

Summary
-
1
Streams collect runoff, heat, and sediment from their watersheds, making them highly vulnerable to anthropogenic disturbances such as urbanization and climate change. Forecasting the effects of these disturbances using process‐based models is critical to identifying the form and magnitude of likely impacts. Here, we integrate a new biotic model with four previously developed physical models (downscaled climate projections, stream hydrology, geomorphology, and water temperature) to predict how stream fish growth and reproduction will most probably respond to shifts in climate and urbanization over the next several decades.
-
2
The biotic submodel couples dynamics in fish populations and habitat suitability to predict fish assemblage composition, based on readily available biotic information (preferences for habitat, temperature, and food, and characteristics of spawning) and day‐to‐day variability in stream conditions.
-
3
We illustrate the model using Piedmont headwater streams in the Chesapeake Bay watershed of the USA, projecting ten scenarios: Baseline (low urbanization; no on‐going construction; and present‐day climate); one Urbanization scenario (higher impervious surface, lower forest cover, significant construction activity); four future climate change scenarios [Hadley CM3 and Parallel Climate Models under medium‐high (A2) and medium‐low (B2) emissions scenarios]; and the same four climate change scenarios plus Urbanization.
-
4
Urbanization alone depressed growth or reproduction of 8 of 39 species, while climate change alone depressed 22 to 29 species. Almost every recreationally important species (i.e. trouts, basses, sunfishes) and six of the ten currently most common species were predicted to be significantly stressed. The combined effect of climate change and urbanization on adult growth was sometimes large compared to the effect of either stressor alone. Thus, the model predicts considerable change in fish assemblage composition, including loss of diversity.
-
5
Synthesis and applications. The interaction of climate change and urban growth may entail significant reconfiguring of headwater streams, including a loss of ecosystem structure and services, which will be more costly than climate change alone. On local scales, stakeholders cannot control climate drivers but they can mitigate stream impacts via careful land use. Therefore, to conserve stream ecosystems, we recommend that proactive measures be taken to insure against species loss or severe population declines. Delays will inevitably exacerbate the impacts of both climate change and urbanization on headwater systems.
Keywords: multiple stressors, urbanization, fish assemblage, headwater stream, siltation, flow regime, temperature regime, urban stream syndrome
Introduction
As topographical low points in the landscape, aquatic systems collect and disperse water, sediment, and heat, integrating changes occurring throughout watersheds. Urban development near streams alters inputs (including nutrients, contaminants and sediment), flashiness of discharge, and temperature (Paul & Meyer 2001; Morgan & Cushman 2005). Climate change is also expected to have far‐reaching impacts on streams, from altering temperature and runoff regimes to increasing the frequency and intensity of droughts and floods (Milly, Dunne & Vecchia 2005; Alcamo, Flörke & Märker 2007). It is difficult to predict the combined effects of climate change and urbanization from empirical data because changes are occurring on broad spatial and temporal scales, and typically, conditions cannot be replicated. Yet, anticipating the combined environmental impacts of such anthropogenic changes is critical to developing proactive strategies to protect ecosystems and the services they provide (Clark et al. 2001; Walsh, Fletcher & Ladson 2005; Palmer et al. 2008a,b). Thus, process‐based models are instructive tools for investigating these complex stressors in a timely manner. Here we present the results from models we developed and parameterized to predict the separate and combined effects of urbanization and climate change on streams in small catchments in the mid‐Atlantic USA (Fig. 1).
Figure 1.
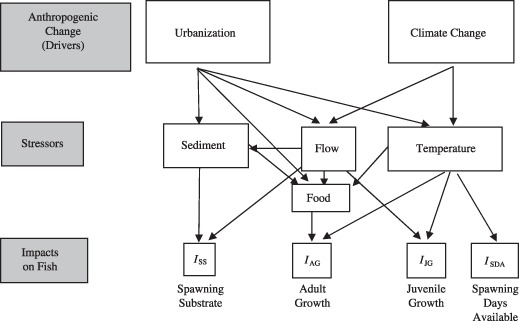
Drivers and stressors leading to indices of impacts on fish growth and reproduction. Arrows signify direct effects.
We ask how climate and urbanization‐induced changes in hydrology, geomorphology, and temperature affect entire assemblages, integrating five submodels: downscaled climate projections, hydrology, geomorphology, water temperature, and fish growth and reproduction. The first four have been described elsewhere (Dettinger et al. 2004; Moglen et al. 2004; Nelson & Palmer 2007; Hejazi & Moglen 2008; Pizzuto et al. 2008). The fifth, Forecasted Indices for Fish (FIF), uses output from the other submodels to integrate with a matrix of species traits to forecast conditions for fish growth and reproduction under selected scenarios of urbanization and climate change. We use the FIF model to predict impacts in Piedmont headwater streams of the Chesapeake Bay watershed; these streams have been the subject of extensive empirical work (e.g. Palmer et al. 2002; Moglen et al. 2004; Moore & Palmer 2005; Allmendinger et al. 2007; Nelson & Palmer 2007) to develop and parameterize the physical submodels.
We focus on fish because they are widely used indicators of environmental quality (Karr 1981; Fausch et al. 1990) and because their life histories and habitat requirements are better known than those of smaller stream biota such as macroinvertebrates (Angermeier 1995). Modelling the entire fish assemblage allows us to compare the vulnerability of functional groups (e.g. feeding, spawning, temperature). While there is a rich literature on stream fishes, the impacts of climate change on their resource base are not well understood. Changes in food availability (an indirect effect) may exert as much control over fish growth and reproduction as do changes in temperature or flow regimes (direct effects). Similarly, the exact relationship between siltation and spawning could not be parameterized from the literature, although the effect is well‐established. We felt it was imperative to include such effects, and made reasonable assumptions and rules based on the best available information; we explicate these assumptions throughout the study.
