

Abstract
Bacillus thuringiensis Cry toxins have been widely used in the control of insect pests either as spray products or expressed in transgenic crops. These proteins are pore forming toxins with a complex mechanism of action that involves the sequential interaction with several toxin-receptors. Cry toxins are specific against susceptible larvae and although they are often highly effective, some insect pests are not affected by them or show low susceptibility. In addition, the development of resistance threatens their effectiveness, so strategies to cope with all these problems are necessary. In this review we will discuss and compare the different strategies that have been used to improve insecticidal activity of Cry toxins. The activity of Cry toxins can be enhanced by using additional proteins in the bioassay like serine protease inhibitors, chitinases, Cyt toxins, or a fragment of cadherin receptor containing a toxin-binding site. On the other hand, different modifications performed in the toxin gene such as site directed mutagenesis, introduction of cleavage sites in specific regions of the protein, and deletion of small fragments from the amino-terminal region lead to improved toxicity or overcome resistance, representing interesting alternatives for insect pest control.
1. Introduction
Bacillus thuringiensis (Bt) bacteria produce insecticidal proteins named Cry toxins, which are effective in killing some insect pests. These toxins are used worldwide, being delivered as sprays or expressed in some transgenic crops. Cry toxins are especially useful due to their insect specificity and because they are harmless to humans. The accepted model of toxin action comprises several steps [2, 45]. The Cry1A protoxins are ingested by the larvae, solubilized in the gut lumen and cleaved by midgut proteases to yield activated 60 kDa monomeric toxins. The activated toxins bind to receptors present in the midgut cells of the larvae. In some lepidopteran larvae, cadherin-like proteins show high affinity for the toxins and function as primary receptors [50]. It was proposed that binding of 60 kDa monomeric toxins to cadherin facilitates additional protease cleavage of the N-terminal end of the toxins including helix α-1 of domain I [22]. This cleavage induces assembly of oligomeric forms of the toxin. The oligomers have increased binding affinity to the secondary receptors, which are glycosylphosphatidyl-inositol (GPI)-anchored proteins, aminopeptidase N and alkaline phosphatase [3, 30]. After the oligomers bind to secondary receptors, they insert into membrane microdomains, creating pores in the apical membrane of midgut cells, which cause osmotic shock and cell death [3, 29,45] (Fig. 1).
Figure 1.

Model of the mode of action of Cry toxins, including the different steps in toxin action that have been modified to enhance its insecticidal activity or to overcome resistance. 1, Crystal solubilization in midgut lumen. 2, Protoxin activation and translocation trough the peritrophic membrane. 3, Toxin binding to primary receptor and cleavage of helix α-1. 4, Oligomerization of the toxin and binding to secondary GPI-anchored receptors. 5, Toxin insertion into the membrane and pore formation. Some strategies used to improve toxin action are represented in this figure such as: i) Chitinase that improves accessibility of the toxin to the epithelial membrane. ii) Serine protease inhibitors that reduce degradation of Cry or of toxin-receptors. iii) Introduction of intramolecular cleavage sites in the toxin that improves binding interaction. iv) Introduction of more binding sites such as the CR12-MPE-peptide from cadherin receptor or the Cyt1A toxin. iv) Deletion of helix α-1 that induces toxin-oligomerization and skip cadherin interaction.
The primary threat to the long-term efficacy of Bt toxins is the evolution of resistance by pest insects [23, 47]. Different pest strains have been selected in laboratory conditions for resistance to Bt toxins and some pests evolved resistance to Bt sprays in the field or in green houses [16, 27, 47, 48, 56]. In addition, it was reported that Helicoverpa zea evolved some resistance to Bt cotton in Arkansas and Mississippi [49]. Understanding the mode of action of Bt toxins and the mechanisms of resistance is essential for designing strategies to improve Bt toxin action and to counter pest resistance.
2. Potentiation of Cry toxin activity by additional proteins
2.1 Serine protease inhibitors
Several serine protease inhibitors are able to potentiate the insecticidal activity of Cry toxins in three different insect orders (Coleoptera, Lepidoptera and Diptera) [24, 35]. The presence of extremely low levels of protease inhibitors enhances the insecticidal activity of some Cry toxins up to 20 fold. The serine protease inhibitors are present in legume seeds and have insecticidal effects when present at high concentrations in the insect diet probably by inhibiting larval digestion, suggesting that they could have evolved as a plant defense mechanism against herbivores [24]. It was reported that a variety of serine protease inhibitors isolated either from plants or animals potentiate the insecticidal activity of several Cry toxins against their targets when they were fed in combination [35]. The potentiation effect of the serine protease inhibitors was observed at a concentration 105 times below their insecticidal level. In addition, genetically modified tobacco plants expressing a protease inhibitor (Curcurbita maxima trypsin protease inhibitor) fused to a truncated Cry1Ab toxin showed a 6 fold increase in specific insecticidal activity against tobacco budworm (Heliothis virescens) when compared with plants expressing only Cry toxin protein [35]. How protease inhibitors enhance Bt toxin activity is not known, although they may inhibit specific gut proteases that normally would inactivate Bt toxins or may limit proteolysis of membrane proteins preventing the degradation of membrane bound receptors, therefore increasing their half-lives and their ability to bind Cry toxins (Fig. 1).
