

Abstract
Curt Richter introduced behavioral control into the concept of homeostasis, thereby opening entire fields of research. The prevailing dogma, and the techniques he used, conspired to lead Richter and others to interpret regulation in strict negative feedback terms. Although this point of view continues to be embraced by many contemporary biologists, we believe that prevailing sentiment favors a broader view in which organisms integrate anticipatory pre-emptive control over regulated variables whenever possible.
Keywords: homeostasis, allostasis, rheostasis, set point, negative feedback, control theory, cephalic responses, glucose regulation, thermal regulation, sodium appetite, calcium appetite
Introduction
Curt Richter was a pioneer and a giant in the field of behavioral neuroendocrinology. His contributions to several specific areas of research are topics of other articles in this series. Here we focus upon Richter’s contributions and views on homeostasis and consider how the field has evolved since his investigations and prescient conclusions. Although neither of us had the pleasure of meeting Richter, our own interests and hypotheses can be seen to derive from the seeds sowed over his illustrious career.
Claude Bernard and Walter Cannon
In the 19th century, Claude Bernard articulated the necessity of maintaining an environment within the body that allows vital processes to proceed independently of physicochemical perturbations in the environment in which an organism resides. In his schema, the various organ systems were viewed as having the capacity to counter harmful deviations in one or another variable in the service of guaranteeing a relatively stable and benevolent milieu intérieur. With survival of the whole organism as the overarching goal, individual components functioned to sustain the temperature, osmolality and/or pH of the fluid bathing each tissue. At another level of regulation, organ systems were coordinated to ensure constant and adequate supplies of essential commodities such as glucose and oxygen to vital tissues, and the assurance of adequate blood flow and pressure.
As interpreted by Langley (Langley 1973), Bernard (Bernard 1870) viewed the stability of the internal environment as a “necessary condition for a free and independent life” (p. 146) and that external variations are at every instant compensated and brought into balance. Consistent with Langley’s interpretation, Yates (Yates 1996) contends that although Bernard’s use of the word “fixité” is frequently translated as “constancy,” it would be better translated as “stability.” It may therefore be that the concept of an unchanging or fixed set point inherent in many conceptions of homeostasis is based on a misunderstanding of Bernard’s original intent, for to quote his countryman, “The living body is stable. It must be in order not to be destroyed, dissolved or disintegrated by the colossal forces, often adverse, which surround it. By an apparent contradiction, it maintains its stability only if it is excitable and capable of modifying itself according to external stimuli, and adjusting its response to the stimulation. In a sense it is stable because it is modifiable. The slight instability is the necessary condition for the true stability of the organism.” ((Richet 1890), translation quoted in (Cannon 1929)). As Mrosovsky (Mrosovsky 1990) notes, Cannon himself recognized this, selecting the word homeostasis rather than homostasis to indicate similarity, with some variability, rather than sameness (Cannon 1929). Cannon later (Cannon 1932) emphasized that rather than emphasizing something set and immobile, homeostasis reflects a condition which may vary but which is relatively stable (see a more thorough discussion in Carpenter (Carpenter 2004)).
An important insight of Bernard was his emphasis upon the centrality of the circulatory system, stating that “… in the living being, the tissues are, in reality, removed from direct external influences and protected by a true internal environment mostly constituted by fluids circulating in the body” ((Bernard 1857a) (p. 87) and that “… blood is therefore a true environment into which all the tissues reject their decomposition products and where they find, for the achievement of their functions, invariable conditions of temperature, humidity, and oxygenation, as well as nitrogenous matter, carbohydrates, and salts without which organs cannot thrive” (Bernard 1857b) (p. 90). Other particularly noteworthy contributions of Bernard include his popularization of the widely held view that all of the organs in a living body act in reciprocal harmony such that separation of any individual part disturbs the entire organism, and the recognition that the brain is an important player in these processes. These major insights laid the foundation for generations of scholars and healthcare providers to interpret and expand the myriad implications. Indeed, monitoring one’s vital signs is generally the first order of business in any professional assessment of one’s health status.
Bernard’s views percolated for several decades but did not achieve widespread acclaim or acceptance within the United States until incorporated into the writings of Walter Cannon in the early 20th century. It was an exciting time for physiology, with the tenet of a fundamental protective function of the nervous system becoming predominant. Sherrington’s work had revealed the overall integrative action of the nervous system including identifying and classifying spinal pathways interconnecting the body and the brain. He championed study of the nature of spinal reflexes with their immediate and automatic ability to respond to sudden inputs. These reflexes were viewed as enabling organisms to withdraw from painful stimuli without the necessity or temporal delay of having to consciously ponder how best to respond in acute and threatening situations. Cannon (Cannon 1932) framed these and many other phenomena into a larger picture that he summarized in his book, The Wisdom of the Body. Coining the term homeostasis, he viewed organisms as engaging in continuously ongoing reflexive processes that ensure an adequate internal environment for every vital activity.
Curt Richter and Homeostasis
Curt Richter embarked on his scientific career at the time that Cannon was popularizing the principles of homeostasis. Having been in John Watson’s lab as a graduate student, Richter became a skilled and creative student of behavior, and over his career there were few areas of behavior that escaped his scrutiny. That is not to say that Richter embraced Watson’s teachings. As Richter’s biographer Schulkin (Schulkin 2005) pointed out, Watson had rejected the concept of instinct whereas Richter believed that instinctive responses represent the basis of the regulatory behaviors animals use to maintain homeostasis. Analogously, whereas Watson co-opted Pavlov’s conditioning paradigm to explain much of behavior, Richter never considered conditioning or learning to be a major force, especially in his consideration of homeostasis (see (Moran & Schulkin 2000)).
On the other hand, Richter did recognize that behavior was a potent and often necessary strategy for controlling vital variables, and his research in this arena moved him far beyond obvious examples such as the drinking and feeding that occur in water-deprived and food-deprived animals, respectively. With regard to the study of homeostasis, he is perhaps best known for uncovering the rich behavioral repertoire animals recruit in the defense of regulated variables such as sodium and calcium levels, body temperature and blood glucose. As Richter himself stated, a preferred strategy he employed was first to eliminate the otherwise predominant physiological regulators of one or another variable so as to better reveal the existence of behavioral regulators of the internal environment (Richter 1942–1943). This often took the form of removing a regulatory hormone-secreting gland. A complementary and quite compelling strategy Richter used was either to administer the hormone whose activity was lost when an endocrine gland was removed or else to implant excised endocrine tissue into animals whose behavior has been altered in order to assess the degree of return of function. For example, thyroidectomized rats cannot generate adequate body heat and consequently build large nests to keep warm, but when they are administered thyroid hormone their temperature increases and they stop building nests altogether (Richter 1937).
