

Abstract
Previous studies have demonstrated anisotropic patterns of hand impedance under static conditions and during movement. Here we show that the pattern of kinematic error observed in studies of dynamics learning is associated with this anisotropic impedance pattern. We also show that the magnitude of kinematic error associated with this anisotropy dictates the amount of motor learning and, consequently, the extent to which dynamics learning generalizes. Subjects were trained to reach to visual targets while holding a robotic device that applied forces during movement. On infrequent trials, the load was removed and the resulting kinematic error was measured. We found a strong correlation between the pattern of kinematic error and the anisotropic pattern of hand stiffness. In a second experiment subjects were trained under force-field conditions to move in two directions: one in which the dynamic perturbation was in the direction of maximum arm impedance and the associated kinematic error was low and another in which the perturbation was in the direction of low impedance where kinematic error was high. Generalization of learning was assessed in a reference direction that lay intermediate to the two training directions. We found that transfer of learning was greater when training occurred in the direction associated with the larger kinematic error. This suggests that the anisotropic patterns of impedance and kinematic error determine the magnitude of dynamics learning and the extent to which it generalizes.
INTRODUCTION
When the limb is displaced, it generates resistive forces that depend on musculoskeletal geometry (Mussa-Ivaldi et al. 1985), reflexes (Bennett 1994), muscle cocontraction (Darainy et al. 2004; Gomi and Osu 1998), and the reciprocal activity of antagonist muscle pairs associated with movement (Darainy et al. 2007; Gomi and Osu 1998; Perreault et al. 2002). This resistance to displacement is termed the mechanical impedance of the limb. Research has shown that impedance can be purposefully modulated to counteract environmental instabilities (Burdet et al. 2001; Darainy et al. 2004; Franklin et al. 2003) and that it limits error early in the learning process (Darainy and Ostry 2008; Milner and Cloutier 1993; Thoroughman and Shadmehr 1999). However, little is known about how the geometric pattern of limb impedance affects motor learning and its generalization. Arm impedance, and specifically arm stiffness, is anisotropic under static conditions (Darainy et al. 2004; Gomi and Osu 1998; Mussa-Ivaldi et al. 1985; Perreault et al. 2002; Tsuji et al. 1995) and during movement (Burdet et al. 2001; Darainy et al. 2007; Frolov et al. 2006; Gomi and Kawato 1997). This means that displacements in some directions are resisted more than displacements in others. Here our goal was to evaluate whether directional asymmetries in impedance affect dynamics learning and generalization.
Since the work of Shadmehr and Mussa-Ivaldi (1994), multijoint reaching movements under conditions in which dynamic forces are applied to the hand have been used to understand how the nervous system controls movement. In these studies the magnitude of kinematic error varies as subjects move in different directions while experiencing the same perturbation (Malfait et al. 2002; Mattar and Ostry 2007; Shadmehr and Holcolmb 1997; Shadmehr and Mussa-Ivaldi 1994). These directional differences in kinematic error are evident on initial movements following unexpected changes in dynamics. Computational simulations of human arm movement suggest that the directional differences in kinematic error may arise from directional asymmetries in arm impedance (Shadmehr and Brashers-Krug 1997; Shadmehr and Mussa-Ivaldi 1994). However, to our knowledge this has not been empirically demonstrated.
The goals of the current study were twofold. In experiment 1 we sought to document the relationship between directional differences in kinematic error and the anisotropic pattern of arm impedance in human subjects. We measured hand stiffness, which is a good model of arm impedance as a whole for the current experimental conditions (see discussion). We also measured kinematic error as subjects made reaching movements to targets. We observed a strong correlation such that dynamic perturbations in directions of high stiffness were associated with low kinematic error (and vice versa). In experiment 2 we investigated whether directional differences in stiffness and kinematic error lead to directional differences in the amount of learning and in the extent to which learning generalizes. Generalization is characterized as the extent to which motor learning affects performance in a novel task. Generalization has been reported in the context of dynamics learning (Malfait et al. 2002; Mattar and Ostry 2007; Shadmehr and Mussa-Ivaldi 1994; Thoroughman and Shadmehr 2000) and following visuomotor transformations (Caithness et al. 2004; Ghahramani and Wolpert 1997; Krakauer et al. 2000). In the present study, subjects learned to compensate for a dynamic perturbation when the limb was deflected in a direction associated with either low stiffness and large kinematic error or high stiffness and small kinematic error. We found that transfer of learning to a test direction was larger following training in directions associated with large kinematic error and low arm impedance. Our results indicate that the magnitude of learning varies with stiffness and the associated kinematic error and that generalization is greater following adaptation to large errors.
METHODS
Subjects
Twenty-eight right-handed subjects, between the ages of 18 and 31 yr, participated in the study. Eight subjects participated in experiment 1 (hand impedance and kinematic error) and 20 subjects participated in experiment 2 (hand impedance and generalization of learning). Subjects had no history of sensory or motor disorders and were naïve regarding the experimental procedure. The McGill University Research Ethics Board approved all experimental procedures.
Experimental setup
Subjects were seated in front of a two-degree-of-freedom planar robotic arm (InMotion2, Interactive Motion Technologies) that they grasped with their right hand (Fig. 1A). For each subject, the position of the seat relative to the robot was adjusted to situate the subject in a standard position with a shoulder angle of 45° (relative to the frontal plane) and elbow angle of 90° (relative to the upper arm). This posture corresponds to a position in the middle of the workspace. We also adjusted the seat height to have 80° of shoulder abduction for all subjects. The subject's forearm was supported against gravity by an air sled and a harness restrained the subject's shoulder and upper body. Eight light-emitting diodes (LEDs) were embedded in a glass-top table in front of the subject and served as visual targets. One LED at the center defined the middle of the workspace. The target LEDs were placed about a circle at a distance of 15 cm from the center LED and were equally spaced at 45° increments (Fig. 1A). The LEDs each illuminated a target zone of 1.5-cm radius. A computer monitor was placed next to the robot and in front of the subject. This monitor was used to give the subject visual feedback about movement duration (too fast, too slow, correct speed; indicated respectively by red, green, and blue signals). Hand position was measured using 16-digit optical encoders (Gurley Precision Instruments) located in the robot arm. Forces applied to the robot handle by the subject were measured using a force–torque sensor (ATI Industrial Automation) that was mounted above the manipulandum handle.
FIG. 1.

