

Abstract
CD74 is a protein whose initial role in antigen presentation was recognized two decades ago. Recent studies have revealed that it has additional functions as a receptor for macrophage migration inhibitory factor and as a receptor for an important human pathogen, Helicobacter pylori (H pylori). The role of CD74 as a receptor is important because after binding of migration inhibitory factor or H pylori, NF-κB and Erk1/2 activation occurs, along with the induction of proinflammatory cytokine secretion. This review provides an up-to-date account of the functions of CD74 and how it might be involved in inflammation and cancer within the gastrointestinal tract.
Keywords: CD74, Invariant chain, Cancer, Inflammation, Helicobacter pylori, Gastrointestinal tract
INTRODUCTION
CD74, also known as the invariant chain or Ii, is a non-polymorphic glycoprotein that has diverse immunological functions. The most well-known function of CD74 is regulating the trafficking of class II major histocompatibility complex (MHC) proteins in antigen presenting cells. More recently, CD74 expression has been examined in cell types other than antigen presenting cells (APCs), such as epithelial cells[1]. Some studies also suggest that CD74 might be expressed independently of class II MHC, indicating additional functions[2]. Various studies have indicated that CD74 is highly expressed in inflammatory disorders and cancers. It also acts as a receptor for macrophage migration inhibitory factor (MIF) and facilitates adhesion of Helicobacter pylori (H pylori) to gastric epithelial cells (GECs)[3,4]. CD74 expression is increased during H pylori infection, chronic inflammatory conditions of the gastrointestinal (GI) tract, and gastric and colon cancers. One critical function it has in carcinogenesis is to act as an accessory signaling molecule for cell proliferation. This molecule is particularly important in the complex immunological mechanisms of the gastrointestinal tract and in the link between chronic inflammation and carcinogenesis in the GI tract.
THE ROLE OF CD74 IN ANTIGEN PRESENTATION
CD74 was initially characterized for its role in regulating class II MHC folding and intracellular sorting and has been studied in most detail in APCs. Newly synthesized CD74 self-assembles as a trimer and this trimer serves as a scaffold onto which class II MHC molecules assemble. CD74 blocks the peptide binding cleft of class II MHC and prevents premature binding of antigenic peptides. Upon endocytosis of antigens, CD74 directs the class II MHC complex to the endosomal pathway using two di-leucine-based signals[5]. Within an endosomal compartment CD74 is digested, leaving just one residual peptide, CLIP (amino acids 91-99), blocking the peptide binding cleft of class II MHC. A class II MHC-like molecule, HLA-DM, then binds to the class II MHC and CLIP is released leaving the peptide binding cleft open for antigenic peptide binding. Class II MHC molecules with bound peptides are then exported to the surface of the antigen presenting cell (APC) for presentation of foreign peptides to T cells. CD74 plays a crucial role in antigen presentation, as class II MHC processing and regulation cannot properly occur in the absence of CD74.
In addition to APCs, other cells of the gastrointestinal tract, such as epithelial cells, express class II MHC proteins and CD74 and act as APCs, which is an unusual trait of the GI tract. We have previously shown that gastric epithelial cells express class II MHC proteins and are capable of processing antigens for presentation to T cells[6,7]. Expression of class II MHC and the potential for antigen presentation have also been shown in intestinal epithelial cells. In one elaborate study by Hershberg et al, the trafficking of these molecules in epithelial cells was followed in a polarized manner outlining a functional system for antigen presentation[8]. Another important group of cells that express class II MHC proteins are the subepithelial intestinal myofibroblasts (IMF)[9,10]. These cells are α-smooth actin positive stromal cells that exist in the lamina propria of the gut[11]. They also act as antigen presenting cells and play an important role in inflammatory diseases and carcinogenesis by releasing cytokines and growth factors and interacting with the immune cells of the lamina propria.
In conventional APCs, CD74 surface expression is low as CD74 is proteolytically removed in endosomes, as we and others[12,13] have shown. However, gastric epithelial cells express readily detectable levels of CD74 on the surface. When we examined human gastric biopsy sections by immunohistochemistry for epithelial expression of CD74, gastric biopsy samples from 44 random patients stained with anti-CD74 monoclonal antibodies (mAb) showed the expression of CD74 in the epithelium. In the samples that were positive for H pylori or had gastritis, CD74 staining was heavily increased[14]. This was corroborated by confocal microscopy studies of gastric epithelial cells grown as a polarized monolayer where expression was higher on the apical side of the cells[1].
CD74 ISOFORMS
CD74 is post-translationally glycosylated and exists in different isoforms. As evidence for this, our previous studies showed proteins with different mobilities when immunoprecipitated and subjected to gel electrophoresis[1]. After chemical deglycosylation, only the isoforms that result from alternative translation initiation or splicing were observed. The most common isoform is 33 kDa (p33), but p35, p41 and p43 isoforms also exist[15]. The p35 isoform contains a longer cytoplasmic tail due to the use of an alternative translation initiation site, while the p41 isoform results from alternative splicing, and p43 has both. Both the p33 and p35 isoforms appear to function in regulating class II MHC antigen presentation. However, the p41 isoform might also play an important role in T cell selection in the thymus[16]. An important posttranslational modification of CD74 commonly seen is the addition of a glycosaminoglycan side chain, chondroitin sulfate. This isoform has been reported to act as an accessory protein during T cell activation through interaction with CD44 on T cells[17].
CD74 AS A RECEPTOR
CD74 has begun to emerge as a more versatile molecule beyond its well-known function of regulating class II MHC trafficking. Multiple studies have revealed that cell surface expression of CD74 is not always dependent on class II MHC[2,18]. This was found to be true in studies of colorectal mucosa and different types of lymphocytes by immunohistochemistry, immunoprecipitation, and a mutant cell line that did not express class II MHC products. The expression of cell surface CD74 in the absence of class II MHC suggests alternative functions for CD74 apart from antigen presentation.
