

Abstract
The development of peripheral to central neural connections within the auditory, visual, and olfactory systems of mice is reviewed to address whether peripheral signaling may play an instructive role during initial synapse formation. For each sensory system, developmental times of histogenesis and the earliest ages of innervation and function are considered for peripheral and selected central relays. For the auditory and visual system, anatomical and functional reports indicate that central connections may form prior to synapse formation in the periphery. However, evidence from the olfactory system suggests that the peripheral olfactory sensory neurons form synaptic connections before more central olfactory connections are established. We find that significant gaps in knowledge exist for embryonic development of these systems in mice and that genetic tools have not yet been systematically directed to address these issues.
Keywords: Development, Synaptogenesis, Comparative, Auditory, Visual, Olfactory
1. Introduction
Neural circuits begin to form during early brain development at embryonic ages. Whether each synaptic relay of the circuit forms autonomously or is directed in some fashion by a previous relay is a key issue and is typically addressed by investigating whether neural activity plays a role in neural development. Here we evaluate the more general question of whether neural signaling, including both electrical and synaptic activity but also non-synaptic biochemical interactions between neurons and along neural pathways, drives circuit formation. This issue is more easily approached in sensory systems because the primary direction of information transfer is specified, in that neural activity originates in the peripheral sensory organ and propagates along neural circuits in the CNS. One means to answer the question of whether sensory systems connect from periphery to cortex, or specifically, does one synaptic relay drive patterning and neural connectivity at subsequent relays, is to examine whether central contacts between neurons form and are functional prior to ability of the sensory periphery to transmit information to more central structures. In this context we evaluate when cellular processes first reach the vicinity of cells with which they will later form classical synaptic contacts. We explored this issue by comparing three relay stations of the ascending neural circuit in sensory systems: (1) receptor cells and associated neurons including ganglion cells, (2) first order CNS target neurons of the sensory ganglion neurons, and (3) second order CNS target neurons of the first order CNS neurons. We present and consider the state of knowledge for three well-studied sensory systems, the auditory, visual and olfactory systems. Given an increasing emphasis on the mouse model for studies of nervous system development, we focus our presentation on data from that species. For each sensory system, we present the birthdates of the circuit-forming cells and information about the structural and functional development of these selected neural connections. We evaluate when the anatomical substrate for each functional connection is established by reporting the earliest evidence from light and/or electron microscopy (EM) studies. We then present the earliest evidence for functional synaptic connections based on electrophysiological recordings, induced expression of immediate early genes, or behavioral readouts of appropriate sensory stimulation.
Although the auditory, visual and olfactory systems are similar in basic organization, they differ in detail. The olfactory system is simplest in peripheral organization, in that the olfactory sensory neuron (OSN) forms the olfactory nerve (without an intervening synapse) and so projects directly into the CNS. The auditory system incorporates a synaptic connection between the receptor cell, or hair cell, and peripheral ganglion cell whose axons form the auditory nerve projection into the CNS. The visual system is most complex in its peripheral organization, whereby the retina is a multi-layered neural network that is part of the CNS, with multiple synaptic connections preceding input to ganglion cells whose axons form the optic nerve projection to the thalamus and other central targets. For the visual and olfactory pathways presented here, the first order target neurons of their respective ganglion neurons project directly to sensory cortex. In contrast, the auditory system has at least four processing stations interposed between the first order CNS target of the auditory nerve and auditory cortex. The systems differ also in the onset of sensitivity to external environmental stimuli. The olfactory system is exposed to airborne odorants at birth, but the auditory and visual systems are not engaged fully until airborne sound is capable of eliciting action potentials in the auditory nerve at postnatal day (P)9 (Mikaelian and Ruben, 1965) and the eyes open at P13-14 (Poole, 1987).
The purpose of this article is to describe and compare the initial formation of synaptic contacts among these three sensory systems. Refinement of central connections that is dependent on neural activity is beyond the scope of this presentation, and we refer the reader to the following references that describe refinement in the auditory (Huang et al., 2007; Kandler, 2004; Rubel et al., 1998), visual (Hooks and Chen, 2007; Huberman et al., 2008; Torborg and Feller, 2005), and olfactory systems (Schwob et al., 1984; Schwob and Price, 1984; Zou et al., 2004). Although this review is focused upon comparisons among sensory systems in the mouse, references to other rodent species are made where data from mouse are incomplete. We found that many questions regarding the timing and nature of neural circuit formation in these systems remain open, so a key purpose of this presentation is to highlight topics that merit further study.
2. Auditory System
The three circuit connections that we consider for the auditory system are: (1) the connection between the inner hair cell and primary afferent spiral ganglion neuron, (2) the connection between the spiral ganglion neuron and a subdivision of its target cell group in the CNS, the ventral subdivision of the cochlear nucleus (VCN), and (3) the connection between the VCN and the next processing station in the auditory pathway, the superior olivary complex (SOC) located in the core of the brainstem (Fig. 1). Some VCN neurons innervate auditory centers other than the SOC, but the vast majority of these projections must first pass through the region of the SOC (Howell et al., 2007; Kandler and Friauf, 1993). Therefore, the potential to form functional connections occurs earliest for connections with the SOC cell groups. Among the many SOC cell groups (Harrison and Warr, 1962; Ollo and Schwartz, 1979), two that are most prominent in all mammalian species are the medial nucleus of the trapezoid body (MNTB) and lateral superior olive (LSO). The development of MNTB and LSO is best studied among SOC cell groups, so they are the focus of our analysis. For more general discussion of peripheral and brainstem auditory system development, refer to the scholarly presentations by Cant (1998) and Pujol et al. (1998).
Figure 1.
