, 35) and stored at −20°C in aliquots of 1 mg/ml. Before use, LPS solutions were sonicated and diluted accordingly. The isolation of immunoglobulin G (IgG) from S. bredanensis serum was performed by standard procedures (33, 39, 40), and the purity of the fraction was evaluated in silver-stained gels (22). The purified dolphin immunoglobulin preparation demonstrated only two bands corresponding to the heavy and light chains of IgG. Antibodies against S. bredanensis IgG were raised in rabbits as described elsewhere (22). Rabbit IgG was purified and conjugated with fluorescein isothiocyanate (FITC; Sigma-Aldrich) or horseradish peroxidase (HRP; Sigma-Aldrich) following previously described protocols (26, 39). Recombinant protein G-HRP was purchased from Pierce Biotechnology, Inc. Conjugates were maintained in the dark and stored in aliquots at −20°C in the presence of 1% lipid-free bovine serum albumin (BSA) and 50% glycerol.
Immunochemical and serological assays.
RBT was performed and evaluated as described elsewhere (1). The cELISA for the diagnosis of brucellosis (Svanova Biotech, Uppsala, Sweden) was performed according to the instruction manual, using the same conditions established for testing bovine sera. In all assays, positive and negative dolphin serum samples were included in the analysis in parallel with the commercial controls included in the kit. The gELISA was performed essentially as reported before for bovine sera (44), with the following modifications. One hundred microliters of control and serum samples diluted 1/250 in phosphate-buffered saline (PBS)-Tween-BSA was added to the wells of polystyrene Immunolon II 96-well microplates (Nalge Nunc International), previously coated with Brucella LPS. After incubation, plates were washed, and a 100-μl volume of a solution of 250 ng/ml protein G-HRP diluted either in PBS-Tween containing 0.1% BSA or in 0.1 M acetate buffer (pH 5), was added to each well; then, the plates were incubated for 1 hour at 37°C in an orbital shaker. The plates were washed with PBS-Tween containing 0.1% BSA; then, 100 μl of the ABTS [2,2′-azinobis(3-ethylbenzthiazolinesulfonic acid)] substrate chromogen (Sigma-Aldrich) was added per well. After incubation at 37°C, plates were read at 405 nm with an ELISA reader.
Indirect ELISA (iELISA) was performed as described previously (30), with modifications. Briefly, polystyrene Immunolon II 96-well microplates were coated with 100 μl/well of LPS solution (25 to 200 μg/ml) for 18 h at room temperature, sealed with adherent plastic covers, and kept at −20°C until used. After five washes with 200 μl of PBS-Tween 20 per well, controls and samples were diluted in PBS-Tween containing 1% BSA, and 100 μl of the solution was added to each well. Microplates were incubated for 1 hour at 37°C in an orbital shaker and then washed again, and 100 μl of the anti-dolphin IgG-HRP diluted in PBS-Tween containing 1% BSA was added per well. The plates were incubated again for 1 hour at 37°C in an orbital shaker. After incubation, the plates were washed in either PBS as described above, and 100 μl of the ABTS substrate chromogen (Sigma-Aldrich) was added per well. After 1 hour incubation at 37°C, plates were read at 405 nm with an ELISA reader.
An immunoenzymatic dot blot (DB) assay was performed in a Bio-Dot microfiltration system (Bio-Rad Laboratories, Inc.) following the manufacturer's instructions. Briefly, 100 μl solution of B. melitensis LPS diluted in PBS was dispensed onto the nitrocellulose membrane (Bio-Rad) in each well and filtered with the aid of a vacuum. After filtration, the membrane was blocked with a solution of PBS containing 1% skim milk (Sigma-Aldrich), and the membrane was washed three times (10 min each) with PBS-Tween 20. Then, 50 μl of the serum samples and controls diluted 1/5,000 in PBS-Tween containing 0.1% BSA was added to each well, and the plates were incubated for 1 hour at room temperature in an orbital shaker. The membrane was then washed as described above, and 50 μl anti-dolphin IgG-HRP diluted in PBS-Tween containing 1% BSA (10 ng/ml) was added to each well. The plates were then incubated for 2 hours at room temperature and washed as described above, and the dots were revealed on an autoradiography film (Eastman Kodak Co.) with SuperSignal chemiluminescent substrate (Thermo Fisher Scientific, Inc.), following manual instructions.
For indirect immunofluorescence (IF), we initially tested B. ceti whole cells as an antigen; however, we did not observe any difference in IF sensitivity or specificity compared to those of B. abortus S19 whole cells. For safety reasons, B. abortus S19, which is attenuated in cattle and lowered risk for brucellosis in humans, was used as the antigen throughout all IF experiments. Live bacteria were dried at 37°C on a glass slide coverslip and fixed with 3.5% paraformaldehyde at room temperature for 15 min. Then, 50 mM NH4Cl was added, and the coverslips were incubated for 10 min at room temperature. Coverslips were rapidly rinsed in PBS, and then, 50 μl of the odontocete serum diluted 1/50 in PBS containing 10% horse serum (HS) was added. After incubation for 30 min at room temperature, coverslips were rinsed three times with PBS and once with PBS containing 0.1% Triton X-100. Then, 50 μl of the FITC-IgG diluted 1/50 in PBS-HS was added to each coverslip and incubated for 30 min at room temperature. Finally, the coverslips were washed as described before, and 10 μl of Mowiol (Merck) was added to the top of each coverslip and turned onto a glass slide. The slides were left overnight at room temperature for polymerization, and samples were evaluated the next day by fluorescence microscopy (Olympus BH-2).
An immunodiffusion assay for the estimation of immunoglobulin cross-reactivity was carried out in 1% agarose gels as described elsewhere (26). Alternatively, Western blotting (WB) of purified IgG and sera was performed on polyvinylidene difluoride membranes (Pall Biosciences, Dreieich, Germany) as described elsewhere (22) and results revealed with anti-dolphin IgG-HRP followed by the SuperSignal chemiluminescent substrate on an autoradiography film. The level of reaction of anti-dolphin IgG-HRP against the IgG of the 17 different species of odontocetes was estimated on photographs by pixel densitometry of the IgG heavy chain, using the public domain ImageJ program (http://rsb.info.nih.gov/ij/). The IgG of the homologous species S. bredanensis was used as an internal standard (100% value), and each experimental value was expressed as the percentage of the standard. The WB experiment was performed at least four times, the relative values for each IgG heavy-chain species were averaged, and the standard error was calculated.
Statistical analysis.
Comparative statistical analysis was performed using public access package Win Episcope 2.0 software (47). The interpretation of the area under the ROC (receiver operating characteristics) curves was made as described by Hanley and McNeil (21). The parameters used to establish the cutoff values and the interpretation of the diagnostic results of brucellosis were based on previous works (30). Briefly, the cutoff values of cELISA, gELISA, and iELISA were adjusted within the limits of the lower percentage of positivity obtained by a positive serum and the higher percentage of positivity obtained by a negative serum. Within these limits, the cutoff value for each ELISA was empirically settled at intersecting values which excluded the minimal number of false positives and false negatives, taking RBT as the reference. This agglutination test was used as the reference because its performance is independent of secondary antibodies and because it is competent in discriminating antibodies in sera from Brucella-infected and noninfected animals (1). Under these conditions, the cutoff values for the cELISA, gELISA, and iELISA were empirically settled at 30.6%, 6.0%, and 7.5% positivity, respectively.