CD74 as a cytokine receptor
Recently, CD74 has emerged as an integral component of a receptor complex for macrophage MIF[3]. MIF is a versatile cytokine-like molecule that mediates both innate and adaptive immunity, plays a role in chronic inflammation, and has also been linked to carcinogenesis. MIF is expressed by the GI tract during inflammatory conditions, H pylori infection, and cancers, suggesting the importance of this interaction[19–21]. H pylori utilizes a type IV secretion system to inject the CagA protein into GECs, which we have shown induces MIF production[19] (Figure 1). CagA has also been shown to induce NF-κB activation and IL-8 production[22]. This protein is not only important in inflammation by increasing proinflammatory cytokine production, such as IL-8, but is also associated with gastric cancer[23]. The current model of this receptor complex suggests that CD44 is required in order for MIF to induce signaling events[24]. This model might require the chondroitin sulfate modified isoform of CD74, since CD44 has been shown to bind only to this isoform thus far.
Figure 1.
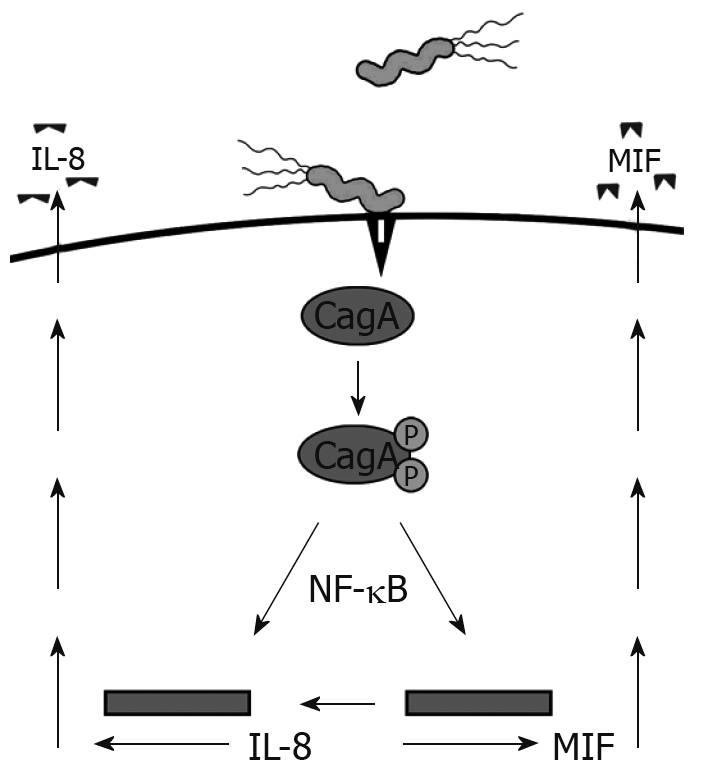
H pylori induces MIF and IL-8 production by injecting CagA into GECs via a type IV secretion system.
Another study has suggested that CD74 complexes with CXCR2, an interleukin-8 (IL-8) receptor, which is commonly expressed on macrophages and functions to recruit leukocytes to sites of infection[25]. CXCR2 is also expressed by the gastric epithelium[26]. Since gastric epithelial cells are central players in the inflammatory response, IL-8 may act via the gastric epithelium in various processes associated with gastric inflammation linked to H pylori infection. The role of CXCR2 on the CD74 receptor complex has only recently been suggested and should be further investigated.
CD74 as a bacterial receptor
CD74 is an interesting example of a host cell receptor usurped by a pathogen because H pylori uses it to adhere to gastric epithelial cells (GECs). H pylori is a gram-negative spiral bacterium that colonizes the human gastroduodenal mucosa. Infection with H pylori usually begins in childhood and persists for decades if untreated. H pylori is recognized as a major contributor to chronic gastritis and peptic ulcer formation and is strongly associated with gastric carcinoma and lymphoma[27,28]. Due to the strength of the evidence supporting an association between adenocarcinomas of the gastric mucosa and H pylori infection, H pylori was classified as a class I carcinogen by the International Agency for Research on Cancer in affiliation with the World Health Organization[29]. Gastric cancer remains among the most common forms of cancer and is the second deadliest cancer worldwide. Gastric cancer accounts for approximately 700 000 deaths annually worldwide and in the US there are 24 000 new cases and 14 000 deaths annually[30]. The prevalence rates of H pylori seropositivity and the incidence of gastric cancer are highly associated within several populations from various countries. For instance, seropositivity can be as high as 80%-100% in some age groups in some countries or in minorities with lower incomes in the United States[31]. These groups have the highest risk of developing gastric cancer and/or gastric ulcers. Thus, H pylori-associated diseases represent a significant global and domestic problem and result in considerable morbidity, mortality, and societal costs.
H pylori adhesion to the gastric epithelium is important in successful colonization of the gastric mucosa. Adherent strains survive in the gastric mucosa, reach high bacterial densities, and can re-colonize, whereas non-adherent strains are cleared[32]. These observations support the notion that adhesion is essential in H pylori persistence and disease induction. An assortment of molecules on epithelial cells have been proposed as receptors for H pylori adherence including carbohydrate moieties [such as Lewisb (Leb) blood group antigen and sialyl-dimeric-Lewisx (Lex)], lysophospholipid, and other structures[33–35]. Our studies have indicated that H pylori also utilizes CD74 to attach to gastric epithelial cells (GEC)[4]. The binding of H pylori to CD74 on gastric epithelial cells was confirmed by a series of independent approaches. For instance, blocking of CD74 with antibodies significantly reduced the binding of H pylori to gastric epithelial cells. Immunoprecipitation revealed that H pylori predominantly binds to the 33 kDa isoform of CD74, but further investigation is needed to test for attachment to the CS modified isoform. As H pylori has been reported to bind to various glycoconjugates, including glycosaminoglycans[36], this isoform of CD74 might contribute to the overall interaction of H pylori with the host gastric epithelium. We also revealed that urease is the protein on H pylori that binds to CD74[37]. This interaction is particularly interesting because many bacteria express urease, so the possibility exists that there might be wider applications of this type of interaction with CD74 depending on urease sequence variation between bacteria.
After adhesion of H pylori to GEC, the expression of cell surface CD74 is increased[14]. We further showed that CD74 expression increases in gastric epithelial cells of infected humans and a recent study confirmed that this increase in CD74 expression also occurred in a mouse model of H pylori infection[38]. Upon H pylori binding to CD74, NF-κB activation occurs resulting in the production of proinflammatory cytokines, including IL-8. IL-8 plays a major role in the proinflammatory immune response to H pylori infection, therefore, the interaction of H pylori with the gastric epithelial cells might be of critical importance in the immune response to infection.