2.2 Chitinase
The larval peritrophic membrane forms a cylindrical sheet that divides the gut epithelium from the lumen. This structure functions as a physical barrier against mechanical damage and invasion of microorganisms. The solubilized Cry toxins need to cross this barrier to bind their receptors, which are located in the apical membrane of the midgut cells. The peritrophic membrane is a network of chitin embedded in a protein-carbohydrate matrix. It was shown that increasing the levels of endochitinases in the larval midgut or the addition of chitinases to Cry toxin preparations increase their efficacy and potency up to 10 fold [12, 44]. It was proposed that chitinases elevate the larvicidal effect by perforating the peritrophic membrane, increasing the accessibility of the Cry toxin to the epithelial membrane where receptors are located (Fig 1).
2.3 Cyt toxins
Cyt proteins are hemolytic and cytolytic toxins produced by some Bacillus thuringiensis strains, particularly by those that show insecticidal activity against mosquitoes. Cyt toxins are also pore-forming proteins, and they are not related phylogenetically to Cry toxins. They have a single α–β domain comprised of two outer layers of α-helix hairpins wrapped around a β-sheet [33]. Cyt toxins do not bind to protein receptors and directly interact with membrane lipids inserting into the membrane to form pores [31, 42] or destroy the membrane by a detergent like interaction [5].
The toxicity of Cyt1Aa to mosquito larvae is, on average, one order of magnitude lower than that of Cry4 or Cry11Aa toxins [9]. However, when present in bioassays against mosquitoes, Cyt proteins synergize the mosquitocidal activity of some Cry toxins including Cry4A, Cry4B and Cry11Aa, and the presence of Cyt toxins overcomes the resistance of Culex quinquefasciatus populations resistant to Cry toxins [52]. Recently, we demonstrated that Cyt1Aa synergizes the toxic activity of Cry11Aa by functioning as a membrane bound receptor of Cry11Aa [40]. The proposed mechanism is that Cyt1Aa inserts into midgut epithelium and exposes protein regions that are recognized by Cry11Aa, thereby facilitating the oligomerization of Cry11Aa and its pore-forming activity [40, 41]. Cry11Aa binds Cyt1Aa using the loop α-8of domain II that is also involved in interacting with its alkaline phosphatase receptor [14, 15]. Mutations in the binding regions of Cry11Aa or in Cyt1Aa affected the specific interaction between these proteins and reduced their synergism [40]. Thus, Cyt1Aa synergizes or suppresses resistance to Cry11Aa in mosquitoes by providing a binding site for Cry11Aa that will result in an efficient formation of the Cry11Aa pre-pore that inserts into membranes and induces pore formation.
2.4 Peptide from cadherin receptor CR12-MPED
Cadherin-like receptors are transmembrane proteins with a cytoplasmic domain and an extracellular ectodomain with several cadherin repeats (CADR) (12 in the case of the cadherin receptor of Manduca sexta named Bt-R1) [50]. The ectodomain contains calcium-binding sites, integrin interaction sequences and cadherin binding sequences. The interaction of Cry toxins with cadherin is quite complex involving multiple binding sites. Three different Cry1A binding sites (named CADR7, CADR11 and CADR12) were mapped in the Bt-R1 receptor molecule by the analysis of truncated derivatives in toxin overlay assays [13, 19, 39], by expressing truncated fragments of cadherin on the surface of Drosophila melanogaster Schneider S2 cells [25], or by employing a library of single chain antibodies displayed in M13 phage [21]. Using synthetic peptides representing the exposed loop regions of domain II of Cry1A toxins, loop 2 was identified as the cognate binding region of the M. sexta Bt-R1 receptor CADR7 site, which corresponds to residues 869HITDTNNK876 [20]. In addition, extensive mutagenesis of loop 2 of Cry1A toxins showed that this loop region is important for receptor interaction and toxicity [28]. The loop α8 and loop 2 of Cry1Ab toxin interact with the CADR11 binding site (1331IPLPASILTVTV1342) [19] and the loop 3 of Cry1Ab and Cry1Ac toxins interact with the CADR12 binding site (1422GVLTLNFQ1440) [18, 55]. It was shown that the CADR12 binding site was essential for Cry1Ab cytotoxicity in S2 cells [25] and that Cry1Ab binds CADR12 with high affinity. In addition, the CADR12 binding region induces the formation of the 250 kDa Cry1Ab and Cry1Ac oligomeric structures [46]. All these data together suggest that the CADR12 binding site has an important role as a functional receptor region for Cry1A toxins.
A cadherin fragment of 23.3 kDa named CR12-MPED showed an important synergistic effect with Cry toxins when fed to lepidopteran larvae [8]. The binding of Cry toxins to CR12-MPED fragment is important for the observed enhancement of activity of Cry protein [8]. An important characteristic of the CR12-MPED fragment is that it also binds to the apical membrane of larvae gut cells with high affinity (Kd= 32 nM) suggesting that the synergistic effect observed when fed with Cry toxins was due to an increase in the number of binding sites in the microvilli membrane of the insect. These binding sites would attract Cry1A toxin molecules to the membrane surface, promoting toxin oligomerization and thereby increase the probability that Cry toxin interacts with its secondary receptors, aminopeptidase N and alkaline phosphatase [3, 30].
