

Abstract
There is evidence that cannabinoid-based medicines that are selective for different targets in the cannabinoid signalling system (e.g. receptors, inactivation mechanism, enzymes) might be beneficial in basal ganglia disorders, namely Parkinson's disease (PD) and Huntington's disease (HD). These benefits not only include the alleviation of specific motor symptoms [e.g. choreic movements with cannabinoid receptor type 1 (CB1)/transient receptor potential vanilloid type 1 agonists in HD; bradykinesia with CB1 antagonists and tremor with CB1 agonists in PD], but also the delay of disease progression due to the neuroprotective properties demonstrated for cannabinoids (e.g. CB1 agonists reduce excitotoxicity; CB2 agonists limit the toxicity of reactive microglia; and antioxidant cannabinoids attenuate oxidative damage). In addition, extensive biochemical, anatomical, physiological and pharmacological studies have demonstrated that: (i) the different elements of the cannabinoid system are abundant in basal ganglia structures and they are affected by these disorders; (ii) the cannabinoid system plays a prominent role in basal ganglia function by modulating the neurotransmitters that operate in the basal ganglia circuits, both in healthy and pathological conditions; and (iii) the activation and/or inhibition of the cannabinoid system is associated with important motor responses that are maintained and even enhanced in conditions of malfunctioning and/or degeneration. In this article we will review the available data regarding the relationship between the cannabinoid system and basal ganglia activity, both in healthy and pathological conditions and will also try to identify future lines of research expected to increase current knowledge about the potential therapeutic benefits of targeting this system in PD, HD and other basal ganglia disorders.
Keywords: cannabinoids, cannabinoid receptors, movement, basal ganglia, basal ganglia disorders, symptom relief, neuroprotection
Function of the cannabinoid signalling system in the basal ganglia
Endocannabinoid ligands, their synthesizing and degrading enzymes, and their receptors are particularly abundant in basal ganglia structures (e.g. caudate-putamen, globus pallidus, substantia nigra) when compared with other brain regions (Herkenham et al., 1991a; Mailleux and Vanderhaeghen, 1992; Tsou et al., 1998a,b; Bisogno et al., 1999; Dinh et al., 2002; Egertova et al., 2003; Köfalvi et al., 2005). This is the logical consequence of the important modulatory function exerted by the cannabinoid signalling system on the control of movement, which is in turn the consequence of the location of specific elements of this signalling system in key groups of neurons and synapses within basal ganglia circuits (see Figure 1 and Gerdeman and Fernández-Ruiz, 2008, for review). Thus, the nuclei that receive striatal efferent outputs, namely the external globus pallidus and substantia nigra pars reticulata/internal globus pallidus, contain high levels of endocannabinoids, either anandamide or 2-arachidonoylglycerol (Bisogno et al., 1999; Di Marzo et al., 2000), as well as the cannabinoid receptor type 1 (CB1) receptor, the cannabinoid receptor type predominantly involved in the control of movement in healthy circumstances (reviewed in Fernández-Ruiz and González, 2005). CB1 receptor binding sites are located presynaptically in nerve terminals of striatal projection neurons, the so-called ‘medium-spiny striatal neurons’ (Herkenham et al., 1991b; Tsou et al., 1998a; Hohmann and Herkenham, 2000; Fusco et al., 2004), which explain the high levels of mRNA transcripts for this receptor type found in the caudate-putamen (Mailleux and Vanderhaeghen, 1992). Indeed, CB1 receptors are located in striatal neurons projecting to both the substantia nigra pars reticulata/internal globus pallidus (the so-called ‘direct’ striatal efferent pathway) and the external globus pallidus (the so-called ‘indirect’ striatal efferent pathway). Both groups of neurons use γ-aminobutiric acid (GABA) as a neurotransmitter, although also express selective phenotypical markers (e.g. enkephalin and D2 receptors in striatopallidal neurons, and dynorphin/substance P and D1 receptors in striatonigral neurons; see Hohmann and Herkenham, 2000; Julián et al., 2003). Intrinsic striatal neurons do not generally contain CB1 receptors, except most of the striatal GABAergic interneurons that are labelled with parvalbumin, and a few interneurons of the cholinergic subpopulation (Hohmann and Herkenham, 2000; Fusco et al., 2004; Uchigashima et al., 2007). CB1 receptors are also present presynaptically in glutamatergic neurons within the basal ganglia circuits, including the afferences to the striatum coming from cortical structures (Köfalvi et al., 2005; Uchigashima et al., 2007) and, in particular from those arising preferentially in the substantia nigra and whose cell bodies are located in the subthalamic nucleus (Mailleux and Vanderhaeghen, 1992; Tsou et al., 1998a). By contrast, nigrostriatal dopaminergic neurons do not contain CB1 receptors, although they do contain the vanilloid TRPV1 receptor (Mezey et al., 2000; de Lago et al., 2004a; Lastres-Becker et al., 2005; Tzavara et al., 2006), a receptor functionally related to the cannabinoid signalling system, which is also evident in different basal ganglia structures [e.g. striatum (Maccarrone et al., 2008), globus pallidus (Cristino et al., 2006)]. Indeed, certain endocannabinoids, like anandamide and N-arachidonoyldopamine, but not 2-arachidonoylglycerol, can also act as endogenous ligands for this TRPV1 cation channel (reviewed in Starowicz et al., 2007). On the other hand, glial cells, mainly astrocytes, might naturally express the cannabinoid receptor type 2 (CB2) receptor type, although the levels of this receptor type would be very small in healthy conditions (Sheng et al., 2005; Sagredo et al., 2009). Interestingly, these receptors are strongly up-regulated in astrocytes and they are induced in microglial cells activated in response to different types of cytotoxic insults, such as those occurring in most basal ganglia disorders (Sagredo et al., 2008).
Figure 1.
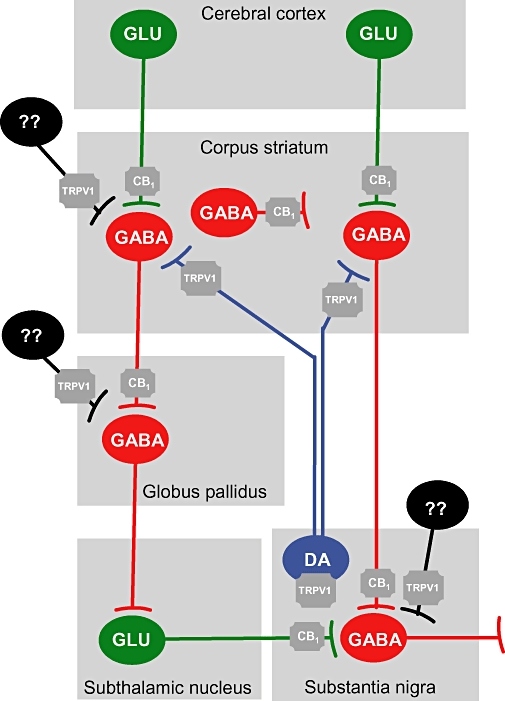
Location of CB1 and TRPV1 receptors in specific neuronal subpopulations within basal ganglia circuits. Regulatory pathways are indicated in blue, whereas inhibitory and excitatory inputs are indicated in red and green respectively. Unknown neurons are shown in black. CB1, cannabinoid receptor type 1; GABA, γ-aminobutiric acid; GLU, glutamate; TRPV1, transient receptor potential vanilloid type 1.
Given the abundance of elements of the cannabinoid signalling system in the basal ganglia, it is likely that the activation or the inhibition of this signalling system might have an important influence on different motor responses (reviewed in Fernández-Ruiz and González, 2005). However, the complex localization of these elements at different sites in the basal ganglia circuits, particularly of the CB1 receptor, may sometimes explain the paradoxical effects observed (reviewed in Fernández-Ruiz and González, 2005; Gerdeman and Fernández-Ruiz, 2008). In general, the substances that activate cannabinoid signalling (e.g. direct receptor agonists, inhibitors of endocannabinoid inactivation) have powerful effects on the control of movement, mostly of an inhibitory nature (Crawley et al., 1993; Fride and Mechoulam, 1993; Wickens and Pertwee, 1993; Romero et al., 1995a,b; Jarbe et al., 1998; Giuffrida et al., 1999; González et al., 1999; Beltramo et al., 2000; McLaughlin et al., 2000; Sañudo-Peña et al., 2000; de Lago et al., 2002; 2004b). However, most studies published so far have found some variation in the magnitude and duration of the motor effects, depending on the characteristics of these compounds in terms of receptor affinity, potency and/or metabolic stability (reviewed in Gerdeman and Fernández-Ruiz, 2008). Most of these effects, if not all, originate through the activation of the CB1 receptor as the motor effects of most cannabinoid agonists are usually prevented by rimonabant and other selective CB1 receptor antagonists (Souilhac et al., 1995; Di Marzo et al., 2001; reviewed in Fernández-Ruiz and González, 2005). However, certain motor effects mediated by anandamide or its analogue AM404 (a compound initially developed as an inhibitor of the endocannabinoid uptake but that also exhibits other properties) were reversed by the TRPV1 receptor antagonist capsazepine, but not by rimonabant. Hence, it would appear that such effects are more closely related to the capacity of these compounds to directly bind to and activate vanilloid TRPV1 receptors rather than their ability to activate CB1 receptors (Lastres-Becker et al., 2002a; 2003a; de Lago et al., 2004a). On the other hand, the inverse agonistic properties of most CB1 receptor antagonists means that their in vivo administration frequently causes hyperlocomotion (Compton et al., 1996), concordant with the motor activity observed in mice deficient in this receptor (Ledent et al., 1999).