THE ROLE OF CD74 IN SIGNALING EVENTS
The role of CD74 in signal transduction was initially hypothesized when it was found to be phosphorylated and associated with proteins that coordinate various signal transduction pathways[39–41]. Interestingly, the observation that CD74 deficient mice have a defect in B cell development that results in decreased levels of follicular B cells provided insights into the important role of signals delivered through CD74 in B cell development. The cytosolic domain of CD74 alone was noted to induce B cell maturation by activation of NF-κB[42]. CD74 appears to promote B cell survival; therefore, it has been implicated in B cell neoplasms such as gastric mucosa-associated lymphoid tissue (MALT) lymphomas associated with H pylori infection.
Our studies have shown that the interaction of H pylori with CD74 on GECs induces NF-κB activation and IL-8 production, as shown in Figure 2. MIF has also been shown to bind to CD74[3]. One study outlines a role for CD44 in CD74 signaling. CD44 was required to initiate ERK1 and ERK2 signaling after MIF binding to CD74. Cells deficient in CD44 or transfected with truncated CD44 were not able to induce ERK signaling[24]. In B cells, MIF binding to CD74 led to AKT, Syk, and NF-κB activation, and proliferation in a CD44-dependant manner[43]. CXCR2 has also been shown to complex with CD74 on monocytes[25]. This study also illustrated that MIF bound directly to CXCR2 and induced monocyte arrest. This study further showed that MIF might interact with CXCR4 on T cells and induce effector T cell arrest. However, it is not yet clear how CD44 is involved in the complexing of CD74 with CXCR2 and what signaling may be induced through CXCR4 since CXCR2 and CXCR4 are G protein-coupled receptors.
Figure 2.
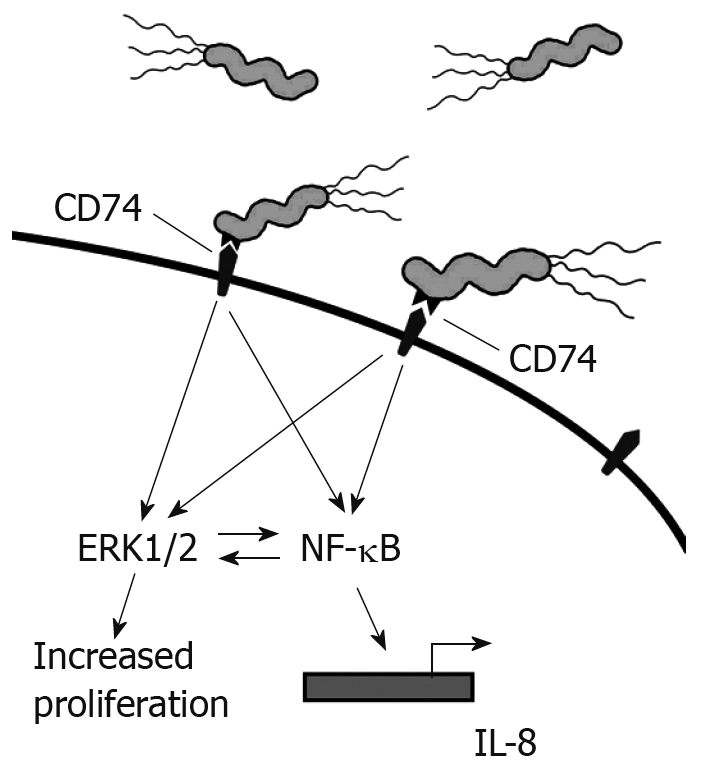
H pylori binds to CD74 on gastric epithelial cells and induces Nf-κB and Erk1/2 activation and IL-8 production.
Other studies suggest that MIF signaling may also occur by non-receptor mediated endocytosis in addition to the above described receptors[44]. In this proposed mechanism, endocytosed or endogenous MIF interacts with the Jun activation domain-binding protein (Jab1), which is a transcriptional activator for AP-1[45]. Activation of AP-1 might affect cell cycle events by inducing degradation of the cyclin dependent kinase inhibitor, which is a tumor suppresser gene.
THE ROLE OF CD74 IN GI INFLAMMATION
CD74 is highly expressed in inflammatory disorders. We have shown it to be expressed on the gastric epithelium and up-regulated during H pylori infection[14]. Others have shown expression to be increased in ulcerative colitis, where overexpression was shown in DNA microarray profiles[46]. Additionally, CD74 is increased during inflammation associated cancers, such as gastric and colon[47,48]. Concurrently, MIF is highly expressed during many inflammatory conditions of the GI tract. We and others have shown that production is increased during H pylori infection[19,49]. MIF is also highly expressed during inflammatory bowel disorders (IBD), such as ulcerative colitis and Crohn’s disease, where it is induced by intestinal bacteria[50]. In one study, elevated MIF levels were found in Crohn’s disease patients at approximately six times higher levels than in healthy controls. Crohn’s disease is an inflammatory bowel disorder where the immune system attacks part of the GI tract, and is accompanied by chronic inflammation. Also, this group went on to study MIF in murine colitis where they found colitis to be dependent on continuous MIF production, as evidenced by the protection from colitis by MIF-deficient mice or blocking MIF with monoclonal antibodies in mice with established colitis leading to reduced inflammation.
MIF or H pylori binding to CD74 induces NF-κB and subsequent cellular responses, such as the secretion of proinflammatory cytokines. MIF also increases inflammatory responses by overriding glucocortocoid suppression of inflammatory immune responses, including cytokines such as IL-1, IL-6, IL-8, and TNF-α[51]. One of the major proinflammatory cytokines produced after engagement of CD74 and receptor complexing is IL-8. IL-8 is a chemoattractant for neutrophils to a site of inflammation or infection. Upon arrival, they endocytose the antigen and form a phagosome in which reactive oxygen species and hydrolytic enzymes are released. While this process is crucial in fighting infections, it might also exacerbate inflammation in H pylori infection and inflammatory bowel disease[52–54]. In another study of glucocortocoid resistant ulcerative colitis, MIF was found to increase IL-8 production through the p38 MAPK pathway with isolated lamina propria mononuclear cells from patient biopsies[55]. When MIF was blocked with monoclonal antibodies after prednisolone treatment, activation of the p38 pathway and IL-8 decreased.