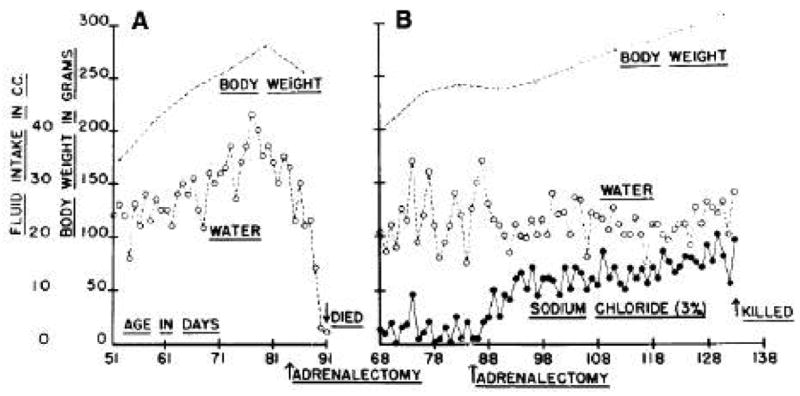
Perhaps the best known example of Richter’s approach to homeostasis is his finding that rats whose adrenal glands are removed, thereby compromising the control of sodium metabolism, compensate for their unrestrained sodium loss by consuming hypertonic saline when given the opportunity and thereby prevent an otherwise certain death (Richter 1936a; Richter 1939). Because intact animals consume only small amounts of such concentrated saline, the homeostatic perturbation induced by adrenalectomy was seen as unmasking an otherwise latent protective homeostatic behavior. In parallel experiments, removal of the parathyroids resulted in the animals compensating behaviorally by consuming more calcium (Richter & Birmingham 1941; Richter & Eckert 1937; Richter & Eckert 1939), and this calcium appetite was reduced to control levels by implanting parathyroid tissue into the anterior chamber of the eye of parathyroidectomized rats (Figure 1), implying that the parathyroids secrete an endocrine factor that helps to maintain plasma calcium (Richter & Eckert 1937).
Figure 1.
Consumption of a 2.4% solution of calcium lactate increases after removal of the parathyroid glands from a rat; and it subsequently decreases following the successful grafting of parathyroid tissue into the anterior chamber of the eye in the same rat (from (Richter 1942–1943)).
Plasma calcium is often reciprocally related to plasma phosphorus, and one manifestation of this is that whereas parathryoidectomized rats have increased calcium appetite, they have reduced phosphorus appetite (Richter & Eckert 1939), providing an elegant example of the inter-relatedness of different homeostatic processes, in this case reciprocally regulating calcium and phosphorus. The principle of dual control over more than one behavior can be also observed when the adrenals are removed. Adrenalectomized rats, besides having increased salt appetite, are extremely hypoactive, with running wheel activity decreasing dramatically. Implanting adrenal tissue, administering adrenal cortical extracts either by injection or orally, or else providing access to sufficient dietary salt, were all found to reduce salt appetite and to restore running wheel activity (Richter 1936b).
Analogously, removal of the pancreas resulted in animals exhibiting polydipsia and hyperphagia (Richter & Schmidt 1939). In this latter instance, the increased drinking compensated for water loss that accompanied sugar excretion in the urine, and the increased eating was to obtain needed amounts of nutrients that can be utilized (see (Richter 1942–1943)). He also found that when given a choice of macronutrients, a different behavioral strategy emerged. Pancreatectomized rats would consume large amounts of fats and proteins and almost no carbohydrates, thus normalizing their blood sugar as well as their caloric intake (Richter & Schmidt 1941). Thus, faced with a life-threatening loss of calories and offered only a single, high-carbohydrate source of food, pancreatectomized rats compromised by eating sufficient calories to live, but at the expense of chronic severe hyperglycemia; and such compromise was unnecessary when a more specific behavioral option was possible. Richter was tireless in documenting and understanding the scope and richness of the behavioral armamentarium that could be used in the service of homeostasis.
Richter’s demonstrations of behavioral maintenance of homeostasis are highly important, and he well-understood how they built upon and extended the sphere of the earlier work of Bernard and Cannon. Like Bernard before him, he emphasized the importance of responses that maintain blood variables at optimal levels. Richter also understood his unique contribution, stating that “both Bernard and Cannon concerned themselves almost entirely with the physiological and chemical regulators of the internal environment,” ((Richter 1942–1943), p. 64), and he believed that his own contribution to the concept of homeostasis was to demonstrate that specific behaviors, which he termed “total organism regulators,” also contribute to maintenance of a constant internal environment (Richter 1942–1943). He came to the conclusion that behavior in any form is aimed at maintaining the internal state, stating “As a result of these findings, and particularly those of Cannon, I had come to believe that almost all aspects of behavior can be fully understood in terms of the concept of homeostasis.” (Richter 1967) (p. 8).
Negative Feedback
Richter’s views on homeostasis reflected prevailing thought on physiology. While he inserted behavior firmly into regulation, his views, like those of Cannon and his other contemporaries, considered only one aspect of the regulatory process; i.e., the ability of an organism to make responses that correct a perturbation to a homeostatic variable. Hence, such responses, whether physiological, chemical or behavioral, were also considered as reflexive or automatic, and they fit into what is now known as a negative feedback model.
The concept of homeostasis has gone through a number of important adjustments over the years. In originally considering the term homeostasis, Cannon believed that under ordinary situations, the variations from the mean position of a regulated variable do not reach dangerous extremes which impair the function of the cells or threaten the existence of the organism, and that before these extremes are reached, responses are automatically called into service which act to bring back toward the mean position the state which has been disturbed (Cannon 1932). A more current conceptualization of this process would involve thresholds that activate effector mechanisms that oppose further deviation of the regulated variable (Ramsay & Woods 1997).
With the application of engineering’s control theory to biological regulation (Weiner 1948), homeostasis became operationally defined as a series of inter-related negative feedback loops. This perspective enjoyed immense popularity and in a review of homeostatic principles, Mrosovsky (1990, p. 4) declared that control theory is firmly entrenched in the field. In an excellent compilation of historical articles on homeostasis, Langley (Langley 1973) defined homeostasis as “… self-regulating negative feedback systems which serve to maintain the constancy of the internal environment of living organisms” (p. 10), and a leading textbook of physiology defined homeostasis to mean “maintenance of static, or constant, conditions in the internal environment” (Guyton 1982). The engineering concept of a negative feedback loop was perfectly consistent with Richter’s writings. It employs a reference signal (set-point) and requires a measured deviation from the reference signal in order to generate the error signal that activates corrective responses.
The face validity and conceptual simplicity of the negative-feedback control model was appealing as the physical sciences became more and more integrated with the life sciences in the latter decades of the twentieth century, but not without a cost. As pointed out by Carpenter, this “particular form of homeostasis, direct negative feedback, began to be taught in schools; it was taught as if it were the only kind of biological control system, and this folk-belief eventually invaded tertiary education” ((Carpenter 2004), p. 187). The legacy of embracing negative feedback as the primary mode of homeostatic control remains apparent in many discussions. Koob and LeMoal (Koob & Le Moal 2001) equate homeostasis with negative feedback, and Berridge (Berridge 2004) contends that anticipatory responses that function to defend a regulated variable from becoming deviated cannot be considered homeostatic because “… there has been no physiological deficit and, hence, no error detection” (p. 182). Carpenter considers it a “travesty” that “for many, homeostasis is synonymous with direct negative feedback with a fixed ‘set point’” ((Carpenter 2004), p. 180). The concept of homeostasis also interacts intimately with the use of the term regulation as it applies to physiological variables. Cabanac (Cabanac 1997) states that the “main property of a regulatory system is that a deviation of the regulated variable triggers a correcting response, which opposes the deviation” (p. 27). It is this corrective response that distinguishes regulation from a passive steady-state (Cabanac 2006).