Experimental setup and arrangement of visual targets. A: the center of the workspace is defined for each subject by a shoulder angle of 45° relative to the frontal plane and an elbow angle of 90° relative to the upper arm. In experiment 1, subjects made movements to each of 8 visual targets. B: 2 groups of naïve subjects were recruited for experiment 2. The top panel shows the training directions (45 and 135°) and the reference direction in which we tested transfer of dynamics learning (90°) for the 1st group of subjects. The bottom panel shows the training (135 and 225°) and reference (180°) directions for the 2nd group of subjects. C: the experimental procedure for experiment 2. The top panel shows the order of target presentation for the 1st group of subjects. Subjects were divided into 2 subgroups of 5 subjects each and completed 9 blocks of reaching movements. The bottom panel shows the order of target presentation for the 2nd group of subjects.
Experiment 1
Experiment 1 was divided into two parts. In the first phase, each subject's hand stiffness was estimated under static conditions in the middle of the workspace. In the second phase, subjects made movements to visual targets while the robot applied a dynamic perturbation to the hand. The experiment was carried out in two sessions over 2 days.
Estimation of stiffness under null-field conditions
Subjects grasped the robot handle with a comfortable grip and held it in the middle of the central target. Subjects were asked to relax as much as possible and not to resist the actions of the robot. No specific instructions were provided with regard to cocontraction. We used a standard procedure that involved position-servo displacement to estimate hand stiffness (Darainy et al. 2007). The robot was used to displace the subject's hand from its resting position. Amplitude of the displacement was 6 mm and its duration was 400 ms. The 400- ms displacement was divided into 100-ms ramp-on, 200-ms hold, and 100-ms ramp-off phases. The displacement was ramped on and off following a minimum jerk trajectory to move the subject's hand smoothly. Robot stiffness and viscosity were set respectively to 4,000 N/m and 100 Ns/m during the perturbation. The effective mass of the robot at the endpoint was about 400 g. No forces or any other manipulations were applied outside of the servo-control interval. Servo-displacements were initiated only if the subject's hand was within 5 mm of the center of the central LED and had a tangential velocity <1 mm/s for a period lasting between 500 and 1,000 ms (selected randomly for each trial). A set of 32 position-servo displacements in eight directions about a circle (4 displacements in each direction) was used for stiffness estimation.
Movements to visual targets
The second phase of experiment 1 involved reaching movements toward visual targets that were placed in front of the subjects. Subjects were trained to make 15-cm reaching movements to each of eight visual targets (Fig. 1A). The order of target presentation was different for each subject. The sequence progressed through the eight targets in a clockwise order, beginning with the 0° target for Subject 1, the 45° target for Subject 2, and so forth. At the start of each trial, subjects held their hand inside the central start location for 1,200 ± 300 ms. The target was then illuminated and subjects were asked to move toward the target in one smooth motion. Subjects were asked to move to the target in as straight a line as possible. At the end of each movement, subjects received feedback based on movement duration. The feedback was used only to encourage subjects to move with the desired duration, although trials were not dropped from analysis for failing to satisfy all the criteria. The desired movement duration, measured from the time the hand left the central position to the time the hand entered the target location, was 600 ± 50 ms. Based on the same criteria, the average movement duration for all subjects was 561 ± 49 ms (mean ± SD). At the conclusion of every trial, the robot moved the subject's hand back to the center.
Each subject completed 200 reaching movements toward each of the eight targets. The first 40 movements were made under null-field conditions. On 10% of null-field trials (selected randomly) a clockwise velocity dependent curl field (see following text) was unexpectedly applied to the hand. We will refer to these movements as unexpected force-field trials. Subjects then made 120 movements in the training phase. On 90% of training movements, the robot applied the clockwise curl field. On the remaining 10% (selected randomly), the robot motors were unexpectedly turned off and subjects made movements under null-field conditions (catch trials). The final 40 movements in each direction were carried out under null-field conditions to wash out the effects of training.
The clockwise force field was applied according to
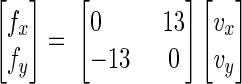 |
(1) |
In this equation, x and y are the lateral and sagittal directions (Fig. 1A), fx and fy are the commanded force to the robot (in newtons), and vx and vy are hand velocities in Cartesian coordinates (in meters/second). The damping matrix coefficients are in newton seconds/meter.
Experiment 2
Subjects were divided into two groups that were trained to make reaching movements to different sets of targets (shown in the top and bottom panels of Fig. 1B). The experimental procedure for both groups is summarized in Fig. 1C. The first group of 10 subjects made movements to the 45, 90, and 135° targets. In all, subjects completed nine blocks of movements (Fig. 1C, top). The first three blocks of the experiment involved 50 reaching movements under null-field conditions to each of the three targets. For half of the subjects (5), the order of target presentation was 45, 90, and 135°, whereas the other 5 started with the 135° target, followed by 90° and finally the 45° target. The fourth, sixth, and eighth blocks of the experiment each involved 120 reaching movements under force-field conditions. The order for the first group was 45, 135, and 90°; the order for the second group was 135, 45, and 90°. On 10% of trials (selected randomly) the robot motors were turned off and subjects were tested under null-field conditions (catch trials). For both groups of subjects, movements in the fifth, seventh, and ninth blocks of the experiment were made toward the 90° target under null-field conditions (50 reaching movements per block). In this way we were able to assess the transfer of learning from the targets selected in the fourth, sixth, and eighth blocks to the 90° target selected in the fifth, seventh, and ninth blocks. These movements also served to wash out the effects of training and thus minimize carryover between experimental conditions.
For the 10 subjects tested with the second set of targets (Fig. 1B, bottom), the method of testing was the same as described earlier. For these subjects the order of testing is shown in the bottom panel of Fig. 1C. For the force-field blocks, half of the 10 subjects moved first to the 135°, followed by the 225° and finally the 180° target. The other 5 subjects moved first to the 225°, followed by the 135° and then the 180° targets.
Data analysis
For both experiments 1 and 2, hand position and forces applied to the handle were sampled at 400 Hz. The resulting signals were low-pass filtered at 40 Hz using a second-order, zero-phase lag Butterworth filter. Position signals were numerically differentiated to provide velocity values. Movement start was scored on a per trial basis at 5% of peak hand velocity. Movement end was defined as the time at which tangential velocity dropped to <5% of its maximum and stayed there for ≥100 ms. The resulting signals were time normalized to have the same number of samples per trial. The perpendicular deviation (PD) of the hand from a straight line connecting movement start and end was calculated to score movement straightness.
In experiment 1 we plotted 50% confidence interval ellipses to visualize directional differences in kinematic error. To calculate these ellipses, we first vectorized the PD data for each subject separately. That is, we plotted PD values in two dimensions, reflecting both the magnitude of PD and the direction in which movements were made. For each subject a 2 × 8 matrix of PD values (x and y components in 8 directions) was created. Across all subjects, we calculated the covariance matrix of these vectorized PDs. This covariance matrix was then used to calculate a 50% confidence interval ellipse (i.e., 50% of the PD values lie within the ellipse).