The motor effects resulting from the activation or blockade of cannabinoid signalling are related to the capacity of this system to modulate the activity of the neurotransmitters that participate in the control of basal ganglia function, namely glutamate and GABA (responsible for the excitatory and inhibitory inputs respectively), and dopamine (which plays a regulatory role; reviewed in Sañudo-Peña et al., 1999; Romero et al., 2002; Fernández-Ruiz and González, 2005; Gerdeman and Fernández-Ruiz, 2008). The identification of CB1 receptors in several GABAergic and glutamatergic synapses within the basal ganglia, as well as the presence of TRPV1 receptors in nigrostriatal dopaminergic neurons, enables endocannabinoids to directly control the function of these three key neurotransmitters. In addition, the preferential presynaptic location of these receptors makes it likely that endocannabinoids mainly control presynaptic events, such as the synthesis, release or reuptake of these neurotransmitters (see Figure 2, and Gerdeman and Fernández-Ruiz, 2008, for review). In fact, different pharmacological (e.g. administration of cannabinoids combined with agonists or antagonists for receptors of these neurotransmitters; Romero et al., 1996; Meschler et al., 2000), electrophysiological (e.g. monitoring the inhibitory or excitatory currents after the application of cannabinoids in vivo; Huang et al., 2001; Gerdeman et al., 2002; Wallmichrath and Szabo, 2002) and neurochemical (e.g. analysis of neurotransmitter synthesis, release and reuptake in vivo or in vitro; Köfalvi et al., 2005) studies support the idea that the main function of cannabinoid signalling within the basal ganglia circuitry is that of a retrograde signalling system in GABAergic and glutamatergic synapses (reviewed in Gerdeman and Fernández-Ruiz, 2008). Accordingly, endocannabinoids and CB1 receptors seem to play a key role in the synaptic depolarization-induced suppression of excitation or inhibition, and also in long-term depression operating at striatal synapses (reviewed in Gerdeman and Fernández-Ruiz, 2008). Anandamide might also be able to directly inhibit certain presynaptic events in dopaminergic synapses within the striatum by acting through TRPV1 receptors located in nigrostriatal dopaminergic neurons (de Lago et al., 2004a). However, it is also possible to modulate dopamine transmission by interacting at the level of the G protein/adenylyl cyclase signal transduction mechanisms shared by both CB1 and D1/D2 dopamine receptors (Di Marzo et al., 2000; Meschler and Howlett, 2001), especially as CB1 receptors colocalize with D1 receptors in striatonigral neurons and with D2 receptors in the neurons projecting to the globus pallidus (reviewed in Fernández-Ruiz and González, 2005).
Figure 2.
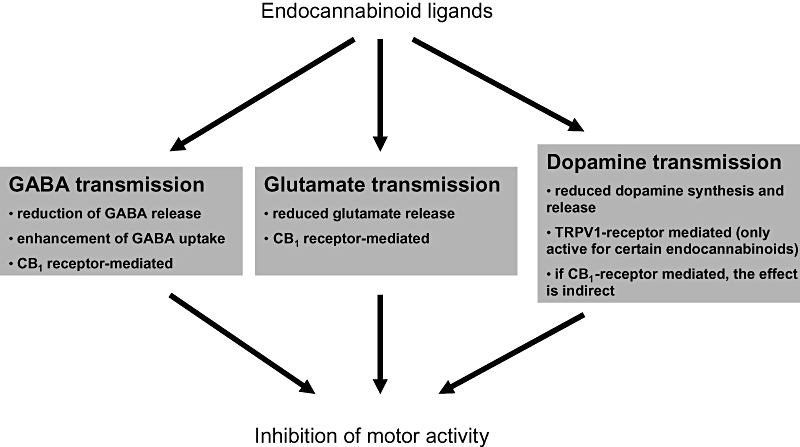
Schematic overview of the modulatory effects of endocannabinoid ligands and their receptors on the activity of the three major neurotransmitters operating in basal ganglia circuits. CB1, cannabinoid receptor type 1; GABA, γ-aminobutiric acid; TRPV1, transient receptor potential vanilloid type 1.
Therefore, the experimental evidence described above: (i) abundant endocannabinoid signalling elements in the basal ganglia; (ii) the important motor effects following the activation or inhibition of this signalling system; and (iii) the modulation of key neurotransmitters within the basal ganglia by endocannabinoids, supports the idea that the cannabinoid signalling system plays an important role in the activity of these brain circuits. Hence, the pharmacological management of this signalling system might serve as a novel therapy for the treatment of motor dysfunction in those disorders affecting basal ganglia structures. This concept is also supported by the fact that the levels of different elements of the cannabinoid system, in particular the CB1 receptor, seem to be significantly altered in various basal ganglia disorders. Such alterations in the proportions of these proteins have been observed in samples obtained from patients (Glass et al., 1993; 2000; Richfield and Herkenham, 1994; Lastres-Becker et al., 2001a; Pisani et al., 2005; Battista et al., 2007) and in different experimental models of these diseases generated by neurotoxin lesion (Zeng et al., 1999; Page et al., 2000; Romero et al., 2000; Lastres-Becker et al., 2001a; 2002a,b; 2004; 2005), by mutation or deletion of specific genes (Lastres-Becker et al., 2002c; García-Arencibia et al., 2008a; Pazos et al., 2008), or by inducing apoptosis in cultured neurons (Lastres-Becker et al., 2005). This cannabinoid-based therapy would not only be aimed at alleviating specific motor symptoms, such as bradykinesia, tremor, dystonia or choreic movements, but also at delaying/arresting the degeneration of the basal ganglia given the well-known neuroprotective/neuroregenerative properties of cannabinoid agonists (reviewed in Fernández-Ruiz et al., 2007; Gerdeman and Fernández-Ruiz, 2008). In general, Parkinson's disease (PD) and Huntington's disease (HD) are the two basal ganglia disorders that have attracted most attention with regards the potential clinical application of cannabinoids (reviewed in Fernández-Ruiz and González, 2005; Sagredo et al., 2007a).
Parkinson's disease is the most prevalent neurodegenerative disorder affecting the basal ganglia. The disease is caused either by environmental factors (e.g. pesticides, metals, antidopaminergic agents) or anomalies affecting the genes encoding several proteins (e.g. α-synuclein, parkin, PINK1), although it is possible that the combination of both factors may also be important. These primary factors would elicit a series of cooperative cytotoxic events (e.g. abnormal proteolysis, protein aggregation, mitochondrial dysfunction, excitotoxicity, oxidative stress and glial activation) that would culminate in the progressive death of dopaminergic neurons located in the substantia nigra pars compacta. These alterations would in turn originate severe dopaminergic denervation of the striatum and the occurrence of different neurological symptoms, such as bradykinesia, rigidity and tremor (see Thomas and Beal, 2007, for a recent review). HD is an inherited (autosomal-dominant), progressive and fatal neurodegenerative disorder caused by an excess of CAG repeats in the gene encoding for huntingtin, which provokes a polyglutamine expansion in the amino-terminal portion of this protein. The expansion originates several conformational changes that result in altered protein–protein interactions, abnormal protein aggregation and proteolysis, and they subsequently provoke transcriptional dysregulation, mitochondrial complex II deficiencies, excitotoxicity, oxidative damage and glial activation. This sequence of cytotoxic events damages specific neuronal subpopulations, particularly the striatal neurons that project to the globus pallidus and the substantia nigra, originating a biphasic pattern of motor abnormalities that evolves from an early hyperkinetic phase (choreic movements) to a late akinetic and more disabling phase (see Roze et al., 2008, for a recent review). Although PD and HD are the two basal ganglia disorders for which the potential therapeutic application of cannabinoids has been best studied, the manipulation of cannabinoid signalling might also be useful in other disorders directly or indirectly affecting the function of the basal ganglia, for example, dystonia (Fox et al., 2002a; Richter and Loscher, 2002), dyskinesias (reviewed in Brotchie, 2003), Gilles de la Tourette's syndrome (reviewed in Müller-Vahl, 2003), Alzheimer's disease (reviewed in Campbell and Gowran, 2007) or multiple sclerosis (reviewed in Baker et al., 2007).
Changes to the cannabinoid signalling system in basal ganglia disorders
Given the hypokinetic profile of the cannabinoid agonists able to activate CB1 receptors, one might expect the cannabinoid signalling system (and particularly that acting through this receptor type) would become hypoactive in the basal ganglia in HD and other hyperkinetic disorders, whereas it would be up-regulated in hypokinetic disorders like PD. If this hypothesis were correct, it would explain the type of motor symptoms that are predominant in these disorders: choreic movements and bradykinesia respectively (see Figure 3). This hypothesis has been confirmed in numerous studies conducted in recent years in which different endocannabinoid elements have been analysed in patients, in particular the CB1 receptor type (Glass et al., 1993; 2000; Richfield and Herkenham, 1994; Lastres-Becker et al., 2001a; Pisani et al., 2005; Battista et al., 2007). Indeed, this system has also been studied in rodents or primates lesioned with neurotoxins (Page et al., 2000; Romero et al., 2000; Lastres-Becker et al., 2001b; 2002b; Gubellini et al., 2002; Fernández-Espejo et al., 2004), mice with mutations or deletion of specific genes (Denovan-Wright and Robertson, 2000; Lastres-Becker et al., 2002c; Centonze et al., 2005; Bisogno et al., 2008; García-Arencibia et al., 2008a), or animals subjected to different pharmacological treatments (Di Marzo et al., 2000), all animal models that totally or partially reproduce aspects of both diseases. However, in these studies it has also been emphasized that these responses are not the only changes experienced by the cannabinoid signalling system in both disorders (see Figure 3 for a schematic representation). Down-regulation in HD and overactivity in PD might be associated with degeneration in specific basal ganglia structures, although these changes or others might also be evident in early phases of these diseases when cell death does not occur or when it is sufficiently infrequent that it does not elicit motor symptoms. For example, fewer CB1 receptors (down-regulation), or even their loss of function (desensitization), have already been documented in post-mortem samples of HD patients at very early stages of the disease (Glass et al., 2000). Likewise, similar events have been described in different transgenic mouse models of HD (e.g. R6/1, R6/2 and HD94) analysed at presymptomatic ages and in the absence of neuronal death (Denovan-Wright and Robertson, 2000; Lastres-Becker et al., 2002c; Naver et al., 2003; McCaw et al., 2004; Centonze et al., 2005), or in rats lesioned with the mitochondrial complex II inhibitor 3-nitropropionic acid at stages prior to the appearance of the first signs of neurodegeneration and neurological deterioration (Lastres-Becker et al., 2004). Similar observations have been made in the case of PD, where despite the up-regulation detected at intermediate and advanced phases of the disease (Romero et al., 2000; Lastres-Becker et al., 2001a), there is also evidence of a reduction in CB1 receptors at early and presymptomatic ages in different knockout mouse models of three different genes linked to the development of PD (e.g. PARK1 encoding α-synuclein, PARK2 encoding parkin and PARK6 encoding the PINK1 protein; García-Arencibia et al., 2008a). Interestingly, at later symptomatic ages the same up-regulatory responses of CB1 receptors were evident in these mice (García-Arencibia et al., 2008a), as observed in samples from patients (Lastres-Becker et al., 2001a) or in laboratory animals lesioned with neurotoxins (Romero et al., 2000). Therefore, down-regulation/desensitization of CB1 receptors seems to be an early event in both HD and PD. Furthermore, this response appears to be associated with neuronal malfunctioning in the absence of cell death, presumably indicating that these losses might be involved in the initiation or early progression of striatal or nigral degeneration (reviewed in Sagredo et al., 2007a). It is possible that such early malfunctioning or loss of CB1 receptors might render basal ganglia neurons more vulnerable to different cytotoxic stimuli that frequently operate in both disorders. This might be so for excitotoxicity as CB1 receptors are involved in the inhibition of glutamate release and thus, their loss or malfunction might enhance extracellular glutamate levels and aggravate the damage caused by this aminoacid (reviewed in van der Stelt et al., 2002; Centonze et al., 2007; Sagredo et al., 2007a; see Figure 3). This would explain why the activation of this receptor type might be used as a neuroprotectant strategy (see below). In addition, the loss and/or malfunctioning of CB1 receptors might also aggravate other cytotoxic stimuli, such as oxidative stress or glial activation, although CB1 receptors seem to be less important in these processes.