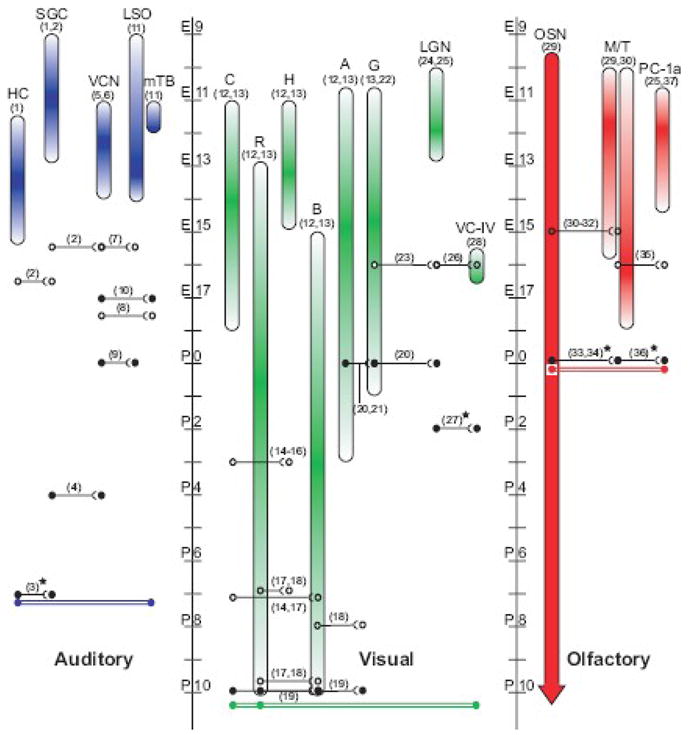
Schematic of sensory system organizations, cellular birthdates, anatomical circuitry, and functional connectivity. The auditory, visual, and olfactory systems discussed in this review are diagrammed from left to right as peripheral to central. Colored vertical bars represent the birthdates of each cell type/nucleus listed, with the darkest color representing the peak in histogenesis. Continuous histogenesis of OSNs is designated by the uniformly colored bar with a downward arrow beyond P10. Open circle connections represent anatomical evidence of connections, either by light or electron microscopy. Closed circle connections represent demonstrated functional connectivity, either by electrophysiological recording or c-fos immunoreactivity. Stars indicate data are from rat. The colored double bar connections represent the earliest known date by which the peripheral-to-central circuit can be functionally active. Abbreviations- E= embryonic day; P= postnatal day; HC= hair cell; SGC= spiral ganglion cell; VCN= ventral cochlear nucleus; LSO= lateral superior olive; mTB= medial nucleus of the trapezoid body; C= cone photoreceptor; R= rod photoreceptor; H= horizontal cell; B= bipolar cell; A= amacrine cell; G= retinal ganglion cell; LGN= lateral geniculate nucleus; VC-IV- layer IV of the visual cortex; OSN= olfactory sensory neuron; M/T= mitral and tufted cells; PC-1a= layer 1a of piriform cortex. Numbers in parenthisis refer to the following references: 1, Ruben, 1967; 2, Koundakjian et al., 2007, 3, Glowatzki and Fuchs, 2002; 4, Wu and Oertel, 1987; 5, Martin and Rickets, 1981; 6, Pierce, 1967; 7, Howell et al., 2007; 8, P Mathers personal communication; 9, Ene et al., 2003; 10, Hoffpauir & Spirou personal communication; 11, Pierce, 1973; 12, Cepko et al., 1996; 13, Young, 1985; 14, Blanks et al., 1974; 15, Olney, 1968; 16, Rich et al., 1997; 17, Sharma et al., 2003; 18, Sherry et al., 2003; 19, Tian and Copenhagen, 2003; 20, Mooney et al., 1996; 21, Wang et al., 2007; 22, Rapaport et al., 2004; 23, Godement et al., 1984; 24, Angevine, 1970; 25, Gardette et al., 1982; 26, Del Rio et al., 2000; 27, Hanganu et al., 2006; 28, Polleux et al., 1997; 29, Hinds, 1968; 30, Blanchart et al., 2006; 31, Blanchart et al., 2008; 32, Hinds and Hinds, 1976; 33, Mair and Gesteland, 1982; 34, Guthrie and Gall, 2003; 35, Walz et al., 2006; 36, Schwob and Price, 1984; 37, Hinds and Angevine, 1965.
Histogenesis
Cochlear hair cells are born between E11.5–15.5 in mice, initially near the cochlear base and in a pattern progressing toward both its basal and apical ends (Chen and Segil, 1999; Ruben, 1967; Woods et al., 2004; birthdates of cells discussed in this paragraph are summarized in Fig. 1). In an elegant set of studies, Koundakjian et al. (2007) utilized genetic labeling techniques to distinguish the location within the otic placode giving rise to auditory and vestibular systems. They showed that spiral ganglion cell birthdays occurred between E9.5–12.5, generally following a basal (high frequency) to apical (low frequency) gradient as previously described (Ruben, 1967). Analysis of brainstem neurons revealed that each cell group is born over the course of several days (Martin and Rickets, 1981; Pierce, 1967; Pierce, 1973). Birth of VCN neurons began later than spiral ganglion neurons but was complete within three days (E11–14). MNTB neurons began this process coincident with VCN neurons, at embryonic day 11 but completed histogenesis at E12. LSO neurons had a longer birth process, extending from E9–14 and peaking at E13. Note that cells in the CNS are born along the ventricular surface and many cell groups, including the SOC, migrate to their eventual location, so their birthdates are distinct from their formation of an appropriately positioned cell group.
Anatomical Connectivity Between Inner Hair Cells and Spiral Ganglion Cells
Using genetic labeling techniques, Koundakjian et al. (2007) were able to distinguish type I and II spiral ganglion neurons and their peripheral processes along the length of the cochlea. Here, we focus on type I spiral ganglion cells, which constitute 95% of the total and innervate inner hair cells (Spoendlin, 1967). At E16.5, peripheral processes of spiral ganglion neurons extended into the inner hair cell zone of the Organ of Corti, where they could potentially innervate inner hair cells. The open circles in figure 1 indicate the earliest age at which anatomical connections could exist. However, EM studies have not been conducted at this age in mice to establish the existence of synaptic structures and functional connections between the cells. Sobkowicz et al. (2003) have observed reciprocal synapses between hair cells and afferent fibers at P1, the earliest age studied, but we are unaware of ultrastructure studies in mice at earlier ages.
Anatomical Connectivity Between Spiral Ganglion and Ventral Cochlear Nucleus Cells
Genetic labeling of spiral ganglion cell bodies and dendrites by Koundakjian et al. (2007) also revealed growth of axons into the cochlear nucleus. At E15.5, just prior to entry of spiral ganglion cell dendrites into the region of inner hair cells, their central processes have already reached the developing cochlear nucleus. These projections are tonotopically correct, in that spiral ganglion neurons that innervate the cochlear base or apex innervate the dorso-medial or ventro-lateral cochlear nucleus, respectively. A comment in the classic study by Pierce (1967) indicates that cochlear nerve fibers enter the ventricular zone where VCN neurons are forming at E11. However, the experimental basis for this conclusion was not presented. Even using the more conservative age of E15.5, and noting that Koundakjian et al. (2007) did not report exploration of earlier ages, the possibility exists, then, that central connections are formed prior to peripheral connections. High magnification light microscopy shows nerve terminal elaborations more consistent with synaptic rather than growth cone profiles in the VCN, but these data were from early postnatal (P1) mice (Limb and Ryugo, 2000).