To estimate static hand stiffness in experiment 1, we measured changes in the hand's position and the forces applied to the handle during position-servo displacement. Differences in both position (dx and dy, in meters) and force (dfx and dfy, in newtons) were averaged over the 50 ms immediately prior to displacement and the last 50 ms of the hold time (i.e., between −50 and 0 ms, and from 250 to 300 ms relative to the onset of the perturbation). Trials were dropped from analysis if the SD of dfx or dfy (for that specific displacement) exceeded a threshold level of 0.15 N. This resulted in the rejection of <1.5% of trials. The following equation shows the linearized relationship between imposed hand displacement and the resulting restoring force
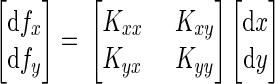 |
(2) |
K is a matrix representing the stiffness of the hand in N/m. Linear regression was used to estimate this matrix.
Statistical analysis
In experiment 1, we used repeated-measures ANOVAs to assess how kinematic error differed depending on movement direction. We also used correlation analyses to describe the relationship between hand stiffness and kinematic error. In experiment 2, we used mixed-factor repeated-measures ANOVAs to assess whether the extent of generalization depended on the direction in which subjects were previously trained. For both experiments, ANOVA was followed by Bonferroni-corrected post hoc comparisons where appropriate. Unless stated otherwise, all statistical interactions were nonreliable (P > 0.05).
RESULTS
Hand impedance and kinematic error
In experiment 1 we assessed the relationship between hand stiffness measured under static conditions and the perpendicular deviation of the hand from a straight line when it encounters a sudden change in environmental dynamics. Figure 2A shows the last of the trials from the initial null-field phase on which the forces were unexpectedly activated. Figure 2B shows the last of the trials from the training phase on which the forces were unexpectedly removed. For both figures, the hand paths represent the mean time-normalized trajectory (±SE) in each of the eight directions, computed across subjects. There are clear directional differences in the extent to which changes in environmental dynamics due to both the introduction and the removal of load result in perpendicular deviation. Whereas movements in the 45 and 225° directions have the lowest PD, movements in the 135 and 315° directions show the largest deviation from straight-line trajectories. PD for movements in the remaining directions (0, 90, 180, 270°) lies intermediate to the extreme PD values. Figure 2C quantifies the data presented in Fig. 2, A and B, showing mean PD at maximum tangential hand velocity (±SE) for the final unexpected force-field trial during null-field trials (dark blue dots) and the last catch trial during training trials (red dots) for all eight directions of movement. Keep in the mind that the data plotted in each of the eight movement directions give the magnitude of kinematic error resulting from loads that act perpendicular to the movement direction. Because the kinematic error on final catch trials is in an opposite direction to error on the last unexpected force-field trials, the sign of the PD for the last catch trial is flipped in Fig. 2C. That is, PD for the final unexpected force-field trial and the last catch trial are shown in Fig. 2C as having the same sign. The figure also shows ellipses plotted in dashed lines that fit the median of the data.
FIG. 2.

The pattern of kinematic error is associated with the anisotropic pattern of hand stiffness. A: mean hand trajectories for the final unexpected force-field movements in the 8 training directions. The shaded areas represent ±1SE. It can be seen that kinematic error following a sudden change in the environmental dynamics was greatest for movements in the 135 and the 315° directions and was the least in the 45 and 225° directions. B: mean hand trajectories (±1SE) for the final catch trials. Again, kinematic error is greatest in the 135 and 315° directions. C: mean (±1SE) perpendicular deviation (PD) on the final unexpected force-field trial (dark blue) and final catch trial (red), measured at the point of maximum tangential hand velocity. The dashed lines show a 50% confidence interval ellipse of PD in the 8 training directions. This ellipse is elongated along the 135–315° axis. D: the mean (±1SE) hand stiffness ellipse for all subjects, measured in the center of the workspace under static conditions. The major axis of the ellipse is oriented at 135.1°. E: for each subject, the magnitude of PD on the final unexpected force-field trial was plotted against the magnitude of the hand's restoring force in the direction perpendicular to the direction of movement (i.e., in the direction of kinematic error caused by the change in environmental dynamics). There is a negative correlation, such that large kinematic error is associated with weaker resistance to perturbations directed perpendicular to the direction of movement. The red line represents a linear fit to the data points. F: magnitude of PD on the final catch trials was plotted against the magnitude of the hand's restoring force in the direction perpendicular to the direction of movement. The red line depicts a linear fit to the data. As in E, there is a negative correlation between kinematic error and hand stiffness.
Figure 2D shows the mean (±SE) static stiffness ellipse measured in the center of the workspace and averaged over all eight subjects. The mean (±SE) static stiffness matrix (in N/m) is
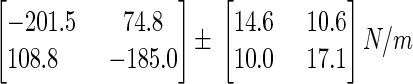 |
The ellipse shape, defined as the ratio of major to minor axis, is 2.81. The ellipse size, defined as the area enclosed by the stiffness ellipse, is 9.1 N2/cm2. The major axis of the average stiffness ellipse is oriented at 135.1°. This direction of stiffness ellipse elongation is similar to the direction of elongation for the 50% confidence ellipses for PD shown in Fig. 2C. The orientation of these ellipses was calculated to be 127.5° for the final unexpected force-field trials and 131.6° for the final catch trials. The eigenvalues of the static stiffness matrix were −283.8 and −102.7 N/m.
We hypothesized that the extent to which an unexpected change in environmental dynamics results in kinematic error would be associated with patterns of hand stiffness. This can be seen by examining the data in Fig. 2, C and D. The results shown in Fig. 2D indicate that the direction of maximum stiffness lies along an axis oriented at 135.1° relative to the frontal plane. The results shown in Fig. 2C also indicate that maximum kinematic error occurred when subjects moved along the same axis (to both the 135 and the 315° targets). These directional differences in kinematic error were found to be reliable (see following text). Recall that the dynamic perturbation acted perpendicular to the movement direction. Thus when subjects moved to targets at 135 or 315°, the force field deflected the limb perpendicular to the movement path in directions of low stiffness. This accounts for the large limb deflection for movements to the 135 and 315° targets.