Figure 3.
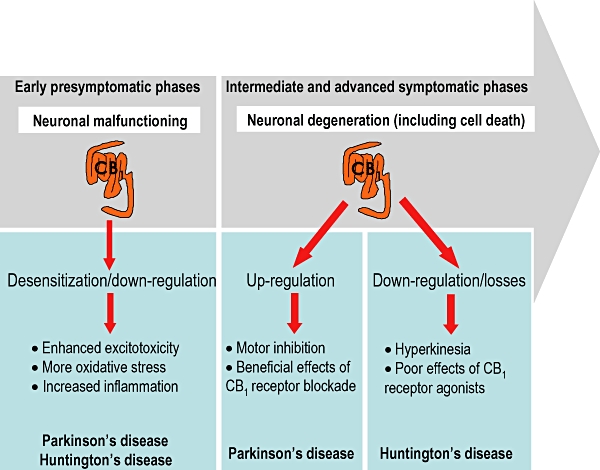
Changes to CB1 receptors in the basal ganglia during the progression of Parkinson's and Huntington's diseases. CB1, cannabinoid receptor type 1.
The loss of CB1 receptors under conditions of neuronal malfunctioning in HD is much more marked when striatal degeneration is already evident. This was evident in post-mortem samples collected from patients at intermediate and advanced phases of the disease (Glass et al., 1993; Richfield and Herkenham, 1994), as well as in transgenic mice analysed at late and symptomatic ages (Pazos et al., 2008) and in rats with striatal atrophy generated by excitotoxins (Page et al., 2000) or mitochondrial toxins (Lastres-Becker et al., 2001b; 2002b). Similarly, the levels of endocannabinoids were also markedly reduced in the brain (Lastres-Becker et al., 2001b; Bisogno et al., 2008), even though some recent studies in patients identified an increase in endocannabinoid levels in the blood due to a reduction in fatty acid amide hydrolase (FAAH) activity (Battista et al., 2007). In conjunction with striatal degeneration, the loss of CB1 receptors seems to be a mere side effect caused by the progressive destruction of CB1 receptor-containing medium-spiny GABAergic neurons, which would explain why compounds targeting this receptor type are not effective against the hyperkinesia characteristic of HD (Lastres-Becker et al., 2003a; de Lago et al., 2006; see Figure 3). However, in conditions of striatal degeneration, the other major cannabinoid receptor type, the CB2 receptor, is significantly up-regulated in glial elements that are recruited to and migrate to lesion sites (e.g. astrocytes and particularly reactive microglial cells; Sagredo et al., 2009), a response already described in other neurodegenerative or neuroinflammatory disorders (reviewed in Fernández-Ruiz et al., 2007; 2008). Interestingly, this response did not exist when animals were analysed at presymptomatic stages, when the major pathological hallmark is neuronal malfunction, supporting the idea that it is a phenomenon directly related to striatal degeneration and susceptible to pharmacological management (see below and Pazos et al., 2008).
The similarities found in the response of CB1 receptors during early presymptomatic phases of HD and PD become inverted in intermediate and advanced phases of both these disorders. Thus, the progression of the nigral pathology and the appearance of major parkinsonian symptoms is associated with overactivity of the cannabinoid signalling system in most basal ganglia structures in patients (e.g. elevated endocannabinoid levels, increased CB1 receptor density and function: Lastres-Becker et al., 2001a; Pisani et al., 2005), as well as in animal models of this disease (Di Marzo et al., 2000; Romero et al., 2000; Lastres-Becker et al., 2001a; Gubellini et al., 2002; Fernández-Espejo et al., 2004; van der Stelt et al., 2005). Such a situation would be in accordance with the hypokinesia that characterizes this disease and with the possibility that CB1 receptor antagonists might be useful for the alleviation of bradykinesia (see below and Figure 3). Interestingly, treatment with levodopa has been found to reverse both the elevation in endocannabinoid levels (Maccarrone et al., 2003; van der Stelt et al., 2005) and the increase in CB1 receptor density (Lastres-Becker et al., 2001a). Another important difference between PD and HD is that CB2 receptors do not appear to be up-regulated in glial elements recruited to the lesioned substantia nigra, either in patients or in animal models of PD. This might explain the failure of CB2 receptor agonists to act as neuroprotectants in parkinsonian animals (García-Arencibia et al., 2007; see below), despite the fact that astrocytosis and reactive microgliosis have been documented in the substantia nigra in both patients and animal models of the disease (reviewed in Sagredo et al., 2007a).
Alleviation of motor symptoms by cannabinoid-related compounds in basal ganglia disorders
In the late 1990s, clinical trials examined whether plant-derived cannabinoids (reviewed in Consroe, 1998) or some of their synthetic analogues (Müller-Vahl et al., 1999) might be used to improve choreic movements in HD patients, in the light of the hypofunctionality of the cannabinoid system in this disease. However, rather than improving hyperkinesia, these studies reported an aggravation of choreic movements (Consroe, 1998; Müller-Vahl et al., 1999). It is possible that this is related to the lack of TRPV1 receptor activity of the cannabinoid agonists used in those clinical trials, as recent studies conducted in animal models have demonstrated that only TRPV1 receptor agonists (e.g. AM404, arvanil, capsaicin) alleviate hyperkinesia and restore neurochemical deficits (Lastres-Becker et al., 2002a; 2003a; de Lago et al., 2005). By contrast, CB1 receptor agonists (e.g. CP55,940) were not particularly effective in combating these symptoms, particular when used at intermediate or advanced phases of the disease (grades 2–4), when there is a progressive loss of these receptors in association with the death of striatal projection neurons. Indeed, these observations were further confirmed in experiments with selective antagonists of both receptor types (Lastres-Becker et al., 2003a). Therefore, TRPV1 receptors appear to be a novel therapeutic target susceptible to clinical exploitation for the treatment of hyperkinesia in HD (see Figure 4). It is possible that the best clinical option might involve developing compounds with a dual capacity that can activate both TRPV1 and CB1 receptors, at least for the first phases of HD (grades 0–2). However, the relative contribution of both targets is likely to vary during the course of the disease due to a progressive loss of CB1 receptors with no concomitant change in TRPV1 receptors (reviewed in Lastres-Becker et al., 2003b).
Figure 4.
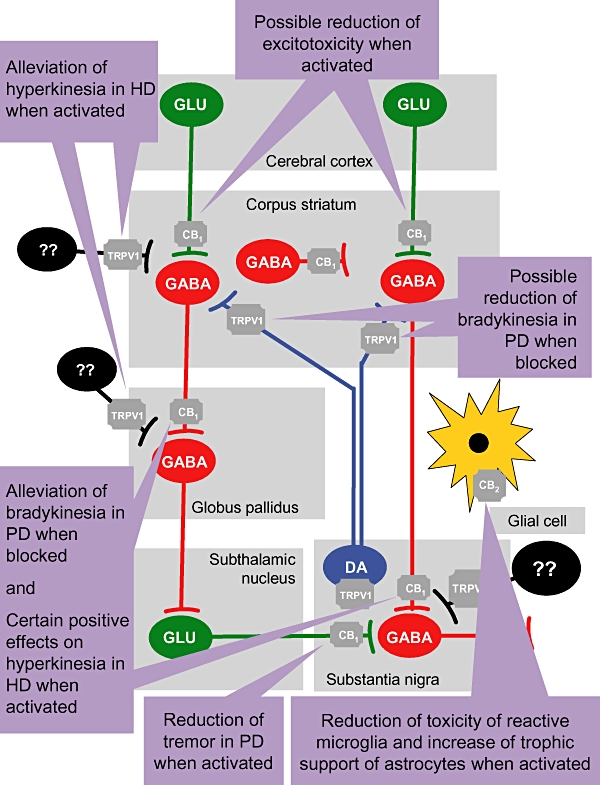
On the scheme shown in Figure 1, a diagram has been superimposed to show the different targets (CB1, CB2 and TRPV1 receptors) that might mediate the ability of cannabinoid-based medicines to alleviate specific symptoms, or to delay/arrest the progression of the disease in basal ganglia disorders. CB1, cannabinoid receptor type 1; DA, dopamine; GABA, γ-aminobutiric acid; GLU, glutamate; HD, Huntington's disease; PD, Parkinson's disease; TRPV1, transient receptor potential vanilloid type 1.