Anatomical Connectivity Between Ventral Cochlear Nucleus and Superior Olivary Complex
The SOC, although comprised of multiple nuclei (rat: Harrison and Warr, 1962; mouse: Ollo and Schwartz, 1979), is first recognizable as a large group of cells at E19 in rat (Kandler and Friauf, 1993) and at E15.5 in mouse (P Mathers, personal communication). VCN axons labeled with lipophilic dyes have been observed entering the future region of the ipsilateral LSO at E13.5 and crossing the midline and entering the vicinity of what will be the contralateral MNTB at E14.5 (Howell et al., 2007). At E15.5, VCN axons begin to branch and extend into the presumptive LSO (Howell et al., 2007). However, the MNTB is not discernible as an individual nucleus within the SOC until E17.5, after MNTB neurons have completed their medial migration (P Mathers, personal communication). Therefore, we indicate that anatomical observations of neural connections from the VCN to the LSO and MNTB have been observed at E15.5 and E17.5, respectively (open circles in Fig. 1). EM evidence for synaptic terminals that contain vesicle release machinery in the SOC was collected at the earliest age from MNTB cells in mice at P0 (Hoffpauir et al., 2006), but EM studies of innervation of the LSO have not been investigated, to our knowledge, in embryonic mice.
Functional Connectivity Between Inner Hair Cells and Spiral Ganglion Cells
Recently, mechanisms for inner hair cell-induced activity in spiral ganglion neurons have been demonstrated. Beginning at E17.5, inner hair cells generate spontaneous Ca2+-based action potentials that could lead to neurotransmitter release (Kros et al., 1998; Marcotti et al., 2003a; Marcotti et al., 2003b). Changes in membrane capacitance induced by imposed depolarization are an indicator of synaptic vesicle fusion (Neher and Marty, 1982). These measurements from inner hair cells reveal a very immature vesicle release mechanism at E16.5 that becomes more robust by E18.5 (Johnson et al., 2005) and at early postnatal ages (Beutner et al., 2001). Therefore, local spontaneous release of neurotransmitter, in the absence of hair cell action potentials, could occur as early as E16.5 in concert with the growth of spiral ganglion dendrites into the vicinity of inner hair cells, but likely at very low release rates. Immunoctyochemistry experiments suggest that spiral ganglion cells express NMDA and AMPA receptor subunits at E13 and E17, respectively (Puyal et al., 2002), although we are unaware of any studies that confirm expression of these receptors on spiral ganglion cell dendrites. Therefore, should synaptic connections form at embryonic ages, neurotransmitter release could potentially evoke postsynaptic responses in spiral ganglion cell as early as E16.5–17.5.
In vivo single cell recordings are difficult to perform on auditory nerve or brainstem neurons at perinatal ages. Consequently, experiments have only been conducted using in vitro cochlear dissection, brain slice or dissociated neuron preparations. In technically difficult experiments, postsynaptic currents are measured via patch electrodes in spiral ganglion cell dendrites contacting inner hair cells. Hair cell depolarization induces postsynaptic currents in spiral ganglion dendrites, but these studies have only been conducted postnatally in rats at ages (P7–P10; Glowatzki and Fuchs, 2002) and mice (P6–P16; Seal et al., 2008). However, the earliest age at which synaptic currents were recorded in mice was not specified. Therefore, we indicate by solid circles in figure 1 that the earliest age at which the hair cell synapse is verified to be functional is P7.
Functional Connectivity Between Spiral Ganglion and Ventral Cochlear Nucleus Cells
Although postsynaptic currents are induced in dendrites, at very young ages the spiral ganglion cell may not propagate an action potential into the CNS. Dissociated spiral ganglion neurons are capable of generating action potentials at P1, but younger, especially embryonic, ages have not been studied (Mo and Davis, 1997). At what age do spiral ganglion neurons drive postsynaptic activity in the cochlear nucleus? Synaptic activation of cochlear nucleus neurons has been measured in mice at P4 (Wu and Oertel, 1987), but younger ages have not been tested, perhaps due to the fragility of the auditory nerve during tissue dissection. Therefore, we indicate by solid circles in figure 1 that the earliest age at which spiral ganglion cell synapses in VCN are verified to be functional is P4.
Functional Connectivity Between Ventral Cochlear Nucleus and Superior Olivary Complex
Functional connectivity from the VCN to LSO has been confirmed in mice at P0 using Ca2+ imaging to assay synaptic responses in the LSO while stimulating VCN axons (Ene et al., 2003). Although functional connectivity has been demonstrated at E18 in rats, the earliest age tested (Kandler and Friauf, 1995), studies at embryonic ages have not been performed in mice. Therefore, we indicate by solid circles in figure 1 that the earliest age at which VCN neuron synapses in the LSO are verified to be functional in mice is P0. Synaptic currents in MNTB cells induced via stimulation of VCN axons have been reported at P1 for mice (Hoffpauir et al., 2006) and P0 for rats (Rodriguez-Contreras et al., 2008). In the only investigation of synaptic activity in the auditory system conducted at embryonic ages, synaptic currents recorded in MNTB neurons were inducible by stimulation of VCN axons when MNTB neurons are first discernable at E17 (Hoffpauir and Spirou, personal communication). Investigation at earlier ages requires reliable identification of MTNB neurons among the SOC population. Therefore, we indicate with solid circles in figure 1 that the earliest age at which VCN synapses in the MNTB are verified to be functional is E17.
3. Visual System
Of the three sensory organs discussed here, the neuronal architecture within the eye is the most complex in terms of cell types, circuitry and processing. Therefore, for the purpose of this review, we focus on the following retinal circuits that together comprise the initial connections in the visual system: (1a) photoreceptor connections onto bipolar cells, (1b) bipolar cell connections onto retinal ganglion cells (RGCs), and (1c) amacrine cell synapses onto RGCs. For the sake of simplicity, we exclude many specific details required for a complete description of retinal development such as bifurcation of the visual message into ON and OFF circuits that involve a myriad of possible interactions throughout the retina. As with the auditory system, we also focus on two sequential central synapses, (2) the initial projection of RGC axons to the lateral geniculate nucleus (LGN) in the thalamus, and (3) the projection from LGN neurons to layer IV of visual cortical area V1 (Fig. 1).