Methods
The FIF model (Fig. 2) uses a daily time‐step with inputs of air temperature, precipitation, and food availability; hydrologic, geomorphic, and water temperature outputs are generated from their respective submodels; and the impacts on fishes are predicted. We simulated Baseline conditions and 9 future scenarios (Urbanization Alone, 4 Climate Change scenarios, 4 Urbanization with Climate Change scenarios). Baseline conditions were based on data from first‐ to third‐order perennial streams in four watersheds currently dominated by residential development, (Fig. 3, also Moglen et al. 2004; Moore & Palmer 2005).
Figure 2.
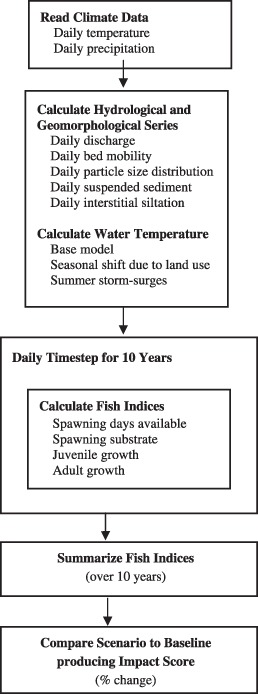
Overview of the FIF (Forecasted Indices for Fish) model.
Figure 3.
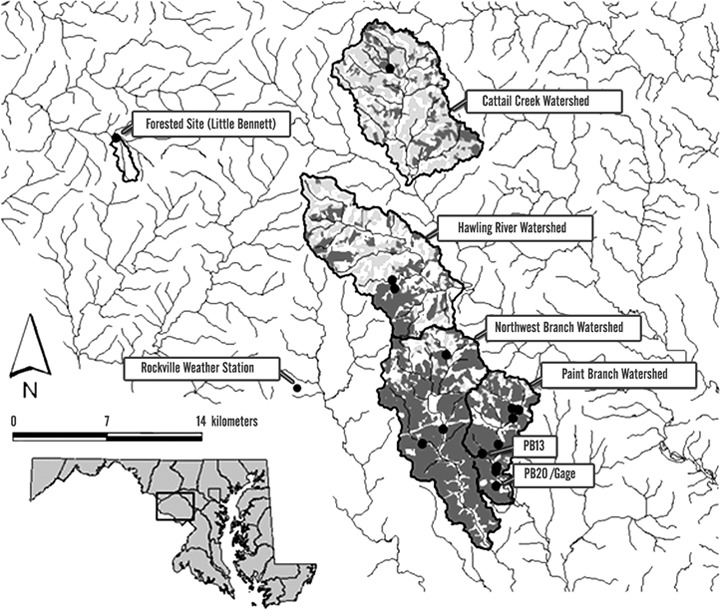
Study site locations (five watersheds outlined, with specific sites indicated by black dots), gauging site, and weather station. Within the watershed boundaries, dark grey represents urban land, light grey represents agricultural land, and white represents forested land.
urbanization
Three land‐use variables drive the hydrologic and geomorphic submodels (Table 1): (i) impervious surface [RESAC (2004); USGS (2005) data bases], which affects infiltration capacity, (ii) % new construction (average change in urban land use over the last 10 years), which affects sediment input, and (iii) % of watershed forested (the sum of deciduous, evergreen, and mixed‐forest land), which affects water temperature and organic input. We did not include agricultural land use in our scenarios, despite its distinct sedimentation regimes, for these two reasons: the output from the hydrologic submodel is similar for agricultural versus residential land use (McCuen & Snyder 1985; Hejazi & Moglen 2008); and the urbanizing watersheds surrounding Washington D.C. have little remaining agricultural land.
Table 1.
Summary of the ten land use × climate change scenarios used to predict impacts on stream fish assemblages
| % impervious | % forested | Presence of riparian buffer | % watershed under construction | Climate | |
|---|---|---|---|---|---|
| Baseline | 10 | 20 | Yes | 0 | Present* |
| Climate Change only | 10 | 20 | Yes | 0 | Future† |
| Urbanization only | 30 | 2 | No | 2 | Present* |
| Urbanization + Climate Change | 30 | 2 | No | 2 | Future† |
*Present climate is taken from the years 1995 to 2004 based on historical simulations by the HadCM3 model and statistically downscaled to match observed historical distributions (Supporting Information, Appendix S1). We used historical simulations to ensure uniformity among the climate drivers.
Four different future climate change scenarios were used, as described in text and Table 3.
Land use in our scenarios reflected conditions in our study sites: 10% impervious surface, intact riparian buffers, and no on‐going construction in the non‐urbanized scenarios; 30% impervious cover, deforested riparian buffers, and 2% new construction per year in the urbanized scenarios.
climate change
In order to obtain plausible, coherent descriptions of future climate, we rely on simulations from two atmosphere–ocean general circulation models – the Hadley Centre Model version 3 (HadCM3; Pope et al. 2000), and the Parallel Climate Model (PCM; Washington et al. 2000). These were run under two sets of future emissions assumptions – the A2 (mid‐high) and B2 (mid‐low), developed by the IPCC Special Report on Emissions Scenarios (Nakićenovićet al. 2000). To project future conditions, we used downscaled climate projections with a daily timestep for the years 2085 to 2094, which allowed us to capture reasonable predictions of day‐to‐day variability and covariance. Further details on the climate projections are provided in Supporting Information, Appendix S1.
physical submodels
The hydrologic submodel (Moglen et al. 2004) is a continuous streamflow model that is conceptually consistent with HSPF (Crawford & Linsley 1966; Bicknell et al. 1997). It predicts daily streamflow over the course of the scenario, thus allowing us to quantify flashiness. The geomorphic submodel (Pizzuto et al. 2008) is a sediment transport model with daily output including particle size distribution, bed load and suspended material discharge, turbidity, and interstitial clogging. The temperature submodel (Nelson & Palmer 2007) predicts minimum and maximum in‐stream temperature based on daily air temperatures derived from the downscaled climate projections, % deforestation, and watershed size. Calibrated models that predict fish‐food availability as a function of flow, temperature, and geomorphic conditions do not exist, so we used data series to represent reasonable estimates of food availability in each scenario (Supporting Information, Appendix S2). These data series were intentionally conservative in terms of predicting impacts of stressors.