RESULTS
A potential drawback for the use of immunoassays that depend upon anti-dolphin antibody conjugates for the diagnosis of marine brucellosis is that pathogenic Brucella infects different species of odontocetes, hypothetically restricting the use of the anti-S. bredanensis IgG-HRP and IgG-FITC conjugates. Therefore, we first tested the reactivity of the antibody and the respective HRP conjugate against sera from 17 species of odontocetes. As shown in Fig. 1A, a strong precipitation line was evident in the immunodiffusion assay against antibodies of all 17 species. A partial identity and somewhat weaker precipitation lines were observed in assays against Kogia sima, K. breviceps, and Mesoplodon europaeus, suggesting slightly lower antibody specificity and potency for immunoglobulins of these three species. Concomitantly, the IgG-HRP conjugate reacted against the heavy and light chains of IgG of the 17 species in WBs. Estimation of the cross-reactivity against heavy-chain bands confirmed the immunodiffusion assay for Kogia spp., demonstrating lower values in species of this genus (Fig. 1B). Two exceptions in the WB reactivity against the Ig-HRP conjugate were the IgG heavy chains of M. europaeus and Stenella longirostris; the former displaying a good cross-reactivity in WB and the latter reacting weaker than other species of the same genus (Fig. 1B). Neither the purified antibody nor the corresponding IgG-HRP conjugate showed cross-reactivity against bovine (Fig. 1A and B), common seal, goat, human, or dog immunoglobulins (not shown). Commercial anti-IgG(H+L)-HRP from T. truncatus (Bethyl Laboratories, Inc.) also demonstrated broad cross-reaction against Odontoceti sera with iELISA (not shown).
It has been shown that the LPS of the Brucella marine strains share the epitopes and extensively cross-react with LPS from other smooth brucellae (4). Consequently, different concentrations of Brucella AC and MC LPS types or equivalent mixtures of both LPSs were assayed. Accordingly, it was determined that the optimal conditions corresponded to 10 μg/ml of purified MC type LPS, a rabbit IgG-HRP conjugate concentration of 10 ng/ml, and a dolphin serum dilution of 1/1,000.
The gELISA has extensively been used for the diagnosis on Brucella infections in marine mammals (28, 46, 48, 49), and the interaction of protein G to antibodies is known to depend on pH conditions (protein G package manual; Pierce Biotechnology). Accordingly, we explored the binding of protein G-HRP against odontocete immunoglobulins in the two recommended buffers and compared it to the binding of bovine antibodies (Fig. 2A). While the antibodies of Brucella-positive bovine serum (dilutions from 1/250 to 1/4,000) efficiently bound protein G-HRP in gELISA using PBS-Tween (pH 7.2) or 0.1 M acetate buffer (pH 5), the immunoglobulins of Brucella-infected odontocete (serum dilution of 1/250) did not bind protein G-HRP using PBS-Tween buffer. Moreover, the kinetics of the protein G-HRP binding to odontocete antibodies in gELISA was slower than that of bovine antibodies. This was not a result of reactions of bovine sera to LPS being stronger than that of dolphin sera, since both Brucella-infected animals showed the same RBT agglutination titer (1/16) and similar optical density readings (∼1.6) when anti-bovine IgG-HRP or anti-dolphin IgG-HRP conjugates were used in iELISA. Consequently, all gELISAs were performed using 0.1 M acetate (pH 5) buffer conditions.
FIG. 2.
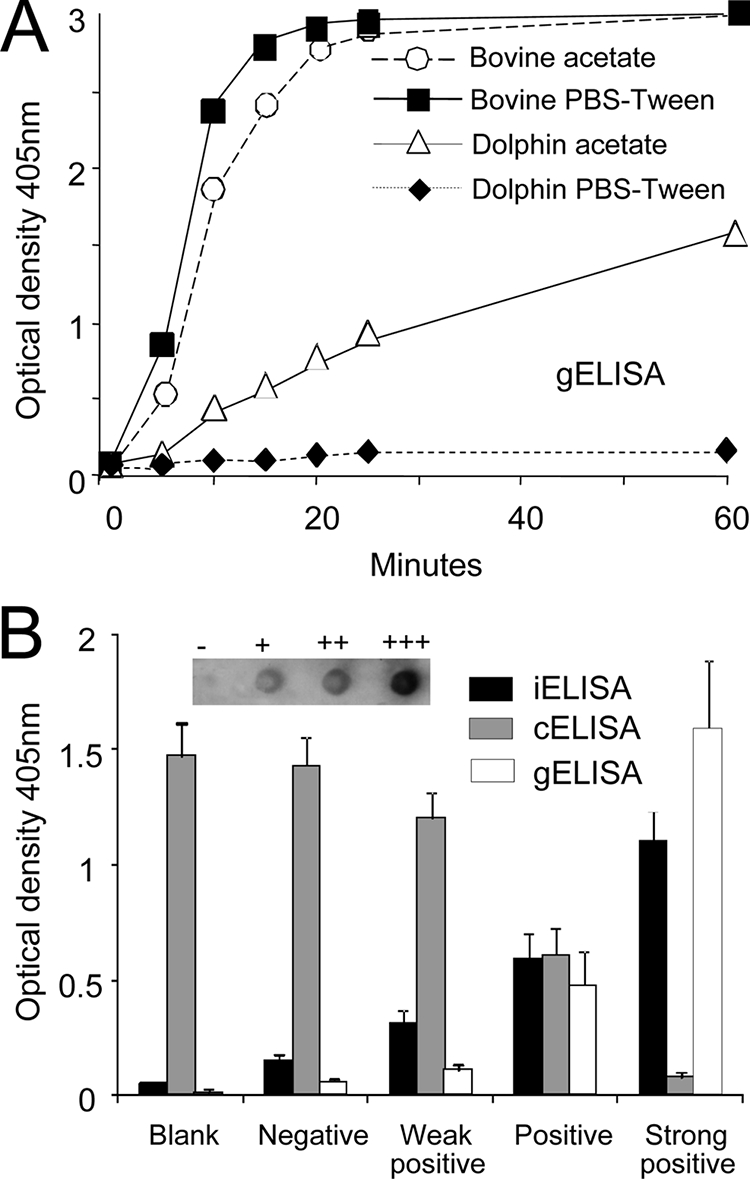
Performance of protein G-HRP against odontocete sera and calibration of positive and negative controls. (A) The performance of protein G-HRP in gELISA against sera from Brucella-infected bovine (Bos taurus) and dolphin (S. coeruleoalba) was tested using two buffer conditions: PBS-Tween (pH 7.7) and 0.1 M acetate buffer (pH 5). The standard error at all points was <5% of the values. (B) Serum samples from dolphins with high (+++), middle (++), low (+), and negative (−) optical density readings were selected as controls for iELISA and cELISA. All positive serum samples were agglutinated by RBT. The standard deviations were derived from eight independent assays in the course of 1 month of monitoring. The DB reaction is included as an in-set image indicating the intensity of the reactions. Values of the three positive and one negative serum samples did not significantly depart from the mean and maintained nonoverlapping limits of variation during 1 month of monitoring in the three ELISAs.
Then, we adjusted the positive and negative odontocete serum samples to the different ELISAs. The optical density values of the positive and negative serum samples did not significantly depart from the mean and maintained nonoverlapping limits of variation during 1 month of monitoring. Taking into account confidence levels of 99%, the upper and lower limits of variation for the ELISAs were close to 10%. Serum samples from odontocetes with high, middle, or low optical density readings were selected as positive controls to be used throughout the experiments in the different serological assays (Fig. 2B). As expected, the intensities of the spots of the controls in DB analysis were the same as those of the ELISAs (Fig. 2B). When the proportionality of the three ELISAs' reactions were determined, it was found that negative serum samples varied by less than 0.02 optical density unit in the corresponding serial dilutions (not shown) and the values of five positive serum samples tested steadily decreased in the iELISA and gELISA (Fig. 3A and B) and increased in the cELISA (Fig. 3C). However, not all the positive serum samples followed the same kinetics. For instance, in the iELISA and gELISA, the two positive serum samples (both from S. coeruleoalba) displaying higher optical densities at lower dilutions showed very different profiles after subsequent dilutions, suggesting that significant variations in avidity may occur in sera from infected animals. Moreover, while the relative positions of some serum samples remained the same in the three ELISAs, the positions of other serum samples did not (Fig. 3).