In contrast to these views and consistent with many contemporary scholars, we feel that it is overly restrictive to limit homeostasis to achieving regulatory control solely via the action of negative feedback. As a concept, homeostasis encompasses the idea that the internal milieu operates optimally when a given bodily variable is within a specified range and that physiological strategies exist to maintain and defend the regulated variable within that range.
Undoubtedly, negative feedback plays an important role in homeostasis as regulated variables are defended from unanticipated disturbances and emergency situations. That said, we believe that equating negative feedback with homeostasis has also unduly influenced how data are interpreted in physiological regulation. For example, if a decrease in blood glucose precedes a bout of eating, the decline could be viewed as a deficit or error signal that elicits feeding as a corrective response in defense of blood glucose (Campfield & Smith 2003; Mayer 1955). This makes perfect sense from a negative feedback perspective. Conversely, if the drop in blood glucose is seen as an anticipatory preparatory response to an upcoming influx of glucose in an impending meal, the interpretation is much different. In this light, eliciting a neurally induced slight decrease of glucose can be viewed as an appropriate preparation to circumvent an otherwise excessive increase of glucose. We (Ramsay et al. 1996; Ramsay & Woods 1997; Strubbe & Woods 2004; Woods 1991; Woods & Ramsay 2000; Woods & Strubbe 1994), as well as other contemporary writers (Dworkin 1993; Schulkin 2003), believe that this better captures the essence of the concept of regulation originally espoused by Bernard.
A stable set-point or reference signal as used in engineering is commonly applied to negative feedback models of physiological regulation. However, in physiology, a set-point is a hypothetical construct that is inferred from measures that assess whether an animal defends a given value of one or another variable using behavioral and/or physiological responses. Furthermore, in actuality these set-points are often adjustable and can be altered by myriad naturalistic factors (e.g. circadian and seasonal changes, hormonal status, nutrition, stress). Viewing homeostasis in terms of a negative feedback loop may lead the observer to interpret a change in a regulated variable as a possible deficit or error signal rather than as an adjustment of the defended level. A similar point is made by Mrosovsky (Mrosovsky 1990) who contends that changes in regulated levels are often mistakenly “… regarded as failures of homeostasis” (p. v).
Anticipatory Responses
There is a rich literature documenting anticipatory responses individuals make in order to preclude changes in regulated variables. Perhaps the best known are meal-anticipatory cephalic responses (Katschinski 2000; Powley 1977; Teff 2000). Initiated by the brain, these responses such as the secretion of insulin, ghrelin and other hormones prior to meal initiation, enable the consumption of a large nutrient load with minimal homeostatic consequences (Drazen et al. 2006; Woods 1991; Woods & Strubbe 1994). It has long been recognized that these anticipatory responses are necessary to circumvent glucose intolerance during meals (Berthoud et al. 1981; Louis-Sylvestre 1978; Nicolaidis 1977; Proietto et al. 1987) and the argument has been made that dysfunction of these responses predisposes to diabetes mellitus and/or obesity (Storlien & Bruce 1989).
This concept is simple and it is fitting to illustrate it using glucose as an example; indeed Langley (Langley 1965) asserts that Bernard’s and Cannon’s interest in the physiology of glucose metabolism was at least partially responsible for the birth of the concept of homeostasis, and Richter himself often used the maintenance of glucose levels in the blood as an example of complex regulatory processes. Blood glucose is normally stabilized within strict limits, even during exercise. The predominant challenge to glucose homeostasis in a non-starved animal is meal intake, during which marked rises in glucose can and often do occur (Grossman 1986; Langhans 1996). To the extent that perturbations can be reliably predicted, an individual can learn to make responses that minimize the perturbations, thus helping maintain homeostasis. Cephalic insulin and other responses mediate this.
In principle, any homeostatically regulated variable is subject to a similar anticipatory process. Body temperature is such a variable, and it is the case that when presented with cues indicating that they are about to be placed into a cold environment, animals raise their temperature in anticipation (Hjeresen et al. 1986; Mansfield et al. 1983). Analogously, animals presented with cues indicating that they are about to receive a pharmacological agent that lowers body temperature also learn to raise their temperature in anticipation (Lê et al. 1979; Mansfield et al. 1983; Mansfield & Cunningham 1980). In both instances, the preemptive rise of temperature mitigates the severity of the impending hypothermia. We believe that these are examples of homeostasis that go well beyond the negative feedback model championed by many.
Regulation vs. Control: Body Adiposity vs. Food Intake
While it is generally accepted that corrective responses triggered by a deviation are indicative of a regulated system (Cabanac 2006), accumulating evidence indicates that regulated variables commonly achieve this stability in the absence of any obvious error signal or deviation. This position was well-stated by Somjen (Somjen 1992), “The conclusion seems almost inescapable: the central nervous system (CNS) anticipates present and future needs on the basis of past experience. By having successfully corrected errors, the CNS learns how to prevent them” (p. 184). As early as the mid-20th century, Kennedy described body adiposity as an example of a regulated variable (Kennedy 1953). He recognized that energy intake is normally well-matched to energy expenditure such that body adiposity is able to be maintained within a fraction of a percent over long intervals. However, meals are typically consumed in free-feeding animals in the absence of any apparent or identifiable deficit (Woods 1991; Woods 2002).
Many writers emphasize the importance of differentiating control versus regulation. Cabanac (Cabanac 2006)discusses the issue in detail, and citing Brobeck (Brobeck 1965), he notes that the distinction has long been recognized as being critical in any discussion of homeostasis. Control refers to the ability to activate effector responses in order to stabilize a regulated variable, either by reversing a perturbation or anticipating and thus helping circumvent or minimize a perturbation. With regard to food intake and body weight, feeding would be considered the controlled effector response, one that is not regulated but helps to stabilize the regulated variable of body adiposity. In this light, although meals are obviously necessary for the defense of body weight in the long run, it has been argued (e.g., (Bolles 1980)) that feeding per se is not regulated because it is not being driven by deficits. In contrast, body adiposity can be considered to be regulated because perturbations in body fat elicit corrective effector responses to restore adiposity (Ramsay et al. 1996; Seeley et al. 1997; Strubbe & van Dijk 2002; Strubbe & Woods 2004; Woods et al. 1974; Woods & Strubbe 1994).
Collier emphasized the distinction between the regulatory aspects of food intake and body weight in a different way. He found that animals are in fact quite flexible with regard to specifics of the parameters of food taking, readily adjusting their daily pattern of meal size and meal number to accommodate environmental constraints, yet always attaining a schedule that allows maintenance (i.e., regulation) of body weight (Collier 1986; Collier et al. 1986; Collier et al. 1999). Collier viewed animals as economists that constantly balance the procurement of food with numerous other behavioral activities, yet still defending body weight. This highlights the necessity of knowing precisely what is being regulated when animals engage in one or another behavioral response as we have discussed elsewhere (Ramsay & Woods 1997; Woods 1991; Woods & Ramsay 2000).