modelling fish response
Thirty‐nine species of fish occur in our study watersheds, with small minnows making up >60% of total individuals (Supporting Information, Appendix S3; MBSS 2004). Information for each species was collected from compilations and surveys listed in Supporting Information, Appendix S3, and extrapolated from related species when necessary. We identified guilds likely to be vulnerable to urbanization and climate change: (i) fish classified as cold or cool water (vs. warm water or cool‐to‐warm water); (ii) fish breeding only at low temperatures or over relatively short seasons (vs. those that breed at a wider range of temperature or season): (iii) fish that use some sort of breeding structure that could trap silt (versus fish that do not use a breeding structure); (iv) fish that do not clean or aerate their eggs (versus cleaners/aerators); (v) fish that feed exclusively on invertebrates and/or other fish (versus those that also feed on detritus or algae). Except for (iii), these guilds are well accepted in the biomonitoring literature (Karr 1981; Rowe et al. 2003).
We developed four indices to summarize the impacts of urbanization and climate change on fishes (Table 2; Supporting Information, Appendix S4). Two indices focused on spawning (I SDA, spawning day availability; I SS, spawning substrate) and two focused on growth (I JG, juvenile growth; I AG, adult growth). Index values ranged from 0 (conditions unacceptable) to 1 (conditions acceptable). To demonstrate the overall impact of the anthropogenic stressors, we calculated the percentage change between the index values for any given stressor scenario vs. the Baseline scenario (hereafter, called impact score). For example, an impact score of −10 points means that the raw scores for a stressor scenario decreased by 10% relative to the baseline scenario.
Table 2.
. Indices developed to summarize the impacts of urbanization and climate change on fish assemblages for the FIF model*
| Indices | Explanation | Environmental factors contributing | Species characteristics contributing |
|---|---|---|---|
| I SDA index of spawning day availability (% days during the typical spawning period for each species when predicted water temperature falls within the spawning temperature range) | Effect of warming on days available for spawning | Daily temperature | Spawning times |
| Spawning temperatures | |||
| I SS index of spawning substrate (% spawning days impacted by siltation†) | Effect of siltation on spawning | Discharge Siltation | Spawning care |
| Spawning mode | |||
| Spawning months | |||
| I SS index of juvenile growth (no of days required to grow to maturity given predicted temperature and likelihood of eggs/young being washed out by high flows‡) | Effect of warming and washout on development time for juveniles | Daily temperature | Spawning times |
| Summer temperature surges | Temperature group | ||
| Time to 40 mm length | |||
| I AG index of adult growth (% days on which ‘positive growth’ was possible§) | Days during which positive growth can occur for adults | Daily temperature | Temperature group |
| Food availability | Adult food preferences | ||
| Summer temperature surges |
*full details and assumption justifications in Supporting Information, Appendix S4.
Siltation impedes flow of interstitial oxygen and depresses hatching rates of eggs (Soulsby et al. 2001; Lapointe et al. 2004). Species that clean and aerate their nests or position their eggs to avoid siltation were assumed less vulnerable (Johnston 1999). The index also reflected the availability of appropriately sized spawning substrate for species that build nests or redds.
We assumed that juveniles had high growth rates only in the middle half of the ‘good growth’ range for adults (Rombough 1997).
Positive growth was possible when water temperature was within ‘good growth’ range for the species’ temperature guild and at least one of the food types eaten by the species (detritus, algae, invertebrates, and/or small fishes) was above a non‐limiting threshold.
We used an independent data set on fish species assemblages across urbanization gradients to validate the use of these indices (Supporting Information, Appendix S5). A corresponding gradient for climate change does not exist, and hence, we could not validate the indices with respect to climate change. Our validation exercise revealed that three of our indices –I AG, I JG, and a composite score averaging all four indices – produced rates of species loss that were consistent with observed patterns across the gradient (i.e. index scores were generally successful in identifying vulnerable species). ISDA produced no trend, perhaps because the number of spawning days does not change much with urbanization, whereas we expect this factor to be more sensitive to climate change scenarios. Since our validation suggested that the fish growth indices (I JG and I AG) best predicted vulnerable species, we only used these two indices in the work reported here. To be conservative, we did not use the composite score, although it exhibited significant trends in third‐order streams, because it contained non‐significant individual index scores.
For each scenario, we counted the number of species that suffered a loss of 10 points or greater for either or both of the growth indices. This quantifies the overlap between those species stressed by each pathway (juvenile or adult growth) and is a rough estimate of the composite effect of a given scenario.
Results
physical submodels
Overall, the climate change conditions could be characterized by change in temperature and rainfall (Table 3). Here, we use the HadCM3 B2 scenario to illustrate physical responses to urbanization and climate change (4, 5). Minimum daily discharge did not vary under the HadCM3 B2 runs, but maximum daily discharge did. High‐flow days moved about 17% more water under Urbanization compared to Baseline, 27% more under Climate Change, and 45% more under simultaneous stressors. By contrast, median discharge, which represents ‘normal’ levels, decreased about 7% compared to Baseline under Urbanization, reflecting lower infiltration and groundwater levels, but increased 40% under Climate Change, reflecting higher annual precipitation.
Table 3.
Climate change driver series used in the FIF model for baseline and future climate scenarios. P, precipitation. Full explanation in text and in Supporting Information, Appendix S1
| Statistic | Baseline | Hadley A2 | Hadley B2 | PCM A2 | PCM B2 |
|---|---|---|---|---|---|
| Mean temperature (ºC March–September) | 17·2 | 20·5 | 21·7 | 15·5 | 15·3 |
| No. of rainfall events in 10 years (>0·1 cm) | 1107 | 1087 | 1093 | 1104 | 1047 |
| Average annual P (cm) | 112·9 | 132·9 | 119·9 | 116·9 | 94·1 |
| Average P event−1 (cm) | 1·02 | 1·22 | 1·10 | 1·05 | 0·90 |
| No. of heavy P events year−1 (>10 cm) | 5 | 13 | 10 | 3 | 0 |
| Max 1‐Day P (cm) | 17·4 | 21·1 | 26·7 | 10·3 | 8·4 |
| Summary compared to present: | |||||
| Average summer temp. | warmer | warmer | |||
| Total P | wetter | drier | |||
| Heavy P events | increased | increased | decreased | decreased | |
Figure 4.