FIG. 3.
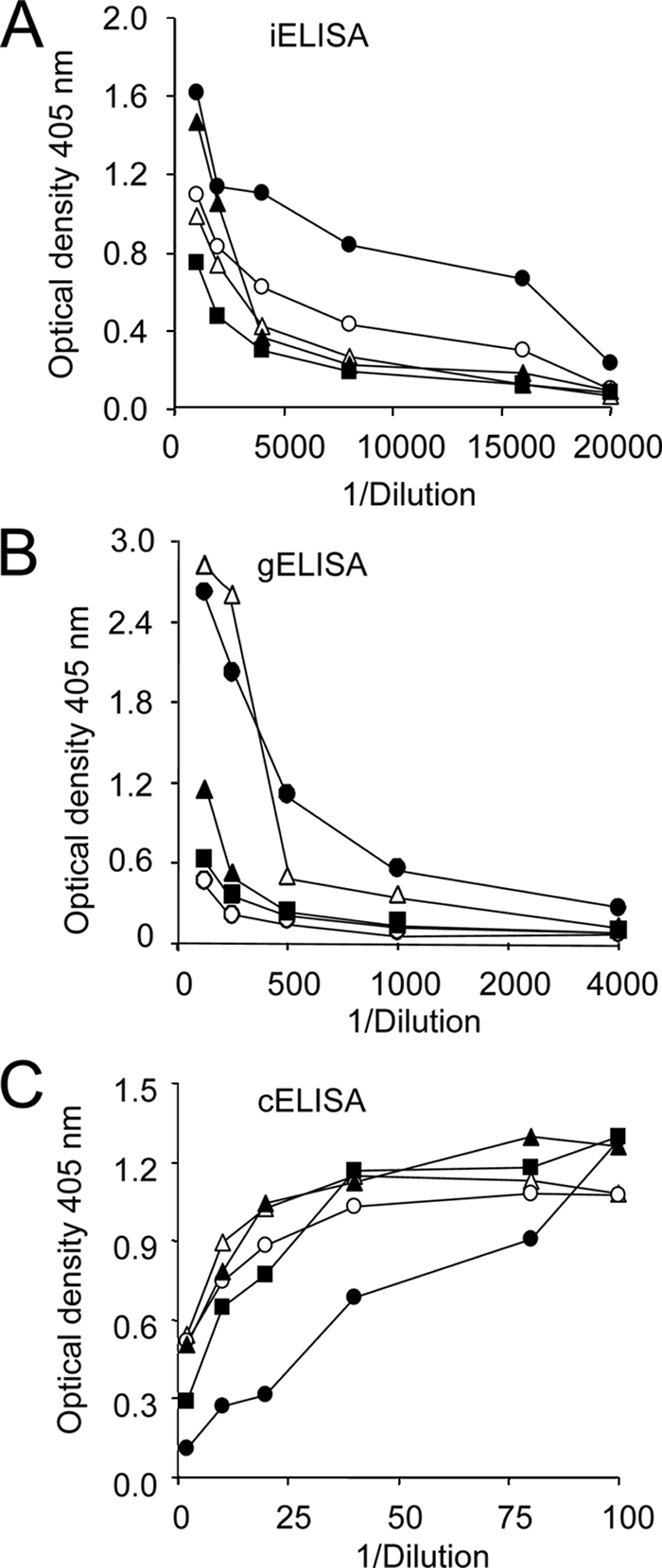
The proportionality of ELISA readings was determined for one culture-positive T. truncatus (white triangles) and four culture-positive S. coeruleoalba serum samples. Serum samples were diluted and tested by ELISA until the optical density did not vary significantly and numbers approached background values of six RBT negative serum samples obtained from resident healthy animals (two S. bredanensis, three T. truncatus, and one S. coeruleoalba). (A) Optical density iELISA readings. The standard deviation at all points was less than 5%. (B) gELISA optical density readings. The standard deviation at all points was less than 5%. (C) cELISA optical density readings. The standard deviation at all points was less than 5%.
Once the performance of the assays was optimally adjusted, we proceeded to compare their diagnostic achievements. The correlation and agreement among the iELISA, gELISA, and cELISA, with respect to RBT, were evaluated in 179 odontocete serum samples from 17 different species (Fig. 4). While the correlation between the iELISA and gELISA was relatively high (Rg/i2 = 0.65), the correlations between these two ELISAs and the cELISA were significantly lower (Rc/i2 = 0.46 and Rc/g2 = 0.37). As expected, high RBT titers were associated mostly with high percentages of positivity in the three ELISAs (Fig. 5). However, one B. ceti culture-positive dolphin (S. coeruleoalba) serum sample, with 1/2 RBT agglutination titer, fell into the negative range of the gELISA (Fig. 5B). Nevertheless, this same sample clearly fell into the positive range of the iELISA and cELISA, together with the remaining six other B. ceti culture-positive serum samples. This undoubtedly indicates that Brucella infection does not necessarily correlate to high ELISA readings and suggests a lower sensitivity for the gELISA.
FIG. 4.
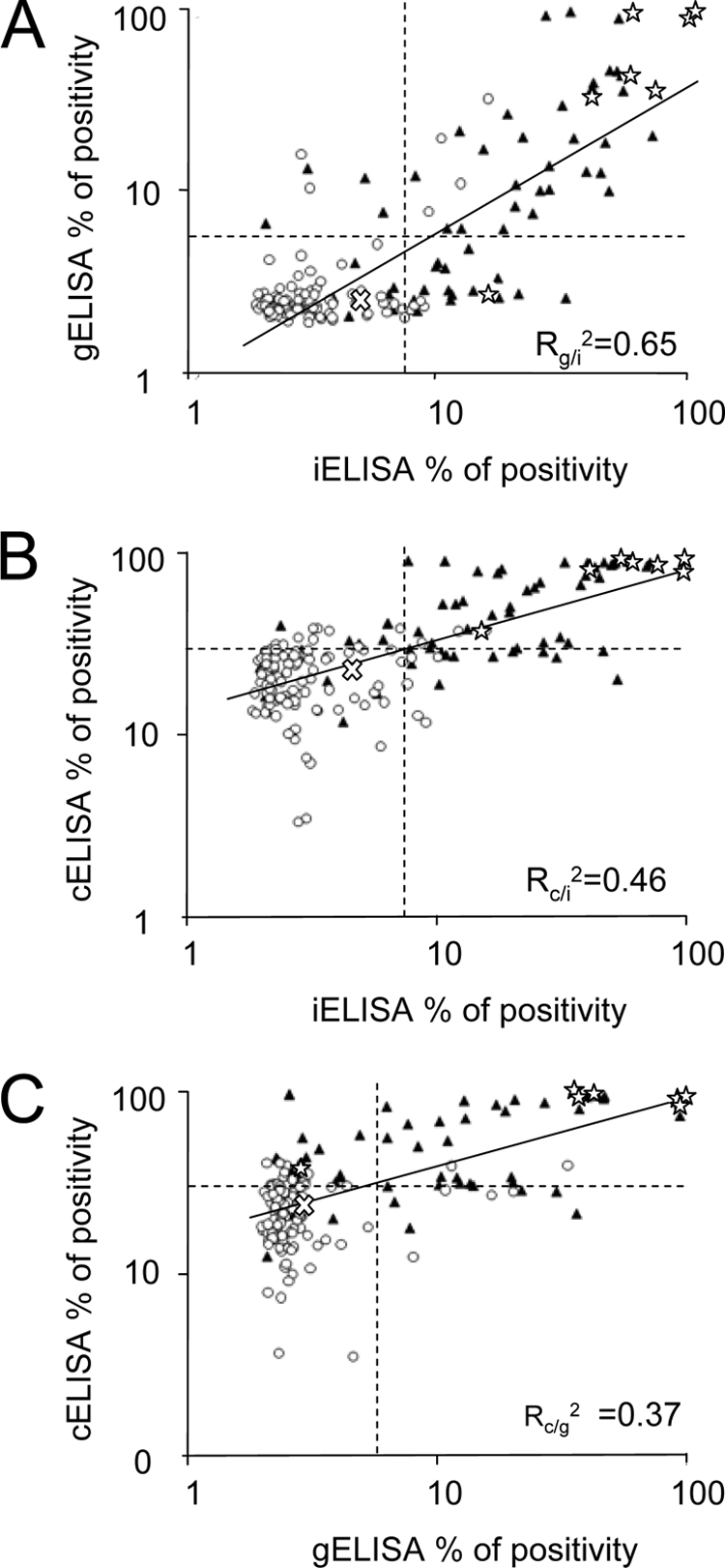
Distribution of positive and negative RBT serum samples in relation to ELISA values and correlation among the three different ELISAs. A total of 179 serum samples from 17 different species of odontocetes were tested by iELISA, gELISA, cELISA, and RBT. (A) Correlation between iELISA and gELISA. (B) Correlation between iELISA and cELISA. (C) Correlation between gELISA and cELISA. The cutoff values indicated by the dotted lines correspond to 7.5%, 6.0%, and 30.6% positivity for iELISA, gELISA, and cELISA, respectively. White circles represent RBT-negative reactions, black triangles RBT-positive reactions, stars serum samples from culture-positive dolphins (all with positive RBT reactions), and the white “X” the maximum value obtained by a negative control serum in the ELISAs. The best-fit correlation line (solid line) and the respective R2 values are indicated. Notice that one B. ceti culture-positive serum sample lies within the negative values of gELISA.