3. Modifications in the Cry toxin gene
3.1 Site directed mutagenesis
Site-directed mutagenesis has been used extensively to explore the participation of specific protein regions of Cry toxin molecules in their mode of action. Most of these mutations adversely affect toxin action but only a few of them have resulted in generation of Cry toxin molecules with improved toxicity. Regarding the pore forming domain (domain I), Wu and Aronson in 1992 described that a mutation in helix α-5, H168R, which caused a three-fold increase in toxic activity against Manduca sexta larvae [52] (Fig 2). Regarding the other domains of the toxin, there are several mutations located in domains II and III, which showed increased toxicity to some larvae [45]. Mutants N372A or N372G of Cry1Ab, located in loop 2 of domain II, resulted in an 8-fold increase in toxicity against Lymantria dispar, and a triple mutant - N372A, A282G and L283S - showed a 36-fold increase in toxicity [43] (Fig 2). Both mutants showed higher affinity to brush border membrane vesicles (BBMV) isolated from this insect, suggesting that improving binding interaction of the toxin with its receptors could be used as a strategy to design more active toxins [43]. In Cry3A toxin, loop 1 plays an important role in toxin binding. Two mutants in this loop, named A1 and A2, containing multiple mutations each (A1: R345A, Y350F, Y351F; and A2: R345A and the deletion of ΔY350Y351), showed three and eleven fold higher toxicity against the coleopteran Tenebrio molitor, respectively, and showed increased in binding affinity [54] (Fig 3). There also are mutations in loop 3 of Cry3A toxin that also showed a moderate increase in toxicity (2.4 fold) to T. molitor larvae [54] (Fig 3). Recently, a phage display system that allows the directed evolution of Cry toxins was characterized [26]. In this system, random mutagenesis could be incorporated in a specific region of the toxin, such as loop 2, and followed by selection of toxin variants with increased affinity to a specific BBMV preparation or to pure receptor sample, such as cadherin receptor [26]. This high troughput technology could be adapted to improve any toxin, selecting toxins with higher binding affinity against the selected target. Finally, rational modification of the loops residues of Cry1Aa toxin (which is active against lepidopteran larvae) to generate a toxin molecule more similar to Cry4Ba (which is active against mosquitoes) in these regions, resulted in a novel toxin with some activity against the mosquito Culex pipiens [34]. The last two examples highlight the potential of site directed mutagenesis in the toxin-binding sites to develop more potent toxins or toxins with changes in specificity.
Figure 2.
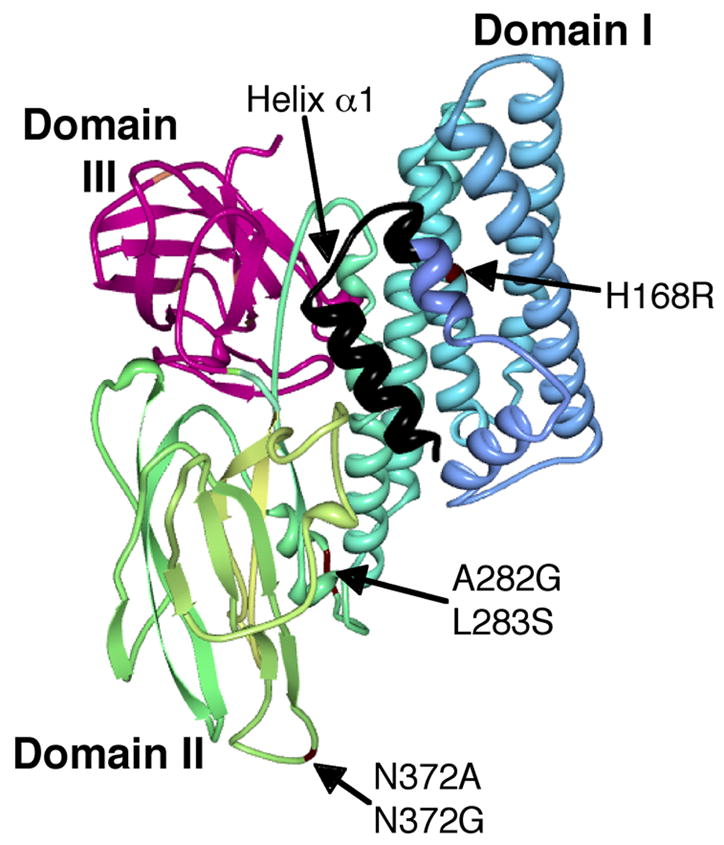
Model of 3D structure of Cry1A toxin, indicating the regions of the toxin that have been modified to improve its insecticidal activity. Point mutations in Domains I and II are labeled in red. Domain III is labeled in dark pink and amino-terminal region deleted in Cry1AMod toxins is labeled in black.
Figure 3.