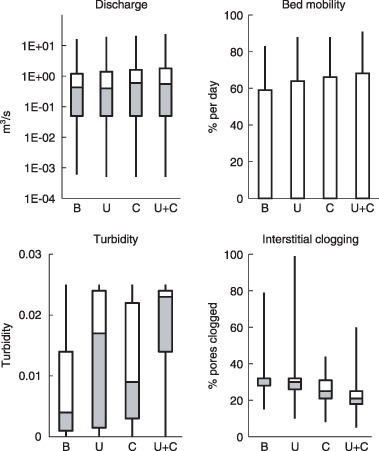
Hydrologic and geomorphic submodel outputs under four scenarios: Baseline (B), Urbanization Alone (U), HadCM3‐B2 Climate Change (C), and Urbanization plus HadCM3‐B2 Climate Change (U+C). Submodels were run for the years 2085–2094. Whiskers represent minimum and maximum values, boxes represent first and third quartiles, and dividing line represents median of Discharge, Bed Mobility, Turbidity, and Siltation over the 10‐year simulations.
Figure 5.
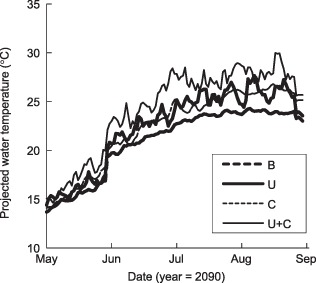
Maximum daily water temperature projected in the year 2090 using scenarios as in Fig. 4.
Minimum and median daily bed mobility (Fig. 4) were each 0%, because flow was not strong enough to move the bed most of the time. Maximum bed mobility was lowest under Baseline conditions, and increased under both Climate Change (due to increased storm size) and Urbanization (due to increased flashiness and a greater influx of sand from soil erosion associated with construction).
Turbidity (Fig. 4) exhibited similar patterns: both frequent extreme precipitation events (Climate Change scenario) and erosion from construction (Urbanization scenario) increased the supply of suspended sediment, and the effect was approximately additive when the two stressors occurred simultaneously. Siltation, or fine‐sediment deposition, was more complicated (Fig. 4). Storm flows and flashiness tend to flush out silt; minimum siltation levels were lower than Baseline under all three stressor scenarios, and median siltation levels were mostly unaffected. Maximum siltation, however, was very sensitive to stressors. Climate Change scenarios exhibited less siltation than Baseline because more episodic flows removed fine sediment from the streambed. The Urbanization scenario had extremely high siltation levels due to elevated inputs from construction. When stressors acted simultaneously, maximum siltation levels were intermediate.
Water temperature was highest in the Simultaneous scenario (Fig. 5). Inter‐scenario differences were most apparent in midsummer, when heated storm runoff made maximum daily temperature much more variable in the Urbanization scenarios. Temperature spikes in urban streams immediately after rainstorms ranged from 3·5 to >7 °C, depending on watershed size, amount of impervious surface, and immediately preceding air temperatures (Nelson & Palmer 2007).
modelling fish response
Across the 9 future scenarios, 8 to 29 of the 39 fish species were negatively affected in terms of juvenile or adult growth (Fig. 6). Urbanization alone stressed relatively few species (8 species exhibited reduced adult growth), while climate change affected most species (22 to 29, depending on the scenario). However, adding urbanization to climate change usually increased the number of stressed species, sometimes quite dramatically, suggesting that considerable change in community composition and loss of diversity could occur under future scenarios. Among the 10 currently most common species, six had at least one impact score of 10% or higher. Almost every recreationally important species (including trouts, basses, and sunfishes, all of which are currently rare), had impact scores ranging from 40 to 90%.
Figure 6.
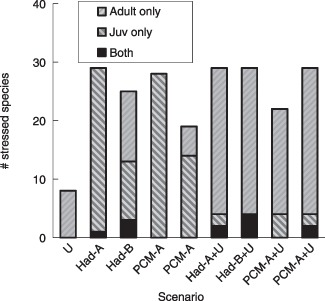
Number of species adversely affected by land use change and/or climate change (>10 point loss in impact score). Striped bars represent species affected in only one of the two indices (indices of adult growth or juvenile growth), while dark bars represent species affected in both indices.
Different scenarios produced distinctive patterns of stress; in each scenario, disparate sets of species were affected by the juvenile vs. adult stress pathways, and few were affected by both pathways. Effects on adult growth dominated under Urbanization and PCM‐based Climate Change scenarios. Effects on juvenile growth dominated under HadCM3‐based Climate Change scenarios. Adding urbanization to climate change generally increased the number of species with reduced adult growth, while it did not increase (and sometimes decreased) the number of species with reduced juvenile growth.
Discussion
We developed and tested a model to explore the effects of urbanization and climate change on stream fish assemblages by focusing on the specific pathways by which these stressors are likely to impact fish reproduction and growth. Each of the four stressor pathways – spawning temperatures, spawning substrate, juvenile growth, and adult growth (Fig. 3) – integrated direct (e.g. thermal tolerance) and indirect (e.g. food availability) effects of changes in the hydrologic and geomorphic regimes. We found that two of these pathways – stress on juvenile growth arising from altered temperature and hydrology, and stress on adult growth arising from altered temperature, siltation, and food resources – are likely to significantly influence species persistence under future land development and climate scenarios. Our results suggest that 50–75% of the fish species in small Piedmont streams will be highly stressed by future scenarios. Most of these species were affected by climate change, but adding increased urbanization to a warming climate exacerbated the number of stressed species (Fig. 6). Declines in abundance are likely for almost all species supporting recreational fisheries in these watersheds, including trout, bass, and sunfish. These predictions are probably conservative because we excluded impacts that we could not validate with an independent data set (e.g. temperature and siltation effects on spawning day availability, siltation effects on spawning substrate).