The overactive cannabinoid transmission in PD and the hypokinetic profile of most cannabinoid agonists does not support the use of these compounds to alleviate the motor disruption in this disease, particularly in terms of bradykinesia, the major parkinsonian symptom. In this case, CB1 receptor antagonists rather than agonists would seem to be a better option (see Figure 4). This issue has been addressed in a series of preclinical (Di Marzo et al., 2000; Meschler et al., 2001) and clinical studies (Mesnage et al., 2004). Although the initial results were controversial, it is possible that the blockade of CB1 receptors might only be effective in special circumstances (e.g. with the use of low doses, at very advanced phases of the disease only and in patients with a poor levodopa response; Fernández-Espejo et al., 2005; González et al., 2006; García-Arencibia et al., 2008b). If this were the case, it would be possible to generate an antiparkinsonian agent for conditions in which classic therapy generally fails. The blockade of TRPV1 receptors might be also useful against the motor inhibition typical of PD, given the inhibition of dopamine activity caused by the activation of these receptors (de Lago et al., 2004a). However, in this case the death of nigrostriatal neurons in which the TRPV1 receptor is located (Lastres-Becker et al., 2005) might represent an important disadvantage. CB1 receptor antagonists might also be useful for delaying and reducing the dyskinetic states caused by chronic levodopa treatment in PD patients and animal models (Brotchie, 2000; 2003; van der Stelt et al., 2005), even though this property has also paradoxically been attributed to CB1 receptor agonists (Brotchie, 1998; Sieradzan et al., 2001; Fox et al., 2002b). Such effects would be consistent with the presence of CB1 receptors in multiple sites (both in excitatory and inhibitory synapses) within basal ganglia circuits, which might facilitate these paradoxical effects (reviewed in Fernández-Ruiz and González, 2005; Gerdeman and Fernández-Ruiz, 2008). Along similar lines, CB1 receptor agonists (or inhibitors of endocannabinoid inactivation) have also been attributed beneficial effects in PD (see Figure 4): (i) in reducing the tremor associated with the overactivity of the subthalamic nucleus (Sañudo-Peña et al., 1998; 1999), although the only clinical trial developed so far produced negative results (Frankel et al., 1990); (ii) by interacting with dopamine agonists to improve certain motor impairments (Brotchie, 1998; Sañudo-Peña et al., 1998; Kreitzer and Malenka, 2007); and (iii) by delaying the progression of nigral degeneration, although this effect may not be caused by their hypokinetic profile, but rather by their neuroprotective properties (see below).
Neuroprotection/neuroregeneration with cannabinoids in basal ganglia disorders
Cannabinoids have also been proposed to offer a novel type of therapy for HD and PD, delaying, arresting or repairing the striatal and nigral damage due to their capacity to buffer excitotoxicity, calcium influx, oxidative injury, cerebro-vasoconstriction, body temperature and/or inflammation in conditions of acute or chronic neurodegeneration (reviewed in Fernández-Ruiz et al., 2005; van der Stelt and Di Marzo, 2005), as well as through their influence on the survival, proliferation and differentiation of neural progenitors (reviewed in Galve-Roperh et al., 2007). Unfortunately, this possibility has not yet progressed beyond the preclinical stage, although it is sufficiently robust to justify an urgent clinical evaluation with some of the already available cannabinoid-based medicines.
In HD, both the early desensitization of CB1 receptors located in certain neuronal subpopulations and the late up-regulation of CB2 receptors in glial elements suggest that compounds selectively targeting both receptor types might be able to attenuate striatal degeneration, as mentioned above. The importance of CB1 receptors has been demonstrated in conditions of excitotoxicity, such as those observed in rats after the lesion of the striatum with the excitotoxin quinolinate (Pintor et al., 2006). By contrast, selective CB2 receptor agonists are effective in conditions of inflammation and glial activation, such as those occurring after the intrastriatal application of the reversible mitochondrial complex II inhibitor malonate in rats (Fernández-Ruiz et al., 2007; 2008; Sagredo et al., 2009). In addition, some plant-derived antioxidant cannabinoids can protect striatal projection neurons from death in conditions of mitochondrial dysfunction and oxidative stress, such as those provoked by the administration of the irreversible mitochondrial complex II inhibitor 3-nitropropionic acid in rats. These cannabinoids include Δ9-tetrahydrocannabinol (Δ9-THC) (Lastres-Becker et al., 2004), which binds to both CB1 and CB2 receptors, and cannabidiol (Sagredo et al., 2007b), which has negligible activity at both receptor types. Given the fact that HD is a neurodegenerative disorder in which the death of striatal neurons is caused by cooperation between different cytotoxic mechanisms, including excitotoxicity, inflammation and oxidative damage, any neuroprotective cannabinoid compound(s) suitable for clinical evaluation in HD should be broad-spectrum cannabinoids, or alternatively combinations of specific existing compounds could be assessed. The objective would be to activate both CB1 and CB2 receptors (e.g. to normalize glutamate homeostasis and to reduce local inflammatory events caused by glial activation), but also to decrease oxidative damage with an antioxidant cannabinoid. Hence, the cannabinoid-based preparation Sativex, a combination of Δ9-THC and cannabidiol (reviewed in Russo and Guy, 2006), might be a good candidate for clinical evaluation in HD.
Despite the fact that the hypokinetic profile of most cannabinoid agonists may represent a disadvantage in the treatment of PD, there is also preclinical evidence indicating that certain cannabinoid agonists may offer neuroprotection in this disease (reviewed in Fernández-Ruiz et al., 2005). This has been studied in rats with hemiparkinsonism generated by unilateral injection of the neurotoxin 6-hydroxydopamine (Lastres-Becker et al., 2005; García-Arencibia et al., 2007). Δ9-THC and cannabidiol were the first cannabinoids shown to be capable of attenuating the damage to nigrostriatal dopaminergic neurons caused by this neurotoxin (Lastres-Becker et al., 2005). As mentioned above, Δ9-THC can bind to and activate both CB1 and CB2 receptors, although cannabidiol has negligible activity at both receptor types. Hence, the neuroprotective effect of both compounds is probably exerted through their cannabinoid receptor-independent antioxidant properties (Lastres-Becker et al., 2005). This was further confirmed by examining the antioxidant properties of additional compounds that are more selective for both receptor types (de Lago et al., 2006; García-Arencibia et al., 2007). Similar findings were recently obtained in an invertebrate model of Parkinsonism (Jiménez-del Río et al., 2008) and in an in vitro model of 6-hydroxydopamine toxicity (Lastres-Becker et al., 2005). Both studies used synthetic cannabinoid agonists (e.g. CP55,940 and HU-210 respectively), which also have well-demonstrated cannabinoid receptor-independent antioxidant properties (Marsicano et al., 2002). Therefore, the cannabinoid receptor-independent antioxidant profile of certain cannabinoids seems to be a key mechanism in the protection of nigrostriatal neurons from death in PD. This observation predisposes the cannabinoid-based medicine, Sativex, to clinical evaluation. By contrast, the studies conducted on the role of CB2 receptors in PD indicate that this receptor is not a relevant target in this disease (García-Arencibia et al., 2007), unlike in HD.
Alternatively, the pharmacological manipulation of the cannabinoid system might also serve to repair the injured tissue in these and other neurodegenerative disorders (reviewed in Galve-Roperh et al., 2007; Maccarrone et al., 2007), particularly given the recent identification of a cannabinoid-sensitive mechanism that participates in the control of adult neurogenesis (Aguado et al., 2005). A compound with this ability would mean that it complied with the aforementioned requirement of these molecules to preserve striatal or nigral neurons from death, permitting injured neurons to be replaced if treatments were initiated after degeneration had already begun, or if the treatment were only effective in delaying but not arresting the progression of the disease. Hence, cannabinoid compounds could be used to activate neurogenic structures (the subventricular zone and the hippocampal dentate gyrus in the adult brain), thereby encouraging the proliferation of cell progenitors, their differentiation to neurons and/or their migration to the damaged structures (striatum or substantia nigra) where they could acquire the phenotype corresponding to striatal medium-spiny neurons or nigrostriatal neurons that are lost in both diseases respectively. Unfortunately, the experimental evidence accumulated so far regarding the possible reparative actions of cannabinoids in both disorders is still minimal. Nevertheless, recent data have demonstrated the presence of a cannabinoid-sensitive population of progenitor cells in the subependymal layer of the normal and HD human brain (Curtis et al., 2006), possibly representing a suitable source for the replacement of the cells lost in this disease.