Histogenesis
All six retinal neuronal classes, as well as the Müller glia, derive from a multipotent retinal progenitor cell (Holt et al., 1988; Turner and Cepko, 1987; Wetts and Fraser, 1988). The birth order of retinal neurons in the mouse exhibits the same stereotypical cellular pattern as that found in other species, with two distinct but overlapping phases of cellular birth (Rapaport et al., 2004; Young, 1985). Early-born cell fates include retinal ganglion cells, horizontal cells, amacrine cells and cone photoreceptors, with ganglion cells the first retinal cells to be born (Polley et al., 1989; Rapaport et al., 2004). Late-born neuronal fates include rod photoreceptors and bipolar cells. The early-born ganglion cells first exit the cell cycle at E10.5, and continue to be generated until early postnatal stages (P1), with the peak of histogenesis around E14.5 (Rapaport et al., 2004; Young, 1985). Late-born rod photoreceptors are born beginning around E13 and continue histogenesis until about P10, with the majority of rods being formed around the birth of the mouse pup. Similar overlaps are seen between early-born amacrine cells, with histogenesis ranging from E10.5 to P3, with a peak around E15, and late-born bipolar cells, which range in birth from E15 to P10, with a peak around P3. Despite these early and late distinctions, significant portions of both classes are still being generated around the birth of the animal. The remaining two neuronal classes within the retina are the horizontal cells, where histogenesis occurs between E11 and E15, with a peak at E13, and the cone photoreceptors, which are generated between E11and E18, with a peak of histogenesis at E14 (Cepko et al., 1996; Young, 1985). Histogenesis in the LGN occurs in the window beginning at E10 and ending at E13, with a peak of birth rate around E12.5 (Angevine, 1970; Gardette et al., 1982). The final element in the circuit we are considering, layer IV of V1 in the visual cortex, is the last to begin but the fastest to complete histogenesis, with birth dates occurring between E15.5 and E16.5, with E16.5 the peak of histogenesis for this layer (Polleux et al., 1997).
Anatomical Connectivity Within the Retina
In the outer plexiform layer (OPL) of the mouse retina, synaptic connections between photoreceptors and their postsynaptic targets, the horizontal cells and bipolar cells, form over a week-long period prior to eye opening. Ultrastructural (Blanks et al., 1974; Olney, 1968) and immunohistochemical (Rich et al., 1997) studies suggest that a small fraction of cone photoreceptors extend toward the presumptive OPL and form synapses with horizontal cells as early as P3. Cone photoreceptors then begin forming synapses with bipolar cells around P7, after bipolar cells have extended processes to the OPL (Blanks et al., 1974; Sharma et al., 2003). Rod synapse formation is delayed, relative to cone synapse formation, with synapses onto horizontal cells first forming around P7 and rod bipolar cell dendrites contacting the rods around P10 (Sharma et al., 2003; Sherry et al., 2003).
In the inner plexiform layer (IPL), bipolar cells and amacrine cells form synapses with each other and onto RGCs. The first synapses formed in the IPL are likely between amacrine and RGCs. Ultrastructural studies of the developing mouse retina indicate that conventional synapses occur in the IPL as early as P2 (Olney, 1968). Given that bipolar cells do not appear to extend axons into the IPL at P3 (Sharma et al., 2003), these synapses are likely presynaptic amacrine cell processes forming synapses onto other amacrine cells and/or ganglion cells. The characteristic ribbon synapses of bipolar cells first appear around P10–P11 (Fisher, 1979; Olney, 1968). However, bipolar cells may form conventional synapses that cannot be distinguished from amacrine-to-ganglion connections at ultrastructural resolution prior to the appearance of presynaptic ribbon complexes. Indeed, immunohistochemical studies suggest that bipolar cells form synapses in the IPL as early as P8 (Sherry et al., 2003). Therefore, we indicate by open circles in figure 1, that the earliest anatomical observations of bipolar cell synapses onto RGCs have been reported for mice at P8.
Anatomical Connections from RGCs to LGN and LGN to Visual Cortex
The RGC axons comprising the optic nerve cross the optic chiasm around E14 and first extend into the contralateral LGN at E16, but the initial growth into the ipsilateral LGN is delayed until E18 (Godement et al., 1984). Thalamocortical projections in mice reach the cortical subplate at E15, but the earliest innervating fibers wait until E16 to invade cortical layers (Del Rio et al., 2000). To our knowledge, there are no ultrastructural studies detailing the initial formation of synaptic contacts within the LGN and visual cortex of mice. However, these light microscopic studies indicate that axons begin to invade both of these central nuclei simultaneously at E16 (open circles in Fig. 1).
Functional Connectivity Within the Retina
In order for ON or OFF RGCs to respond to photic stimuli with their characteristic ON or OFF firing properties, appropriate upstream connections between photoreceptors, bipolar cells and, for the rod circuitry, amacrine cells must be functional. Thus, evidence of ON and OFF responses in RGCs is indicative of functional connectivity for the connections within the retina. Using multi-electrode arrays, Tian and Copenhagen (2003) recorded distinct ON, ON-OFF, and OFF firing patterns from RGCs during light stimulation of a mouse retina preparation as early as P10. Therefore, this study confirmed that the key elements in the retinal ON and OFF circuit were connected and functional as early as P10.
Many studies of early retinal function in mice focus on propagating Ca2+ waves in the inner retina, which indicate functional connectivity among retinal neurons prior to the retina becoming sensitive to light. In multiple mammalian species, multi-electrode array recordings and Ca2+ imaging indicate that waves of activity spread across the retina prior to eye opening (Feller et al., 1996; Feller et al., 1997; Meister et al., 1991; Wong et al., 1993; Wong et al., 1995). Three stages of retinal waves have been proposed. Stage I waves spread across the retina prior to synapse formation and involve signaling through gap junctions, as has been shown in the rabbit retina (Syed et al., 2004). Stage II waves are the most thoroughly studied and are driven by coordinated activity of a particular class of amacrine cell, the interconnected cholinergic starburst amacrine cells, which undergo spontaneous depolarization in the rabbit retina (Zheng et al., 2006) and release acetylcholine and GABA onto other starburst amacrine cells and RGCs (Wang et al., 2007; Zheng et al., 2006). Stage III waves are propagated by activation of ionotropic glutamate receptors and appear as bipolar cells begin to mature and form synapses (Bansal et al., 2000; Wong et al., 2000).