MIF might also contribute to inflammation by regulating Toll Like Receptor 4 (TLR4) expression on immune cells. TLR4 engagement by ligands such as bacterial LPS leads to proinflammatory cytokine production. This mechanism might be especially important in IBD, in which intestinal bacteria are a major contributor to the induction of the strong inflammatory response. In an in vivo study in mice, TLR4 expression in colonic tissue was not seen in MIF knockout mice, although it was in wild-type mice[56]. When the MIF knockout mice were administered rMIF, TLR4 expression was restored and further increased in colonic mice. In a human study of macrophages, neutralizing MIF or deleting the MIF gene resulted in decreased expression of TLR4 and a decreased response to LPS and gram negative bacteria; in broader experiments a decreased response to staphylococcal toxic shock and septic shock was demonstrated[57]. In addition, these cells did not respond well to LPS or gram negative bacteria and had a decreased expression of TLR4. The role of this receptor in the inflammation seen during H pylori infection is not clear because although H pylori LPS has been suggested to induce only weak responses, there are some studies suggesting it might contribute to the overall immune response to H pylori[58,59].
In addition to the role IL-8 plays in inflammation, we have previously shown that IL-8 increases expression of CD74 by gastric epithelial cells, both at the cell surface and mRNA levels[14] (Figure 3). Similarly, we found increased CD74 expression in vivo. Most of the H pylori infected samples and the samples with gastritis for reasons other than H pylori infection had much higher expression of CD74 than uninfected samples not exhibiting signs of gastritis. Other studies have further shown the expression of CD74 increased in ulcerative colitis and Crohn’s Disease[46]. Increased CD74 expression could then go on to intensify inflammation by providing more free receptors for MIF or H pylori attachment.
Figure 3.
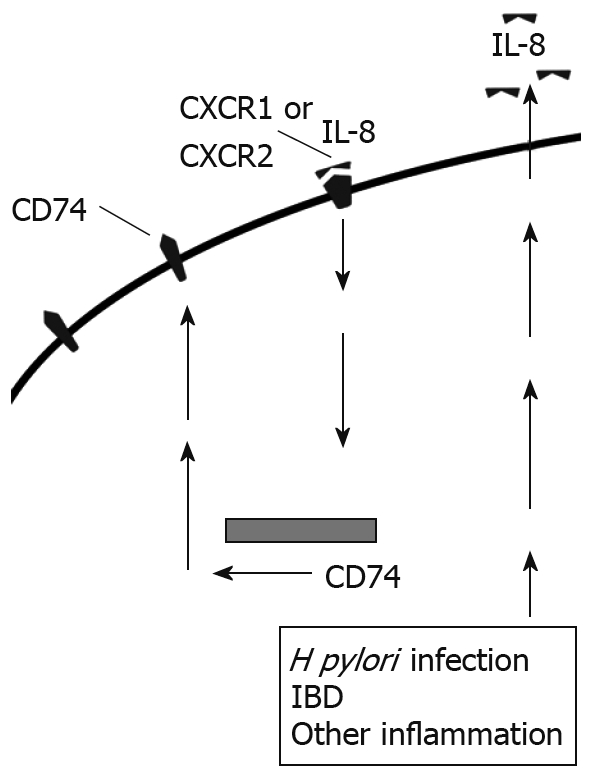
IL-8 produced by epithelial cells binds to CXCR1 and CXCR2 and up-regulates CD74 expression in an autocrine manner.
THE ROLE OF CD74 IN GI CANCERS
CD74 has a strong link to carcinogenesis, as does MIF. This ligand/receptor combination might be an important link between chronic inflammation and carcinogenesis. CD74 expression and MIF production have been shown to be increased during H pylori infection and gastric cancer[14,47]. High expression has also been noted in colon cancer along with highly increased serum concentrations of MIF in patients with colorectal cancers[60,61]. The contribution of CD74 to carcinogenesis is multifaceted. High levels of CD74 expression associated with class II MHC expression might prevent tumor antigen presentation by blocking the peptide binding cleft and preventing antigenic peptide binding for presentation to T cells, rendering tumors less immunogenic. One study suggested this to be the case with colon neoplasms where expression was even increased from low to high grade neoplasms[48]. Chronic inflammation and IL-8 production leads to a prolonged increase in CD74 expression, which might not only decrease antigen presentation, but also exacerbate IL-8 production upon engagement by MIF or H pylori. MIF binding to the CD74 receptor complex also promotes proliferation of epithelial cells[19,62]. Long term increased proliferation overtakes natural cell cycle events and sets the stage for carcinogenesis.
MIF binding to CD74 might contribute to carcinogenesis in chronic conditions through the up-regulation of proinflammatory cytokines, including IL-8, which up-regulates CD74 and has its own mechanisms leading to increased proliferation, tumor growth, and angiogenesis. MIF or IL-8 binding to their receptors on epithelial cell surfaces induces the shedding of EGFR ligands in a mellatoprotease-dependent manner, and activation of EGFR through engagement of these ligands[63]. We and others have shown that this pathway is activated during H pylori infection[62,64,65]. Additionally, we found that EGFR expression is up-regulated in gastric epithelial cells during H pylori infection by MIF and IL-8" after the word infection. The EGFR is highly expressed in various cancers and is involved in pro-inflammatory responses and pro-carcinogenic events, including cell proliferation, migration, and invasion. Expression and activation of this receptor is well-documented in gastric and intestinal cancers[66,67]. One study suggests a correlation between EGFR expression on tumor cells, proliferation, and prognosis in gastric cancer[68]. Another study showed that treatment with antisense RNA for EGFR inhibited gastric tumor growth in a mouse model[69]. Blocking EGFR or its ligands is being studied in order to develop more effective treatments for GI cancers.
MIF also increases epithelial cell proliferation after binding to CD74. One way MIF might affect proliferation and cell cycle events is by regulating p53 tumor suppressor. Numerous cell cycle and apoptosis genes are controlled by p53. We have shown that phosphorylation of the p53 is decreased after MIF binding to CD74[19]. Others have shown that MIF can interact directly with p53 and prevent translocation to the nucleus where it becomes activated and acts as a transcription factor for apoptotic genes[70]. When p53 is blocked from transport to the nucleus, apoptotic pathways are decreased and proliferation increases. Suppression of p53 in macrophages results in a more robust inflammatory response, implying a further link between p53 and inflammation[71].