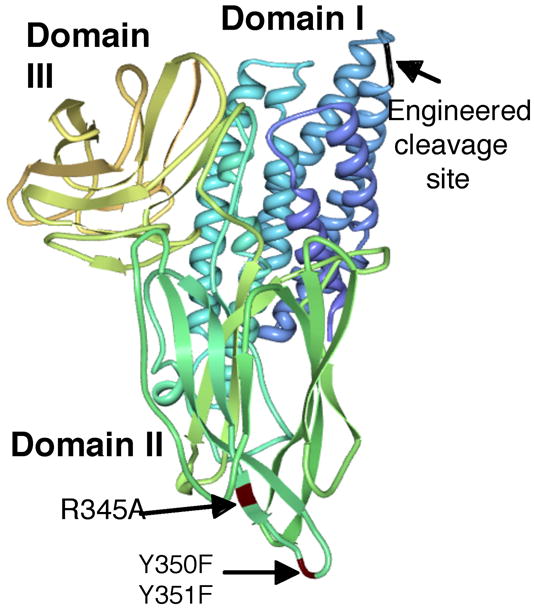
Model of 3D structure of Cry3A toxin. Point mutations in Domain II that improve its insecticidal activity are labeled in red. An engineered proteolytic site included in Domain I is labeled in black.
3.2 Hybrid toxins
In addition to site-directed toxin mutants, the construction of hybrid toxins which include mainly substitutions of domain III have resulted in production of novel toxins with a wider target spectrum or higher toxicity than either of the parental toxins from which were derived (Fig 2). For example several Cry1 toxins with low or no specificity against Spodoptera exigua including Cry1Ab, Cry1Ac, Cry1Ba and Cry1Ea, become active when their domain III is replaced by that of Cry1Ca [11]. The Cry1Ab toxin is not toxic to S. exigua but the hybrid toxin containing domain III of Cry1C showed a 10 times higher toxicity against S. exigua than the parental Cry1C toxin [10]. In another example, replacing residues 450–612 (domain III) of Cry1Aa by those of Cry1Ac resulted in a 300-fold increase in toxicity against Heliothis virescens [6]. The exchanges of domain III alter binding of the toxins to the insect membrane proteins on ligand blots [10]. In the case of Cry1Ac toxin, it is recognized that a cavity in domain III of this toxin interacts with N-acetylgalactosamine (GalNAc) carbohydrate located in the APN receptor [4, 32]. In fact, it has been proposed that Domain III shuffling may have occurred in nature as an evolutionary mechanism to generate toxins with new specificities [1].
3.3 Introduction of specific proteolytic cleavage sites
Activation of Cry toxins is an essential step in their acquisition of toxicity. The activation process in the larval gut has been extensively studied using Cry1 toxins and involves the removal of 27–29 amino acids at the N terminus and 500–600 amino acids at the C terminus. The coleopteran specific Cry3 toxins lack the large C-terminal domain and it has been proposed that midgut activation involves only removal of 57 amino acids from N-terminal region [2, 45]. However, a study of the activation of Cry3A toxin using different proteolytical treatments showed that this protein was cleaved resulting in several polypeptides that remain associated under non-denaturing conditions [7]. Activation with chymotrypsin cleaved the Cry3A toxin at the beginning of helix α-4 producing a 49-kDa major polypeptide and other small fragments, but the individual components were not separated by gel filtration chromatography [7]. The chymotrypsinated protein showed an increase in solubility and retained full activity against Leptinotarsa decemlineata larvae. The Cry3A toxin showed low toxicity against Diabrotica virgifera, an important pest of corn, and it has been documented that introduction of a chymotrypsin/cathepsin G site in the loop between helix α-3 and helix α-4 (see Fig. 3) increased three fold the toxicity of Cry3A against neonate D. virgifera larvae [51]. The authors proposed that enhanced cleavage of this toxin at this proteolytic site permitted the subsequent binding of the activated toxin to the receptors present in the midgut cells.
3.4 Deletion of small regions of the toxin
According to the prevailing model of toxin action, binding of Cry toxins to cadherin receptor is essential for removal of the helix α-1 and this cleavage then promotes oligomerization of the toxin to form a pre-pore structure. Mutations in midgut cadherin that bind Cry1Ac are linked with insect resistance and represent the most common mechanism of resistance [17, 37, 56].
Deletion in the amino terminal region including helix α-1 of Cry1A toxins (named Cry1AMod toxins; see Fig 1 and 2) resulted in Cry toxins that form oligomers in the absence of cadherin receptor. Such modified toxins kill insects that had developed resistance to Cry1A toxins caused by mutations in the cadherin gene. The modified toxins were also effective against insects which had acquired reduced susceptibility to native Bt toxins due to diminished expression of cadherin protein by cadherin gene silencing with RNA interference [46]. Recently we demonstrated that the oligomeric structure formed by the Cry1AMod toxins have pore formation activity, with a pore-forming ability similar to that of the wild type toxin [Muñoz-Garay, personal communication].
In the Cry2A protoxin includes a 49-residue fragment at the amino terminal region that precedes helix α-1 (Fig. 4), which is cleaved upon activation with trypsin [38]. The removal of this fragment by proteolysis serves to expose a formerly occluded region of the toxin that corresponds to the binding region of the Cry2 toxin [38]. It was proposed that this amino terminal cleavage is a rate-limiting step in the toxin binding of Cry2A, and a deletion of 42 residues of the amino-terminal region resulted in a 3 fold enhancement of Cry2A toxicity against Spodoptera littoralis and Agrotis ipsilon [36] (Fig 4). In addition to this deletion, two point mutations (K63F and K64P) increased toxicity by up to six-fold [36]. It is important to mention that the 42 amino acid deletion does not include the residues of helix α-1 of domain I.