In mice, the current model for retinal wave development suggests that stage I waves begin at E16 and continue through birth (Bansal et al., 2000). Unlike the stage I waves described in rabbit, cholinergic signaling appears to play a role in stage I retinal waves in mice, as nicotinic acetylcholine receptor antagonists inhibit propagation of large-area Ca2+ waves across the retina at embryonic ages (Bansal et al., 2000). Smaller, local areas of correlated Ca2+ increases persist in the presence of nicotinic acetylcholine antagonists and might be explained by intercellular signaling via gap junctions, but studies confirming this have not yet been performed in mice. Stage II waves in mice occur from P0 until P11 and can be completely inhibited by blocking cholinergic transmission with curare, a nicotinic acetylcholine receptor antagonist (Bansal et al., 2000). RGCs undergo bursts of action potentials during these retinal waves as early as P0, and postsynaptic currents have been recorded from starburst amacrine cells and ganglion cells (Mooney et al., 1996; Wang et al., 2007). Thus, although Olney (1968) indicates that evidence of synapses in the IPL prior to P2 was equivocal, electrophysiology studies indicate that RGCs and amacrine cells are both innervated by amacrine cells by P0, the earliest age examined. Stage III waves in mice occur from P12 until P20 and are completely blocked by ionotropic glutamate receptor antagonists (Bansal et al., 2000), indicating that, as with the ferret retina (Wong et al., 2000), bipolar cells play a functional role in driving stage III retinal waves.
In summary, amacrine cell to RGC connections appear to be the first functional contacts in the retina. Electrophysiology data indicate that the amacrine-to-RGC synapses are functional at P0 (closed circles in Fig. 1). Ca2+ imaging data show correlated cholinergic-dependent Ca2+ increases at E16, but studies have not yet confirmed whether RGCs are capable of spiking prior to P0. On and OFF RGC responses recorded at P10 indicate that all synaptic connections between photoreceptors and RGCs are functional by P10, which we indicate with closed circles in figure 1.
Functional Connectivity between RGCs and LGN Neurons
Mooney et al. (1996) confirmed that RGC axons functionally innervate the LGN as early as P0 by inhibiting stimulus-evoked RGC activity in LGN neurons with local application of lidocaine on the optic nerve. Interestingly, correlated periodic bursts of activity were also recorded from the optic nerve and LGN neurons in mice as early as P0. These recordings indicate that the RGCs are capable of generating and conducting action potentials at this age. Therefore, we indicate by solid circles in figure 1 the earliest age at which connection from RGCs to the LGN are established to be functional. The robust response at P0 suggests that synaptogenesis occurs at embryonic ages, but additional studies are necessary to determine when functional communication between RGCs and LGN neurons initiates.
Functional Connectivity Between LGN and Visual Cortex
In the visual cortex of rats, correlated bursts of activity, termed spindle bursts, have been recorded using extracellular multi-electrode arrays and whole-cell voltage clamp techniques. These bursts are highly correlated with retinal waves and can be evoked upon direct stimulation of the optic nerve. Spindle bursts, evoked either by retinal waves or optic nerve stimulation, occur as early as, but not prior to, P2 (Hanganu et al., 2006). Although similar experiments have not been performed in mice, functional connectivity during the first postnatal week is inferred from experiments where genetic or pharmacological blockade of retinal waves during the first postnatal week inhibits proper mapping of LGN projections in the visual cortex of mice, as assayed at P8 (Cang et al., 2005). Therefore, LGN directed activity likely occurs during the first postnatal week in mice, but when this activity initiates remains unknown. Based on the available data for rat, we estimate connections between LGN and the visual cortex are functional at P2 (closed circles in Fig. 1).
4. Olfactory System
Olfaction initiates in the nasal epithelium where combinations of olfactory sensory neurons (OSNs) reside, each encoding a single receptor subtype that selectively binds to a particular odorant molecule (Bozza et al., 2002; Buck and Axel, 1991; Chess et al., 1994). The mouse genome encodes approximately 1200 different OSN receptor types for OSNs. Although OSN cell types are relatively scattered across 4 epithelial zones, OSN axons are targeted by individual receptor type to particular mitral and tufted cell dendrite groups in the CNS at specialized junctional zones known as glomeruli (Miyamichi et al., 2005; Mombaerts et al., 1996; Ressler et al., 1993). In contrast to the auditory and visual systems, the olfactory nerve projects to CNS targets directly from its peripheral receptors, such that the first olfactory synapse is comparable to initial CNS targets of the other systems. A single glomerulus therefore constitutes the first synaptic relay of the olfactory system, and consists of an anatomically distinct region of axon terminations of same-receptor OSNs from the periphery together with local mitral/tufted (MT) cell dendrites. MT cells are the only long projecting neurons of the olfactory bulb. Among other CNS targets, MT cells innervate the primary olfactory cortex, comprised of the periamygdaloid area and piriform cortex (Hinds, 1968; Price, 1973; Schwob and Price, 1984). The three layer piriform cortex connects peripheral olfactory signals to diverse regions, and may function as an associative area for olfactory information coding (Haberly, 2001). In the olfactory system, our focus is: (1) the first synaptic relay between OSN axons and MT cells at the glomeruli of the olfactory bulb, and (2) MT connectivity with the piriform cortex layer 1a via the lateral olfactory tract. Note that these connections correspond to the second and third connections in the auditory and visual systems. In order to more accurately generalize our results across a diversity of studies, we have excluded separate discussion of tufted cells from our analyses, because reports of their exclusive connectivity and function apart from mitral cells are lacking.
Histogenesis
Neurons supporting early synaptic relays of the mouse olfactory system arise in a relatively parallel fashion (Fig. 1). OSN, MT cells, and primary olfactory cortical cells all share initial appearance between E9.5 and E10.5 (Blanchart et al., 2006; Gardette et al., 1982; Hinds and Angevine, 1965; Hinds, 1968). Peak histogenesis of mitral cells and piriform cortical neurons are also similar, appearing in greatest number at E12. Histogenesis is complete before birth for mitral (E16), tufted (E18), and their pyramidal cell targets of the piriform cortex (E14.5); however, OSNs have persistent histogenesis (Gardette et al., 1982; Hinds and Angevine, 1965; Hinds, 1968).