ANOVAs on the data presented in Fig. 2C reveal that the magnitude of kinematic error varies with the direction of movement, both for the final unexpected force-field trials and for the last catch trials. ANOVA on the final unexpected force-field trials indicated that these directional differences in PD were reliable [F(7,49) = 14.318, P < 0.001]. Post hoc comparisons showed that the largest perturbations occurred when loads acted in directions of lowest stiffness, that is, when subjects moved to targets at 135 and 315°. In particular, PD for movements in the 45° direction was reliably smaller than PD for movements in the 135° (P < 0.05) and 315° (P < 0.05) directions. In addition, PD for movements in the 225° direction was reliably smaller than PD for movements in the 135° (P < 0.01) and 315° (P < 0.01) directions. Another ANOVA on the final catch trials yielded similar results. There were reliable differences in the magnitude of PD that depended on the direction of movement [F(7,49) = 13.539, P < 0.001]. Post hoc comparisons showed that PD for the 45° direction was reliably smaller than PD for the 135° (P < 0.01) and 315° (P < 0.05) directions. Additionally, PD for the 225° direction was reliably smaller than PD for the 135° (P < 0.01) and 315° (P < 0.05) directions.
To demonstrate that the effect of stiffness on kinematic error is a graded one, we performed a second analysis involving repeated-measures ANOVAs. We grouped PDs by directions of movement associated with 1) maximum arm stiffness (135 and 315°), 2) minimum arm stiffness (45 and 225°), and 3) intermediate stiffness (0, 90, 180, and 270°). These analyses revealed that PD on final unexpected force-field trials [F(2,14) = 74.293, P < 0.001] and on final catch trials [F(2,14) = 42.073, P < 0.001] depended on the directions in which movements were made. For both analyses, post hoc comparisons revealed that in each of the three directions (high, low, and intermediate stiffness), PD was reliably different from the remaining two directions (i.e., for all comparisons, P < 0.01).
To further investigate the relationship between kinematic error and stiffness, we performed the following analysis. The goal was to assess whether kinematic deviations arising from the force field can be accounted for by directional differences in arm stiffness. We used the stiffness matrices calculated for each subject to determine the resistive forces that would have been generated by the arm following a 1-cm displacement perpendicular to the eight directions of movement (i.e., in the directions in which forces were applied by the velocity-dependent force field). Note that since the relationship between displacement and force was assumed to be linear, a displacement of any magnitude could have been chosen. We then evaluated the relationship between kinematic error perpendicular to each direction of movement and the computed resistive force due to the stiffness of the arm in the same perpendicular direction. We found that for both the introduction and removal of the force field, kinematic error is negatively correlated with the magnitude of the restoring force in the direction of the limb deflection. That is, the larger the restoring force in the direction of the dynamic perturbation, the smaller the kinematic error during movement—a relationship depicted in Fig. 2, E and F. The ordinate shows the hand's restoring force perpendicular to the direction of movement (i.e., in the direction that the hand was deviated by the force field) and the abscissa shows PD on the final unexpected force-field trial (Fig. 2E) or the final catch trial (Fig. 2F). There is a clear negative correlation between PD on the final unexpected force-field trial and the calculated restoring force perpendicular to the direction of movement (r = −0.55, P < 0.001). There is also strong negative correlation between PD on the final catch trial and the associated restoring force in the direction of the error (r = −0.68, P < 0.001). The solid lines show linear fits to the 64 data points (8 subjects × 8 directions) in each figure.
The results of experiment 1 raise the possibility that the extent of learning in each direction, quantified as the reduction in kinematic error over training, was different. In experiment 2 we directly tested whether the magnitude of learning and the extent to which learning generalizes depend on the direction in which subjects make training movements.
Hand impedance and generalization of learning
Our results from experiment 1 showed that the magnitude of kinematic error following an unexpected change in environmental dynamics was associated with hand stiffness. Our goal in experiment 2 was to evaluate whether directional differences in impedance and the associated differences in kinematic error result in differences in the amount of motor learning and the extent to which it generalizes. We hypothesize that movements in directions associated with larger kinematic error will result in a greater degree of learning and thus greater generalization of learning to movements in neighboring directions.
To test our hypothesis, we recruited two groups of 10 subjects each for experiment 2. We tested each group on a separate set of targets, as shown in Fig. 1B. The logic of the design is that each group of subjects made movements in one direction associated with high hand impedance and in one direction associated with low hand impedance; transfer of learning was assessed in an intervening direction associated with intermediate stiffness. If generalization is dependent on the extent to which subjects are required to correct for kinematic error in the training direction, then greater generalization should be observed when loads in the training condition are applied in directions of low stiffness (and accordingly subjects have to compensate for large kinematic errors). The first group of subjects made movements to targets located in the 45, 90, and 135° directions, whereas the second group of subjects was asked to move to the 135, 180, and 225° targets. The directions of maximum and minimum hand stiffness correspond roughly with the 135–315 and the 45–225° axes, respectively. The 90 and 180° directions are associated with intermediate hand stiffness and we used these directions to measure generalization of learning. As in experiment 1, subjects’ hands deviated most when moving in the 135° direction and least when moving in the 45 or 225° directions. The observed deviations are consistent with stiffness in the direction of applied force, which in each case is perpendicular to the direction of movement. It should be noted that the actual direction of maximum stiffness for the subjects who participated in experiment 1 was 135.1°, a result that agrees with previous estimates of hand stiffness obtained by our group and by others (Darainy et al. 2004, 2006; Gomi and Kawato 1997; Mussa-Ivaldi et al. 1985; Tsuji et al. 1995).
Figure 3 shows the results for the 10 subjects tested in Group 1. Figure 3, A–C shows the perpendicular deviation of the hand from a straight-line trajectory measured at maximum tangential hand velocity. Panels A, B, and C of Fig. 3 each show movements in a single training direction (135, 45, and 90°, respectively) and the subsequent effect of training on movements in the reference direction (90°). The three columns in each panel show movements from three blocks in the experiment (see Fig. 1C). The first column shows movements in the training direction under null-field conditions (light blue points). The second column shows movements in the training direction under force-field conditions (dark blue points) and also catch trials (red points). The third column shows movements in the reference direction (90°) under null-field conditions (green points). The goal of the experiment was to examine how PD on aftereffect movements in the reference direction differed, depending on the extent of adaptation in the training directions. Figure 3, A–C also shows that there are differences in the magnitude of PD across the three training directions. This is consistent with our findings in experiment 1. The figures also show that PD on aftereffect movements in the reference direction differs depending on the training direction. When training movements and aftereffect movements were both made in the reference direction, aftereffects were the largest. Aftereffects following training in the 135° direction, which is associated with larger PD and thus greater adaptation, are larger than the aftereffects that follow training in the 45° direction. This difference occurs despite an equal separation of 45° from the reference direction. Figure 3D quantifies this effect. It shows mean PD on the initial aftereffect movement, following training in the 45, 90, and 135° directions, respectively.