CONCLUSION
CD74 is a much more versatile molecule than originally thought, playing many important roles in the immune system. Of crucial importance is the role it plays in class II MHC processing and the regulation of antigen presentation. This is important in the GI tract because epithelial cells and subepithelial myofibroblasts express CD74 and act as antigen presenting cells. Furthermore, the expression of CD74 on the cell surface might increase chronic inflammatory responses important in both H pylori infection and IBD. As a receptor for MIF, CD74 also plays and crucial role in chronic inflammation and might represent a major link between chronic inflammation and carcinogenesis in gastric and intestinal cancers. Development of therapeutics for cancer involving blocking CD74 might provide effective treatments for GI cancers.
Supported by The National Institutes of Health Grant K22AI068712, the Texas Board of Higher Education, and the John Sealy Memorial Endowment Fund for Biomedical Research
Peer reviewer: Dr. Francesco Costa, Dipartimento di Medicina Interna, U.O. di Gastroenterologia, Università di Pisa, Via Roma, 67-56122 Pisa, Italy
S- Editor Li LF L- Editor Stewart GJ E- Editor Ma WH
References
- 1.Barrera CA, Beswick EJ, Sierra JC, Bland D, Espejo R, Mifflin R, Adegboyega P, Crowe SE, Ernst PB, Reyes VE. Polarized expression of CD74 by gastric epithelial cells. J Histochem Cytochem. 2005;53:1481–1489. doi: 10.1369/jhc.4A6552.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Henne C, Schwenk F, Koch N, Moller P. Surface expression of the invariant chain (CD74) is independent of concomitant expression of major histocompatibility complex class II antigens. Immunology. 1995;84:177–182. [PMC free article] [PubMed] [Google Scholar]
- 3.Leng L, Metz CN, Fang Y, Xu J, Donnelly S, Baugh J, Delohery T, Chen Y, Mitchell RA, Bucala R. MIF signal transduction initiated by binding to CD74. J Exp Med. 2003;197:1467–1476. doi: 10.1084/jem.20030286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Beswick EJ, Bland DA, Suarez G, Barrera CA, Fan X, Reyes VE. Helicobacter pylori binds to CD74 on gastric epithelial cells and stimulates interleukin-8 production. Infect Immun. 2005;73:2736–2743. doi: 10.1128/IAI.73.5.2736-2743.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Barrera C, Ye G, Espejo R, Gunasena S, Almanza R, Leary J, Crowe S, Ernst P, Reyes VE. Expression of cathepsins B, L, S, and D by gastric epithelial cells implicates them as antigen presenting cells in local immune responses. Hum Immunol. 2001;62:1081–1091. doi: 10.1016/s0198-8859(01)00281-6. [DOI] [PubMed] [Google Scholar]
- 6.Pieters J, Bakke O, Dobberstein B. The MHC class II-associated invariant chain contains two endosomal targeting signals within its cytoplasmic tail. J Cell Sci. 1993;106(Pt 3):831–846. doi: 10.1242/jcs.106.3.831. [DOI] [PubMed] [Google Scholar]
- 7.Barrera C, Espejo R, Reyes VE. Differential glycosylation of MHC class II molecules on gastric epithelial cells: implications in local immune responses. Hum Immunol. 2002;63:384–393. doi: 10.1016/s0198-8859(02)00386-5. [DOI] [PubMed] [Google Scholar]
- 8.Hershberg RM, Cho DH, Youakim A, Bradley MB, Lee JS, Framson PE, Nepom GT. Highly polarized HLA class II antigen processing and presentation by human intestinal epithelial cells. J Clin Invest. 1998;102:792–803. doi: 10.1172/JCI3201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Barrera CA, Pinchuk IV, Saada JI, Suarez G, Bland DA, Beswick E, Adegboyega PA, Mifflin RC, Powell DW, Reyes VE. Class II MHC-expressing myofibroblasts play a role in the immunopathogenesis associated with staphylococcal enterotoxins. Ann N Y Acad Sci. 2004;1029:313–318. doi: 10.1196/annals.1309.022. [DOI] [PubMed] [Google Scholar]
- 10.Saada JI, Pinchuk IV, Barrera CA, Adegboyega PA, Suarez G, Mifflin RC, Di Mari JF, Reyes VE, Powell DW. Subepithelial myofibroblasts are novel nonprofessional APCs in the human colonic mucosa. J Immunol. 2006;177:5968–5979. doi: 10.4049/jimmunol.177.9.5968. [DOI] [PubMed] [Google Scholar]
- 11.Powell DW, Adegboyega PA, Di Mari JF, Mifflin RC. Epithelial cells and their neighbors I. Role of intestinal myofibroblasts in development, repair, and cancer. Am J Physiol Gastrointest Liver Physiol. 2005;289:G2–G7. doi: 10.1152/ajpgi.00075.2005. [DOI] [PubMed] [Google Scholar]
- 12.Blum JS, Cresswell P. Role for intracellular proteases in the processing and transport of class II HLA antigens. Proc Natl Acad Sci USA. 1988;85:3975–3979. doi: 10.1073/pnas.85.11.3975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Reyes VE, Lu S, Humphreys RE. Cathepsin B cleavage of Ii from class II MHC alpha- and beta-chains. J Immunol. 1991;146:3877–3880. [PubMed] [Google Scholar]
- 14.Beswick EJ, Das S, Pinchuk IV, Adegboyega P, Suarez G, Yamaoka Y, Reyes VE. Helicobacter pylori-induced IL-8 production by gastric epithelial cells up-regulates CD74 expression. J Immunol. 2005;175:171–176. doi: 10.4049/jimmunol.175.1.171. [DOI] [PubMed] [Google Scholar]