Anatomical connectivity between olfactory sensory neurons and mitral/tufted cells; development of glomeruli
Formation of the first olfactory projection relay occurs following simultaneous entry and synaptogenesis between OSN axons and newly formed dendrites of MT cells within the glomerular layer of the olfactory bulb at E15–16 (Blanchart et al., 2006; Blanchart et al., 2008; Hinds and Hinds, 1976). Formation of glomerular-like structures, at this stage termed “protoglomeruli” because they lack encapsulating cell layers, is achieved by E17 as assayed by light microscopy techniques (Blanchart et al., 2006). Electron microscopic confirmation of synaptic contacts between OSN and MT cells at E15 is coincident with the developing protoglomerulus at E15–16 (Fig. 1, open circles; Blanchart et al., 2008; Hinds and Hinds, 1976).
Anatomical connectivity between Mitral/Tufted cells and primary olfactory cortex
Development and outgrowth of the lateral olfactory tract (LOT) from MT cells, the second olfactory relay, is apparent at E11.5 (Walz et al., 2006), soon after the onset of histogenesis of OSNs. Terminations of the LOT are first revealed by light microscopic observation within the piriform cortex, proximal to piriform dendrites, at E16 (Fig. 1, open circles; Walz et al., 2006). EM observations confirm the absence of synapses between pyramidal cells of olfactory cortex and the LOT at E15, but later ages were not examined (Derer et al., 1977). In the rat, EM examination revealed that piriform layer 1a synapses first appear infrequently at E19, coincident with E19 formation of glomeruli in rat (Schwob and Price, 1984), increasing in density with age to almost 40% of adult levels by P10 (Moriizumi et al., 1995).
Functional connectivity between olfactory sensory neurons and mitral/tufted cells; activation of glomeruli
Few studies have examined activity-related properties of prenatal olfactory bulb neurons, and sensory deprivation studies have been limited to postnatal windows, well after initial circuit and synapse establishment (Benson et al., 1984; Brunjes, 1994). The earliest observation of mitral cell activity is at P0, with mitral cells already exhibiting selective excitation by particular odorants in rats (Mair and Gesteland, 1982). Distinct glomerular regions are activated in response to odorants immediately following birth (<12 hours), as measured by activation of the immediate early gene, c-fos, in rat (Fig. 1, closed circles; Guthrie and Gall, 2003). Single-unit action potentials of OSNs in rat can occur infrequently as generalized odorant responsiveness first at E16, about 3 days prior to glomerular formation in rat, but become more specific and pronounced by E19 (Gesteland et al., 1982). If OSN activation in mouse occurs in a temporally parallel fashion, initial activity would be present at about E13–14 (3 days before mouse glomerular formation).
Functional connectivity between mitral/tufted cells and primary olfactory cortex
Investigations of functional connectivity between the olfactory bulb and olfactory cortex in mice have not been performed, but studies in rat have been directed towards this question. The earliest reported olfactory associated activity in the piriform cortex was observed by Schwob et al. (1984), who recorded monosynaptic EPSP waveforms in the piriform cortex of rats at P0 in response to LOT stimulation (closed circles in Fig 1). Therefore, anatomical and functional connectivity between MT cells and primary olfactory cortex are in place by birth. Experience-driven responsiveness in the primary olfactory cortex has been implicated as early as P3 in rats (the earliest age tested), with activity of c-fos enhanced in stereotypic patterns of the anterior piriform cortex in response to odorant stimulation trials (Illig, 2007).
5. Unresolved Issues in Sensory System Development
The title of this article posed the question: Does the brain connect before the periphery can direct? Multiple observations might suggest it does not. Soon after final mitosis, many neurons extend an axon toward their target even as their cell body migrates toward its final position in the brain (Yee et al., 1999). Axon guidance cues then position the axon growth cone into an approximate location to innervate its target (Tessier-Lavigne and Goodman, 1996). The role of neurotransmitter release in synapse formation has been addressed by preventing vesicle fusion. Knockout of munc 13-1/2 proteins involved in exocytosis reveals formation of synapses in culture in the absence of neurotransmitter release (Varoqueaux et al., 2002). In a munc 18-1 knockout mouse, synaptic contacts in certain regions of the brain, including cerebellar cortex, initially form then degenerate quickly (Verhage et al., 2000). These studies suggest that axon guidance and synapse formation occur independently from neurotransmitter release. However, the sensory systems described here were not evaluated in these studies. Indeed, the munc 18-1 knockout exhibits severe brainstem degeneration by E14.5, so auditory system neurons may die prior to formation of synaptic contacts. Therefore, conclusions from these studies should be determined separately for each neural system. In addition, assembly of pre- or postsynaptic structures may occur before cells are capable of generating action potentials. Alternatively, communication between cells, especially at the earliest ages, need not occur via classical synaptic junctions. For example, one could speculate that membrane-associated or secreted factors could induce signal transduction cascades that signal along dendrites and axons to affect other cells. In sensory systems, then, the signals to form contacts could be dependent on peripherally generated signals that are transmitted either synaptically or non-synaptically. In the following sections we summarize the relevant connectivity and functional data for each sensory system then compare features across systems. We conclude with consideration how genetic models for receptor cell ablation or silencing have been or could be utilized to shed light upon this issue.
Auditory Circuit Formation
In the auditory system, light microscopic evidence (open circles in Fig. 1) suggests that the two central synapses considered here could form simultaneously at E15.5, and innervation of the VCN and the SOC, especially ipsilateral innervation of the LSO, may occur even earlier. Synaptic connections between hair cells and spiral ganglion cells may form up to one day later at E16.5. Therefore, the brain may form initial contacts prior to formation of contacts in the periphery. Ultrastructural evidence for cellular contacts, including synaptic contacts, in mice is lacking at embryonic ages. These estimations of the earliest age at which information could flow from the periphery to the CNS do not assume that the nature of information transmission need be electrical or through conventional synapses. As with EM studies, functional assays are limited, for the most part, to postnatal ages (solid circles in Fig. 1). The only functional evidence for synaptic activity at embryonic ages demonstrates synaptic currents in MNTB neurons at E17. Even when synaptic connections are functional, cells may be unable to generate and propagate action potentials as the earliest evidence that spiral ganglion and VCN neurons can generate action potentials is at P1. Due to experimental difficulty, the earliest age at which the hair cell to spiral ganglion synapse has been studied is P7. Hence, this is the age at which we indicate experimental evidence supports the conclusion that neural activity can propagate from the periphery into the brain (double red bars in Fig. 1). Investigation of synaptic ultrastructure and function at embryonic ages, then, will fill important gaps in our understanding of early circuit formation for the auditory pathway.