FIG. 3.

Training and transfer movements for targets located at 135, 45, and 90°. Data in A–C are plotted as follows. Data points show mean perpendicular deviation of the hand (±1SE) from a straight line connecting movement start and end, measured at maximum tangential velocity. Following null-field movements (50 trials, light blue points) in a given training direction, subjects made force-field training movements (120 trials, dark blue points) in the same direction. On 10% of training movements, the force field was unexpectedly removed (catch trials, red points). Subjects then made transfer movements in the 90° reference direction (50 trials, green points). A: training movements made in the 135° direction, followed by transfer movements made in the 90° reference direction. B: training movements made in the 45° direction, followed by transfer movements in the 90° direction. C: training and subsequent transfer movements were made in the 90° direction. D: perpendicular deviation on the 1st transfer movement in the 90° reference direction following training in the 3 directions indicated. Subjects trained in the 135° direction, in which the force field acted in the direction of minimum hand stiffness, showed more transfer to the 90° reference direction than subjects who made training movements in the 45° direction.
Figure 4 shows a similar result for the second group of subjects, who were tested relative to a reference direction located at 180°. Following null-field movements in each of the training directions, subjects made training movements under force-field conditions in the 135° (Fig. 4A), 225° (Fig. 4B), or 180° directions (Fig. 4C). Following each training session subjects immediately made movements toward the 180° target under null-field conditions to quantify transfer of learning to the reference direction. The results were similar to those presented in Fig. 3. PD on the initial aftereffect movement was largest when training and aftereffect movements were made in the reference direction (180°). Aftereffects were larger following training in the 135° direction, in which the kinematic deviations were in a direction associated with low hand impedance than they were following training in the 225° direction (Fig. 4D). Once again this suggests that following training in directions associated with larger kinematic error and thus greater adaptation, transfer of dynamics learning is also greater.
FIG. 4.

Training and transfer movements for targets located at 135, 225, and 180°. Data are plotted in the same manner as in Fig. 3. A: training movements made in the 135° direction, followed by transfer movements made in the 180° reference direction. B: training movements made in the 225° direction, followed by transfer movements in the 180° direction. C: training and subsequent transfer movements were made in the 180° direction. D: perpendicular deviation on the 1st transfer movement in the 180° reference direction following training in the 3 directions indicated. Subjects trained in the 225° direction, in which the force field displaced the limb in the direction of maximum hand stiffness, showed less transfer to the 180° reference direction than subjects who made training movements in the 135° direction.
To statistically quantify our results, we performed a three-way mixed-factor ANOVA on initial aftereffect movements for subjects tested relative to both the 90 and the 180° reference directions. This ANOVA allowed us to assess whether the pattern of kinematic error on aftereffect movements was different, depending on which reference direction was used, on the order in which subjects made movements to the training targets, or depending on whether subjects made training movements in directions associated with high stiffness, low stiffness, or in the reference direction. Our analysis revealed that the pattern of perpendicular deviation on initial aftereffect movements did not differ between subjects tested in the 90 versus the 180° reference directions [F(1,16) = 0.637, P > 0.4]. Moreover, the pattern of kinematic error on aftereffect movements did not differ, depending on the order in which subjects made movements to the training targets [F(1,16) = 0.50, P > 0.4]. Instead, the magnitude of kinematic error on aftereffect movements in the reference direction depended only on the direction in which subjects had trained just prior to testing [F(2,32) = 19.5, P < 0.001]. Post hoc comparisons revealed that aftereffects following training to targets at 45 and 225°, where the limb was deflected in directions of high stiffness, were smaller than those following training in the other directions (P < 0.01 in both cases). Furthermore, our post hoc tests revealed that the magnitude of initial aftereffects following training in the reference directions (90 and 180°) was larger than that following training at 135°, where the limb was displaced in a direction of low stiffness. This comparison was marginally reliable by a post hoc test (P = 0.05).
In experiment 1, we found that kinematic error differed, depending on the direction of movement, for both initial movements in the force field and for final catch trials following adaptation. This result suggests that there may be directional differences in the extent of learning. In experiment 2 we directly tested this possibility. A paired-samples t-test revealed that the magnitude of kinematic error for the initial movement in the force field was larger when the limb was displaced in directions associated with low hand stiffness than it was when displacements occurred in directions associated with high hand stiffness (P < 0.01). By the end of training, a paired-samples t-test on the final 10 training movements revealed that these directional differences in kinematic error had been eliminated (P > 0.1), suggesting that in both directions the same asymptotic level of performance was achieved. This suggests that the magnitude of dynamics learning, defined as the reduction of kinematic error, varies depending on movement direction. Subjects were then tested in a common reference direction and differences in the magnitude of aftereffects were observed, as detailed earlier. Importantly, the training directions were both located 45° from the reference direction. Thus differences in aftereffect magnitude cannot be attributed to differences in the distance between the training and reference directions (see discussion and Fig. 5). Instead, the difference in aftereffect magnitude suggests that the extent of learning was different when subjects trained in high versus low stiffness directions.
FIG. 5.

Generalization gradients can be rendered symmetrical when the extent of transfer is normalized by the magnitude of learning. A: mean (±1SE) normalized transfer of learning for subjects trained relative to the 90° reference direction in experiment 2. PD on the initial aftereffect trial is normalized by the magnitude of PD on the final catch trial during training. A symmetrical generalization gradient results from this normalization procedure. B: as in A, mean (±1SE) normalized transfer of learning for subjects trained relative to the 180° reference direction.
Control analyses for experiment 1
We performed an ANOVA to determine whether there were directional differences in subjects’ performance at the end of the 120 training movements. We found that PD, averaged over the final 10 force-field trials, differed depending on movement direction [F(7,49) = 4.628, P < 0.01]. Post hoc comparisons revealed that movements toward the 90° target showed more deviation at the end of training than movements toward the 0° target (P < 0.05). However, this was the only reliable difference. Movements in all other directions showed deviation at the end of training that did not differ. This suggests that in all but one case, subjects’ performance reached the same asymptotic level following 120 training movements. This occurred in spite of the differences in initial deflection that are associated with the anisotropic pattern of hand stiffness (Fig. 2A).