Concluding remarks and future perspectives
The studies reviewed here support the view that the cannabinoid signalling system is a key modulatory element in the activity of the basal ganglia. This concept is supported by different anatomical, electrophysiological, pharmacological and biochemical data. Indeed, we have shown that the cannabinoid system is impaired in different disorders that directly or indirectly affect the basal ganglia, which supports the idea of developing novel pharmacotherapies with compounds that selectively target specific elements of the cannabinoid system. The development of such compounds would not only provide novel therapeutic agents capable of minimizing the frequent side effects observed when classic cannabinoids are used in patients, but also help to elucidate the exact role played by the cannabinoid system in the pathogenesis of these disorders. In the light of the neuroprotectant/neuroregenerative properties demonstrated for certain cannabinoids, these treatments should not only aim to alleviate specific symptoms (e.g. hyperkinesia in HD, tremor and bradykinesia in PD), but also attempt to delay/arrest disease progression and to repair the damaged structures. However, most of the studies that have examined the therapeutic potential of these compounds in basal ganglia disorders have been conducted in animal models, with very few clinical trials carried out to date. Therefore, the importance of this intercellular signalling system needs further clinical research to be carried out in patients with the aim of validating the results found in animal and cellular models. Such studies should examine the potential of hybrid vanilloid/cannabinoid agonists to reduce hyperkinesia in HD patients. Another challenge in HD is to develop a broad-spectrum cannabinoid, or a combination of selective compounds, which might reduce excitotoxicity while limiting the activation of microglia and the generation of reactive oxygen intermediates. In this respect, given what is already known about the nature of their location and pharmacological properties, it will be important to establish in greater detail both the part that CB2 receptors play in HD and the potential therapeutic benefits of targeting them with a medicine. This receptor type might also be important in PD, although novel tools will have to be developed to determine whether this receptor is up-regulated or not in this disease (e.g. more selective anti-CB2 antibodies, selective ligands useful for in vivo analysis). In the absence of such information, it might be reasonable to test combinations of antioxidant cannabinoids as novel neuroprotectant therapies in patients. Alternatively, the capacity of CB1 and TRPV1 receptor antagonists to alleviate bradykinesia should be considered in patients who do not benefit from classical levodopa therapy.
Acknowledgments
The studies included in this review have been made possible by grants from MEC (SAF2006-11333), CIBERNED (CB06/05/0089) and CAM (S-SAL-0261/2006). The author is indebted to all authors that collaborated in these studies, to Patricia Rodríguez-Valsero and Yolanda García-Movellán for technical and administrative assistance and to Mark Sefton for language editing.
Glossary
Abbreviations
- CB1
cannabinoid receptor type 1
- CB2
cannabinoid receptor type 2
- FAAH
fatty acid amide hydrolase
- GABA
γ-aminobutiric acid
- HD
Huntington's disease
- PD
Parkinson's disease
- Δ9-THC
Δ9-tetrahydrocannabinol
- TRPV1
transient receptor potential vanilloid type 1
Conflict of interest
The author has not any conflict of interest. All agencies that supported the studies included in this review had no further role in study design, the collection, analysis and interpretation of data, in the writing of the report, or in the decision to submit the article for publication.
References
- Aguado T, Monory K, Palazuelos J, Stella N, Cravatt B, Lutz B, et al. The endocannabinoid system drives neural progenitor proliferation. FASEB J. 2005;19:1704–1706. doi: 10.1096/fj.05-3995fje. [DOI] [PubMed] [Google Scholar]
- Baker D, Jackson SJ, Pryce G. Cannabinoid control of neuroinflammation related to multiple sclerosis. Br J Pharmacol. 2007;152:649–654. doi: 10.1038/sj.bjp.0707458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battista N, Bari M, Tarditi A, Mariotti C, Bachoud-Lévi AC, Zuccato C, et al. Severe deficiency of the fatty acid amide hydrolase (FAAH) activity segregates with the Huntington's disease mutation in peripheral lymphocytes. Neurobiol Dis. 2007;27:108–116. doi: 10.1016/j.nbd.2007.04.012. [DOI] [PubMed] [Google Scholar]
- Beltramo M, Rodríguez de Fonseca F, Navarro M, Calignano A, Gorriti MA, Grammatikopoulos G, et al. Reversal of dopamine D2 receptor responses by an anandamide transport inhibitor. J Neurosci. 2000;20:3401–3407. doi: 10.1523/JNEUROSCI.20-09-03401.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisogno T, Berrendero F, Ambrosino G, Cebeira M, Ramos JA, Fernández-Ruiz J, et al. Brain regional distribution of endocannabinoids: implications for their biosynthesis and biological function. Biochem Biophys Res Commun. 1999;256:377–380. doi: 10.1006/bbrc.1999.0254. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Martire A, Petrosino S, Popoli P, Di Marzo V. Symptom-related changes of endocannabinoid and palmitoylethanolamide levels in brain areas of R6/2 mice, a transgenic model of Huntington's disease. Neurochem Int. 2008;52:307–313. doi: 10.1016/j.neuint.2007.06.031. [DOI] [PubMed] [Google Scholar]
- Brotchie JM. Adjuncts to dopamine replacement: a pragmatic approach to reducing the problem of dyskinesia in Parkinson's disease. Mov Disord. 1998;13:871–876. doi: 10.1002/mds.870130603. [DOI] [PubMed] [Google Scholar]
- Brotchie JM. The neural mechanisms underlying levodopa-induced dyskinesia in Parkinson's disease. Ann Neurol. 2000;47:S105–S114. [PubMed] [Google Scholar]
- Brotchie JM. CB1 cannabinoid receptor signalling in Parkinson's disease. Curr Opin Pharmacol. 2003;3:54–61. doi: 10.1016/s1471-4892(02)00011-5. [DOI] [PubMed] [Google Scholar]
- Campbell VA, Gowran A. Alzheimer's disease; taking the edge off with cannabinoids? Br J Pharmacol. 2007;152:655–662. doi: 10.1038/sj.bjp.0707446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Centonze D, Rossi S, Prosperetti C, Tscherter A, Bernardi G, Maccarrone M, et al. Abnormal sensitivity to cannabinoid receptor stimulation might contribute to altered gamma-aminobutyric acid transmission in the striatum of R6/2 Huntington's disease mice. Biol Psychiatry. 2005;57:1583–1589. doi: 10.1016/j.biopsych.2005.03.008. [DOI] [PubMed] [Google Scholar]
- Centonze D, Finazzi-Agro A, Bernardi G, Maccarrone M. The endocannabinoid system in targeting inflammatory neurodegenerative diseases. Trends Pharmacol Sci. 2007;28:180–187. doi: 10.1016/j.tips.2007.02.004. [DOI] [PubMed] [Google Scholar]
- Compton DR, Aceto MD, Lowe J, Martin BR. In vivo characterization of a specific cannabinoid antagonis (SR141716A): inhibition of Δ9-tetrahydrocannabinol-induced responses and apparent agonist activity. J Pharmacol Exp Ther. 1996;277:586–594. [PubMed] [Google Scholar]
- Consroe P. Brain cannabinoid systems as targets for the therapy of neurological disorders. Neurobiol Dis. 1998;5:534–551. doi: 10.1006/nbdi.1998.0220. [DOI] [PubMed] [Google Scholar]
- Crawley JN, Corwin RL, Robinson JK, ChC F, Devane WA, Axelrod J. Anandamide, an endogenous ligand of the cannabinoid receptor, induces hypomotility and hypothermia in vivo in rodents. Pharmacol Biochem Behav. 1993;46:967–972. doi: 10.1016/0091-3057(93)90230-q. [DOI] [PubMed] [Google Scholar]
- Cristino L, de Petrocellis L, Pryce G, Baker D, Guglielmotti V, Di Marzo V. Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neuroscience. 2006;139:1405–1415. doi: 10.1016/j.neuroscience.2006.02.074. [DOI] [PubMed] [Google Scholar]
- Curtis MA, Faull RL, Glass M. A novel population of progenitor cells expressing cannabinoid receptors in the subependymal layer of the adult normal and Huntington's disease human brain. J Chem Neuroanat. 2006;31:210–215. doi: 10.1016/j.jchemneu.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Denovan-Wright EM, Robertson HA. Cannabinoid receptor messenger RNA levels decrease in subset neurons of the lateral striatum, cortex and hippocampus of transgenic Huntington's disease mice. Neuroscience. 2000;98:705–713. doi: 10.1016/s0306-4522(00)00157-3. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Hill MP, Bisogno T, Crossman AR, Brotchie JM. Enhanced levels of endocannabinoids in the globus pallidus are associated with a reduction in movement in an animal model of Parkinson's disease. FASEB J. 2000;14:1432–1438. doi: 10.1096/fj.14.10.1432. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Lastres-Becker I, Bisogno T, De Petrocellis L, Milone A, Davis JB, et al. Hypolocomotor effects in rats of capsaicin and two long chain capsaicin homologues. Eur J Pharmacol. 2001;420:123–131. doi: 10.1016/s0014-2999(01)01012-3. [DOI] [PubMed] [Google Scholar]