General treatments of the ecological impacts expected from climate change are now common (e.g., Eaton & Scheller 1996; Parmesan & Yohe 2003) and the impacts of urbanization on streams are well documented (Power 1997; Bledsoe & Watson 2001; Walsh et al. 2005), but we believe our work is the first to project stream impacts at the watershed scale most relevant to management. Even so, our findings are likely to have broad applicability because we studied species that are common across much of the eastern USA, described these fishes via traits that occur in many other species and regions, and modelled pathways through which either climate or land‐use change may exert local ecological impacts. However, we expect details regarding the magnitude of species‐specific effects to vary considerably among localities. Since climate change forecasts and urbanizing trends are similar for most of the coastal mid‐Atlantic, our results are certainly applicable to many mid‐Atlantic watersheds. Extending these results to other ecoregions could be done if climate change predictions are similar and watersheds are, like ours, moderately to heavily urbanized.
model realism and underlying assumptions
Our findings are instructive only to the extent that our assumptions regarding urbanization and climate change are reasonable. Our Urbanization scenario is probably conservative, given that Maryland's population is projected to increase substantially, doubling in some localities, within the next several decades (Freece 2006). Climate change projections are less certain, which is why we used multiple sets of climate change assumptions. The HadCM3 and PCM models used here are both widely applied and accepted, and our emissions scenarios are in the mid‐range of accepted future scenarios (IPCC 2007). Recent evaluations of these models for Maryland agree on increases in average summer air temperature (up to 5 °C) and in frequencies of spring floods, summer droughts, and summer heat waves, especially in urban areas (Boesch 2008). Given the potentially severe ecological impacts, we believe that policies to protect people and ecosystems cannot wait for uncertainties to be eliminated by additional research. Instead, we have attempted to synthesize reasonable assumptions and the best available scientific knowledge into projections that can aid ecosystem managers.
Our findings are probably sensitive to our assumptions about food availability. Lacking robust models to estimate availability of important foods, particularly stream invertebrates (Morin 1997), we used data from our study sites, literature‐based values, and expert judgment to develop reasonable but intentionally conservative estimates of abundance for detritus, algae, and small invertebrates. Much of our projected impact on adult fish growth stemmed from food limitation for obligate invertivores. While it is well‐established that invertebrate abundance declines under urbanization, effects of climate change are largely unknown (Poff, Brinson & Day 2002). We assumed that flashier flow would deplete invertebrates and their foods, and that high summer temperature combined with low summer flow would exacerbate this effect. However, it is also possible that warmer water temperatures could increase invertebrate growth or maturation rates, resulting in more smaller‐bodied adults. Clearly, more empirical work is needed to distinguish which effects will prevail.
Other knowledge gaps also impaired our forecasting. Both urban stream ‘flashiness’ and summer droughts may cause significant low flows and interact with altered sediment regimes to cause severe siltation. However, despite the widely recognized importance of low flow and siltation for streams (Rabeni & Smale 1995; Waters 1995; Matthews & Marsh‐Matthews 2003; Roy et al. 2003), models to predict siltation have not been developed (Kondolf 2000). Likewise, despite many studies of temperature‐induced mortality for fishes (Lutterschmidt & Hutchison 1997), sublethal effects of large temperature changes are not well understood. Temperature surges have long been assumed to occur in urban streams (e.g., Pluhowski 1970), but have only recently been documented (Nelson & Palmer 2007). Temperature surges may disproportionately affect fish eggs and young, which have greater surface‐to‐volume ratios and less ability to regulate body temperatures or avoid unsuitable water temperatures compared to adults. In our scenarios, shifts in maximum temperatures had little impact because temperatures rarely exceeded critical thermal maxima, but this may change as our understanding of temperature surges improves.
Finally, our model simplifies reality in several ways that tend to make our impact findings more conservative. First, we ignore spatial relationships (e.g. habitat connectivity), believing species reintroduction to be unimportant in small drainages without significant north–south river systems (i.e. our rivers do not extend across climatic gradients). We also ignored non‐native species introductions, which increase with human occupation of ecosystems but are not readily predicted by changes in land use or climate, and we ignored the myriad pollutants, associated with urbanization, that adversely affect stream biota (Wheeler, Angermeier & Rosenberger 2005). Finally, we do not include the effect of channelization, a frequent alteration of urban streams. Clearly, channelization would exacerbate temperature, lack of spawning substrate, and flooding issues.
relative effects of urbanization and climate change
Our results showed that only eight species were strongly affected by urbanization alone; all are obligate invertivores that were stressed by the decline in invertebrates due to siltation, urban flashiness, and lack of leaf litter input. Our indices of spawning day availability and spawning substrate predicted negative impacts on many more species under urbanization, but we excluded these indices because we could not validate them. Additionally, our watersheds have already lost species due to urbanization; impacts would be greater in a newly urbanizing area. Nevertheless, the pattern of higher impacts on juvenile vs. adult growth suggests that urbanization alone may allow most species to persist regionally if they can move among non‐urbanized refugia. But without refugia, adding the impacts of urbanization to climate change will probably drive more species to regional extinction.
Our projected impacts on fishes varied among climate change scenarios. In the HadCM3‐based scenarios, average summer temperature increased 3–4 °C and the number of scouring extreme precipitation events doubled. Many species experienced decreased juvenile growth, but food availability for adults could remain unchanged or even increase due to lower siltation and higher water temperature. In addition, strong temperature changes associated with the HadCM3‐based climate change scenarios suggest significant stress on species with brief spawning periods, as shown in Moglen et al. (2004). In contrast to the HadCM3 projections, the PCM‐based climate change scenarios projected little change in precipitation, fewer scouring extreme precipitation events, and only slight changes in temperature. Although juvenile growth rates were largely unaffected in the PCM scenarios, high siltation depressed growth of adult fishes. More precise predictions of biotic impacts are not feasible until the inherent uncertainties of climate projections are reduced.