Richter viewed homeostatic responses as reflexive behaviors elicited by perturbations of regulated variables. Loss of plasma sodium elicits salt appetite; loss of calcium elicits calcium appetite; loss of carbohydrate calories through the urine elicits food intake and especially carbohydrate intake; loss of body temperature elicits nest building; and so on. In his schema, these behaviors were conceptualized as respondents made in the service of regulating and hence maintaining the integrity of Bernard’s internal milieu.
It is curious that Richter rarely discussed the operant aspects of the same behaviors that he believed were instinctively recruited to regulate homeostatic variables. To use food intake as an example, while it is certainly the case that eating can be elicited by experimentally lowering blood glucose or its utilization (Grossman 1986; Lotter & Woods 1977; MacKay et al. 1940; Mayer 1955; Mayer & Thomas 1967; Smith & Epstein 1969), or even compromising the ability of an animal to glean calories from oxidizing fats (Langhans 1996; Scharrer & Langhans 1986), as discussed above most instances of eating or meal taking occur in the absence of such homeostatic perturbations, and these examples of elicited eating are generally viewed as metabolic emergencies that bear little resemblance to most instances of food taking (Grossman 1986; Langhans 1996; Woods 1991).
The rich literature on the ease of manipulating food-procuring responses via operant procedures argues that whereas behaviors certainly can be recruited in the service of homeostasis as espoused by Richter, the same behaviors are easily modifiable to meet the overall needs of the organism. Indeed, the very behavior touted as the poster child of innate homeostatic regulation, increased salt appetite when plasma sodium is decreased, is superimposed upon an already high basal intake (i.e., a high operant level) of mild sodium solutions relative to pure water, as Richter himself noted (Richter 1939). Further, learning has been found to interact with the initial bout of salt intake in response to an experimentally induced lowering of plasma sodium (Krieckhaus & Wolf 1968; Weisinger et al. 1970). The point is that Richter perhaps over-zealously viewed behaviors that participate in the control of homeostatic variables as being solely examples of innate regulation.
To summarize, we believe that a more appropriate position is to describe the role of feeding within an expanded view of homeostatic regulation that is not limited to a negative feedback model. In this situation, feeding is not a regulated variable, but a controlled effector/response mechanism that, among other functions, stabilizes the regulated variable of adiposity. In the expanded view of homeostasis, feeding can occur without needing to be elicited by an error signal and in some instances can be seen as creating error signals on its own (Woods 1991; Woods & Strubbe 1994). Nonetheless, eating is a behavior that functions to stabilize adiposity over long intervals.
Expanding the Homeostatic Umbrella
The issues discussed above raise several fundamental questions about homeostasis and regulation. When corrective or compensatory responses are activated in anticipation of an impending disturbance, thus preventing the deviation of a regulated variable, should this be considered homeostatic regulation even though it does not fit within a negative feedback framework? Our view is that it should. Similarly, if a prolonged homeostatic challenge initially perturbs a regulated variable and thus creates an error signal that in turn elicits a corrective response, must that response cease once the error signal is sufficiently attenuated only to be re-elicited as the error signal recurs due to the ongoing presence of the disturbance? Such a system would result in oscillation of the regulated variable. To prevent these oscillations, one possibility is that the corrective response could persist after correction of the triggering deficit. Such a persistent response in the absence of a continuing perturbation presents a different challenge to negative feedback conceptions of homeostasis. Hammel (Hammel 1990), a pioneer in homeostatic research on thermoregulation, considered this issue and believed that the continued presence of a corrective response after the regulated variable had recovered from the perturbation was “inexplicable within the framework of Cannon’s concept of homeostasis and negative feedback” (p. 175). Thus, when confronted with empirical data demonstrating this phenomenon, Hammel (Hammel 1989; Hammel 1990) proposed that the negative feedback view of homeostasis should be augmented by the addition of a positive feedback loop or autofacilitation to account for a corrective response that persists in the continued presence of a homeostatic challenge even though the error signal is no longer present. The point is that contemporary versions of control theory offer many sophisticated approaches to regulation beyond those provided by simple negative feedback alone.
Some endogenous signals are early warning predictors of impending perturbations to regulated variables, and these signals can activate corrective responses directly. An instructive example comes from thermoregulation where a drop in skin temperature signals an impending reduction in core temperature unless a compensatory response (e.g., increased metabolic rate; shivering) is initiated to circumvent it. These anticipatory responses can sometimes result in overcompensation (Brown & Brengelmann 1970; Gordon 2005). If core temperature is the actual regulated variable, preemptive thermoregulatory responses are activated without the need of an error signal from the perturbed regulated variable. As stated previously, a narrow view of what constitutes homeostasis would not consider these responses homeostatic because they are not triggered by an error signal. However, by broadening the sensory input used by the regulatory system to include shell temperature as well as core temperature, one could consider skin temperature change as providing a signal to the regulatory system that can be used to help stabilize the regulated variable and thus accommodate to objectives of homeostasis without the need for an error signal in the regulated variable per se.
An alternate approach suggested by Mrosovsky (Mrosovsky 1990) is that cooling of the body’s shell causes a rheostatic increase in the defended level of the variable (i.e., the ‘set-point’) which generates an error signal due to the mis-match between the regulated variable and the new set-point such that effector mechanisms are activated that will defend core temperature against hypothermia. However, an error signal does not appear to be necessary to activate corrective responses. Learned thermoregulatory behavioral responses made in anticipation of thermal challenges (e.g., putting on one’s winter coat before going outside in the snow) occur in the absence of a thermal error signal and serve to prevent an impending perturbation of the regulated variable. From our perspective, homeostasis should include these responses that do not require elicitation by an error signal.
Set-Points
As discussed above, another area of confusion regarding the concept of homeostasis relates to the understanding of set-points. In physiological regulation, the term is used metaphorically to indicate that a system operates “as if” there was an engineering type of set-point or reference signal, and this analogy can be misleading (Berridge 2004; Gordon et al. 2001). Physiologically plausible models that give regulated systems the appearance of a set-point propose that there are thresholds for activating homeostatic responses and that they exist within hierarchically arranged, and possibly interacting, physiological control systems (Bligh 1998; Bligh 2006; Brengelmann & Savage 1997; Kanosue et al. 1997; Romanovsky 2004; Satinoff 1978). When intact animals encounter naturally occurring homeostatic challenges, it is typical to find that physiological responses and motivated behaviors are activated in a coordinated manner to defend a regulated variable (Cabanac 2006). However, the specific value/range that is defended at a given moment can vary according to myriad factors (Hammel 1970; Satinoff 1996). In his book on rheostasis, Mrosovsky (Mrosovsky 1990) describes this phenomenon in detail as “.. a convenient term for designating changes in regulated levels” (p. v). The point is that an interpretational complexity arises in studies on homeostasis because an observed change in a regulated variable can result from a forced deviation away from its defended value by an externally arising disturbance or else from a rheostatic adjustment of the value to a new defended level.
Gordon (Gordon 1983; Gordon 2005) refers to changes in a regulated variable that reflect a deviation away from a defended level because of an external disturbance as being “forced” while changes in a variable that are due to a rheostatic adjustment of the defended value are termed “regulated.” Gordon suggests that “forced” and “regulated” changes in a measure can be distinguished by inference because behavioral and physiological homeostatic mechanisms will be activated to oppose or compensate for a “forced” change whereas effector mechanisms will facilitate a change that is “regulated.”