FIG. 5.
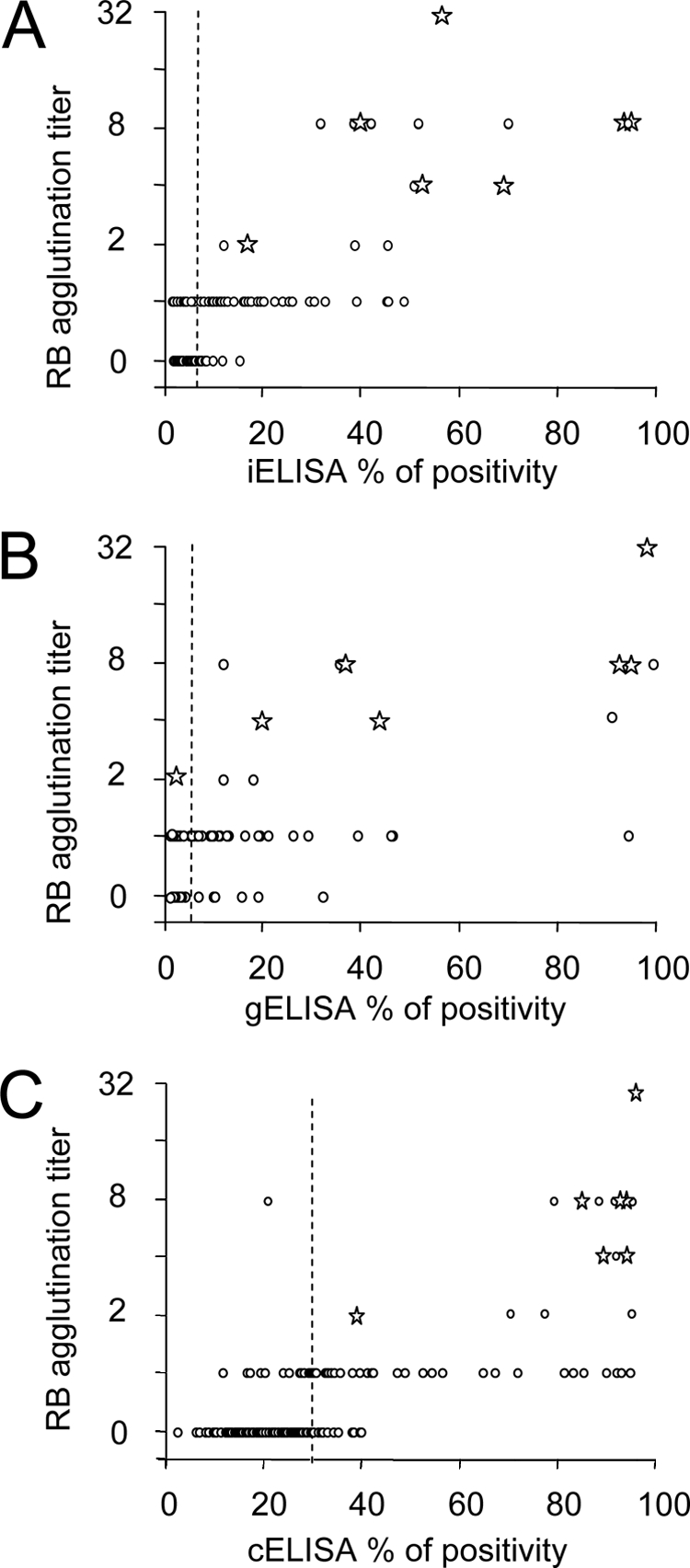
Relationships between ELISA values and RBT titers. A total of 179 serum samples from 17 different species of odontocetes were tested by iELISA, gELISA, cELISA, and RBT. (A) Relationship between RBT titers and iELISA. (B) Relationship between RBT titers and gELISA. (C) Relationship between RBT titers and cELISA. White circles correspond to RBT titles, dotted lines indicate cutoff values, and the stars correspond to serum samples from culture-positive dolphins.
The predicted specificities and sensitivities for the ELISAs were estimated by ROC analysis, using the RBT as the reference (Fig. 6). The theoretical cutoff of 30.7% for the cELISA and 5.9% for the gELISA, corresponding to the intersections of higher sensitivities and specificities on the ROC curves, matched up with the empirically established values of 30.6% and 6.0%, respectively. However, this was not the case for the iELISA, in which the empirically established cutoff value (7.5%) favored specificity over sensibility, barely affecting the kappa value. Indeed, according to the estimated kappa values, the performance of iELISA was superior to the performances of gELISA and cELISA in all comparisons (Table 2). Moreover, the smoother transition of the ROC curve and the narrower distribution of the assigned positive reactions observed in the iELISA compared to those of the gELISA and cELISA also indicated superior performance of the former. As expected, the highest kappa values were obtained when the three assays were used as the reference; however, the number of cumulative tested serum samples (consistently positive or negative for the combined tests) was reduced from 179 to 130, indicating that the performance of the IF and DB assays is low compared to that of the iELISA. The fact that the same anti-dolphin antibody was used in iELISA, IF, and DB biased the outcome in favor of the iELISA, when the latter two assays were included as reference tests.
FIG. 6.

Determination of ELISAs' sensitivities and specificities, using RBT as the reference assay. Cutoff values expressed as percentages of positivity were theoretically determined on ROC curves according to values approaching the highest sensitivity and specificity with reference to predictive percentages. Kappa values (κ) for each experiment are indicated. Black bars designate negative serum samples, and white bars designate positive serum samples. Se, sensitivity; Sp, specificity.