Figure 4.
Model of 3D structure of Cry2A toxin, indicating in black the N-terminal region of the toxin that was deleted to improve its insecticidal activity.
Deletions of small fragments at their amino termini resulted either in increased toxicity or in toxins that could be useful for countering insect resistance to native Cry1A toxins either applied as spray products or expressed in transgenic crops. Such altered toxins may be particularly effective in insects that became resistant due to changes in the first toxin receptor.
4. Final Remarks
Bt toxins have been employed successfully in recent years to control the most important insect agricultural pests, but not all insect pests are targets for Cry toxins and it is anticipated that insect resistance to Bt toxins will become more frequent in the coming years and could limit the utility of Bt-crops in the near future.
In this review we have presented different experimental strategies that have been used to improve Cry toxin action. In most cases, the improvement of toxins came as a direct result of understanding their mode of action in the different insect orders. Figure 1 shows a model of the mode of action in different insect orders. The activation process can be modified by addition of protease inhibitors or by introduction of intramolecular cleavage sites in the toxin. The toxin-receptor binding interaction can be increased by improving the contact of the toxin with its receptors, for example by using of chitinases or by including additional binding sites as shown when Cry toxin activity is potentiated by the use of Cyt toxins and by the use of a small fragment of cadherin-like receptor. In addition, the affinity of the binding interaction can be also improved by site-directed mutagenesis or by designing of novel hybrid toxins. Figures 2–4 show structural models of different Cry toxins, indicating the regions of the toxin that have been modified to improve their insecticidal activity. Knowledge of the role of the cadherin receptor in toxin activation and oligomerization formed the basis for the design of modified toxin proteins that have great potential to deal with insect resistance.
Continued analysis of the molecular basis of toxin action including studies of insect specificity, of insect-resistance to Cry toxins and of the role of receptor molecules in toxicity will provide new ways for a rational design of Cry toxins to control insect pests important in agriculture or in human health. Such developments will extend the useful lifespan of this technology.
Acknowledgments
The research work of our groups was supported in part by DGAPA/UNAM IN210208-N, CONACyT U48631-Q and J45863-Q, USDA 2006-35302-17365 and NIH 1R01 AI066014.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Bravo A. Phylogenetic relationships of the Bacillus thuringiensis δ-endotoxin family proteins and their functional domains. J Bacteriol. 1997;179:2793–2801. doi: 10.1128/jb.179.9.2793-2801.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bravo A, Gill SS, Soberón M. Mode of action of Bacillus thuringiensis toxins and their potential for insect control. Toxicon. 2007;49:423–435. doi: 10.1016/j.toxicon.2006.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bravo A, Gómez I, Conde J, Muñoz-Garay C, Sánchez J, Miranda R, Zhuang M, Gill SS, Soberón M. Oligomerization triggers binding of a Bacillus thuringiensis Cry1Ab pore-forming toxin to aminopeptidase N receptor leading to insertion into membrane microdomains. Biochim Biophys Acta. 2004;1667:38–46. doi: 10.1016/j.bbamem.2004.08.013. [DOI] [PubMed] [Google Scholar]
- 4.Burton SL, Ellar DJ, Li J, Derbyshire DJ. N-acetylgalactosamine on the putative insect receptor aminopeptidase N is recognized by a site on the domain III lectin-like fold of a Bacillus thuringiensis insecticidal toxin. J Mol Biol. 1999;287:1011–1022. doi: 10.1006/jmbi.1999.2649. [DOI] [PubMed] [Google Scholar]
- 5.Butko P. Cytolytic toxin Cyt1Aa and its mechanism of membrane damage: data and hypothesis. Appl Environ Microbiol. 2003;69:2415–2422. doi: 10.1128/AEM.69.5.2415-2422.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Caramori T, Albertini AM, Galizzi A. In vivo generation of hybrids between two Bacillus thuringiensis insect-toxin-encoding genes. Gene. 1991;98:37–44. doi: 10.1016/0378-1119(91)90101-g. [DOI] [PubMed] [Google Scholar]
- 7.Carrol J, Convents D, Van Damme J, Boets A, Van Rie J, Ellar DJ. Intramolecular proteolytic cleavage of Bacillus thuringiensis Cry3A δ-endotoxin may facilitate its coleopteran toxicity. J Invertebr Pathol. 1996;70:41–49. doi: 10.1006/jipa.1997.4656. [DOI] [PubMed] [Google Scholar]