Implications of Homeostatic Principles
The practical aspect of adjustments of regulated variables is that they may compromise health. Features of modern society may inadvertently challenge our regulatory systems in ways that do not lead to improved viability or quality of life. For example, certain aspects of the modern environment are considered to have raised the defended level of adiposity, contributing to the current ‘epidemic’ of obesity (Hill et al. 2003; Schwartz et al. 2003). Chronically elevated adiposity in turn is often associated with maintenance and defense of elevated blood glucose (i.e., type-2 diabetes mellitus) and elevated blood pressure, both of which can be life-threatening (Landsberg 2006). Understanding why and how so-called set-points or defended levels of medically important variables change, and how the regulation of one variable interacts with the regulation of others when the homeostatic objectives of each system are in conflict (for greater discussion see Mrosovsky, 1990), are daunting but critical challenges facing modern science.
There is thus a tremendous potential to improve health through a better understanding of homeostasis. Evolution has created complex robust regulatory mechanisms that can be fragile when confronted with ecologically novel challenges that are an increasingly common byproduct of modern society (Csete & Doyle 2002). Indeed, the case might be made that many of our major health problems like drug addiction and obesity may be the result of fragilities in these regulatory mechanisms. Advances in conceptualizing the neural organization of homeostatic systems have important clinical implications (Craig 2002). In 1990, Mrosovsky (Mrosovsky 1990) p. 147) was optimistic about the therapeutic potential of working in concert with, rather than against, our homeostatic systems, and the application of these ideas is coming closer to fruition in some areas (Gordon 2005; Gordon et al. 2001). In 1986, Moore-Ede (Moore-Ede 1986) stated that “No single thought has been more central to the development of physiological thought than the principle of homeostasis” (p. R737), and more recently, Carpenter (Carpenter 2004) stated “… ‘homeostasis’ has become a kind of respectable euphemism for the P-word” (physiology) (p. 180).
Yet, in many instances restricted conceptualizations of homeostasis have lacked the necessary explanatory power to account for the variety of homeostatic-relevant phenomena. As a consequence, many contemporary investigators have modified the concept to make it more inclusive of the data, providing terms such as predictive homeostasis, reactive homeostasis, homeorheusis, homeorhesis, homeokinetics, rheostasis, homeodynamic regulation, heterodynamic regulation, heterostasis, allostasis, and allodynamic regulation. One of the newer derivations of homeostatic concepts is allostasis (Schulkin 2003; Sterling 2004), which Schulkin ((Schulkin 2004), p. 7) describes as “the process by which an organism achieves internal viability through a bodily change of state (especially central motive state).” However, it should be noted that at present there is no generally agreed upon definition for allostasis (Berridge 2004; Power 2004). Not convinced of the value of allostasis, Carpenter describes it as a “… much-promoted but vaguely characterized cluster of concepts that represent nothing that has not always been part of the ordinary conceptual basis of homeostatic control.” ((Carpenter 2004), p. 180). In our view, it is too early to assess whether the concept of allostasis will prove of value in understanding regulatory physiology.
Hammel (Hammel 1988) observed that, “Research in regulatory biology, wherein the nervous system of the vertebrate body is engaged, has diminished markedly in recent years, although the concepts enunciated by Claude Bernard, Walter Cannon, their peers, and their successors require major revision” (p. 835). Some might argue that the paucity of research in this area relates to Carpenter’s ((Carpenter 2004), p. 180) contention that “… a generation of molecularly oriented biologists have grown up unfamiliar with the ideas of these pioneers, or with even the rudiments of control theory. Among today’s physiologists there is generally more pious lip service paid to homeostasis than genuine understanding of control theory.” However, our view is different in that sophisticated applications of control theory are being used to understand cellular regulatory systems (e.g., (Hucka et al. 2003; Kurata et al. 2006)). The application of these principles at the higher level of whole animal physiological regulation has diminished because it has not been a goal of the growing number of scientists working at a more molecular level. Schultz (Schultz 1996) makes the point that understanding the emergent properties of integrated functioning of physiological systems will not be accomplished through the reductionistic investigation of its component parts. He states “…. the study of integrated systems cannot be supplanted, or rendered unnecessary or superfluous, by advances at lower levels of organization. Physiology is not a derivative science.” ((Schultz 1996), p. 246) Scientists with an integrative research agenda will be needed to investigate higher level questions in biological regulation. Indeed, there is a growing recognition of the need to revitalize and train individuals in a “systems” or “integrative” approach to biology and physiology (Cowley 2005), as well as in experimental psychology (Cabanac 2006).
We believe that there needs to be an increased research focus on advancing our knowledge of CNS-mediated regulatory principles particularly within the broader context of adaptive control (also see Carpenter (Carpenter 2004)). Understanding how these highly complex homeostatic regulatory pathways operate will yield great health dividends as many of major health problems (e.g., obesity, drug addiction) may be amenable to therapeutic intervention at the level of the homeostatic control systems. Research that encompasses a broad view of homeostasis that goes beyond simple feedback circuit models will be required to make progress in this area.
Looking Back
An interesting retrospective exercise is to ponder whether Curt Richter would have embraced a broader perspective on the principles of homeostasis. Certainly there were no predominant theoretical models espousing the kind of deviations from strict negative feedback that we discuss above, and dogma during much of Richter’s career favored a strict interpretation. Further, his career-long reluctance to consider learning as a strategy to facilitate regulation argues against certain directions his thinking might have taken. Nonetheless, all indications are that Richter had an unusually creative and open mind, and that had nuances of control theory been championed by others, he would have been at the forefront in designing clever behavioral paradigms to uncover even further expansions of the homeostatic envelope.