When we combined effects of urbanization and climate change, more species were predicted to experience depressed adult growth (2 to 14 species, depending on the scenario). This impact reflected increased siltation in the PCM‐based climate change scenarios and increased flashiness in the HadCM3‐based scenarios. However, fewer species were predicted to experience depressed juvenile growth, which is surprising since this impact reflects elevated water temperature and flood frequency, both of which we expected to increase when urbanization was added to climate change. The observed decrease in impact on juvenile growth appears to be due to changes in the timing of reproduction and subsequent juvenile growth. The PCM scenarios without urbanization involve little change in water temperature, while the PCM scenarios with urbanization project higher water temperature. This difference shifts the timing of reproduction and decreases the time required for juvenile development.
The strong impacts of climate change implied in our results may seem surprising, given that water temperatures are generally well‐buffered. However, climate change also entails changes in total precipitation and the distribution of that precipitation in terms of number of events. These hydrologic changes alone will induce shifts in sediment loading, interstitial siltation, primary productivity, and so on. In addition, rather than a uniform increase of a few degrees, climate change will shift the seasonality of temperature regimes, causing further ecological repercussions. Our results do not point to definitive conclusions regarding the relative magnitude of climate‐change versus urbanization impacts on streams, but they do indicate that both are severe and that they may have synergistic impacts. This finding is significant because it suggests that wise decisions regarding ecosystem management must consider both stressors simultaneously.
implications
Proactive and reactive management actions may enable society to adapt to climate change, but reactive management may cost more because of damage to infrastructure such as roads or bridges (Palmer et al. 2008a,b). Therefore, many natural resource managers prefer proactive approaches such as increasing stormwater management to reduce peak flows and temperature stress in developed basins, or preferably, acquiring land near streams to free floodplains of infrastructure, thereby allowing re‐growth of riparian vegetation. Where land is not already heavily developed, land preservation programmes could be used to protect headwater streams. More aggressive efforts to reduce sediment and pollutant loads, including the use of staging during construction and establishing vegetative buffers along streams and roads would also reduce the stresses on stream biota.
Continued degradation of headwater streams poses a serious loss of valuable public resources. Although hydroelectric dams and major highways are unlikely to be damaged when headwater streams deteriorate, our study suggests that other less quantifiable costs will indeed be high. In particular, impacts of urbanization or climate change are likely to reduce local biodiversity (for which headwater streams are a hotspot; Meyer et al. 2007) and provision of ecosystem services (e.g. drinkable, swimmable, or fishable waters). Furthermore, urbanization combined with climate change will be more environmentally costly than either impact alone. Minimizing these costs will require resource managers, policymakers, and the public to collectively reassess the socioeconomic importance of urban streams, as well as the adequacy of programmes to protect them under future stress.
The interactions between climate change and land‐use change are interesting because of the contrasting spatial grains over which the two processes operate, and the concomitant differences in institutional scale needed to manage them. Impacts of land‐use change are integrated across watersheds. Even if management goals emphasize protecting receiving waters, such as the Chesapeake Bay, jurisdiction over land use is shared by, at most, a few states. Climate change, by contrast, results from global emissions of greenhouse gases, and can only truly be controlled by international agreement. Likewise, climate change involves time lags of decades or centuries, while land use is immediately manageable and has acute, immediate ecological effects during transitions (e.g. during construction). Finally, effects of climate change are just beginning to be recognized, while those of urbanization have been well documented in some areas for several decades, including cycles of construction and riparian disturbance.
Clearly, the consequences of urbanization and climate change for stream ecosystems have the potential to be quite significant. Society must decide, deliberately or by default, which of those consequences are acceptable. If societal goals favour conservation of stream ecosystems, our results suggest that proactive policy and/or management actions to advance conservation should have high priority in regions expected to undergo further urbanization.
Supporting information
Appendix S1. Climate projections and downscaling
Appendix S2. Submodels for components of the food web
Appendix S3. Table of fish species and their characteristics
Appendix S4. Modelling individual stressor mechanisms
Appendix S5. Testing the utility of the fish indices
Please note: Wiley‐Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Supporting info item
Acknowledgements
This work was supported by grants from the Environmental Protection Agency: Science to Achieve Results (STAR award R828012) and the Global Climate Change Program (EPA numbers: 1W0594NAEX and R83038701). We thank Montgomery County, Maryland, Department of Environmental Protection for their valuable collaboration and data contributions. We also thank Nancy Bockstael, Laura Craig, William Fagan, Holly Menninger, Dave Richardson, Catriona Rogers, and the entire Palmer laboratory group for stimulating discussions and valuable suggestions. Mention of trade names or commercial products does not imply endorsement by the US government. This is contribution number 4230 from the University of Maryland Center for Environmental Science.
Re‐use of this article is permitted in accordance with the Creative Commons Deed, Attribution 2·5, which does not permit commercial exploitation.