- 8.Chen J, Hua G, Jurat-Fuentes JL, Abdullah MA, Adang M. Synergism of Bacillus thuringiensis toxins by a fragment of a toxin-binding cadherin. Proc Nat Acad Sci USA. 2007;104:13901–13906. doi: 10.1073/pnas.0706011104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Crickmore N, Bone EJ, Williams JA, Ellar DJ. Contribution of the individual components of the δ-endotoxin crystal to the mosquitocidal activity of Bacillus thuringiensis subsp israelensis. FEMS Microbiol Lett. 1995;131:249–254. [Google Scholar]
- 10.deMaagd RA, Kwa MSG, van der Klei H, Yamamoto T, Schipper B, Vlak JM, Stiekema WJ, Bosch D. Domain III substitution in Bacillus thuringiensis delta-endotoxin Cry1Ab results in superior toxicity for Spodoptera exigua and altered membrane protein recognition. Appl Environ Microbiol. 1996;62:1537–1543. doi: 10.1128/aem.62.5.1537-1543.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.deMaagd RA, Weemen-Hendriks M, Stiekema W, Bosch D. Bacillus thuringiensis delta-endotoxin Cry1C domain III can function as a specificity determinant for Spodoptera exigua in different but not all Cry1-Cry1C hybrids. Appl Environ Microbiol. 2000;66:1559–1563. doi: 10.1128/aem.66.4.1559-1563.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ding X, Luo Z, Xia L, Gao B, Sun Y, Zhang Y. Improving the insecticidal activity by expression of a recombinant cry1Ac gene with chitinase-encoding gene in acrystalliferous Bacillus thuringiensis. Curr Microbiol. 2008;56:442–447. doi: 10.1007/s00284-008-9112-1. [DOI] [PubMed] [Google Scholar]
- 13.Dorsch JA, Candas M, Griko NB, Maaty WSA, Midbo EG, Vadlamudi RK, Bulla LA., Jr Cry1A toxins of Bacillus thuringiensis bind specifically to a region adjacent to the membrane-proximal extracellular domain of Bt-R1 in Manduca sexta: involvement of a cadherin in the entomopathogenicity of Bacillus thuringiensis. Insect Biochem Mol Biol. 2002;32:1025–1036. doi: 10.1016/s0965-1748(02)00040-1. [DOI] [PubMed] [Google Scholar]
- 14.Fernandez LE, Aimanova KG, Gill SS, Bravo A, Soberón M. A GPI-anchored alkaline phosphatase is a functional midgut receptor of Cry11Aa toxin in Aedes aegypti larvae. Biochem J. 2006;394:77–84. doi: 10.1042/BJ20051517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fernández LE, Pérez C, Segovia L, Rodríguez MH, Gill SS, Bravo A, Soberón M. Cry11Aa toxin from Bacillus thuringiensis binds its receptor in Aedes aegypti mosquito larvae trough loop α-8 of domain II. FEBS Lett. 2005;79:3508–3514. doi: 10.1016/j.febslet.2005.05.032. [DOI] [PubMed] [Google Scholar]
- 16.Ferré J, Van Rie J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Ann Rev Entomol. 2002;47:501–533. doi: 10.1146/annurev.ento.47.091201.145234. [DOI] [PubMed] [Google Scholar]
- 17.Gahan LJ, Gould F, Heckel DG. Identification of a gene associated with Bt resistance in Heliothis virescens. Science. 2001;293:857–860. doi: 10.1126/science.1060949. [DOI] [PubMed] [Google Scholar]
- 18.Gómez I, Arenas I, Benitez I, Miranda-Ríos J, Becerril B, Grande R, Almagro JC, Bravo A, Soberón M. Specific epitopes of Domains II and III of Bacillus thuringiensis Cry1Ab toxin involved in the sequential interaction with cadherin and aminopeptidase-N receptors in Manduca sexta. J Biol Chem. 2006;281:34032–34039. doi: 10.1074/jbc.M604721200. [DOI] [PubMed] [Google Scholar]
- 19.Gomez I, Dean DH, Bravo A, Soberón M. Molecular basis for Bacillus thuringiensis Cry1Ab toxin specificity: Two structural determinants in the Manduca sexta Bt-R1 receptor interact with loops α-8 and 2 in domain II of Cry1Ab toxin. Biochemistry. 2003;42:10482–10489. doi: 10.1021/bi034440p. [DOI] [PubMed] [Google Scholar]
- 20.Gomez I, Miranda-Rios J, Rudiño-Piñera E, Oltean DI, Gill SS, Bravo A, Soberón M. Hydropathic complementarity determines interaction of epitope 869HITDTNNK876 in Manduca sexta Bt-R1 receptor with loop 2 of domain II of Bacillus thuringiensis Cry1A toxins. J Biol Chem. 2002;277:30137–30143. doi: 10.1074/jbc.M203121200. [DOI] [PubMed] [Google Scholar]
- 21.Gomez I, Oltean DI, Sanchez J, Bravo A, Gill S, Soberón M. Mapping the epitope in Cadherin-like receptors involved in Bacillus thuringiensis Cry1A toxin interaction using phage display. J Biol Chem. 2001;276:28906–28912. doi: 10.1074/jbc.M103007200. [DOI] [PubMed] [Google Scholar]