Acknowledgments
We owe an enormous debt to Gerry Smith, both for inviting us to contribute to this retrospective consideration of Curt Richter as well as for the infusion of his encyclopedic knowledge and sage editorial advice into earlier versions of the manuscript. The writing was supported in part by NIH awards DK17844, DK067550 and DA016047.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bernard C. Lectures on the physiological properties and the pathological alterations of the liquids of the organism: First lecture. In: Langley LL, editor. Homeostasis: Origins of the Concept, 1973. Stroudsberg, PA: Dowden, Hutchinson & Ross, Inc; 1857a. pp. 84–88. [Google Scholar]
- Bernard C. Lectures on the physiological properties and the pathological alterations of the liquids of the organism: Third lecture. In: Langley LL, editor. Homeostasis: Origins of the Concept. Stroudsberg, PA: Dowden, Hutchinson & Ross, Inc; 1857b. pp. 89–100. [Google Scholar]
- Bernard C. Lessons on the phenomena of life common to animals and vegetables. Second lecture, the three forms of life. In: Langley LL, editor. Homeostasis: Origins of the Concept, 1973. Stroudsberg, PA: Dowden, Hutchinson & Ross, Inc; 1870. pp. 129–151. [Google Scholar]
- Berridge KC. Motivation concepts in behavioral neuroscience. Physiology and Behavior. 2004;81:179–209. doi: 10.1016/j.physbeh.2004.02.004. [DOI] [PubMed] [Google Scholar]
- Berthoud HR, Bereiter DA, Trimble ER, Siegel EG, Jeanrenaud B. Cephalic phase, reflex insulin secretion. Diabetologia. 1981;20(Suppl):393–401. doi: 10.1007/BF00254508. [DOI] [PubMed] [Google Scholar]
- Bligh J. Mammalian homeothermy: An integrative thesis. Journal of Thermal Biology. 1998;23:143–258. [Google Scholar]
- Bligh J. A theoretical consideration of the means whereby the mammalian core temperature is defended at a null zone. Journal of Applied Physiology. 2006;100:1332–1337. doi: 10.1152/japplphysiol.01068.2005. [DOI] [PubMed] [Google Scholar]
- Bolles RC. Some functionalistic thoughts about regulation. In: Toates FM, Halliday TW, editors. Analysis of Motivational Processes. New York: Academic Press; 1980. pp. 63–75. [Google Scholar]
- Brengelmann GL, Savage MV. Temperature regulation in the neutral zone. Annals of the New York Academy of Sciences. 1997;813:39–50. doi: 10.1111/j.1749-6632.1997.tb51670.x. [DOI] [PubMed] [Google Scholar]
- Brobeck JR. Exchange, control, and regulation. In: Yamamoto WS, Brobeck JR, editors. Physiological Controls and Regulations. Philadelphia: Saunders; 1965. pp. 1–14. [Google Scholar]
- Brown AC, Brengelmann GE. The interaction of peripheral and central input in the temperature regulation system. In: Hardy JD, Gagge AP, Stolwijk JAJ, editors. Physiological and Behavioral Temperature Regulation. Springfield: Charles C. Thomas; 1970. pp. 684–702. [Google Scholar]
- Cabanac M. Regulation and modulation in biology. A reexamination of temperature regulation. Annals of the New York Academy of Sciences. 1997;813:21–31. doi: 10.1111/j.1749-6632.1997.tb51668.x. [DOI] [PubMed] [Google Scholar]
- Cabanac M. Adjustable set point: to honor Harold T. Hammel. Journal of Applied Physiology. 2006;100:1338–1346. doi: 10.1152/japplphysiol.01021.2005. [DOI] [PubMed] [Google Scholar]
- Campfield LA, Smith FJ. Blood glucose dynamics and control of meal initiation: a pattern detection and recognition theory. Physiological Reviews. 2003;83:25–58. doi: 10.1152/physrev.00019.2002. [DOI] [PubMed] [Google Scholar]
- Cannon WB. Organization for physiological homeostasis. Physiological Reviews. 1929;9:399–431. [Google Scholar]
- Cannon WB. The Wisdom of the Body. New York: Norton; 1932. [Google Scholar]
- Carpenter RH. Homeostasis: a plea for a unified approach. Advances in Physiology Education. 2004;28:180–187. doi: 10.1152/advan.00012.2004. [DOI] [PubMed] [Google Scholar]
- Collier G. The dialogue between the house economist and the resident physiologist. Nutrition and Behavior. 1986;3:9–26. [Google Scholar]
- Collier GH, Johnson DF, Hill WL, Kaufman LW. The economics of the law of effect. Journal of the Experimental Analysis of Behavior. 1986;48:113–136. doi: 10.1901/jeab.1986.46-113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collier GH, Johnson DF, Mitchell C. The relation between meal size and the time between meals: Effects of cage complexity and food cost. Physiology and Behavior. 1999;67:339–346. doi: 10.1016/s0031-9384(99)00086-4. [DOI] [PubMed] [Google Scholar]
- Cowley A. Global manpower needs for integrative systems physiologists. Physiologist. 2005;48:1, 3–8. [PubMed] [Google Scholar]
- Craig AD. How do you feel? Interoception: the sense of the physiological condition of the body. Nature Reviews Neuroscience. 2002;3:655–666. doi: 10.1038/nrn894. [DOI] [PubMed] [Google Scholar]
- Csete ME, Doyle JC. Reverse engineering of biological complexity. Science. 2002;295:1664–1669. doi: 10.1126/science.1069981. [DOI] [PubMed] [Google Scholar]
- Drazen DL, Vahl TP, D’Alessio DA, Seeley RJ, Woods SC. Effects of a fixed meal pattern on ghrelin secretion: evidence for a learned response independent of nutrient status. Endocrinology. 2006;147:23–30. doi: 10.1210/en.2005-0973. [DOI] [PubMed] [Google Scholar]
- Dworkin BR. Learning and Physiological Regulation. Chicago: University of Chicago Press; 1993. [Google Scholar]
- Gordon CJ. A review of terms for regulated vs. forced, neurochemical-induced changes in body temperature. Life Sciences. 1983;32:1285–1295. doi: 10.1016/0024-3205(83)90802-0. [DOI] [PubMed] [Google Scholar]
- Gordon CJ. Temperature and Toxicology: An Integrative, Comparative and Environmental Approach. Boca Raton: CRC Press; 2005. [Google Scholar]
- Gordon CJ, Kimm-Brinson KL, Padnos B, Ramsdell JS. Toxicon. 2001;39:1367–1374. doi: 10.1016/s0041-0101(01)00092-7. [DOI] [PubMed] [Google Scholar]
- Grossman SP. The role of glucose, insulin and glucagon in the regulation of food intake and body weight. Neuroscience and Biobehavioral Reviews. 1986;10:295–315. doi: 10.1016/0149-7634(86)90015-1. [DOI] [PubMed] [Google Scholar]
- Guyton AC. Human Physiology and Mechanisms of Disease. Philadelphia: W. B. Saunders Co; 1982. [Google Scholar]
- Hammel HT. Concept of the adjustable set temperature. In: Hardy JD, Gagge AP, Stolwijk JAJ, editors. Physiological and Behavioral Temperature Regulation. Springfield: Charles C. Thomas; 1970. pp. 676–683. [Google Scholar]
- Hammel HT. Anesthetics and body temperature regulation. Anesthesiology. 1988;68:833–5. [PubMed] [Google Scholar]