We also performed an analysis to ensure that the directional differences in PD were not due to underlying differences in movement kinematics. Specifically, we performed two-way, repeated-measures ANOVAs on peak velocity and movement duration to assess whether there were differences in either variable that depended on movement direction. We computed averages across all trials in each of four phases of the experiment: initial null-field movements, training movements, catch trials, and aftereffect movements. ANOVA revealed that there were directional differences in peak velocity in two phases of the experiment [i.e., we found a reliable interaction; F(21,147) = 8.194, P < 0.01]. However, our post hoc comparisons revealed that these differences were not systematic. Out of the 112 possible comparisons between 8 directions across the 4 experimental phases (i.e., [8 choose 2] × 4), only 6 pairwise differences were reliable. There was no specific pattern in the phases and the directions in which the reliable differences were found. Similarly, the second ANOVA on movement duration found that in three phases of experiment there are some instances in which duration depended on the direction of movement [i.e., there was a reliable interaction; F(21,147) = 1.821, P < 0.05]. Again, post hoc comparisons found that only 8 of 112 possible pairwise comparisons were reliable. Interestingly, the differences identified by the two ANOVAs involved different groups on all but one occasion, again emphasizing the lack of a systematic effect of movement direction on movement kinematics.
In experiment 1, each subject was tested in all eight training directions. This raises the possibility that the directional differences in kinematic error when the force field was activated (shown in Fig. 2, A and C) were due to carryover of learning from training in previous directions. We designed our experiment to minimize this possibility. First, the initial direction in which subjects were trained was different for each of the eight subjects and each subject made movements in the eight directions in a different order. Second, we included 40 null-field trials following training in a given direction to wash out the effects of learning. Finally, subjects performed 40 further null-field trials prior to activation of the force field in the next direction in the testing sequence. To verify that there were no effects of previous training on the kinematic error that we observed at the beginning of force-field training, we performed the following analysis. We examined kinematic error on the final 10% of null-field trials prior to the activation of the force field (i.e., on the final 4 null-field trials). A one-way, repeated-measures ANOVA revealed that immediately prior to encountering the force field, subjects in the eight directions showed no differences in kinematic error [F(7,49) = 1.588, P > 0.1]. This suggests that the difference in initial deviations detailed in Fig. 2, A and C cannot be attributed to carryover effects from previous training in other directions.
Control analyses for experiment 2
In the final phase of experiment 2, subjects made movements to the 90 or the 180° target to determine the magnitude of aftereffects following direct training to these targets. To verify that these movements were unaffected by the preceding sequence of training and testing we performed the following control study. We recruited two groups of 10 subjects each and had them make movements to the 90 or the 180° target without any prior training in other directions. The procedure was otherwise the same as that in the main experiment. Subjects initially made movements while the robot applied no forces. The force field was then turned on and subjects made 120 training movements. On 10% of trials, the forces were unexpectedly turned off (catch trials). To determine whether prior training affected performance we performed two-way, mixed-factor ANOVAs on movement duration, maximum velocity, and PD at maximum tangential hand velocity. We computed averages for each of these measures during four intervals throughout testing: on the final 10 null-field movements, on the initial 10 movements in the force field, on the final 10 movements in the force field, and on catch trials. In both the 90 and the 180° directions, we sought to determine whether subjects with prior training and naïve subjects performed differently. Our analyses revealed that in the 90° direction, subjects with prior training and naïve subjects performed no differently across the four phases of the experiment for each of the performance measures (i.e., there were no interactions). Compared with naïve controls, subjects with prior training performed no differently in terms of movement duration [F(1,18) = 1.935, P > 0.3], peak velocity [F(1,18) = 1.163, P > 0.2], or PD [F(1,18) = 0.053, P > 0.8]. We likewise found that in the 180° direction, subjects with or without prior training performed no differently across the four phases of the experiment (again, we found no interaction for any measure). Naïve subjects and subjects with prior experience did not differ in terms of movement duration [F(1,18) = 4.098, P > 0.05], peak velocity [F(1,18) = 2.361, P > 0.1], or maximum perpendicular deviation [F(1,18) = 2.568, P > 0.1]. These results indicate that movements in the 90 and 180° directions in the final phase of experiment 2 were not significantly affected by previous training in other directions.
In experiment 2, we tested for transfer of learning in either the 90° (Fig. 1C, Group 1) or the 180° (Fig. 1C, Group 2) direction following training in directions associated with low or high arm impedance. Each test for transfer of learning involved 50 movements in the reference direction. This enabled us both to assess transfer and to wash out the effects of previous training and return performance in the reference direction back to baseline levels. To determine the effectiveness of these washout trials, we performed the following analysis. We performed two one-way, repeated-measures ANOVAs on kinematic error over the final 10% of washout trials. Following training in the 45, 90, or 135° directions, ANOVA revealed that there were no differences in PD on the final five trials in the 90° direction [F(2,18) = 0.206, P > 0.8]. Likewise, there were no differences in kinematic error for the final movements in the 180° direction following training in the 135, 180, or 225° directions [F(2,18) = 0.177, P > 0.8]. These results suggest that, following tests for transfer of learning, any residual effects of training were effectively washed out and thus our subsequent tests for transfer were uncontaminated by carryover from previous conditions.
DISCUSSION
Musculoskeletal geometry has a substantial impact on the mechanical impedance of the arm. Arm impedance is anisotropic such that in certain directions, external disturbances are resisted more than in other directions (Burdet et al. 2001; Darainy et al. 2004; Gomi and Kawato 1997; Mussa-Ivaldi et al. 1985; Perreault et al. 2002). In this study, we have assessed the extent to which this anisotropic pattern of hand impedance influences dynamics learning and its generalization. In experiment 1, we tested for the presence of a correlation between arm impedance and the extent to which the force field perturbed the hand from a straight-line trajectory during reaching movements. We found a strong correlation, such that the subject's arm deviates most when perturbed in directions in which arm impedance is the least and deviates least when perturbed in directions of high impedance. This suggests that arm impedance contributes to the pattern of limb displacement that occurs in the context of unexpected changes in environmental dynamics.
Evidence consistent with the idea that movement error in a new dynamic environment is linked to stiffness can be found elsewhere. The anisotropic pattern of hand deflection has been previously shown to vary with limb configuration. Malfait and Ostry (2002) reported a systematic rotation of the pattern of hand deflection when they shifted the center of workspace from the left to right. In that study the shoulder angle changed from 90° at the left to 0° at the right relative to the frontal plane and the elbow angle was held constant at 90° relative to the upper arm. The pattern of kinematic errors in response to a dynamic perturbation likewise rotated corresponding to the change in shoulder angle. Hand stiffness patterns have been shown to rotate in exactly the same fashion in relation to the angle at the shoulder (Mussa-Ivaldi et al. 1985). These two pieces of evidence together strengthen the suggestion that the anisotropic pattern of impedance and the anisotropic pattern of arm deflection are linked.