TABLE 2.
Kappa values of diagnostic tests for detection of anti-Brucella antibodies in odontocetes
| Test | Kappa values witha:
|
||||||||
|---|---|---|---|---|---|---|---|---|---|
| iELISA (n = 179) | cELISA (n = 179) | gELISA (n = 178) | RBT (n = 179) | IF (n = 179) | DB (n = 179) | IF + RBT (n = 147) | IF + DB (n = 149) | IF + DB + RBT (n = 130) | |
| iELISA | 0.617 | 0.661 | 0.763 | 0.758 | 0.631 | 0.939 | 0.833 | 0.950 | |
| cELISA | 0.417 | 0.638 | 0.578 | 0.519 | 0.755 | 0.663 | 0.799 | ||
| gELISA | 0.607 | 0.527 | 0.440 | 0.708 | 0.586 | 0.722 | |||
| RBT | 0.619 | 0.597 | 0.735 | ||||||
| IF | 0.664 | ||||||||
| DB | 0.759 | ||||||||
The number of samples in the columns combining assays corresponds to a cumulative value from which both samples are consistent in the result (either all positives or all negatives).
On the one hand, the cELISA and gELISA do not depend upon anti-dolphin conjugates and use either monoclonal antibody-HRP reacting against the Brucella LPS common epitope to compete (cELISA) or protein G-HRP, which is a “universal” heavy-chain IgG binding protein (17, 44); on the other hand, the HRP conjugate antibody raised against S. bredanensis displayed lower reactivity against Kogia sp. immunoglobulins (Fig. 1). Therefore, it was necessary to compare the performances of the ELISAs against both types of sera. As shown in Fig. 7, the iELISA displayed higher kappa predictive values, whether or not the serum samples belonged to Kogia spp. or to the homologous S. bredanensis, than those of gELISA and cELISA. Decreasing Kogia sp. serum dilutions onefold slightly augmented the optical densities of the assigned positives without increasing the background or the optical density of the negatives (not shown). Finally, the HRP conjugate also delivered high iELISA readings (percent positivity of >82%) with the heterologous serum samples from six culture-positive S. coeruleoalba and one T. truncatus (Fig. 4), supporting the broad cross-reaction of this anti-IgG antibody against odontocete immunoglobulins.
FIG. 7.
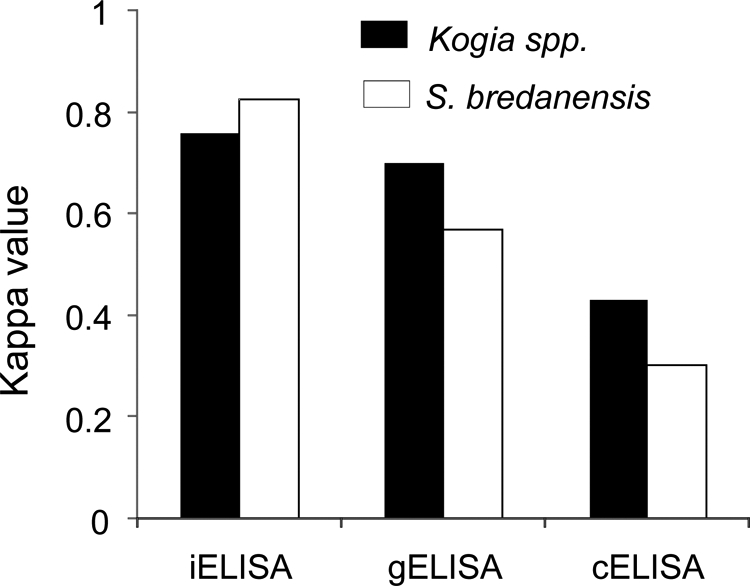
Comparative performances of iELISA, gELISA, and cELISA with serum samples from Kogia spp. and S. bredanensis, using RBT as the reference.
The frequency of assigned positive reactions in the different serological assays was determined for 179 serum samples from 17 odontocete species (Table 1). The most frequent positive serological reactions were found in members of the genera Stenella, Steno, and Tursiops, but no clear correlation to sex or age was observed (Table 3). The highest proportion of positive reactions was observed with the DB test, followed by IF. Thirteen dolphins out of 15 diagnosed with meningoencephalitis were considered positive in all reactions. Cases of meningoencephalitis were conspicuous in S. coeruleoalba (10 out of 11 reactors), six from which Brucella was isolated from brain fluids (25).
TABLE 3.
Distribution of positive serum samples according to sex and age in different odontocetes
| Assay | No. of positive samples
|
|||||
|---|---|---|---|---|---|---|
| Sex
|
Age
|
|||||
| Female (n = 84) | Male (n = 95) | Adult (n = 85) | Juvenile (n = 48) | Calf (n = 24) | Unknown (n = 22) | |
| iELISA | 37 | 21 | 35 | 15 | 7 | 1 |
| cELISA | 32 | 26 | 37 | 14 | 2 | 5 |
| gELISA | 29 | 16 | 26 | 13 | 2 | 4 |
| RBT | 38 | 25 | 33 | 21 | 4 | 5 |
| IF | 43 | 28 | 40 | 20 | 3 | 8 |
| DB | 51 | 38 | 48 | 23 | 4 | 14 |
DISCUSSION
Brucellosis in cetaceans has been shown to be a significant cause of strandings and a potential zoonosis (19, 25). Therefore, the rapid and correct identification of infected animals becomes a relevant practice for many marine wildlife services, aquariums, and fisheries. Agglutination tests, cELISA, and gELISA have been used for the detection of antibodies in cetaceans and for presumptive diagnosis of Brucella infections. One study performed on Pacific bottlenose dolphins from the Solomon Islands (46) reported up to 70% antibody prevalence using gELISA, while others using the same test reported between 8 and 14% prevalence in three species of mysticetes from the North Atlantic Ocean (48). In a retrospective study of 593 North American odontocetes of five species and four mysticetes of three species, the prevalence of antibodies corresponded to 4.1% and 5.5%, respectively, as determined by cELISA (36). Serum samples from 76 odontocetes of five species collected in Peruvian fisheries and from strandings on the Spanish coast revealed 41% prevalence, estimated by cELISA and gELISA (49). Similarly, Jepson et al. (28) reported 35.3% antibody prevalence in 68 serum samples from five different species of odontocetes from northern Europe, using cELISA and gELISA. The a priori rationale for using these assays lies in the fact that they do not depend upon specific antibody conjugates directed against immunoglobulins from the species to be explored. Although these works are valuable in many aspects, none of them have critically evaluated the performance of these serological assays in cetaceans. As stated before, one of the problems confronted is the absence of a significant collection of negative and positive serum samples from cetaceans that could serve to determine the specificity and sensitivity of immunoassays.
The advantages of RBT as the reference test in naturally infected populations rest in its simplicity, consistency, and straightforward interpretation (1). The RBT false-positive reactions are commonly due to the use of plasma instead of sera, heavily hemolyzed sera, or cross-reactions; and the false-negative results usually correspond to overall low avidity or reduced titers of agglutinating antibodies. In spite of these drawbacks, the RBT remains robust in the absence of other independent assays, and its use as the reference is justified (1, 29). Moreover, the fact that the seven serum samples from B. ceti culture animals presented agglutination titers of ≥1/2 provided additional support for the use of RBT as the reference assay and a starting point for understanding the serological reactions in Brucella-infected cetaceans.