- Hammel HT. Homeostasis: Embracing negative feedback enhanced and sustained by positive feedback. In: Paganelli CV, Fahri LE, editors. Physiological Function in Special Environments. New York: Springer-Verlag; 1989. pp. 191–202. [Google Scholar]
- Hammel HT. Negative plus positive feedback. In: Bligh J, Voigt K, editors. Thermoreception and Temperature Regulation. New York: Springer-Verlag; 1990. pp. 174–182. [Google Scholar]
- Hill JO, Wyatt HR, Reed GW, Peters JC. Obesity and the environment: where do we go from here? Science. 2003;299:853–5. doi: 10.1126/science.1079857. [DOI] [PubMed] [Google Scholar]
- Hjeresen DL, Reed DR, Woods SC. Tolerance to hypothermia induced by ethanol depends on specific drug effects. Psychopharmacology. 1986;89:45–51. doi: 10.1007/BF00175187. [DOI] [PubMed] [Google Scholar]
- Hucka M, Finney A, Sauro HM, Bolouri H, Doyle JC, Kitano H, Arkin AP, Bornstein BJ, Bray D, Cornish-Bowden A, Cuellar AA, Dronov S, Gilles ED, Ginkel M, Gor V, Goryanin II, Hedley WJ, Hodgman TC, Hofmeyr JH, Hunter PJ, Juty NS, Kasberger JL, Kremling A, Kummer U, Le Novere N, Loew LM, Lucio D, Mendes P, Minch E, Mjolsness ED, Nakayama Y, Nelson MR, Nielsen PF, Sakurada T, Schaff JC, Shapiro BE, Shimizu TS, Spence HD, Stelling J, Takahashi K, Tomita M, Wagner J, Wang J. The systems biology markup language (SBML): a medium for representation and exchange of biochemical network models. Bioinformatics. 2003;19:524–31. doi: 10.1093/bioinformatics/btg015. [DOI] [PubMed] [Google Scholar]
- Kanosue K, Romanovsky AA, Hosono T, Chen XM, Yoda T. “Set-point” revisited. In: Nielsen-Johannsen B, Nielsen R, editors. Thermal Physiology. Copenhagen: The August-Krogh Institute; 1997. pp. 39–43. [Google Scholar]
- Katschinski M. Nutritional implications of cephalic phase gastrointestinal responses. Appetite. 2000;34:189–96. doi: 10.1006/appe.1999.0280. [DOI] [PubMed] [Google Scholar]
- Kennedy GC. The role of depot fat in the hypothalamic control of food intake in the rat. Proceedings of the Royal Society of London (Biology) 1953;140:579–592. doi: 10.1098/rspb.1953.0009. [DOI] [PubMed] [Google Scholar]
- Koob GF, Le Moal M. Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology. 2001;24:97–129. doi: 10.1016/S0893-133X(00)00195-0. [DOI] [PubMed] [Google Scholar]
- Krieckhaus EE, Wolf G. Acquisition of sodium by rats: Interaction of innate mechanisms and latent learning. Journal of Comparative and Physiological Psychology. 1968;65:197–201. doi: 10.1037/h0025547. [DOI] [PubMed] [Google Scholar]
- Kurata H, El-Samad H, Iwasaki R, Ohtake H, Doyle JC, Grigorova I, Gross CA, Khammash M. Module-based analysis of robustness tradeoffs in the heat shock response system. PLoS Computational Biology. 2006;2:e59. doi: 10.1371/journal.pcbi.0020059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landsberg L. A teleological view of obesity, diabetes and hypertension. Clinical and Experimental Pharmacology and Physiology. 2006;33:863–867. doi: 10.1111/j.1440-1681.2006.04455.x. [DOI] [PubMed] [Google Scholar]
- Langhans W. Metabolic and glucostatic control of feeding. Proceedings of the Nutrition Society. 1996;55:497–515. doi: 10.1079/pns19960044. [DOI] [PubMed] [Google Scholar]
- Langley LL. Homeostasis. New York: Reinhold; 1965. [Google Scholar]
- Langley LL, editor. Homeostasis: Origins of the Concept. Stroudsberg, PA: Dowden, Hutchinson & Ross, Inc; 1973. [Google Scholar]
- Lê AD, Poulos CX, Cappell H. Conditioned tolerance to the hypothermic effect of ethyl alcohol. Science. 1979;206:1109–1110. doi: 10.1126/science.493999. [DOI] [PubMed] [Google Scholar]
- Lotter EC, Woods SC. Injections of insulin and changes of body weight. Physiology and Behavior. 1977;18:293–297. doi: 10.1016/0031-9384(77)90136-6. [DOI] [PubMed] [Google Scholar]
- Louis-Sylvestre J. Feeding and metabolic patterns in rats with truncular vagotomy or with transplanted beta-cells. American Journal of Physiology. 1978;235:E119–E125. doi: 10.1152/ajpendo.1978.235.2.E119. [DOI] [PubMed] [Google Scholar]
- MacKay EM, Calloway JW, Barnes RH. Hyperalimentation in normal animals produced by protamine insulin. Journal of Nutrition. 1940;20:59–66. [Google Scholar]
- Mansfield JG, Benedict RS, Woods SC. Response specificity of behaviorally augmented tolerance to ethanol supports a learning interpretation. Psychopharmacology. 1983;79:94–98. doi: 10.1007/BF00427791. [DOI] [PubMed] [Google Scholar]
- Mansfield JG, Cunningham CL. Conditioning and extinction of tolerance to the hypothermic effect of ethanol in rats. Journal of Comparative and Physiological Psychology. 1980;94:962–969. doi: 10.1037/h0077824. [DOI] [PubMed] [Google Scholar]
- Mayer J. Regulation of energy intake and the body weight: The glucostatic and lipostatic hypothesis. Annals of the New York Academy of Sciences. 1955;63:14–42. doi: 10.1111/j.1749-6632.1955.tb36543.x. [DOI] [PubMed] [Google Scholar]
- Mayer J, Thomas DW. Regulation of food intake and obesity. Science. 1967;156:328–37. doi: 10.1126/science.156.3773.328. [DOI] [PubMed] [Google Scholar]
- Moore-Ede MC. Physiology of the circadian timing system: predictive versus reactive homeostasis. American Journal of Physiology. 1986;250:R737–R752. doi: 10.1152/ajpregu.1986.250.5.R737. [DOI] [PubMed] [Google Scholar]
- Moran TH, Schulkin J. Curt Richter and regulatory physiology. American Journal of Physiology. 2000;279:R357–R363. doi: 10.1152/ajpregu.2000.279.2.R357. [DOI] [PubMed] [Google Scholar]
- Mrosovsky N. The Physiology of Change. New York: Oxford University Press; 1990. Rheostasis. [Google Scholar]
- Nicolaidis S. Sensory-neuroendocrine reflexes and their anticipatory optimizing role on metabolism. In: Kare MR, Maller O, editors. The Chemical Senses and Nutrition. New York: Academic Press; 1977. pp. 123–140. [Google Scholar]
- Power ML. Viability as opposed to stability: An evolutionary perspective on physiological regulation. In: Schulkin J, editor. Allostasis, Homeostasis, and the Costs of Physiological Adaptation. Cambridge: Cambridge University Press; 2004. pp. 343–364. [Google Scholar]
- Powley TL. The ventromedial hypothalamic syndrome, satiety, and a cephalic phase hypothesis. Psychological Review. 1977;84:89–126. [PubMed] [Google Scholar]
- Proietto J, Rohner-Jeanrenaud F, Ionescu E, Jeanrenaud B. Role of the oropharynx in regulation of glycemia. Diabetes. 1987;36:791–795. doi: 10.2337/diab.36.7.791. [DOI] [PubMed] [Google Scholar]
- Ramsay DS, Seeley RJ, Bolles RC, Woods SC. Ingestive homeostasis: The primacy of learning. In: Capaldi ED, editor. Why We Eat What We Eat. Washington, D. C: American Psychological Association; 1996. pp. 11–27. [Google Scholar]