References
- Alcamo, J. , Flörke, M. & Märker, M. (2007) Future long‐term changes in global water resources driven by socio‐economic and climatic changes. Hydrological Sciences Journal, 52, 247–275. [Google Scholar]
- Allmendinger, N.E. , Pizzuto, J.E. , Moglen, G.E. & Lewicki, M. (2007) A sediment budget for an urbanized watershed, 1951–1996, Montgomery County, Maryland, USA. Journal of the American Water Resources Association 43, 1–16. [Google Scholar]
- Angermeier, P.L. (1995) Ecological attributes of extinction‐prone species – loss of fresh‐water fishes of Virginia. Conservation Biology, 9, 143–158. [Google Scholar]
- Bicknell, B.R. , Imhoff, J.C. , Kittle, J.L. , Donigan, A.S. & Johanson, R.C. (1997) Hydrologic Simulation Program – Fortran, User's Manual for Version 11. EPA/600/R‐97/080 . US EPA Ecosystems Research Division, Environmental Research Laboratory, Athens GA, USA. [Google Scholar]
- Bledsoe, B.P. & Watson, C.C. (2001) Effects of urbanization on channel instability. Journal of the American Water Resources Association, 37, 255–270. [Google Scholar]
- Boesch D.F., ed. (2008) Global Warming in the Free State: Comprehensive Assessment of Climate Change Impacts in Maryland. University of Maryland Center for Environmental Science, Cambridge, MD, USA. [Google Scholar]
- Clark, J. , Carpenter, S.R. , Barber, M. , Collins, S. , Dobson, A. , Foley, J.A. , Lodge, D.M. , Pascual, M. , Pielke, R. , Pizer, W. , Pringle, C. , Reid, W.V. , Rose, K.A. , Sala, O. , Schlesinger, W.H. , Wall, D.H. & Wear, D. (2001) Ecological forecasts: an emerging imperative. Science, 293, 657–660. [DOI] [PubMed] [Google Scholar]
- Crawford, N.H. & Linsley, R.K. (1966) Digital Simulation in Hydrology: Stanford Watershed Model IV, Technical Report 39. Stanford University, Department of Civil Engineering, Stanford, CA, USA. [Google Scholar]
- Dettinger, M.D. , Cayan, D.R. , Meyer, M.K. & Jeton, A.E. (2004) Simulated hydrologic responses to climate variations and change in the Merced, Carson, and American River basins, Sierra Nevada, California, 1900–2099. Climatic Change, 62, 283–317. [Google Scholar]
- Eaton, J.G. & Scheller, R.M. (1996) Effects of climate warming on fish thermal habitat in streams of the United States. Limnology and Oceanography, 41, 1109–1115. [Google Scholar]
- Fausch, K.D. , Lyons, J. , Karr, J.R. & Angermeier, P.L. (1990) Fish communities as indicators of environmental degradation. American Fisheries Society Symposium, 8, 123–144. [Google Scholar]
- Frece, J.W. (2006) Today's Vision, Tomorrow's Reality: Summary Report of the ‘Reality Check Plus’ Growth Visioning Exercises. National Center for Smart Growth Research and Education . Available at http://www.realitycheckmaryland.org/files/pdf/TVTR‐Final.pdf.
- Hejazi, M.I. & Moglen, G.E. (2008) The effects of climate and land use change on flow duration in the Maryland Piedmont Region. Hydrological Processes, 22, 4710–4722. [Google Scholar]
- IPCC (2007) Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge UK. [Google Scholar]
- Johnston, C.E. (1999) The relationship of spawning mode to conservation of North American minnows (Cyprinidae). Environmental Biology of Fishes, 55, 21–30. [Google Scholar]
- Karr, J.R. (1981) Assessment of biotic integrity using fish communities. Fisheries, 6, 21–27. [Google Scholar]
- Kondolf, G.M. (2000) Assessing salmonid spawning gravel quality. Transactions of the American Fisheries Society, 129, 262–281. [Google Scholar]
- Lapointe, M. , Bergeron, N. , Berube, F. , Pouliot, M. & Johnston, P. (2004) Interactive effects of substrate sand and silt contents, redd‐scale hydraulic gradients, and interstitial velocities on egg‐to‐emergence survival of Atlantic salmon (Salmo salar ). Canadian Journal of Fisheries and Aquatic Sciences, 61, 2271–2277. [Google Scholar]
- Lutterschmidt, W.I. & Hutchison, V.H. (1997) The critical thermal maximum: history and critique. Canadian Journal of Zoology, 75, 1561–1574. [Google Scholar]
- Matthews, W.J. & Marsh‐Matthews, E. (2003) Effects of drought on fish across axes of space, time and ecological complexity. Freshwater Biology, 48, 1232–1253. [Google Scholar]
- MBSS (2004) Maryland Biological Stream Survey . URL: http:dnr.state.md.us/streams/mbss/. Accessed in December 2004.
- McCuen, R.H. & Synder, W.M. (1985) Hydrologic Modeling: Statistical Methods and Applications. Prentice‐Hall, Englewood Cliffs, NJ, USA. [Google Scholar]
- Meyer, J.L. , Strayer, L. , Wallace, J.B. , Eggert, S.L. , Helfman, G.S. & Leonard, N.E. (2007) The contribution of headwater streams to biodiversity in river networks. Journal of the American Water Resources Association, 43, 86–103. [Google Scholar]
- Milly, P.C.D. , Dunne, K.A. & Vecchia, A.V. (2005) Global pattern of trends in streamflow and water availability in a changing climate. Nature, 438, 347–350. [DOI] [PubMed] [Google Scholar]
- Moglen, G.E. , Nelson, K.C. , Palmer, M.A. , Pizzuto, J.E. , Rogers, C.E. & Hejazi, M.I. (2004) Hydro‐ecologic responses to land use in small urbanizing watersheds within the Chesapeake Bay Watershed Ecosystems and Land Use Change (ed. DeFries R.). American Geophysical Union, Washington D.C. [Google Scholar]
- Moore, A.M. & Palmer, M.A. (2005) Invertebrate biodiversity in agricultural and urban headwater streams: implications for conservation and management. Ecological Applications, 15, 1169–1177. [Google Scholar]
- Morgan, R.P. & Cushman, S.E. (2005) Urbanization effects on stream fish assemblages in Maryland, USA. Journal of the North American Benthological Society, 24, 643–655. [Google Scholar]
- Morin, A. (1997) Empirical models predicting population abundance and productivity in lotic systems. Journal of the North American Benthological Society, 16, 319–337. [Google Scholar]
- Nakićenović, N. , Alcamo, J. , Davis, G. , De Vries, B. , Fenhann, J. , Gaffin, S. , Gregory, K. , Grübler, A. , Jung, T.Y. , Kram, T. et al (2000) IPCC Special Report on Emissions Scenarios. Cambridge University Press, Cambridge, UK. [Google Scholar]
- Nelson, K.C. & Palmer, M.A. (2007) Stream temperature surges under urbanization and climate change: data, models, and responses. Journal of the American Water Resources Association, 43, 440–452. [Google Scholar]
- Palmer, M.A. , Moglen, G.E. , Bockstael, N.E. , Brooks, S. , Pizzuto, J.E. , Wiegand, C. & VanNess, K. (2002) The ecological consequences of changing land use for running waters: the suburban Maryland case. Yale Bulletin of Environmental Science, 107, 85–113. [Google Scholar]
- Palmer, M.A. , Lettenmeier, D. , Poff, L. , Postel, S. , Richter, B. & Warner, R. (2008a). Adaptation options for climate‐sensitive ecosystems and resources: Wild and scenic rivers. Synthesis and Assessment Product 4·4. US Climate Change Science Program.