Despite the potentially later age at which the hair cell to spiral ganglion cell synapse forms, cochlear-induced activity may originate in supporting and ganglion cells at earlier ages in a manner analogous to retinal Ca2+ waves. Prior to the onset of sensitivity to airborne sound in rats, spontaneous currents in a transient supporting cell population, called Kolliker’s Organ, correlate with inward currents in inner hair cells and excitatory postsynaptic currents in spiral ganglion cell dendrites. Pharmacologic manipulation indicates that a likely mechanism is inner hair cell activation by ATP release from Kolliker’s Organ followed by synaptic activation of spiral ganglion neurons. These phenomena have been described in rats for ages P7–10 (Tritsch et al., 2007), but have not been explored at earlier ages, especially during embryogenesis. These waves, too, should be investigated at ages prior to E16.5, because such mechanisms may provide a substrate to guide central contact formation from the periphery before hair cell-to-spiral ganglion synapses form.
Visual Circuit Formation
We consider P10 to be the earliest age at which activity in the extreme periphery could be relayed to the visual cortex (double bars in Fig. 1) based on the following functional data: (1) Stimulation of the optic nerve evokes responses in the LGN at P0, (2) stimulation of the optic nerve evokes activity in the visual cortex of rats at P2, and (3) photic stimulation of photoreceptors produces ON and OFF responses in RGCs at P10. Light microscopy studies indicate that a receptor-to-cortical circuit could be complete when the first bipolar cells form synapses onto RGCs at P8, 12 days after the LGN and cortex initially receive their afferent projections (open circles in Fig. 1). Unlike the auditory and olfactory systems, connections in the visual system, particularly in the retina are established over a protracted period during postnatal ages. However, the connections to more central visual centers considered here may form simultaneously at embryonic ages, similar to the auditory and olfactory systems. Note however, that these critical time points for anatomical connectivity were determined using only light microscopy. EM data for the visual system therefore is lacking, likely because proper identification of synapses at ultrastructural resolution is difficult and requires labeling of presynaptic neurons.
The earliest demonstration of retinal function at P10 (Tian and Copenhagen, 2003) coincides with the emergence of bipolar cell ribbon synapses in the IPL and anatomical observations of rod synapse formation in the OPL (Olney, 1968). Although the complete outer-to-inner retina circuit is not complete until P8 or later, Ca2+ imaging of retinal wave activity suggests that amacrine and ganglion cells, both early-born cell fates, may form connections by E16, coincident with RGC innervation of the LGN. Although it is currently unknown which cell types initiate and/or propagate these early waves in mice, blocking nicotinic acetylcholine receptors attenuates propagation of large area waves throughout the retina (Bansal et al., 2000) and implies that some form of intercellular communication occurs at these early ages. It is possible that the smaller, local regions of Ca2+ waves that persist during cholinergic blockade are propagated through gap junctions, like those observed in rabbit (Syed et al., 2004), and the waves that require cholinergic transmission utilize chemical synapses that are sufficiently sparse to make them difficult to detect. What role might stage I waves play prior to birth? If RGCs can fire action potentials at E16, then these waves may provide a mechanism for retinal neurons to influence initial synapse formation in the LGN. However, if the signals from the waves of activity in the retina are not communicated beyond the LGN to the cortex until postnatal ages, as in rat, then stage I retinal waves may be instructive only for connections within the LGN. Additional studies are necessary to determine if and how amacrine and ganglion cells communicate at the time of stage I waves and whether RGCs are capable of firing action potentials to relay information to the LGN prior to birth.
Olfactory Circuit Formation
Dating for anatomic connectivity in the olfactory system appears well established for the circuit considered here. Light and electron microscopy studies indicate that a complete anatomical circuit exists as early as E16 from OSN to piriform cortex (open circles in Fig. 1). Within the CNS a sequential, peripheral-to-central, pattern for connectivity is observed for the olfactory system, with initial connectivity between OSNs and MT cells at E15 (Blanchart et al., 2006; Hinds and Hinds, 1976), and initial connectivity between LOT and primary olfactory cortex at E16 (Moriizumi et al., 1995; Schwob and Price, 1984). Therefore, anatomical evidence is consistent with the possibility that peripheral activity could play an instructive role during synaptogenesis at both glomeruli and primary olfactory cortex.
Functional connectivity has not been explored for prenatal ages in the olfactory system; therefore the earliest time points for active circuit maturation remain uncertain. OSN activity in rat is initiated 3 days prior to glomerular formation, the equivalent of E13–14 in mouse (Gesteland et al., 1982). Functional connectivity between OSNs and MT cells (Guthrie and Gall, 2003; Mair and Gesteland, 1982), activity of MT cells (Mair and Gesteland, 1982), and responsiveness of piriform cortex to LOT stimulation (Schwob et al., 1984), have all been demonstrated first at P0 (solid black circles and double red bars in Fig. 1). Together, this evidence indicates that by birth, the essential ingredients are in place for rudimentary olfactory sensory experience. EM data demonstrate that synapses are present in glomeruli at E15 (Blanchart et al., 2008; Hinds and Hinds, 1976). Extrapolating from rat EM data, the LOT forms synapses with piriform pyramidal cells at E16–E17 (Moriizumi et al., 1995). Because anatomical evidence of connectivity includes ultrastructural descriptions of synapses, dating for complete functional maturation of the olfactory system could be moved to E16, provided OSNs and MTs are capable of action potential generation and conduction. Functional maturation of the olfactory circuitry at E16 strengthens the prospect of peripheral activation driving central synaptogenesis before birth. Indeed, evidence for prenatal olfactory sensation has been described in several behavioral studies. Presentation of odorant chemicals to late embryonic rats (E19–20) increases activity and facial wiping behaviors (Smotherman and Robinson, 1990), and olfactory bulb transection at E19–20 greatly reduces responsiveness to a chemical stimulus (Smotherman et al., 1987). These behavioral data are consistent with the hypothesis that rodent OSNs are capable of chemical sensation and can transmit that information through central pathways slightly before birth. Regardless, it is clear that early postnatal olfactory experience is critical to support proper infant feeding behaviors and survival (for review see: Leon, 1992; Sullivan, 2003), therefore consistent with rudimentary olfactory sensation present at least by birth.