- Dinh TP, Freund TF, Piomelli D. A role for monoglyceride lipase in 2-arachidonoylglycerol inactivation. Chem Phys Lipids. 2002;121:149–158. doi: 10.1016/s0009-3084(02)00150-0. [DOI] [PubMed] [Google Scholar]
- Egertova M, Cravatt BF, Elphick MR. Comparative analysis of fatty acid amide hydrolase and CB1 cannabinoid receptor expression in the mouse brain: evidence of a widespread role for fatty acid amide hydrolase in regulation of endocannabinoid signaling. Neuroscience. 2003;119:481–496. doi: 10.1016/s0306-4522(03)00145-3. [DOI] [PubMed] [Google Scholar]
- Fernández-Espejo E, Caraballo I, Rodriguez de Fonseca F, Ferrer B, El Banoua F, Flores JA, et al. Experimental parkinsonism alters anandamide precursor synthesis, and functional deficits are improved by AM404: a modulator of endocannabinoid function. Neuropsychopharmacology. 2004;29:1134–1142. doi: 10.1038/sj.npp.1300407. [DOI] [PubMed] [Google Scholar]
- Fernández-Espejo E, Caraballo I, de Fonseca FR, El Banoua F, Ferrer B, Flores JA, et al. Cannabinoid CB1 antagonists possess antiparkinsonian efficacy only in rats with very severe nigral lesion in experimental parkinsonism. Neurobiol Dis. 2005;18:591–601. doi: 10.1016/j.nbd.2004.10.015. [DOI] [PubMed] [Google Scholar]
- Fernández-Ruiz J, González S. Cannabinoid control of motor function at the basal ganglia. In: Pertwee RG, editor. Handbook of Experimental Pharmacology – 168 – Cannabinoids. Heidelberg: Springer-Verlag; 2005. pp. 479–507. [DOI] [PubMed] [Google Scholar]
- Fernández-Ruiz J, González S, Romero J, Ramos JA. Cannabinoids in neurodegeneration and neuroprotection. In: Mechoulam R, editor. Cannabinoids as Therapeutics (MDT) Basel: Birkhaüser Verlag; 2005. pp. 79–109. [Google Scholar]
- Fernández-Ruiz J, Romero J, Velasco G, Tolón RM, Ramos JA, Guzmán M. Cannabinoid CB2 receptor: a new target for the control of neural cell survival? Trends Pharmacol Sci. 2007;28:39–45. doi: 10.1016/j.tips.2006.11.001. [DOI] [PubMed] [Google Scholar]
- Fernández-Ruiz J, Pazos MR, García-Arencibia M, Sagredo O, Ramos JA. Role of CB2 receptors in neuroprotective effects of cannabinoids. Mol Cell Endocrinol. 2008;286:S91–S96. doi: 10.1016/j.mce.2008.01.001. [DOI] [PubMed] [Google Scholar]
- Fox SH, Kellett M, Moore AP, Crossman AR, Brotchie JM. Randomised, double-blind, placebo-controlled trial to assess the potential of cannabinoid receptor stimulation in the treatment of dystonia. Mov Disord. 2002a;17:145–149. doi: 10.1002/mds.1280. [DOI] [PubMed] [Google Scholar]
- Fox SH, Henry B, Hill M, Crossman A, Brotchie J. Stimulation of cannabinoid receptors reduces levodopa-induced dyskinesia in the MPTP-lesioned nonhuman primate model of Parkinson's disease. Mov Disord. 2002b;17:1180–1187. doi: 10.1002/mds.10289. [DOI] [PubMed] [Google Scholar]
- Frankel JP, Hughes A, Lees AJ, Stern GM. Marijuana for parkinsonian tremor. J Neurol Neurosurg Psychiatry. 1990;53:436–436. doi: 10.1136/jnnp.53.5.436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fride E, Mechoulam R. Pharmacological activity of the cannabinoid receptor agonist, anandamide, a brain constituent. Eur J Pharmacol. 1993;231:313–314. doi: 10.1016/0014-2999(93)90468-w. [DOI] [PubMed] [Google Scholar]
- Fusco FR, Martorana A, Giampa C, De March Z, Farini D, D'Angelo V, et al. Immunolocalization of CB1 receptor in rat striatal neurons: a confocal microscopy study. Synapse. 2004;53:159–167. doi: 10.1002/syn.20047. [DOI] [PubMed] [Google Scholar]
- Galve-Roperh I, Aguado T, Palazuelos J, Guzman M. The endocannabinoid system and neurogenesis in health and disease. Neuroscientist. 2007;13:109–114. doi: 10.1177/1073858406296407. [DOI] [PubMed] [Google Scholar]
- García-Arencibia M, González S, de Lago E, Ramos JA, Mechoulam R, Fernández-Ruiz J. Evaluation of the neuroprotective effect of cannabinoids in a rat model of Parkinson's disease: importance of antioxidant and cannabinoid receptor-independent properties. Brain Res. 2007;1134:162–170. doi: 10.1016/j.brainres.2006.11.063. [DOI] [PubMed] [Google Scholar]
- García-Arencibia M, García C, Kurz A, Rodríguez-Navarro JA, Gispert-Sánchez S, Mena MA, et al. Cannabinoid CB1 receptors are early down-regulated followed by a further up-regulation in the basal ganglia of mice with deletion of specific PARK genes. J Neural Transm. 2008a doi: 10.1007/978-3-211-92660-4_22. in press. [DOI] [PubMed] [Google Scholar]
- García-Arencibia M, Ferraro L, Tanganelli S, Fernández-Ruiz J. Enhanced striatal glutamate release after the administration of rimonabant to 6-hydroxydopamine-lesioned rats. Neurosci Lett. 2008b;438:10–13. doi: 10.1016/j.neulet.2008.04.041. [DOI] [PubMed] [Google Scholar]
- Gerdeman GL, Ronesi J, Lovinger DM. Postsynaptic endocannabinoid release is critical to long-term depression in the striatum. Nat Neurosci. 2002;5:446–451. doi: 10.1038/nn832. [DOI] [PubMed] [Google Scholar]
- Gerdeman GL, Fernández-Ruiz J. The endocannabinoid system in the physiology and pathophysiology of the basal ganglia. In: Kofalvi A, editor. Cannabinoids and the Brain. New York, NY: Springer-Verlag; 2008. pp. 423–483. [Google Scholar]
- Giuffrida A, Parsons LH, Kerr TM, Rodríguez de Fonseca F, Navarro M, Piomelli D. Dopamine activation of endogenous cannabinoid signaling in dorsal striatum. Nat Neurosci. 1999;2:358–363. doi: 10.1038/7268. [DOI] [PubMed] [Google Scholar]
- Glass M, Faull RLM, Dragunow M. Loss of cannabinoid receptors in the substantia nigra in Huntington's disease. Neuroscience. 1993;56:523–527. doi: 10.1016/0306-4522(93)90352-g. [DOI] [PubMed] [Google Scholar]
- Glass M, Dragunow M, Faull RLM. The pattern of neurodegeneration in Huntington's disease: a comparative study of cannabinoid, dopamine, adenosine and GABA-A receptor alterations in the human basal ganglia in Huntington's disease. Neuroscience. 2000;97:505–519. doi: 10.1016/s0306-4522(00)00008-7. [DOI] [PubMed] [Google Scholar]
- González S, Romero J, de Miguel R, Lastres-Becker I, Villanúa MA, Makriyannis A, et al. Extrapyramidal and neuroendocrine effects of AM404, an inhibitor of the carrier-mediated transport of anandamide. Life Sci. 1999;65:327–336. doi: 10.1016/s0024-3205(99)00251-9. [DOI] [PubMed] [Google Scholar]
- González S, Scorticati C, Garcia-Arencibia M, de Miguel R, Ramos JA, Fernandez-Ruiz J. Effects of rimonabant, a selective cannabinoid CB1 receptor antagonist, in a rat model of Parkinson's disease. Brain Res. 2006;1073(1074):209–219. doi: 10.1016/j.brainres.2005.12.014. [DOI] [PubMed] [Google Scholar]
- Gubellini P, Picconi B, Bari M, Battista N, Calabresi P, Centonze D, et al. Experimental parkinsonism alters endocannabinoid degradation: implications for striatal glutamatergic transmission. J Neurosci. 2002;22:6900–6907. doi: 10.1523/JNEUROSCI.22-16-06900.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, Little MD, Melvin LS, Johnson MR, de Costa DR, et al. Characterization and localization of cannabinoid receptors in rat brain: a quantitative in vitro autoradiographic study. J Neurosci. 1991a;11:563–583. doi: 10.1523/JNEUROSCI.11-02-00563.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, de Costa BR, Richfield EK. Neuronal localization of cannabinoid receptors in the basal ganglia of the rat. Brain Res. 1991b;547:267–264. doi: 10.1016/0006-8993(91)90970-7. [DOI] [PubMed] [Google Scholar]
- Hohmann AG, Herkenham M. Localization of cannabinoid CB1 receptor mRNA in neuronal subpopulations of rat striatum: a double-label in situ hybridization study. Synapse. 2000;37:71–80. doi: 10.1002/(SICI)1098-2396(200007)37:1<71::AID-SYN8>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- Huang CC, Lo SW, Hsu KS. Presynaptic mechanisms underlying cannabinoid inhibition of excitatory synaptic transmission in rat striatal neurons. J Physiol. 2001;532:731–748. doi: 10.1111/j.1469-7793.2001.0731e.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarbe TU, Sheppard R, Lamb RJ, Makriyannis A, Lin S, Goutopoulos A. Effects of Δ9-tetrahydrocannabinol and (R)-methanandamide on open-field behavior in rats. Behav Pharmacol. 1998;9:169–174. [PubMed] [Google Scholar]
- Jiménez-del Río M, Daza-Restrepo A, Velez-Pardo C. The cannabinoid CP55,940 prolongs survival and improves locomotor activity in Drosophila melanogaster against paraquat: implications in Parkinson's disease. Neurosci Res. 2008;61:404–411. doi: 10.1016/j.neures.2008.04.011. [DOI] [PubMed] [Google Scholar]
- Julián MD, Martin AB, Cuellar B, Rodriguez de Fonseca F, Navarro M, Moratalla R, et al. Neuroanatomical relationship between type 1 cannabinoid receptors and dopaminergic systems in the rat basal ganglia. Neuroscience. 2003;119:309–318. doi: 10.1016/s0306-4522(03)00070-8. [DOI] [PubMed] [Google Scholar]