- Ramsay DS, Woods SC. Biological consequences of drug administration: Implications for acute and chronic tolerance. Psychological Review. 1997;104:170–193. doi: 10.1037/0033-295x.104.1.170. [DOI] [PubMed] [Google Scholar]
- Richet C. Dictionnaire de Physiologie. Paris. 1890;4:721. [Google Scholar]
- Richter CP. Increased salt appetite in adrenalectomized rats. American Journal of Physiology. 1936a;115:155–161. [Google Scholar]
- Richter CP. The spontaneous activity of adrenalectomized rats treated with replacement and other therapy. Endocrinology. 1936b;20:657–666. [Google Scholar]
- Richter CP. Hypophyseal control of behavior. Cold Spring Harbor Symposia on Quantitative Biology. 1937;5:258–268. [Google Scholar]
- Richter CP. Salt taste thresholds of normal and adrenalectomized rats. Endocrinology. 1939;24:3764–3766. [Google Scholar]
- Richter CP. Total self-regulatory functions in animals and human beings. Harvey Lecture Series. 1942–1943;38:63–103. [Google Scholar]
- Richter CP. Sleep and activity: Their relation to the 24-hour clock. Proceedings of the Association of Research in Nervous and Mental Diseases. 1967;45:8–29. [PubMed] [Google Scholar]
- Richter CP, Birmingham JR. Calcium appetite of parathyroidectomized rats used to bioassay substances which affect blood calcium. Endocrinology. 1941;29:655–666. [Google Scholar]
- Richter CP, Eckert JF. Increased calcium appetite of parathyroidectomized rats. Endocrinology. 1937;21:50–54. [Google Scholar]
- Richter CP, Eckert JF. Mineral appetite of parathyroidectomized rats. American Journal of Medical Science. 1939;198:9–16. [Google Scholar]
- Richter CP, Schmidt ECH. Increased fat and decreased carbohydrate appetite of pancreatectomized rats. Endocrinology. 1941;28:179–192. [Google Scholar]
- Richter CP, Schmidt ECH., Jr Behavior and anatomical changes reproduced in rats by pancreatectomy. Endocrinology. 1939;25:698–706. [Google Scholar]
- Romanovsky AA. Do fever and anapyrexia exist? Analysis of set point-based definitions. American Journal of Physiology. 2004;287:R992–R995. doi: 10.1152/ajpregu.00068.2004. [DOI] [PubMed] [Google Scholar]
- Satinoff E. Neural organization and evolution of thermal regulation in mammals. Science. 1978;201:16–22. doi: 10.1126/science.351802. [DOI] [PubMed] [Google Scholar]
- Satinoff E. Behavioral thermoregulation in the cold. In: Fregly MJ, Blatteis CM, editors. Handbook of Physiology, vol 1, section 4: Environmental Physiology. New York: Oxford University Press; 1996. pp. 481–505. [Google Scholar]
- Scharrer E, Langhans W. Control of food intake by fatty acid oxidation. American Journal of Physiology. 1986;250:R1003–R1006. doi: 10.1152/ajpregu.1986.250.6.R1003. [DOI] [PubMed] [Google Scholar]
- Schulkin J. Allostasis: a neural behavioral perspective. Hormones and Behavior. 2003;43:21–7. doi: 10.1016/s0018-506x(02)00035-1. discussion 28–30. [DOI] [PubMed] [Google Scholar]
- Schulkin J. Allostasis, Homeostasis, and the Costs of Physiological Adaptation. Cambridge: Cambridge University Press; 2004. [Google Scholar]
- Schulkin J. Curt Richter. A Life in the Laboratory. Baltimore: The Johns Hopkins University Press; 2005. [Google Scholar]
- Schultz SG. Homeostasis, Humpty Dumpty, and integrative biology. News in Physiological Sciences. 1996;11:238–246. [Google Scholar]
- Schwartz MW, Woods SC, Seeley RJ, Barsh GS, Baskin DG, Leibel RL. Is the energy homeostasis system inherently biased toward weight gain? Diabetes. 2003;52:232–238. doi: 10.2337/diabetes.52.2.232. [DOI] [PubMed] [Google Scholar]
- Seeley RJ, Ramsay DS, Woods SC. Regulation of food intake: Interactions between learning and physiology. In: Bouton ME, Fanselow MS, editors. Learning, Motivation, and Cognition: The Functional Behaviorism of Robert C. Bolles. Washington, D. C: American Psychological Association; 1997. pp. 99–115. [Google Scholar]
- Smith GP, Epstein AN. Increased feeding in response to decreased glucose utilization in rat and monkey. American Journal of Physiology. 1969;217:1083–1087. doi: 10.1152/ajplegacy.1969.217.4.1083. [DOI] [PubMed] [Google Scholar]
- Somjen GG. The missing error signal - Regulation beyond negative feedback. News in Physiological Sciences. 1992;7:184–185. [Google Scholar]
- Sterling P. Principles of allostasis: optimal design, predictive regulation, pathophysiology, and rational therapeutics. In: Schulkin J, editor. Allostasis, Homeostasis and the Costs of Physiological Adaptation. Cambridge: Canbridge University Press; 2004. pp. 17–64. [Google Scholar]
- Storlien LH, Bruce DG. Mind over metabolism: the cephalic phase in relation to non-insulin-dependent diabetes and obesity. Biological Psychology. 1989;28:3–23. doi: 10.1016/0301-0511(89)90108-7. [DOI] [PubMed] [Google Scholar]
- Strubbe JH, van Dijk G. The temporal organization of ingestive behaviour and its interaction with regulation of energy balance. Neuroscience and Biobehavioral Reviews. 2002;26:485–498. doi: 10.1016/s0149-7634(02)00016-7. [DOI] [PubMed] [Google Scholar]
- Strubbe JH, Woods SC. The timing of meals. Psychological Review. 2004;111:128–141. doi: 10.1037/0033-295X.111.1.128. [DOI] [PubMed] [Google Scholar]
- Teff K. Nutritional implications of the cephalic-phase reflexes: endocrine responses. Appetite. 2000;34:206–213. doi: 10.1006/appe.1999.0282. [DOI] [PubMed] [Google Scholar]
- Weiner N. Cybernetics; Control and Communication in the Animal and Machine. New York: John Wiley and Sons; 1948. [Google Scholar]
- Weisinger RS, Woods SC, Skorupski JD. Sodium deficiency and latent learning. Psychonomic Science. 1970;19:307–308. [Google Scholar]
- Woods SC. The eating paradox: How we tolerate food. Psychological Reviews. 1991;98:488–505. doi: 10.1037/0033-295x.98.4.488. [DOI] [PubMed] [Google Scholar]
- Woods SC. The house economist and the eating paradox. Appetite. 2002;38:161–165. doi: 10.1006/appe.2001.0468. [DOI] [PubMed] [Google Scholar]
- Woods SC, Decke E, Vasselli JR. Metabolic hormones and regulation of body weight. Psychological Review. 1974;81:26–43. doi: 10.1037/h0035927. [DOI] [PubMed] [Google Scholar]
- Woods SC, Ramsay DS. Pavlovian influences over food and drug intake. Behavioral Brain Research. 2000;110:175–182. doi: 10.1016/s0166-4328(99)00194-1. [DOI] [PubMed] [Google Scholar]
- Woods SC, Strubbe JH. The psychobiology of meals. Psychonomic Bulletin and Review. 1994;1:141–155. doi: 10.3758/BF03200770. [DOI] [PubMed] [Google Scholar]
- Yates FE. Homeostasis. In: Birren JE, editor. Encyclopedia of Gerontology: age, aging, and the aged. Vol. 1. San Diego, CA: Academic Press, Inc; 1996. pp. 679–686. [Google Scholar]