The “false-negative” reactions observed in iELISA and gELISA, could be due to (i) the presence of small amounts of IgM or IgA agglutinating antibodies that may escape detection of anti-IgG-HRP and protein G-HRP, (ii) agglutinating antibodies directed against non-LPS epitopes (e.g., proteins) that are precluded in the ELISAs, or (iii) both. However, in contrast to protein G, which primarily binds the heavy chain of IgGs, the anti-dolphin IgG used in the iELISA also reacts against the light chain, increasing the probability for detecting other immunoglobulin isotypes and, therefore, increasing the serological sensitivity. The lower sensitivity of cELISA in relation to that of iELISA has been documented (29), and it has been connected to the displacement of low-avidity antibodies by the competing monoclonal antibody against the C epitope of LPS used in this test.
The “false-positive” reactions detected in iELISA and gELISA may be due to antibodies directed against deep Brucella LPS determinants not readily exposed in the RBT, but accessible in purified LPS used for coating the plates. Indeed, it has been demonstrated that bovines and other animals are capable of generating antibodies against deep determinants of LPS such as the core and lipid A moieties (41), which are shared by other Alphaproteobacteria related to Brucella (8, 50). Since Alphaproteobacteria, such as Bartonella species, also infect odontocetes (23), cross-reactions against these deep LPS epitopes may arise. The relatively higher specificity of cELISA seems to be linked to the competing monoclonal antibody which specifically recognizes the C epitope of the LPS. However, if the cutoff is adjusted, the iELISA displays specificity comparable to or better than that of the cELISA without reaching the low sensitivity levels of the later assay.
In spite of the relatively good correlation between the iELISA and the gELISA (Rg/i2 = 0.65), the former assay displayed a significantly higher sensitivity (SeiELISA = 86 versus SegELISA = 58). It is noteworthy that one serum sample from a B. ceti culture-positive dolphin was recorded as negative by the gELISA and a significant number of RBT positive serum samples that tested as positive were also marked as negatives in the gELISA, suggesting that protein G-HRP failed to detect lower avidity or reduced antibody titers. Moreover, caution with the buffer system has to be taken at the time of using protein G-HRP for the diagnosis of brucellosis in odontocetes, since regular neutral conditions do not work. In this regard, it may be that the use of an acidic pH buffer system contributes to the reduced sensitivity of the gELISA, since low-avidity antibodies may be displaced. In small ruminants, the gELISA has been shown to be less sensitive than ELISAs that use anti-IgG conjugates or other serological tests for the diagnosis of Brucella infections (9, 12). Therefore, the unrestricted use of gELISA for odontocete brucellosis testing calls for attention, since its sensitivity seems to be lower in these animals than in bovines.
Taking into account the performances of iELISA, gELISA, and cELISA in cattle (17, 30, 44), the correlation and agreement among these three tests for odontocete sera were much lower than predicted. This result was not anticipated, and generally, the three assays display relatively good correlation in bovines (17, 10, 30, 44), and the RBT test commonly displays similar agreements with the three ELISAs for the detection of anti-Brucella antibodies (kappa values ranging between 0.70 and 0.75). In our hands, the iELISA was highly reproducible and performed very similarly to what has been observed in cattle using the RBT. On the contrary, our experience using bovine sera has shown us that the cELISA results are more variable from test to test and the assay is very sensitive to the conditions of test sera (including storage), conjugate, and controls than are the iELISA results. Although the last two variables can be controlled, the quality of sera depends upon collection, an event that is not always controlled at the time of bleeding stranded odontocetes (25). The variability of the cELISA is somewhat illustrated by the irregular shape of the ROC curve and in the broader distribution of positive and negative odontocete serum samples in comparison to the iELISA. Alternatively, it may also be that the overall avidity of cetacean antibodies against Brucella LPS works differently in the various ELISAs, accounting for these discrepancies, or both. To some extent, this phenomenon was also illustrated by the different relative locations of positive serum on the titration curves (Fig. 3). Finally, the interpretation of IF and DB tests, mainly with serum samples possessing borderline reactions, were less obvious than the ELISAs and RBT. This fact may explain the higher proportion of “false-positive” reactions observed with DB and IF tests than that observed with iELISA, in spite of using the same anti-dolphin IgG conjugate.
In bovine, caprine, or ovine brucellosis the source of smooth Brucella LPS seems to have little relevance for diagnostic purposes (34). In the case of odontocete sera, it seems that MC type LPS performed slightly better than did AC type LPS in the iELISA, with respect to time of reading, optical densities, and differences between positive and negative sera. However, the final “positive” or “negative” results using AC type LPS instead of MC type LPS did not significantly diverge, except in some borderline readings (not shown). It has been demonstrated that the density and content of the common epitopes in B. ceti LPS vary with respect to the classical LPSs from terrestrial smooth brucellae (4). This factor may influence the generation of antibodies in Brucella-infected odontocetes, which may recognize more readily the common epitopes in MC than in AC type LPSs. However, we doubt that this variable accounts for the significant differences observed between the various ELISAs. In any case, our results also illustrate the relevance of testing the Brucella LPS antigen for the standardization of immunoenzymatic assays in cetaceans.
It has been demonstrated that antibodies made against Bos taurus extensively cross-react with immunoglobulins of other Bovinae and, therefore, are useful in the serological testing and disease diagnosis (including brucellosis) of other species of this subfamily (24). As shown here and by others (38), in odontocetes, the cross-reactivity of immunoglobulins is extensive, thereby allowing the use of a single species anti-IgG antibody for discriminating the presence of anti-Brucella LPS antibodies. The fact that kappa values of the iELISA were considerably higher than those of the gELISA and cELISA regardless of whether the serum sample tested belonged to the homologous species (S. bredanensis) or to the heterologous species (Kogia spp.) reinforced this concept. In spite of the fact that the cELISA does not depend upon an anti-odontocete IgG conjugate, the agreement with RBTs was lower with Kogia sp. and S. bredanensis serum samples than that observed in gELISA and iELISA. Moreover, the anti-odontocete IgG conjugate showed good reactivity against Physeter macrocephalus (sperm whale), a member of the Physeteridae family, which is phylogenetically closer to Kogiidae than to Delphinidae. Based on this, the prediction is that this broadly cross-reacting antibody would detect immunoglobulins from other odontocetes, including those of the Monodontidae (narwhals) and the fresh water dolphins from the Iniidae, Pontoporiidae, Lipotidae, and Platanistidae families. Indeed, all these families are phylogenetically closer to the family Delphinidae, of which S. bredanensis is a member (37). Regarding the very low cross-reactivity between odontocete immunoglobulins and gray whale (Eschrichtius robustus) and humpback whale (Megaptera novaeangliae) antibodies (38), it is unlikely that this same anti-dolphin IgG would work for Mysticeti (baleen whales), an order that belongs to a different phylogenetic branch among cetaceans (37). Finally, based on limited experience with commercial anti-dolphin IgG(H+L) antibodies, we predict that extensive cross-reactions with odontocete sera will be observed with these reagents. Therefore, they may be practical for establishing straightforward iELISAs and other diagnostic tests.
One significant observation was the correlation between the high titers of antibodies against Brucella antigens and the presence of neurological symptoms in dolphins. Out of a total of 15 dolphins that stranded with diagnoses of neurological pathology, 13 displayed positive RBT and iELISA values above 85% positivity. Among these, 10 were S. coeruleoalba, three were T. truncatus, and seven of these animals were B. ceti culture positive. In one T. truncatus, the isolation was performed retrospectively from frozen tissues stored at −80C° after serological diagnosis, sustaining the potential predictive value of iELISA and RBT assays, mainly when they are used in concert.
Acknowledgments
We thank José-María Blasco, Gobierno de Aragón, Spain, for his advice in the standardization of the iELISA; Juan-José Romero from Escuela de Medicina Veterinaria, Universidad Nacional, Costa Rica, for recommending the suitable statistical package; and Frances Gulland from The Marine Mammal Center, Sausalito, CA, for the seal samples. We also thank Fundación KETO for the logistic support during this investigation.