- 15.Warmerdam PA, Long EO, Roche PA. Isoforms of the invariant chain regulate transport of MHC class II molecules to antigen processing compartments. J Cell Biol. 1996;133:281–291. doi: 10.1083/jcb.133.2.281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wright RJ, Bikoff EK, Stockinger B. The Ii41 isoform of invariant chain mediates both positive and negative selection events in T-cell receptor transgenic mice. Immunology. 1998;95:309–313. doi: 10.1046/j.1365-2567.1998.00595.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Naujokas MF, Morin M, Anderson MS, Peterson M, Miller J. The chondroitin sulfate form of invariant chain can enhance stimulation of T cell responses through interaction with CD44. Cell. 1993;74:257–268. doi: 10.1016/0092-8674(93)90417-o. [DOI] [PubMed] [Google Scholar]
- 18.Momburg F, Koretz K, Von Herbay A, Moller P. Nonimmune human cells can express MHC class II antigens in the absence of invariant chain--an immunohistological study on normal and chronically inflamed small intestine. Clin Exp Immunol. 1988;72:367–372. [PMC free article] [PubMed] [Google Scholar]
- 19.Beswick EJ, Pinchuk IV, Suarez G, Sierra JC, Reyes VE. Helicobacter pylori CagA-dependent macrophage migration inhibitory factor produced by gastric epithelial cells binds to CD74 and stimulates procarcinogenic events. J Immunol. 2006;176:6794–6801. doi: 10.4049/jimmunol.176.11.6794. [DOI] [PubMed] [Google Scholar]
- 20.Ishiguro Y, Yamagata K, Sakuraba H, Munakata A, Nakane A, Morita T, Nishihira J. Macrophage migration inhibitory factor and activator protein-1 in ulcerative colitis. Ann N Y Acad Sci. 2004;1029:348–349. doi: 10.1196/annals.1309.040. [DOI] [PubMed] [Google Scholar]
- 21.Wilson JM, Coletta PL, Cuthbert RJ, Scott N, MacLennan K, Hawcroft G, Leng L, Lubetsky JB, Jin KK, Lolis E, et al. Macrophage migration inhibitory factor promotes intestinal tumorigenesis. Gastroenterology. 2005;129:1485–1503. doi: 10.1053/j.gastro.2005.07.061. [DOI] [PubMed] [Google Scholar]
- 22.Kim SY, Lee YC, Kim HK, Blaser MJ. Helicobacter pylori CagA transfection of gastric epithelial cells induces interleukin-8. Cell Microbiol. 2006;8:97–106. doi: 10.1111/j.1462-5822.2005.00603.x. [DOI] [PubMed] [Google Scholar]
- 23.Shmuely H, Passaro D, Figer A, Niv Y, Pitlik S, Samra Z, Koren R, Yahav J. Relationship between Helicobacter pylori CagA status and colorectal cancer. Am J Gastroenterol. 2001;96:3406–3410. doi: 10.1111/j.1572-0241.2001.05342.x. [DOI] [PubMed] [Google Scholar]
- 24.Shi X, Leng L, Wang T, Wang W, Du X, Li J, McDonald C, Chen Z, Murphy JW, Lolis E, et al. CD44 is the signaling component of the macrophage migration inhibitory factor-CD74 receptor complex. Immunity. 2006;25:595–606. doi: 10.1016/j.immuni.2006.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bernhagen J, Krohn R, Lue H, Gregory JL, Zernecke A, Koenen RR, Dewor M, Georgiev I, Schober A, Leng L, et al. MIF is a noncognate ligand of CXC chemokine receptors in inflammatory and atherogenic cell recruitment. Nat Med. 2007;13:587–596. doi: 10.1038/nm1567. [DOI] [PubMed] [Google Scholar]
- 26.Backhed F, Torstensson E, Seguin D, Richter-Dahlfors A, Rokbi B. Helicobacter pylori infection induces interleukin-8 receptor expression in the human gastric epithelium. Infect Immun. 2003;71:3357–3360. doi: 10.1128/IAI.71.6.3357-3360.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Solnick JV, Tompkins LS. Helicobacter pylori and gastroduodenal disease: pathogenesis and host-parasite interaction. Infect Agents Dis. 1992;1:294–309. [PubMed] [Google Scholar]
- 28.Talley NJ, Zinsmeister AR, Weaver A, DiMagno EP, Carpenter HA, Perez-Perez GI, Blaser MJ. Gastric adenocarcinoma and Helicobacter pylori infection. J Natl Cancer Inst. 1991;83:1734–1739. doi: 10.1093/jnci/83.23.1734. [DOI] [PubMed] [Google Scholar]
- 29.Infection with Helicobacter pylori. IARC Monogr Eval Carcinog Risks Hum. 1994;61:177–240. [PMC free article] [PubMed] [Google Scholar]
- 30.Cancer Database. Available from: URL: http://www-dep.iarc.fr/ 2005. [Google Scholar]
- 31.Blaser MJ. Helicobacter pylori phenotypes associated with peptic ulceration. Scand J Gastroenterol Suppl. 1994;205:1–5. [PubMed] [Google Scholar]
- 32.Hayashi S, Sugiyama T, Asaka M, Yokota K, Oguma K, Hirai Y. Modification of Helicobacter pylori adhesion to human gastric epithelial cells by antiadhesion agents. Dig Dis Sci. 1998;43:56S–60S. [PubMed] [Google Scholar]
- 33.Boren T, Falk P, Roth KA, Larson G, Normark S. Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science. 1993;262:1892–1895. doi: 10.1126/science.8018146. [DOI] [PubMed] [Google Scholar]
- 34.Ilver D, Arnqvist A, Ogren J, Frick IM, Kersulyte D, Incecik ET, Berg DE, Covacci A, Engstrand L, Boren T. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science. 1998;279:373–377. doi: 10.1126/science.279.5349.373. [DOI] [PubMed] [Google Scholar]
- 35.Van den Brink GR, Tytgat KM, Van der Hulst RW, Van der Loos CM, Einerhand AW, Buller HA, Dekker J. H pylori colocalises with MUC5AC in the human stomach. Gut. 2000;46:601–607. doi: 10.1136/gut.46.5.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ascencio F, Fransson LA, Wadstrom T. Affinity of the gastric pathogen Helicobacter pylori for the N-sulphated glycosaminoglycan heparan sulphate. J Med Microbiol. 1993;38:240–244. doi: 10.1099/00222615-38-4-240. [DOI] [PubMed] [Google Scholar]