- Köfalvi A, Rodrigues RJ, Ledent C, Mackie K, Vizi ES, Cunha RA, et al. Involvement of cannabinoid receptors in the regulation of neurotransmitter release in the rodent striatum: a combined immunochemical and pharmacological analysis. J Neurosci. 2005;25:2874–2884. doi: 10.1523/JNEUROSCI.4232-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreitzer AC, Malenka RC. Endocannabinoid-mediated rescue of striatal LTD and motor deficits in Parkinson's disease models. Nature. 2007;445:643–647. doi: 10.1038/nature05506. [DOI] [PubMed] [Google Scholar]
- de Lago E, Fernández-Ruiz J, Ortega-Gutierrez S, Viso A, Lopez-Rodríguez ML, Ramos JA. UCM707, a potent and selective inhibitor of endocannabinoid uptake, potentiates hypokinetic and antinociceptive effects of anandamide. Eur J Pharmacol. 2002;449:99–103. doi: 10.1016/s0014-2999(02)01996-9. [DOI] [PubMed] [Google Scholar]
- de Lago E, de Miguel R, Lastres-Becker I, Ramos JA, Fernández-Ruiz J. Involvement of vanilloid-like receptors in the effects of anandamide on motor behavior and nigrostriatal dopaminergic activity: in vivo and in vitro evidence. Brain Res. 2004a;1007:152–159. doi: 10.1016/j.brainres.2004.02.016. [DOI] [PubMed] [Google Scholar]
- de Lago E, Ligresti A, Ortar G, Morera E, Cabranes A, Pryce G, et al. In vivo pharmacological actions of two novel inhibitors of anandamide cellular uptake. Eur J Pharmacol. 2004b;484:249–257. doi: 10.1016/j.ejphar.2003.11.027. [DOI] [PubMed] [Google Scholar]
- de Lago E, Urbani P, Ramos JA, Di Marzo V, Fernández-Ruiz J. Arvanil, a hybrid endocannabinoid and vanilloid compound, behaves as an antihyperkinetic agent in a rat model of Huntington's disease. Brain Res. 2005;1050:210–216. doi: 10.1016/j.brainres.2005.05.024. [DOI] [PubMed] [Google Scholar]
- de Lago E, Fernández-Ruiz J, Ortega-Gutierrez S, Cabranes A, Pryce G, Baker D, et al. UCM707, an inhibitor of the anandamide uptake, behaves as a symptom control agent in models of Huntington's disease and multiple sclerosis, but fails to delay/arrest the progression of different motor-related disorders. Eur Neuropsychopharmacol. 2006;16:7–18. doi: 10.1016/j.euroneuro.2005.06.001. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Cebeira M, de Ceballos M, Zeng B-Y, Jenner P, Ramos JA, et al. Increased cannabinoid CB1 receptor binding and activation of GTP-binding proteins in the basal ganglia of patients with Parkinson's syndrome and of MPTP-treated marmosets. Eur J Neurosci. 2001a;14:1827–1832. doi: 10.1046/j.0953-816x.2001.01812.x. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Fezza F, Cebeira M, Bisogno T, Ramos JA, Milone A, et al. Changes in endocannabinoid transmission in the basal ganglia in a rat model of Huntington's disease. Neuroreport. 2001b;12:2125–2129. doi: 10.1097/00001756-200107200-00017. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Hansen HH, Berrendero F, de Miguel R, Pérez-Rosado A, Manzanares J, et al. Alleviation of motor hyperactivity and neurochemical deficits by endocannabinoid uptake inhibition in a rat model of Huntington's disease. Synapse. 2002a;44:23–35. doi: 10.1002/syn.10054. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Gomez M, De Miguel R, Ramos JA, Fernández-Ruiz J. Loss of cannabinoid CB1 receptors in the basal ganglia in the late akinetic phase of rats with experimental Huntington's disease. Neurotox Res. 2002b;4:601–608. doi: 10.1080/10298420290030514. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Berrendero F, Lucas JJ, Martín-Aparicio E, Yamamoto A, Ramos JA, et al. Loss of mRNA levels, binding and activation of GTP-binding proteins for cannabinoid CB1 receptors in the basal ganglia of a transgenic model of Huntington's disease. Brain Res. 2002c;929:236–242. doi: 10.1016/s0006-8993(01)03403-5. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, de Miguel R, De Petrocellis L, Makriyannis A, Di Marzo V, Fernández-Ruiz J. Compounds acting at the endocannabinoid and/or endovanilloid systems reduce hyperkinesia in a rat model of Huntington's disease. J Neurochem. 2003a;84:1097–1109. doi: 10.1046/j.1471-4159.2003.01595.x. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, De Miguel R, Fernández-Ruiz J. The endocannabinoid system and Huntington's disease. Curr Drug Target CNS Neurol Disord. 2003b;2:335–347. doi: 10.2174/1568007033482751. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Bizat N, Boyer F, Hantraye P, Fernández-Ruiz J, Brouillet E. Potential involvement of cannabinoid receptors in 3-nitropropionic acid toxicity in vivo. Neuroreport. 2004;15:2375–2379. doi: 10.1097/00001756-200410250-00015. [DOI] [PubMed] [Google Scholar]
- Lastres-Becker I, Molina-Holgado F, Ramos JA, Mechoulam R, Fernández-Ruiz J. Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: relevance to Parkinson's disease. Neurobiol Dis. 2005;19:96–107. doi: 10.1016/j.nbd.2004.11.009. [DOI] [PubMed] [Google Scholar]
- Ledent C, Valverde O, Cossu G, Petitet F, Aubert JF, Beslot F, et al. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science. 1999;283:401–404. doi: 10.1126/science.283.5400.401. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Gubellini P, Bari M, Picconi B, Battista N, Centonze D, et al. Levodopa treatment reverses endocannabinoid system abnormalities in experimental parkinsonism. J Neurochem. 2003;85:1018–1025. doi: 10.1046/j.1471-4159.2003.01759.x. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Battista N, Centonze D. The endocannabinoid pathway in Huntington's disease: a comparison with other neurodegenerative diseases. Prog Neurobiol. 2007;81:349–379. doi: 10.1016/j.pneurobio.2006.11.006. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Rossi S, Bari M, De Chiara V, Fezza F, Musella A, et al. Anandamide inhibits metabolism and physiological actions of 2-arachidonoylglycerol in the striatum. Nat Neurosci. 2008;11:152–159. doi: 10.1038/nn2042. [DOI] [PubMed] [Google Scholar]
- McCaw EA, Hu H, Gomez GT, Hebb AL, Kelly ME, Denovan-Wright EM. Structure, expression and regulation of the cannabinoid receptor gene (CB1) in Huntington's disease transgenic mice. Eur J Biochem. 2004;271:4909–4920. doi: 10.1111/j.1432-1033.2004.04460.x. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Delevan CE, Carnicom S, Robinson JK, Brener J. Fine motor control in rats is disrupted by Δ9-tetrahydrocannabinol. Pharmacol Biochem Behav. 2000;66:803–809. doi: 10.1016/s0091-3057(00)00281-1. [DOI] [PubMed] [Google Scholar]
- Mailleux P, Vanderhaeghen JJ. Distribution of neuronal cannabinoid receptor in the adult rat brain: a comparative receptor binding radioautography and in situ hybridization histochemistry. Neuroscience. 1992;48:655–668. doi: 10.1016/0306-4522(92)90409-u. [DOI] [PubMed] [Google Scholar]
- Marsicano G, Moosmann B, Hermann H, Lutz B, Behl C. Neuroprotective properties of cannabinoids against oxidative stress: role of the cannabinoid receptor CB1. J Neurochem. 2002;80:448–456. doi: 10.1046/j.0022-3042.2001.00716.x. [DOI] [PubMed] [Google Scholar]
- Meschler JP, Howlett AC. Signal transduction interactions between CB1 cannabinoid and dopamine receptors in the rat and monkey striatum. Neuropharmacology. 2001;40:918–926. doi: 10.1016/s0028-3908(01)00012-0. [DOI] [PubMed] [Google Scholar]
- Meschler JP, Conley TJ, Howlett AC. Cannabinoid and dopamine interaction in rodent brain: effects on locomotor activity. Pharmacol Biochem Behav. 2000;67:567–573. doi: 10.1016/s0091-3057(00)00390-7. [DOI] [PubMed] [Google Scholar]
- Meschler JP, Howlett AC, Madras BK. Cannabinoid receptor agonist and antagonist effects on motor function in normal and 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine (MPTP)-treated non-human primates. Psychopharmacology. 2001;156:79–85. doi: 10.1007/s002130100728. [DOI] [PubMed] [Google Scholar]
- Mesnage V, Houeto JL, Bonnet AM, Clavier I, Arnulf I, Cattelin F, et al. Neurokinin B, neurotensin, and cannabinoid receptor antagonists and Parkinson's disease. Clin Neuropharmacol. 2004;27:108–110. doi: 10.1097/00002826-200405000-00003. [DOI] [PubMed] [Google Scholar]
- Mezey E, Toth ZE, Cortright DN, Arzubi MK, Krause JE, Elde R, et al. Distribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and human. Proc Natl Acad Sci USA. 2000;97:3655–3660. doi: 10.1073/pnas.060496197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller-Vahl KR. Cannabinoids reduce symptoms of Tourette's syndrome. Expert Opin Pharmacother. 2003;4:1717–1725. doi: 10.1517/14656566.4.10.1717. [DOI] [PubMed] [Google Scholar]
- Müller-Vahl KR, Schneider U, Emrich HM. Nabilone increases choreatic movements in Huntington's disease. Mov Disord. 1999;14:1038–1040. doi: 10.1002/1531-8257(199911)14:6<1038::aid-mds1024>3.0.co;2-7. [DOI] [PubMed] [Google Scholar]
- Naver B, Stub C, Moller M, Fenger K, Hansen AK, Hasholt L, et al. Molecular and behavioral analysis of the R6/1 Huntington's disease transgenic mouse. Neuroscience. 2003;122:1049–1057. doi: 10.1016/j.neuroscience.2003.08.053. [DOI] [PubMed] [Google Scholar]