This work was funded by grants from FIDA-2006, from the Universidad Nacional, FS-CONARE UNA/UCR, Costa Rica, NeTropica 8-N-2005, and from MICIT/CONICIT of Costa Rica.
Footnotes
Published ahead of print on 22 April 2009.
REFERENCES
- 1.Alton, G. G., L. M. Jones, R. D. Angus, and J. M. Verger. 1988. Techniques for the brucellosis laboratory. Institut National de la Recherche Agronomique, Paris, France.
- 2.Antonioli, C., and M. A. Reveley. 2005. Randomised controlled trial of animal facilitated therapy with dolphins in the treatment of depression. BMJ 3311231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aragón, V., R. Díaz, E. Moreno, and I. Moriyón. 1996. Characterization of Brucella abortus and Brucella melitensis native haptens as outer membrane O-type polysaccharides independent from smooth lipopolysaccharide. J. Bacteriol. 1781070-1079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Baucheron, S., M. Grayon, M. S. Zygmunt, and A. Cloeckaert. 2002. Lipopolysaccharide heterogeneity in Brucella strains isolated from marine mammals. Res. Microbiol. 153277-280. [DOI] [PubMed] [Google Scholar]
- 5.Brew, S. D., L. L. Perrett, J. A. Stack, and A. P. MacMillan. 1999. Human exposure to Brucella recovered from a sea mammal. Vet. Rec. 144483. [PubMed] [Google Scholar]
- 6.Clavareau, C., V. Wellemans, K. Walravens, M. Tryland, J. M. Verger, M. Grayon, A. Cloeckaert, J. J. Letesson, and J. Godfroid. 1998. Phenotypic and molecular characterization of a Brucella strain isolated from a minke whale (Balaenoptera acutorostrata). Microbiology 1443267-3273. [DOI] [PubMed] [Google Scholar]
- 7.Cloeckaert, A., J. M. Verger, M. Grayon, J. Y. Paquet, B. Garin-Bastuji, G. Foster, and J. Godfroid. 2001. Classification of Brucella spp. isolated from marine mammals by DNA polymorphism at the omp2 locus. Microbes Infect. 3729-738. [DOI] [PubMed] [Google Scholar]
- 8.Delpino, M. V., C. A. Fossati, and P. C. Baldi. 2004. Occurrence and potential diagnostic applications of serological cross-reactivities between Brucella and other alpha-proteobacteria. Clin. Diagn. Lab. Immunol. 11868-873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Díaz-Aparicio, E., C. Marín, B. Alonso-Urmeneta, V. Aragón, S. Pérez-Ortiz, M. Pardo, J. M. Blasco, R. Díaz, and I. Moriyón. 1994. Evaluation of serological tests for diagnosis of Brucella melitensis infection of goats. J. Clin. Microbiol. 321159-1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Díaz-Aparicio, E., V. Aragón, C. Marín, B. Alonso, M. Font, E. Moreno, S. Pérez-Ortiz, J. M. Blasco, R. Díaz, and I. Moriyón. 1993. Comparative analysis of Brucella serotype A and M Yersinia enterocolitica O:9 polysaccharides for serological diagnosis of brucellosis in cattle, sheep and goats. J. Clin. Microbiol. 313136-3141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ewalt, D. R., J. B. Payeur, B. M. Martin, D. R. Cummins, and W. G. Miller. 1994. Characterization of a Brucella species from a bottlenose dolphin (Tursiops truncatus). J. Vet. Diagn. Investig. 6448-452. [DOI] [PubMed] [Google Scholar]
- 12.Ficapal, A., B. Alonso-Urmeneta, J. Velasco, I. Moriyón, and J. M. Blasco. 1995. Diagnosis of Brucella ovis infection of rams with an ELISA using protein G as conjugate. Vet. Rec. 137145-147. [DOI] [PubMed] [Google Scholar]
- 13.Forbes, L. B., O. Nielsen, L. Measures, and D. R. Ewalt. 2000. Brucellosis in ringed seals and harp seals from Canada. J. Wildl. Dis. 36595-598. [DOI] [PubMed] [Google Scholar]
- 14.Foster, G., A. P. MacMillan, J. Godfroid, F. Howie, H. M. Ross, A. Cloeckaert, R. J. Reid, S. Brew, and I. A. Patterson. 2002. A review of Brucella spp. infection of sea mammals with particular emphasis on isolates from Scotland. Vet. Microbiol. 90563-580. [DOI] [PubMed] [Google Scholar]
- 15.Foster, G., B. S. Osterman, J. Godfroid, I. Jacques, and A. Cloeckaert. 2007. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol. 572688-2693. [DOI] [PubMed] [Google Scholar]
- 16.Foster, G., K. L. Jahans, R. J. Reid, and R. M. Ross. 1996. Isolation of Brucella species from cetaceans, seals and an otter. Vet. Rec. 138583-586. [DOI] [PubMed] [Google Scholar]
- 17.Gall, D., A. Colling, O. Mariño, E. Moreno, K. Nielsen, B. Pérez, and L. Samartino. 1998. Enzyme immunoassays for the serological diagnosis of bovine brucellosis. Trial in Latin America. Clin. Diagn. Lab. Immunol. 5654-661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Garner, M. M., D. M. Lambourn, S. J. Jeffries, P. B. Hall, J. C. Rhyan, D. R. Ewalt, L. M. Polzin, and N. F. Cheville. 1997. Evidence of Brucella infection in Parafilaroides lungworms in a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn. Investig. 9298-303. [DOI] [PubMed] [Google Scholar]
- 19.González, L., I. A. Patterson, R. J. Reid, G. Foster, M. Barberán, J. M. Blasco, S. Kennedy, F. E. Howie, J. Godfroid, A. P. MacMillan, A. Schock, and D. Buxton. 2002. Chronic meningoencephalitis associated with Brucella sp. infection in live-stranded striped dolphins (Stenella coeruleoalba). J. Comp. Pathol. 126147-152. [DOI] [PubMed] [Google Scholar]
- 20.Groussaud, P., S. J. Shankster, M. S. Koylass, and A. M. Whatmore. 2007. Molecular typing divides marine mammal strains of Brucella into at least three groups with distinct host preferences. J. Med. Microbiol. 561512-1518. [DOI] [PubMed] [Google Scholar]
- 21.Hanley, J. A., and B. J. McNeil. 1983. A method of comparing the areas under receiver operating characteristic curves derived from the same cases. Radiology 148839-843. [DOI] [PubMed] [Google Scholar]
- 22.Harlow, E., and D. Lane. 1988. Antibodies: a laboratory manual, 1st ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
- 23.Harms, C. A., R. G. Maggi, E. B. Breitschwerdt, C. L. Clemons-Chevis, M. Solangi, D. S. Rotstein, P. A. Fair, L. J. Hansen, A. A. Hohn, G. N. Lovewell, W. A. McLellan, D. A. Pabst, T. K. Rowles, L. H. Schwacke, F. I. Townsend, and R. S. Wells. 2008. Bartonella species detection in captive, stranded and free-ranging cetaceans. Vet. Res. 3959. [DOI] [PubMed] [Google Scholar]
- 24.Henning, D., and K. Nielsen. 1992. Cross-reactivity of monoclonal antibodies to bovine immunoglobulins with immunoglobulins of other species. Vet. Immunol. Immunopathol. 34235-243. [DOI] [PubMed] [Google Scholar]
- 25.Hernández-Mora, G., R. González-Barrientos, J. A. Morales, E. Chaves-Olarte, C. Guzmán-Verri, E. Barquero-Calvo, M. J. De-Miguel, C. M. Marín, J. M. Blasco, and E. Moreno. 2008. Neurobrucellosis in stranded dolphins, Costa Rica. Emerg. Infect. Dis. 141430-1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hudson, L., and F. C. Hay. 1976. Practical immunology. Blackwell Scientific, Oxford, England.