In experiment 2, we assessed how asymmetries in arm impedance led to directional differences both in the extent of dynamics learning and in the extent to which dynamics learning generalizes. Subjects learned to compensate for a force field that perturbed the arm in directions of low stiffness (resulting in maximum kinematic error) or in directions of high stiffness (resulting in minimum kinematic error). Following each training block the generalization of motor learning was assessed under null-field conditions in a reference direction that lay halfway between the two training directions. The results show that the magnitude of aftereffects—and thus transfer of learning—is greater when training movements were made in the direction associated with low stiffness and large kinematic error. This suggests that when adaptation involved the elimination of large kinematic errors, the extent of dynamics learning and, consequently, the extent to which learning generalized were greater.
In a dynamics learning task such as the one studied here, there are a number of factors that can contribute to the reduction of kinematic error. One is the mechanical impedance of limb due to limb geometry and the coactivation of antagonist muscles (impedance control). Kinematic error can also be reduced by the production of time-varying forces that compensate for loads due to the dynamic perturbation (feedforward learning). It has been shown that subjects rely extensively on feedforward learning when the forces applied to the arm are predictable and when the arm's interaction with the environment is stable (Shadmehr and Mussa-Ivaldi 1994). It has also been shown that under these conditions, cocontraction persists throughout training (Darainy and Ostry 2008). The present experiments indicate that limb impedance and feedforward control contribute to the reduction of kinematic error in a fashion that has ramifications both for extent of learning and for generalization to new movements. In directions of high impedance, kinematic error is reduced by hand impedance and, as a result, there is less kinematic error that must be accounted for by feedforward learning. Thus while impedance has the beneficial effect of reducing error, it reduces both the extent of feedforward learning and the extent to which it generalizes.
A number of recent studies on the generalization of motor learning merit comment (Donchin et al. 2003; Mattar and Ostry 2007; Thoroughman and Shadmehr 2000). The generalization functions reported in these studies have typically been shown to be symmetrical around a reference direction. Here, our data show an asymmetric generalization function for transfer of motor learning. In Mattar and Ostry (2007) the reference direction was positioned at either 135 or 315°. These two directions correspond with the directions of maximum arm stiffness. The training directions flanked the reference direction on either side and would have been associated with similar arm stiffness. Therefore according to the results reported here a symmetrical generalization function would be expected given the particular directions studied. In other studies (Donchin et al. 2003; Thoroughman and Shadmehr 2000), the generalization functions were unitless and were calculated as the average sensitivity of the current trial in a reference direction to errors on a previous catch trial in directions farther away. Importantly, movements in all possible directions were combined to create these average generalization functions. Thus the asymmetries reported here that are due to directional differences in hand impedance were likely masked by averaging across the multiple directions studied in the previous work.
Care must be taken when interpreting the asymmetry in our generalization gradient. Our results indicate that there is a difference in the absolute magnitude of transfer of learning from directions associated with high impedance and directions associated with low impedance. If subjects learn to compensate for the dynamic perturbation in a direction associated with large kinematic error and thus large adaptation, movements in the reference direction are affected more than if subjects train in a direction associated with low kinematic error. Here, this result is based on the absolute magnitude of kinematic error on movements in the test direction. In previous studies, generalization has been assessed as the relative transfer of learning to a reference direction (Donchin et al. 2003; Thoroughman and Shadmehr 2000; Thoroughman and Taylor 2005). To compare the present results with those reported previously, we have computed the relative amount of generalization by normalizing the magnitude of transfer by the magnitude of final catch-trial error in the training direction. The results are shown in Fig. 5. We also normalized the magnitude of transfer by the magnitude of initial kinematic error in the force field and by the magnitude of learning, defined as the reduction in kinematic error over training. Results of these analyses were qualitatively similar to those described in the following text. Figure 5, A and B shows normalized perpendicular deviation in the reference direction following training in the three training directions. Thus when we examine transfer as a proportion of the extent to which subjects learned to compensate for the perturbation in different training directions, the generalization gradient is rendered symmetrical. To quantitatively analyze the data shown in Fig. 5, we performed a three-way mixed-factor ANOVA on initial normalized aftereffect movements, followed by post hoc tests where appropriate. The only significant difference that we found was the effect of training direction on the initial normalized aftereffect [F(2,32) = 17.53, P < 0.001]. Post hoc comparisons found that training in the transfer direction resulted in larger normalized aftereffects than those following training in the other two directions (P < 0.01 in both cases). There are no differences in the size of normalized aftereffects when subjects trained in the direction of maximum arm impedance or the direction of minimum arm impedance (P > 0.05). Our original results suggest, however, that because the magnitude of kinematic error is not symmetrical across movement directions, there is an asymmetry in the effect of previous training on movements in the reference direction when examined in absolute (nonnormalized) terms.
In the present study, we have measured arm stiffness in the middle of the workspace under static conditions. These measurements were used to demonstrate that the extent to which the dynamic perturbation resulted in kinematic error is correlated with the arm impedance in the direction of the displacement. We believe that in the present study, stiffness is a good approximation of arm impedance as a whole. In general terms, arm impedance is a nonlinear function of the hand's position, velocity, acceleration, and additional higher-order derivatives. To simplify the estimation of arm impedance, it is usually modeled using a second-order linear differential equation in which stiffness, viscosity, and inertia matrices are assumed to be the essential components of impedance. The orientation of each of these components of arm impedance appears to be closely related under static conditions. A study by Tsuji and colleagues (1995) demonstrated that the magnitude of hand viscosity covaries well with hand stiffness and that the direction of maximum viscosity corresponds closely to the direction of maximum stiffness. In addition, Tsuji et al. (1995) showed that the direction of maximum arm inertia aligns well with the orientation of the lower arm segment. During the stiffness measurements taken in the current experiment, the lower arm segment lay along an axis of about 135–315°. Thus the orientation of the lower arm segment and therefore the orientation of maximum inertia lay close to the direction of maximum hand stiffness and, presumably, the direction of maximum viscosity as well (135.1°). Thus the close correspondence between directions of maximum stiffness, inertia, and viscosity supports the idea that, in this study, the orientation of arm stiffness can approximate the orientation of static arm impedance as a whole.