- Page KJ, Besret L, Jain M, Monaghan EM, Dunnett SB, Everitt BJ. Effects of systemic 3-nitropropionic acid-induced lesions of the dorsal striatum on cannabinoid and mu-opioid receptor binding in the basal ganglia. Exp Brain Res. 2000;130:142–150. doi: 10.1007/s002210050016. [DOI] [PubMed] [Google Scholar]
- Pazos MR, Sagredo O, Fernández-Ruiz J. The endocannabinoid system in Huntington's disease. Curr Pharm Des. 2008;14:2317–2325. doi: 10.2174/138161208785740108. [DOI] [PubMed] [Google Scholar]
- Pintor A, Tebano MT, Martire A, Grieco R, Galluzzo M, Scattoni ML, et al. The cannabinoid receptor agonist WIN 55,212-2 attenuates the effects induced by quinolinic acid in the rat striatum. Neuropharmacology. 2006;51:1004–1012. doi: 10.1016/j.neuropharm.2006.06.013. [DOI] [PubMed] [Google Scholar]
- Pisani A, Fezza F, Galati S, Battista N, Napolitano S, Finazzi-Agro A, et al. High endogenous cannabinoid levels in the cerebrospinal fluid of untreated Parkinson's disease patients. Ann Neurol. 2005;57:777–779. doi: 10.1002/ana.20462. [DOI] [PubMed] [Google Scholar]
- Richfield EK, Herkenham M. Selective vulnerability in Huntington's disease: preferential loss of cannabinoid receptors in lateral globus pallidus. Ann Neurol. 1994;36:577–584. doi: 10.1002/ana.410360406. [DOI] [PubMed] [Google Scholar]
- Richter A, Loscher W. Effects of pharmacological manipulations of cannabinoid receptors on severity of dystonia in a genetic model of paroxysmal dyskinesia. Eur J Pharmacol. 2002;454:145–151. doi: 10.1016/s0014-2999(02)02477-9. [DOI] [PubMed] [Google Scholar]
- Romero J, García L, Cebeira M, Zadrozny D, Fernández-Ruiz J, Ramos JA. The endogenous cannabinoid receptor ligand, anandamide, inhibits the motor behaviour: role of nigrostriatal dopaminergic neurons. Life Sci. 1995a;56:2033–2040. doi: 10.1016/0024-3205(95)00186-a. [DOI] [PubMed] [Google Scholar]
- Romero J, de Miguel R, García-Palomero E, Fernández-Ruiz J, Ramos JA. Time-course of the effects of anandamide, the putative endogenous cannabinoid receptor ligand, on extrapyramidal function. Brain Res. 1995b;694:223–232. doi: 10.1016/0006-8993(95)00835-e. [DOI] [PubMed] [Google Scholar]
- Romero J, García-Palomero E, Fernández-Ruiz J, Ramos JA. Involvement of GABA(B) receptors in the motor inhibition produced by agonists of brain cannabinoid receptors. Behav Pharmacol. 1996;7:299–302. doi: 10.1097/00008877-199605000-00011. [DOI] [PubMed] [Google Scholar]
- Romero J, Berrendero F, Pérez-Rosado A, Manzanares J, Rojo A, Fernández-Ruiz J, et al. Unilateral 6-hydroxydopamine lesions of nigrostriatal dopaminergic neurons increased CB1 receptor mRNA levels in the caudate-putamen. Life Sci. 2000;66:485–494. doi: 10.1016/s0024-3205(99)00618-9. [DOI] [PubMed] [Google Scholar]
- Romero J, Lastres-Becker I, de Miguel R, Berrendero F, Ramos JA, Fernández-Ruiz J. The endogenous cannabinoid system and the basal ganglia. biochemical, pharmacological, and therapeutic aspects. Pharm Ther. 2002;95:137–152. doi: 10.1016/s0163-7258(02)00253-x. [DOI] [PubMed] [Google Scholar]
- Roze E, Saudou F, Caboche J. Pathophysiology of Huntington's disease: from huntingtin functions to potential treatments. Curr Opin Neurol. 2008;21:497–503. doi: 10.1097/WCO.0b013e328304b692. [DOI] [PubMed] [Google Scholar]
- Russo E, Guy GW. A tale of two cannabinoids: the therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med Hypotheses. 2006;66:234–246. doi: 10.1016/j.mehy.2005.08.026. [DOI] [PubMed] [Google Scholar]
- Sagredo O, García-Arencibia M, de Lago E, Finetti S, Decio A, Fernández-Ruiz J. Cannabinoids and neuroprotection in basal ganglia disorders. Mol Neurobiol. 2007a;36:82–91. doi: 10.1007/s12035-007-0004-3. [DOI] [PubMed] [Google Scholar]
- Sagredo O, Ramos JA, Decio A, Mechoulam R, Fernández-Ruiz J. Cannabidiol reduced the striatal atrophy caused 3-nitropropionic acid in vivo by mechanisms independent of the activation of cannabinoid receptors. Eur J Neurosci. 2007b;26:843–851. doi: 10.1111/j.1460-9568.2007.05717.x. [DOI] [PubMed] [Google Scholar]
- Sagredo O, González S, Aroyo I, Pazos MR, Benito C, Lastres-Becker I, et al. Cannabinoid CB2 receptor agonists protect the striatum against malonate toxicity by limiting the influence of reactive microglia on neuronal homeostasis: relevance for Huntington's disease. Glia. 2009 doi: 10.1002/glia.20838. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sañudo-Peña MC, Patrick SL, Khen S, Patrick RL, Tsou K, Walker JM. Cannabinoid effects in basal ganglia in a rat model of Parkinson's disease. Neurosci Lett. 1998;248:171–174. doi: 10.1016/s0304-3940(98)00368-1. [DOI] [PubMed] [Google Scholar]
- Sañudo-Peña MC, Tsou K, Walker JM. Motor actions of cannabinoids in the basal ganglia output nuclei. Life Sci. 1999;65:703–713. doi: 10.1016/s0024-3205(99)00293-3. [DOI] [PubMed] [Google Scholar]
- Sañudo-Peña MC, Romero J, Seale GE, Fernández-Ruiz J, Walker JM. Activational role of cannabinoids on movement. Eur J Pharmacol. 2000;391:269–274. doi: 10.1016/s0014-2999(00)00044-3. [DOI] [PubMed] [Google Scholar]
- Sheng WS, Hu S, Min X, Cabral GA, Lokensgard JR, Peterson PK. Synthetic cannabinoid WIN55,212-2 inhibits generation of inflammatory mediators by IL-1β-stimulated human astrocytes. Glia. 2005;49:211–219. doi: 10.1002/glia.20108. [DOI] [PubMed] [Google Scholar]
- Sieradzan KA, Fox SH, Hill M, Dick JP, Crossman AR, Brotchie JM. Cannabinoids reduce levodopa-induced dyskinesia in Parkinson's disease: a pilot study. Neurology. 2001;57:2108–2111. doi: 10.1212/wnl.57.11.2108. [DOI] [PubMed] [Google Scholar]
- Souilhac J, Poncelet M, Rinaldi-Carmona M, Le-Fur G, Soubrie P. Intrastriatal injection of cannabinoid receptor agonists induced turning behavior in mice. Pharmacol Biochem Behav. 1995;51:3–7. doi: 10.1016/0091-3057(94)00396-z. [DOI] [PubMed] [Google Scholar]
- Starowicz K, Nigam S, Di Marzo V. Biochemistry and pharmacology of endovanilloids. Pharmacol Ther. 2007;114:13–33. doi: 10.1016/j.pharmthera.2007.01.005. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Di Marzo V. Cannabinoid receptors and their role in neuroprotection. Neuromolecular Med. 2005;7:37–50. doi: 10.1385/NMM:7:1-2:037. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Veldhuis WB, Maccarrone M, Bär PR, Nicolay K, Veldink GA, et al. Acute neuronal injury, excitotoxicity, and the endocannabinoid system. Mol Neurobiol. 2002;26:317–346. doi: 10.1385/MN:26:2-3:317. [DOI] [PubMed] [Google Scholar]
- van der Stelt M, Fox SH, Hill M, Crossman AR, Petrosino S, Di Marzo V, et al. A role for endocannabinoids in the generation of parkinsonism and levodopa-induced dyskinesia in MPTP-lesioned non-human primate models of Parkinson's disease. FASEB J. 2005;19:1140–1142. doi: 10.1096/fj.04-3010fje. [DOI] [PubMed] [Google Scholar]
- Thomas B, Beal MF. Parkinson's disease. Hum Mol Genet. 2007;16:R183–R194. doi: 10.1093/hmg/ddm159. [DOI] [PubMed] [Google Scholar]
- Tsou K, Brown S, Sañudo-Peña MC, Mackie K, Walker JM. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience. 1998a;83:393–411. doi: 10.1016/s0306-4522(97)00436-3. [DOI] [PubMed] [Google Scholar]
- Tsou K, Nogueron MI, Muthian S, Sañudo-Peña M, Hillard CJ, Deutsch DG, et al. Fatty acid amide hydrolase is located preferentially in large neurons in the rat central nervous system as revealed by immunohistochemistry. Neurosci Lett. 1998b;254:137–140. doi: 10.1016/s0304-3940(98)00700-9. [DOI] [PubMed] [Google Scholar]
- Tzavara ET, Li DL, Moutsimilli L, Bisogno T, Di Marzo V, Phebus LA, et al. Endocannabinoids activate transient receptor potential vanilloid 1 receptors to reduce hyperdopaminergia-related hyperactivity: therapeutic implications. Biol Psychiatry. 2006;59:508–515. doi: 10.1016/j.biopsych.2005.08.019. [DOI] [PubMed] [Google Scholar]
- Uchigashima M, Narushima M, Fukaya M, Katona I, Kano M, Watanabe M. Subcellular arrangement of molecules for 2-arachidonoyl-glycerol-mediated retrograde signaling and its physiological contribution to synaptic modulation in the striatum. J Neurosci. 2007;27:3663–3676. doi: 10.1523/JNEUROSCI.0448-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallmichrath I, Szabo B. Cannabinoids inhibit striatonigral GABAergic neurotransmission in the mouse. Neuroscience. 2002;113:671–682. doi: 10.1016/s0306-4522(02)00109-4. [DOI] [PubMed] [Google Scholar]
- Wickens AP, Pertwee RG. Δ9-Tetrahydrocannabinol and anandamide enhance the ability of muscimol to induce catalepsy in the globus pallidus of rats. Eur J Pharmacol. 1993;250:205–208. doi: 10.1016/0014-2999(93)90646-y. [DOI] [PubMed] [Google Scholar]
- Zeng BY, Dass B, Owen A, Rose S, Cannizzaro C, Tel BC, et al. Chronic L-DOPA treatment increases striatal cannabinoid CB1 receptor mRNA expression in 6-hydroxydopamine-lesioned rats. Neurosci Lett. 1999;276:71–74. doi: 10.1016/s0304-3940(99)00762-4. [DOI] [PubMed] [Google Scholar]