- 27.Jahans, K. L., G. Foster, and E. S. Broughton. 1997. The characterisation of Brucella strains isolated from marine mammals. Vet. Microbiol. 57373-382. [DOI] [PubMed] [Google Scholar]
- 28.Jepson, P. D., S. Brew, A. P. MacMillan, J. R. Baker, J. Barnett, J. K. Kirkwood, T. Kuiken, I. R. Robinson, and V. R. Simpson. 1997. Antibodies to Brucella in marine mammals around the coast of England and Wales. Vet. Rec. 141513-515. [DOI] [PubMed] [Google Scholar]
- 29.Mainar-Jaime, R. C., P. M. Muñoz, M. J. de Miguel, M. J. Grilló, C. M. Marín, I. Moriyón, and J. M. Blasco. 2005. Specificity dependence between serological tests for diagnosing bovine brucellosis in Brucella-free farms showing false positive serological reactions due to Yersinia enterocolitica O:9. Can. Vet. J. 46913-916. [PMC free article] [PubMed] [Google Scholar]
- 30.Marín, C. M., E. Moreno, I. Moriyón, R. Díaz, and J. M. Blasco. 1999. Performance of competitive and indirect enzyme-linked immunosorbent assay, gel immunoprecipitation with native hapten polysaccharide and standard serological tests in the diagnosis of sheep brucellosis. Clin. Diagn. Lab. Immunol. 6269-272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.May-Collado, L. J., and I. Agnarsson. 2006. Cytochrome b and Bayesian inference of whale phylogeny. Mol. Phylogenet. Evol. 38344-354. [DOI] [PubMed] [Google Scholar]
- 32.McDonald, W. L., R. Jamaludin, G. Mackereth, M. Hansen, S. Humphrey, P. Short, T. Taylor, J. Swingler, C. E. Dawson, A. M. Whatmore, E. Stubberfield, L. L. Perrett, and G. Simmons. 2006. Characterization of a Brucella sp. as a marine mammal type isolated from a New Zealand patient with spinal osteomyelitis. J. Clin. Microbiol. 444363-4370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.McKinney, M. M., and A. Parkinson. 1987. A simple non-chromatographic procedure to purify immunoglobulins from serum and ascites fluid. J. Immunol. Methods 96271-278. [DOI] [PubMed] [Google Scholar]
- 34.Moreno, E., and I. Moriyón. 2006. The genus Brucella, p. 315-456. In M. Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (ed.), The prokaryotes, vol. 5. Springer-Verlag, New York, NY. [Google Scholar]
- 35.Moreno, E., M. W. Pitt, L. M. Jones, G. G. Schuring, and D. T. Berman. 1979. Purification and characterization of smooth and rough lipopolysaccharides from Brucella abortus. J. Bacteriol. 138361-369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nielsen, O., R. E. Stewart, K. Nielsen, L. Measures, and P. Duignan. 2001. Serologic survey of Brucella spp. antibodies in some marine mammals of North America. J. Wildl. Dis. 3789-100. [DOI] [PubMed] [Google Scholar]
- 37.Nikaido, M., F. Matsuno, H. Hamilton, R. L. Brownell, Jr., Y. Cao, W. Ding, Z. Zuoyan, A. M. Shedlock, R. E. Fordyce, M. Hasegawa, and N. Okada. 2001. Retroposon analysis of major cetacean lineages: the monophyly of toothed whales and the paraphyly of river dolphins. Proc. Natl. Acad. Sci. USA 987384-7389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nollens, H. H., C. Ruiz, M. T. Walsh, F. M. Gulland, G. Bossart, E. D. Jensen, J. F. McBain, and J. F. Wellehan. 2008. Cross-reactivity between immunoglobulin G antibodies of whales and dolphins correlates with evolutionary distance. Clin. Vaccine Immunol. 151547-1554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Page, M., and R. Thorpe. 1998. IgG purification. Methods Mol. Biol. 8095-111. [DOI] [PubMed] [Google Scholar]
- 40.Reik, L. M. 1987. A simple non-chromatographic purification procedure for monoclonal antibodies. J. Immunol. Methods 100123-130. [DOI] [PubMed] [Google Scholar]
- 41.Rojas, N., O. Zamora, J. Cascante, D. Garita, and E. Moreno. 2001. Comparison of the antibody response in adult cattle against different epitopes of Brucella abortus lipopolysaccharide. J. Vet. Med. B Infect. Dis. Vet. Public Health 48623-629. [DOI] [PubMed] [Google Scholar]
- 42.Ross, H. M., G. Foster, R. J. Reid, K. L. Jahans, and A. P. MacMillan. 1994. Brucella species infection in sea-mammals. Vet. Rec. 134359. [DOI] [PubMed] [Google Scholar]
- 43.Ross, H. M., K. L. Jahans, A. P. MacMillan, R. J. Reid, P. M. Thompson, and G. Foster. 1996. Brucella species infection in North Sea seal and cetacean populations. Vet. Rec. 138647-648. [DOI] [PubMed] [Google Scholar]
- 44.Saegerman, C., L. De Waele, D. Gilson, J. Godfroid, P. Thiange, P. Michel, B. Limbourg, T. K. Vo, J. Limet, J. J. Letesson, and D. Berkvens. 2004. Evaluation of three serum i-ELISAs using monoclonal antibodies and protein G as peroxidase conjugate for the diagnosis of bovine brucellosis. Vet. Microbiol. 10091-105. [DOI] [PubMed] [Google Scholar]
- 45.Sohn, A. H., W. S. Probert, C. A. Glaser, N. Gupta, A. W. Bollen, J. D. Wong, E. M. Grace, and W. C. Mc Donald. 2003. Human neurobrucellosis with intracerebral granuloma caused by a marine mammal Brucella spp. Emerg. Infect. Dis. 9485-488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tachibana, M., K. Watanabe, S. Kim, Y. Omata, K. Murata, T. Hammond, and M. Watarai. 2006. Antibodies to Brucella spp. in Pacific bottlenose dolphins from the Solomon Islands. J. Wildl. Dis. 42412-414. [DOI] [PubMed] [Google Scholar]
- 47.Thrusfield, M., C. Ortega, I. de Blas, J. P. Noordhuizen, and K. Frankena. 2001. Win Episcope 2.0: improved epidemiological software for veterinary medicine. Vet. Rec. 148567-572. [DOI] [PubMed] [Google Scholar]
- 48.Tryland, M., L. Kleivane, A. Alfredsson, M. Kjeld, A. Arnason, S. Stuen, and J. Godfroid. 1999. Evidence of Brucella infection in marine mammals in the North Atlantic Ocean. Vet. Rec. 144588-592. [DOI] [PubMed] [Google Scholar]
- 49.Van Bressem, M. F., K. Van Waerebeek, J. A. Raga, J. Godfroid, S. D. Brew, and A. P. MacMillan. 2001. Serological evidence of Brucella species infection in odontocetes from the South Pacific and the Mediterranean. Vet. Rec. 148657-661. [DOI] [PubMed] [Google Scholar]
- 50.Velasco, J., R. Díaz, M. J. Grilló, M. Barberán, C. Marín, J. M. Blasco, and I. Moriyón. 1997. Antibody and delayed-type hypersensitivity responses to Ochrobactrum anthropi cytosolic and outer membrane antigens in infections by smooth and rough Brucella spp. Clin. Diagn. Lab. Immunol. 4279-284. [DOI] [PMC free article] [PubMed] [Google Scholar]