- 37.Beswick EJ, Pinchuk IV, Minch K, Suarez G, Sierra JC, Yamaoka Y, Reyes VE. The Helicobacter pylori urease B subunit binds to CD74 on gastric epithelial cells and induces NF-kappaB activation and interleukin-8 production. Infect Immun. 2006;74:1148–1155. doi: 10.1128/IAI.74.2.1148-1155.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kobayashi M, Lee H, Schaffer L, Gilmartin TJ, Head SR, Takaishi S, Wang TC, Nakayama J, Fukuda M. A distinctive set of genes is upregulated during the inflammation-carcinoma sequence in mouse stomach infected by Helicobacter felis. J Histochem Cytochem. 2007;55:263–274. doi: 10.1369/jhc.6A7097.2006. [DOI] [PubMed] [Google Scholar]
- 39.Anderson HA, Roche PA. Phosphorylation regulates the delivery of MHC class II invariant chain complexes to antigen processing compartments. J Immunol. 1998;160:4850–4858. [PubMed] [Google Scholar]
- 40.Kuwana T, Peterson PA, Karlsson L. Exit of major histocompatibility complex class II-invariant chain p35 complexes from the endoplasmic reticulum is modulated by phosphorylation. Proc Natl Acad Sci USA. 1998;95:1056–1061. doi: 10.1073/pnas.95.3.1056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Spiro RC, Quaranta V. The invariant chain is a phospho-rylated subunit of class II molecules. J Immunol. 1989;143:2589–2594. [PubMed] [Google Scholar]
- 42.Matza D, Wolstein O, Dikstein R, Shachar I. Invariant chain induces B cell maturation by activating a TAF(II)105-NF-kappaB-dependent transcription program. J Biol Chem. 2001;276:27203–27206. doi: 10.1074/jbc.M104684200. [DOI] [PubMed] [Google Scholar]
- 43.Gore Y, Starlets D, Maharshak N, Becker-Herman S, Kaneyuki U, Leng L, Bucala R, Shachar I. Macrophage migration inhibitory factor induces B cell survival by activation of a CD74-CD44 receptor complex. J Biol Chem. 2008;283:2784–2792. doi: 10.1074/jbc.M703265200. [DOI] [PubMed] [Google Scholar]
- 44.Kleemann R, Grell M, Mischke R, Zimmermann G, Bernhagen J. Receptor binding and cellular uptake studies of macrophage migration inhibitory factor (MIF): use of biologically active labeled MIF derivatives. J Interferon Cytokine Res. 2002;22:351–363. doi: 10.1089/107999002753675785. [DOI] [PubMed] [Google Scholar]
- 45.Kleemann R, Hausser A, Geiger G, Mischke R, Burger-Kentischer A, Flieger O, Johannes FJ, Roger T, Calandra T, Kapurniotu A, et al. Intracellular action of the cytokine MIF to modulate AP-1 activity and the cell cycle through Jab1. Nature. 2000;408:211–216. doi: 10.1038/35041591. [DOI] [PubMed] [Google Scholar]
- 46.Lawrance IC, Fiocchi C, Chakravarti S. Ulcerative colitis and Crohn’s disease: distinctive gene expression profiles and novel susceptibility candidate genes. Hum Mol Genet. 2001;10:445–456. doi: 10.1093/hmg/10.5.445. [DOI] [PubMed] [Google Scholar]
- 47.Ishigami S, Natsugoe S, Tokuda K, Nakajo A, Iwashige H, Aridome K, Hokita S, Aikou T. Invariant chain expression in gastric cancer. Cancer Lett. 2001;168:87–91. doi: 10.1016/s0304-3835(01)00503-1. [DOI] [PubMed] [Google Scholar]
- 48.Jiang Z, Xu M, Savas L, LeClair P, Banner BF. Invariant chain expression in colon neoplasms. Virchows Arch. 1999;435:32–36. doi: 10.1007/s004280050391. [DOI] [PubMed] [Google Scholar]
- 49.Xia HH, Lam SK, Chan AO, Lin MC, Kung HF, Ogura K, Berg DE, Wong BC. Macrophage migration inhibitory factor stimulated by Helicobacter pylori increases proliferation of gastric epithelial cells. World J Gastroenterol. 2005;11:1946–1950. doi: 10.3748/wjg.v11.i13.1946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.de Jong YP, Abadia-Molina AC, Satoskar AR, Clarke K, Rietdijk ST, Faubion WA, Mizoguchi E, Metz CN, Alsahli M, ten Hove T, et al. Development of chronic colitis is dependent on the cytokine MIF. Nat Immunol. 2001;2:1061–1066. doi: 10.1038/ni720. [DOI] [PubMed] [Google Scholar]
- 51.Calandra T, Bernhagen J, Metz CN, Spiegel LA, Bacher M, Donnelly T, Cerami A, Bucala R. MIF as a glucocorticoid-induced modulator of cytokine production. Nature. 1995;377:68–71. doi: 10.1038/377068a0. [DOI] [PubMed] [Google Scholar]
- 52.Ina K, Kusugami K, Yamaguchi T, Imada A, Hosokawa T, Ohsuga M, Shinoda M, Ando T, Ito K, Yokoyama Y. Mucosal interleukin-8 is involved in neutrophil migration and binding to extracellular matrix in inflammatory bowel disease. Am J Gastroenterol. 1997;92:1342–1346. [PubMed] [Google Scholar]
- 53.Leakey A, La Brooy J, Hirst R. The ability of Helicobacter pylori to activate neutrophils is determined by factors other than H pylori neutrophil-activating protein. J Infect Dis. 2000;182:1749–1755. doi: 10.1086/317611. [DOI] [PubMed] [Google Scholar]
- 54.Mazzucchelli L, Hauser C, Zgraggen K, Wagner H, Hess M, Laissue JA, Mueller C. Expression of interleukin-8 gene in inflammatory bowel disease is related to the histological grade of active inflammation. Am J Pathol. 1994;144:997–1007. [PMC free article] [PubMed] [Google Scholar]
- 55.Ishiguro Y, Ohkawara T, Sakuraba H, Yamagata K, Hiraga H, Yamaguchi S, Fukuda S, Munakata A, Nakane A, Nishihira J. Macrophage migration inhibitory factor has a proinflammatory activity via the p38 pathway in glucocorticoid-resistant ulcerative colitis. Clin Immunol. 2006;120:335–341. doi: 10.1016/j.clim.2006.05.010. [DOI] [PubMed] [Google Scholar]