- 22.Gómez I, Sánchez J, Miranda R, Bravo A, Soberón M. Cadherin-like receptor binding facilitates proteolytic cleavage of helix α-1 in domain I and oligomer pre-pore formation of Bacillus thuringiensis Cry1Ab toxin. FEBS Lett. 2002;513:242–246. doi: 10.1016/s0014-5793(02)02321-9. [DOI] [PubMed] [Google Scholar]
- 23.Gould F. Sustainability of transgenic insecticidal cultivars: integrating pest genetics and ecology. Ann Rev Entomol. 1998;43:701–726. doi: 10.1146/annurev.ento.43.1.701. [DOI] [PubMed] [Google Scholar]
- 24.Hilder VA, Gatehouse AMR, Sheerman SE, Barrer RF, Boulter D. A novel mechanism of insect resistance engineered into tobacco. Nature. 1987;330:160–163. [Google Scholar]
- 25.Hua G, Jurat-Fuentes JL, Adang MJ. Bt-R1 extracellular cadherin repeat 12 mediates Bacillus thuringiensis Cry1Ab binding and cytotoxicity. J Biol Chem. 2004 Jul;279:28051–28056. doi: 10.1074/jbc.M400237200. [DOI] [PubMed] [Google Scholar]
- 26.Ishikawa H, Hoshino Y, Motoki Y, Kawahara T, Kitajima M, Kitami M, Watanabe A, Bravo A, Soberón M, Honda A, Yaoi K, Sato R. A system for the directed evolution of the insecticidal protein from Bacillus thuringiensis. Mol Biotechnol. 2007;36:90–102. doi: 10.1007/s12033-007-0001-9. [DOI] [PubMed] [Google Scholar]
- 27.Janmaat AF, Myers JH. Rapid evolution and the cost of resistance to Bacillus thuringiensis in greenhouse populations of cabbage loppers, Trichoplusia ni. Proc Roy Soc Lond B. 2003;270:2263–2270. doi: 10.1098/rspb.2003.2497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jenkins JL, Dean DH. Exploring the mechanism of action of insecticidal proteins by genetic engineering methods. In: Setlow JK, editor. Genetic Engineering: Principles and Methods. New York: Plenum Press; 2000. pp. 33–54. [DOI] [PubMed] [Google Scholar]
- 29.Jiménez-Juárez N, Muñoz-Garay C, Gómez I, Saab-Rincon G, Damian-Almazo JY, Gill SS, Soberón M, Bravo A. Bacillus thuringiensis Cry1Ab mutants affecting oligomer formation are non-toxic to Maduca sexta larvae. J Biol Chem. 2007;282:21222–21229. doi: 10.1074/jbc.M701314200. [DOI] [PubMed] [Google Scholar]
- 30.Jurat-Fuentes JL, Adang MJ. Characterization of a Cry1Ac-receptor alkaline phosphatase in susceptible and resistant Heliothis virescens larvae. Eur J Biochem. 2004;271:3127–3135. doi: 10.1111/j.1432-1033.2004.04238.x. [DOI] [PubMed] [Google Scholar]
- 31.Knowles BH, Blatt MR, Tester M, Horsnell JM, Carroll J, Menestrina G, Ellar DJ. A cytolytic delta-endotoxin from Bacillus thuringiensis var. israelensis forms cation-selective channels in planar lipid bilayers. FEBS Lett. 1989;244:259–262. doi: 10.1016/0014-5793(89)80540-x. [DOI] [PubMed] [Google Scholar]
- 32.Lee MK, You TH, Gould FL, Dean DH. Identification of residues in domain III of Bacillus thuringiensis Cry1Ac toxin that affect binding and toxicity. Appl Environ Microbiol. 1999;65:4513–4520. doi: 10.1128/aem.65.10.4513-4520.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li J, Koni PA, Ellar DJ. Structure of the mosquitocidal delta-endotoxin CytB from Bacillus thuringiensis ssp. kyushuensis and implications for membrane pore formation. J Mol Biol. 1996;257:129–152. doi: 10.1006/jmbi.1996.0152. [DOI] [PubMed] [Google Scholar]
- 34.Liu XS, Dean DH. Redesigning Bacillus thuringiensis Cry1Aa toxin into a mosquito toxin. Prot Eng Design Selec. 2006;19:107–111. doi: 10.1093/protein/gzj009. [DOI] [PubMed] [Google Scholar]
- 35.MacIntosh SC, Kishore GM, Perlak FJ, Marrone PG, Stone TB, Sims SR, Fuchs RL. Potentiation of Bacillus thuringiensis insecticidal activity by serine protease inhibitors. J Agric Food Chem. 1990;38:1145–1152. [Google Scholar]
- 36.Mandal CC, Gayen S, Basu A, Ghosh KS, Dasgupta S, Maiti MK, Sen SK. Prediction based protein engineering of domain I of Cry2A entomocidal toxin of Bacillus thuringiensis for the enhancement of toxicity against lepidopteran insects. Prot Eng Design Selec. 2007;20:599–606. doi: 10.1093/protein/gzm058. [DOI] [PubMed] [Google Scholar]
- 37.Morin S, Biggs RW, Shriver L, Ellers-Kirk C, Higginson D, Holley D, Gahan L, Heckel DG, Carriere Y, Dennehy TJ, Brown JK, Tabashnik BE. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc Nat Acad Sci USA. 2003;100:5004–5009. doi: 10.1073/pnas.0831036100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Morse RJ, Yamamoto T, Strud RM. Structure of cry2A suggests an unexpected receptor binding epitope. Structure. 2001;9:409–417. doi: 10.1016/s0969-2126(01)00601-3. [DOI] [PubMed] [Google Scholar]
- 39.Nagamatsu Y, Koike T, Sasaki K, Yoshimoto A, Furukawa Y. The cadherin-like protein is essential to specificity determination and cytotoxic action of the Bacillus thuringiensis insecticidal. FEBS Lett. 1999;460:385–390. doi: 10.1016/s0014-5793(99)01327-7. [DOI] [PubMed] [Google Scholar]