- Palmer, M.A. , Reidy, C.A. , Nilsson, C. , Flörke, M. , Alcamo, J. , Lake, P.S. & Bond, N. (2008b) Climate change and the world's river basins: anticipating management options. Frontiers in Ecology and the Environment. 6: 81–89. [Google Scholar]
- Parmesan, C. & Yohe., G. (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature, 37–42. [DOI] [PubMed] [Google Scholar]
- Paul, M.J. & Meyer, J.L. (2001) Streams in the urban landscape. Annual Review of Ecology and Systematics, 32, 333–365. [Google Scholar]
- Pizzuto, J.E. , Moglen, G.E. , Palmer, M.A. & Nelson, K.C. (2008) Two model scenarios illustrating the effects of land use and climate change on gravel riverbeds of suburban Maryland, U.S.A Gravel Bed Rivers VI: From Process Understanding to the Restoration of Mountain Rivers (eds Rinaldi M., Habersack H. & Piegay H.). Elsevier, Amsterdam, The Netherlands. [Google Scholar]
- Pluhowski, E.J. (1970) Urbanization and its effect on the temperature of the streams on Long Island, New York. U S Geological Survey, Professional Paper 627‐D.
- Poff, N.L. , Brinson, M.M. & Day, J.W. (2002) Aquatic Ecosystems and Global Climate Change. Pew Center on Global Climate Change, Arlington VA, USA. [Google Scholar]
- Pope, V.D. , Gallani, M.L. , Rowntree, P.R. & Stratton, R.A. (2000) The impact of new physical parameterizations in the Hadley Centre climate model HadCM3. Climate Dynamics, 16, 123–146. [Google Scholar]
- Power, M. (1997) Assessing the effects of environmental stressors on fish populations. Aquatic Toxicology, 39, 151–169. [Google Scholar]
- Rabeni, C.F. & Smale, M.A. (1995) Effects of siltation on stream fishes and the potential mitigating role of the buffering riparian zone. Hydrobiologia, 303, 211–219. [Google Scholar]
- RESAC (2004) Land cover mapping of the Chesapeake Bay Watershed. Department of Geography, University of Maryland. URL: http:geog.umd. edu/resac/. Accessed in December 2004. [Google Scholar]
- Rombough, P.J. (1997) The effect of temperature on embryonic and larval development Global Warming: Implications for Freshwater and Marine Fish (eds Wood C.M. McDonald &. D.G.). Cambridge University Press, Cambridge, UK. [Google Scholar]
- Rowe, D.K. , Dean, T.L. , Williams, E. & Smith, J.P. (2003) Effects of turbidity on the ability of juvenile rainbow trout, Oncorhynchus mykiss, to feed on limnetic and benthic prey in laboratory tanks. New Zealand Journal of Marine and Freshwater Research, 37, 45–52. [Google Scholar]
- Roy, A.H. , Rosemond, A.D. , Paul, M.J. , Leigh, D.S. & Wallace, J.B. (2003) Stream macroinvertebrate response to catchment urbanization (Georgia, USA). Freshwater Biology, 48, 329–346. [Google Scholar]
- Soulsby, C. , Youngson, A.F. , Moir, H.J. & Malcolm, I.A. (2001) Fine sediment influence on salmonid spawning habitat in a lowland agricultural stream: a preliminary assessment. Science of the Total Environment, 265, 295–307. [DOI] [PubMed] [Google Scholar]
- USGS (2005) The USGS Land Cover Institute (LCI): National Land Cover Dataset 1992. URL: http:usgs.gov/natllandcover.php. Accessed in March 2005.
- Walsh, C.J. , Fletcher, T.D. & Ladson, A.R. (2005) Stream restoration in urban catchments through redesigning stormwater systems: looking to the catchment to save the stream. Journal of the North American Benthological Society, 24, 690–705. [Google Scholar]
- Walsh, C.J. , Roy, A.H. , Feminella, J.W. , Cottingham, P.D. , Groffman, P.M. & Morgan, R.P. (2005) The urban stream syndrome: current knowledge and the search for a cure. Journal of the North American Benthological Society, 24, 706–723. [Google Scholar]
- Washington, W.M. , Weatherly, J.W. , Meehl, G.A. , Semtner, A.J. , Bettge, T.W. , Craig, A.P. , Strand, W.G. , Arblaster, J. , Wayland, V.B. , James, R. & Zhang, Y. (2000) Parallel Climate Model (PCM) control and transient simulations. Climate Dynamics, 16, 755–774. [Google Scholar]
- Waters, T.F. (1995) Sediment in Streams: Sources, Biological Effects, and Control. American Fisheries Society, Bethesda, MD, USA. [Google Scholar]
- Wheeler, A.P. , Angermeier, P.L. & Rosenberger, A.E. (2005) Impacts of new highways and subsequent landscape urbanization on stream habitat and biota. Reviews in Fisheries Science, 13: 141–164. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1. Climate projections and downscaling
Appendix S2. Submodels for components of the food web
Appendix S3. Table of fish species and their characteristics
Appendix S4. Modelling individual stressor mechanisms
Appendix S5. Testing the utility of the fish indices
Please note: Wiley‐Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Supporting info item