- 56.Ohkawara T, Takeda H, Miyashita K, Nishiwaki M, Nakayama T, Taniguchi M, Yoshiki T, Tanaka J, Imamura M, Sugiyama T, et al. Regulation of Toll-like receptor 4 expression in mouse colon by macrophage migration inhibitory factor. Histochem Cell Biol. 2006;125:575–582. doi: 10.1007/s00418-005-0092-y. [DOI] [PubMed] [Google Scholar]
- 57.Roger T, Froidevaux C, Martin C, Calandra T. Macrophage migration inhibitory factor (MIF) regulates host responses to endotoxin through modulation of Toll-like receptor 4 (TLR4) J Endotoxin Res. 2003;9:119–123. doi: 10.1179/096805103125001513. [DOI] [PubMed] [Google Scholar]
- 58.Ishihara S, Rumi MA, Kadowaki Y, Ortega-Cava CF, Yuki T, Yoshino N, Miyaoka Y, Kazumori H, Ishimura N, Amano Y, et al. Essential role of MD-2 in TLR4-dependent signaling during Helicobacter pylori-associated gastritis. J Immunol. 2004;173:1406–1416. doi: 10.4049/jimmunol.173.2.1406. [DOI] [PubMed] [Google Scholar]
- 59.Smith MF Jr, Mitchell A, Li G, Ding S, Fitzmaurice AM, Ryan K, Crowe S, Goldberg JB. Toll-like receptor (TLR) 2 and TLR5, but not TLR4, are required for Helicobacter pylori-induced NF-kappa B activation and chemokine expression by epithelial cells. J Biol Chem. 2003;278:32552–32560. doi: 10.1074/jbc.M305536200. [DOI] [PubMed] [Google Scholar]
- 60.Degener T, Momburg F, Moller P. Differential expression of HLA-DR, HLA-DP, HLA-DQ and associated invariant chain (Ii) in normal colorectal mucosa, adenoma and carcinoma. Virchows Arch A Pathol Anat Histopathol. 1988;412:315–322. doi: 10.1007/BF00750257. [DOI] [PubMed] [Google Scholar]
- 61.Yasasever V, Camlica H, Duranyildiz D, Oguz H, Tas F, Dalay N. Macrophage migration inhibitory factor in cancer. Cancer Invest. 2007;25:715–719. doi: 10.1080/07357900701560695. [DOI] [PubMed] [Google Scholar]
- 62.Beswick EJ, Reyes VE. Macrophage migration inhibitory factor and interleukin-8 produced by gastric epithelial cells during Helicobacter pylori exposure induce expression and activation of the epidermal growth factor receptor. Infect Immun. 2008;76:3233–3240. doi: 10.1128/IAI.01534-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Itoh Y, Joh T, Tanida S, Sasaki M, Kataoka H, Itoh K, Oshima T, Ogasawara N, Togawa S, Wada T, et al. IL-8 promotes cell proliferation and migration through metalloproteinase-cleavage proHB-EGF in human colon carcinoma cells. Cytokine. 2005;29:275–282. doi: 10.1016/j.cyto.2004.11.005. [DOI] [PubMed] [Google Scholar]
- 64.Joh T, Kataoka H, Tanida S, Watanabe K, Ohshima T, Sasaki M, Nakao H, Ohhara H, Higashiyama S, Itoh M. Helicobacter pylori-stimulated interleukin-8 (IL-8) promotes cell proliferation through transactivation of epidermal growth factor receptor (EGFR) by disintegrin and metalloproteinase (ADAM) activation. Dig Dis Sci. 2005;50:2081–2089. doi: 10.1007/s10620-005-3011-0. [DOI] [PubMed] [Google Scholar]
- 65.Keates S, Keates AC, Nath S, Peek RM Jr, Kelly CP. Transactivation of the epidermal growth factor receptor by cag+ Helicobacter pylori induces upregulation of the early growth response gene Egr-1 in gastric epithelial cells. Gut. 2005;54:1363–1369. doi: 10.1136/gut.2005.066977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Svrcek M, Cosnes J, Tiret E, Bennis M, Parc Y, Flejou JF. Expression of epidermal growth factor receptor (EGFR) is frequent in inflammatory bowel disease (IBD)-associated intestinal cancer. Virchows Arch. 2007;450:243–244. doi: 10.1007/s00428-006-0338-7. [DOI] [PubMed] [Google Scholar]
- 67.Tokunaga A, Onda M, Okuda T, Teramoto T, Fujita I, Mizutani T, Kiyama T, Yoshiyuki T, Nishi K, Matsukura N. Clinical significance of epidermal growth factor (EGF), EGF receptor, and c-erbB-2 in human gastric cancer. Cancer. 1995;75:1418–1425. doi: 10.1002/1097-0142(19950315)75:6+<1418::aid-cncr2820751505>3.0.co;2-y. [DOI] [PubMed] [Google Scholar]
- 68.Jonjic N, Kovac K, Krasevic M, Valkovic T, Ernjak N, Sasso F, Melato M. Epidermal growth factor-receptor expression correlates with tumor cell proliferation and prognosis in gastric cancer. Anticancer Res. 1997;17:3883–3888. [PubMed] [Google Scholar]
- 69.Hirao T, Sawada H, Koyama F, Watanabe A, Yamada Y, Sakaguchi T, Tatsumi M, Fujimoto H, Emoto K, Narikiyo M, et al. Antisense epidermal growth factor receptor delivered by adenoviral vector blocks tumor growth in human gastric cancer. Cancer Gene Ther. 1999;6:423–427. doi: 10.1038/sj.cgt.7700058. [DOI] [PubMed] [Google Scholar]
- 70.Jung H, Seong HA, Ha H. Critical role of cysteine residue 81 of macrophage migration inhibitory factor (MIF) in MIF-induced inhibition of p53 activity. J Biol Chem. 2008;283:20383–20396. doi: 10.1074/jbc.M800050200. [DOI] [PubMed] [Google Scholar]
- 71.Mitchell RA, Liao H, Chesney J, Fingerle-Rowson G, Baugh J, David J, Bucala R. Macrophage migration inhibitory factor (MIF) sustains macrophage proinflammatory function by inhibiting p53: regulatory role in the innate immune response. Proc Natl Acad Sci USA. 2002;99:345–350. doi: 10.1073/pnas.012511599. [DOI] [PMC free article] [PubMed] [Google Scholar]