- 40.Pérez C, Fernandez LE, Sun J, Folch JL, Gill SS, Soberón M, Bravo A. Bti Cry11Aa and Cyt1Aa toxins interactions support the synergism-model that Cyt1Aa functions as membrane-bound receptor. Proc Nat Acad Sci USA. 2005;102:18303–18308. doi: 10.1073/pnas.0505494102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pérez C, Muñoz-Garay C, Portugal LC, Sánchez J, Gill SS, Soberón M, Bravo A. Bacillus thuringiensis subsp. israelensis Cyt1Aa enhances activity of Cry11Aa toxin by facilitating the formation of a pre-pore oligomeric structure. Cellular Microbiol. 2007;9:2931–2937. doi: 10.1111/j.1462-5822.2007.01007.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Promdonkoy B, Ellar DJ. Membrane pore architecture of a cytolytic toxin from Bacillus thuringiensis. Biochem J. 2000;350:275–282. [PMC free article] [PubMed] [Google Scholar]
- 43.Rajamohan F, Alzate O, Cotrill JA, Curtiss A, Dean DH. Protein engineering of Bacillus thuringiensis delta-endotoxin: mutations at domain II of CryIAb enhance receptor affinity and toxicity toward gypsy moth larvae. Proc Natl Acad Sci USA. 1996;93:14338–14343. doi: 10.1073/pnas.93.25.14338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Regev A, Keller M, Strizhov N. Synergistic activity of a Bacillus thuringiensis delta-endotoxin and a bacterial endochitinase against Spodoptera littoralis larvae. Appl Environ Microbiol. 1996;62:3581–3586. doi: 10.1128/aem.62.10.3581-3586.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Schnepf E, Crickmore N, Van Rie J, Lereclus D, Baum JR, Feitelson J, Zeigler D, Dean DH. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol Mol Biol Rev. 1998;62:705–806. doi: 10.1128/mmbr.62.3.775-806.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Soberón M, Pardo-López L, López I, Gómez I, Tabashnik B, Bravo A. Engineering Modified Bt Toxins to Counter Insect Resistance. Science. 2007;318:1640–1642. doi: 10.1126/science.1146453. [DOI] [PubMed] [Google Scholar]
- 47.Tabashnik BE. Evolution of resistance to Bacillus thuringiensis. Annu Rev Entomol. 1994;39:47–79. [Google Scholar]
- 48.Tabashnik BE, Dennehy TJ, Carriere Y. Delayed resistance to transgenic cotton in pink bollworm. Proc Nat Acad Sci. 2005;102:15389–15393. doi: 10.1073/pnas.0507857102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tabashnik BE, Gassmann AJ, Crowder DW, Carriere Y. Insect resistance to Bt crops: evidence versus theory. Nature Biotecchnol. 2008;26:199– 202. doi: 10.1038/nbt1382. [DOI] [PubMed] [Google Scholar]
- 50.Vadlamudi RK, Weber E, Ji I, Ji TH, Bulla LA., Jr Cloning and expression of a receptor for an insecticidal toxin of Bacillus thuringiensis. J Biol Chem. 1995;270:5490–5494. doi: 10.1074/jbc.270.10.5490. [DOI] [PubMed] [Google Scholar]
- 51.Walters FS, Stacy ChM, Lee MK, Palekar N, Chen JS. An engineered chymotrypsin/cathepsin G site in domain I renders Bacillus thuringiensis Cry3A active against western corn rootworm larvae. Appl Environ Microbiol. 2008;74:367–374. doi: 10.1128/AEM.02165-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wirth MC, Georghiou GP, Federici BA. CytA enables CryIV endotoxins of Bacillus thuringiensis to overcome high levels of CryIV resistance in the mosquito Culex quinquefasciatus. Proc Natl Acad Sci USA. 1997;94:10536–10540. doi: 10.1073/pnas.94.20.10536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wu D, Aronson AI. Localized mutagenesis defines regions of the Bacillus thuringiensis delta-endotoxin involved in toxicity and specificity. J Biol Chem. 1992;267:2311–2327. [PubMed] [Google Scholar]
- 54.Wu SJ, Koller CN, Miller DL, Bauer LS, Dean DH. Enhanced toxicity of Bacillus thuringiensis Cry3A delta-endotoxin in coleopterans by mutagenesis in a receptor binding loop. FEBS Lett. 2000;473:227–232. doi: 10.1016/s0014-5793(00)01505-2. [DOI] [PubMed] [Google Scholar]
- 55.Xie R, Zhuang M, Ross LS, Gómez I, Oltean DI, Bravo A, Soberón M, Gill SS. Single amino acid mutations in the cadherin receptor from Heliothis virescens affect its toxin binding ability to Cry1A toxins. J Biol Chem. 2005;280:8416–8425. doi: 10.1074/jbc.M408403200. [DOI] [PubMed] [Google Scholar]
- 56.Xu X, Yu L, Wu Y. Disruption of a cadherin gene associated with resistance to Cry1Ac δ-endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl Environ Microbiol. 2005;71:948–954. doi: 10.1128/AEM.71.2.948-954.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]