One should also consider whether stiffness measured under static conditions at the movement start position provides an adequate measure for the interpretation of kinematic error during movement. In a previous study Gomi and Kawato (1997) estimated hand stiffness at different points during reaching movements. Their results showed that arm stiffness in the vicinity of the start position closely matches arm stiffness under static conditions. Popescu et al. (2003) also showed similar results for single joint movements. Therefore early in the course of movement, where feedforward commands are assumed to play the largest role in the ongoing movement, arm impedance can be approximated by static arm stiffness. Because our measures of kinematic error were taken early on in the movement path, stiffness measured under static conditions approximates what the arm's impedance would have been at this point of the movement.
In summary, we have shown that limb impedance can play a major role in dynamics learning and its generalization. Specifically, we have shown that kinematic error in a novel dynamic environment is associated with hand stiffness. In directions of low stiffness and kinematic error, the magnitude of learning is consequently greater than that in directions of high stiffness. The extent to which dynamics learning generalizes is affected by stiffness and the magnitude of learning, such that generalization is greater following compensation for large kinematic error.
GRANTS
This research was supported by National Institute of Child Health and Human Development Grant HD-48924, Natural Sciences and Engineering Research Council of Canada, and Fonds québécois de la recherche sur la nature et les technologies Québec.
REFERENCES
- Bennett 1994.Bennett DJ Stretch reflex responses in the human elbow joint during a voluntary movement. J Physiol 474: 339–351, 1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burdet et al. 2001.Burdet E, Osu R, Franklin DW, Milner TE, Kawato M. The central nervous system stabilizes unstable dynamics by learning optimal impedance. Nature 414: 446–449, 2001. [DOI] [PubMed] [Google Scholar]
- Caithness et al. 2004.Caithness G, Osu R, Bays P, Chase H, Klassen J, Kawato M, Wolpert DM, Flanagan JR. Failure to consolidate the consolidation theory of learning for sensorimotor adaptation tasks. J Neurosci 24: 8662–8671, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darainy et al. 2004.Darainy M, Malfait N, Gribble PL, Towhidkhoh F, Ostry DJ. Learning to control arm stiffness under static conditions. J Neurophysiol 92: 3344–3350, 2004. [DOI] [PubMed] [Google Scholar]
- Darainy and Ostry 2008.Darainy M, Ostry DJ. Muscle cocontraction following dynamics learning. Exp Brain Res 190: 153–163, 2008. [DOI] [PubMed] [Google Scholar]
- Darainy et al. 2007.Darainy M, Towhidkhoh F, Ostry DJ. Control of hand impedance under static conditions and during reaching movement. J Neurophysiol 97: 2676–2685, 2007. [DOI] [PubMed] [Google Scholar]
- Franklin et al. 2003.Franklin DW, Osu R, Burdet E, Kawato M, Milner TE. Adaptation to stable and unstable dynamics achieved by combined impedance control and inverse dynamics model. J Neurophysiol 90: 3270–3282, 2003. [DOI] [PubMed] [Google Scholar]
- Frolov et al. 2006.Frolov AA, Prokopenko RA, Dufossè M, Ouezdou FB. Adjustment of the human arm viscoelastic properties to the direction of reaching. Biol Cybern 94: 97–109, 2006. [DOI] [PubMed] [Google Scholar]
- Ghahramani and Wolpert 1997.Ghahramani Z, Wolpert DM. Modular decomposition in visuomotor learning. Nature 386: 392–395, 1997. [DOI] [PubMed] [Google Scholar]
- Gomi and Kawato 1997.Gomi H, Kawato M. Human arm stiffness and equilibrium-point trajectory during multi-joint movement. Biol Cybern 76: 163–171, 1997. [DOI] [PubMed] [Google Scholar]
- Gomi and Osu 1998.Gomi H, Osu R. Task-dependent viscoelasticity of human multijoint arm and its spatial characteristics for interaction with environments. J Neurosci 18: 8965–8978, 1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krakauer et al. 2000.Krakauer JW, Pine ZM, Ghilardi MF, Ghez C. Learning of visuomotor transformations for vectorial planning of reaching trajectories. J Neurosci 20: 8916–8924, 2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malfait et al. 2002.Malfait N, Shiller DM, Ostry DJ. Transfer of motor learning across arm configurations. J Neurosci 22: 9656–9660, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattar and Ostry 2007.Mattar AA, Ostry DJ. Modifiability of generalization in dynamics learning. J Neurophysiol 98: 3321–3329, 2007. [DOI] [PubMed] [Google Scholar]
- Milner and Cloutier 1993.Milner TE, Cloutier C. Compensation for mechanically unstable loading in voluntary wrist movement. Exp Brain Res 94: 522–532, 1993. [DOI] [PubMed] [Google Scholar]
- Mussa-Ivaldi et al. 1985.Mussa-Ivaldi FA, Hogan N, Bizzi E. Neural, mechanical, and geometric factors subserving arm posture in humans. J Neurosci 5: 2732–2743, 1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perreault et al. 2002.Perreault EJ, Kirsch RF, Crago PE. Voluntary control of static endpoint stiffness during force regulation tasks. J Neurophysiol 87: 2808–2816, 2002. [DOI] [PubMed] [Google Scholar]
- Popescu et al. 2003.Popescu F, Hidler JM, Rymer WZ. Elbow impedance during goal-directed movements. Exp Brain Res 152: 17–28, 2003. [DOI] [PubMed] [Google Scholar]
- Scheidt et al. 2000.Scheidt RA, Reinkensmeyer DJ, Conditt MA, Rymer WZ, Mussa-Ivaldi FA. Persistence of motor adaptation during constrained, multi-joint, arm movements. J Neurophysiol 84: 853–862, 2000. [DOI] [PubMed] [Google Scholar]
- Shadmehr and Brashers-Krug 1997.Shadmehr R, Brashers-Krug T. Functional stages in the formation of human long-term motor memory. J Neurosci 17: 409–419, 1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shadmehr and Mussa-Ivaldi 1994.Shadmehr R, Mussa-Ivaldi FA. Adaptive representation of dynamics during learning of a motor task. J Neurosci 14: 3208–3224, 1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thoroughman and Shadmehr 1999.Thoroughman KA, Shadmehr R. Electromyographic correlates of learning an internal model of reaching movements. J Neurosci 19: 8573–8588, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thoroughman and Shadmehr 2000.Thoroughman KA, Shadmehr R. Learning of action through adaptive combination of motor primitives. Nature 407: 742–747, 2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thoroughman and Taylor 2005.Thoroughman KA, Taylor JA. Rapid reshaping of human motor generalization. J Neurosci 25: 8948–8953, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuji et al. 1995.Tsuji T, Morasso PG, Goto K, Ito K. Human hand impedance characteristics during maintained posture. Biol Cybern 72: 475–485, 1995. [DOI] [PubMed] [Google Scholar]