Given evidence of early wave-like activity from both the auditory and visual systems that could be involved at the periphery in directing early patterning, it is tempting to speculate that such a mechanism could exist at the olfactory epithelium as well. Recently, Ca2+ waves have been observed in sustentacular cells in the olfactory epithelium (Hegg et al., 2008). Sustentacular cells form a layer at the apical surface of the olfactory epithelium and may serve to provide structural, functional, and trophic support for OSNs (Getchell and Getchell, 1992). Spontaneous calcium waves in the olfactory epithelium were observed as early as P0 in mice. These waves appear to depend on connexin hemichannels that are responsive to ATP (Hegg et al., 2008; Vogalis et al., 2005). Although sustentacular cell hemichannels do not appear to be developmentally regulated and persist into adulthood, it is possible that they could have distinct developmental roles at prenatal ages relating to olfactory circuit formation, a possibility that has not been examined.
Comparisons among sensory systems
For all three systems, evidence for functional connections at each relay along the sensory pathway was collected primarily at postnatal ages (solid circles and double-bar symbols in Fig. 1). From these functional data, then, it is not possible to determine the temporal order of circuit formation or dependence upon upstream elements. These data highlight, rather, the importance of conducting physiological studies at early, especially embryonic, ages for all sensory systems. Considering light microscopy information available from the auditory and visual systems (open circles in Fig. 1), two central connections that we consider here may form simultaneously and prior to peripheral connections with their ganglion cells. The olfactory system differs in that the main input to mitral cells may form prior to innervation of cortex. For the olfactory system, connectivity information is based on ultrastructural studies, so the anatomical template for synaptic connectivity has been established. These data highlight the importance of conducting ultrastructural studies of the visual and auditory systems at embryonic ages. One caveat to these inferred temporal sequences is that the temporal resolution of most embryonic studies occurs at one-day intervals, so conclusions distinguishing precedence from simultaneity must be tempered by this consideration.
An emerging theme from the systems compared here is that initial formation of functional circuitry may occur well before each sensory system encounters appropriate environmental stimuli. For the auditory system functional contacts could begin to form 10–11 days before ear canal opening. In the visual system, connections from cone photoreceptors through bipolar to ganglion cells are in place at least 6 days prior to eye opening. In the olfactory system, functional activity from the periphery to the cortex could be in place about 3.5 days before birth, as early as E16, when mouse appropriate ages are extrapolated from rat EM and neuronal activation evidence. The establishment of circuitry across sensory systems of the mouse several days prior to their first use may indicate a need for protracted development of the basic topography of circuits in the absence of normal environmental stimulation and perhaps a margin of safety, especially for the olfactory system, for functional sensation in the event of early birth.
Given the absence of physiological data at embryonic ages, consider that, at least for the auditory and visual systems, the relay of instructive signals from one cell to the next may not require synaptic transmission and propagation of action potentials along axons during initial circuit formation (open circles in Fig. 1). Instead, information transfer may only require cells to be in proximity to one another or physically in contact, and that their axons extend to the next relay station. In this case, cells may communicate via receptor-mediated intracellular signal transduction cascades and axonal transport. Such a scenario could reverse the implied order of events for the auditory system if ganglion cells are activated directly, for example by ATP release from Kolliker’s Organ. ATP-generated waves would have to occur at E15.5 or earlier in order to activate spiral ganglion neurons directly and induce formation of central connections. Purinergic receptors are present on spiral ganglion neurons at E18, but earlier ages have not been investigated (Huang et al., 2006). In the visual system, the onset of stage I retinal waves in mice at E16, an age coincident with the potential formation of initial circuits in the LGN and visual cortex, could activate signal transduction pathways or induce gene expression in ganglion cells. Subsequent transport of signals further into the CNS might induce formation of these more central connections.
Genetic techniques have been applied in these sensory systems to silence or ablate particular cell types to determine their role in subsequent central circuit formation. Because initial formation of sensory circuits considered here occurs by the beginning of the third postnatal week, later postnatal assessments of central circuitry may overlook key events in the initial formation of neural connections. Below we provide a brief overview of the different strategies that have been utilized to examine the necessity of more peripheral signaling in central areas of auditory, visual, and olfactory systems.
In the auditory system, perhaps the best means for silencing hair cell activity is through knockout of its L-type Ca+ channel (Brandt et al., 2003). In this mutant model, inner hair cell synapses form normally and presynaptic inputs to the MNTB are functional, prior to postnatal degeneration of spiral ganglion cells (Erazo-Fischer et al., 2007; Glueckert et al., 2003; Nemzou N et al., 2006). Although these observations imply activity-independent formation of this auditory circuit, it should be noted that hair cells are not completely silenced in L-type Ca+ channel knockouts. Low levels of spontaneous vesicle release remain and non-synaptic vesicle secretion may occur (Brandt et al., 2003). Knockout of otoferlin, a hair cell protein that forms part of the SNARE complex for vesicle release, also is an effective way to reduce synaptic activity (Beurg et al., 2008; Roux et al., 2006). However, some vesicle release does occur in this animal. Hair cells but not brainstem auditory neurons utilize the vesicular glutamate transporter isoform 3 (VGLUT3). The VGLUT3 knockout mouse lacks glutamate release, measured as early as P6, and has altered synaptic ribbon morphology (Seal et al., 2008). Therefore, it may currently be the best model to remove synaptically-induced activity in spiral ganglion neurons. Note that synaptic vesicle fusion could occur in this animal, with release of other vesicle constituents.
The importance of non-synaptic communication can be assessed in animals whereby circuit elements, such as receptor cells, are prevented from forming. Hair cells can be genetically ablated with preservation of spiral ganglion neurons with the knockout of Math1, a transcription factor necessary for hair cell formation (Bermingham et al., 1999; Chen et al., 2002; Woods et al., 2004). Unfortunately, the VCN fails to form in these animals (Wang et al., 2005), so the central connections considered here cannot be tested. These observations point out the importance of evaluating more central stations along the sensory pathway following genetic elimination of circuit elements.
Genetic silencing of the retinal ganglion cell does not appear to have been achieved to date. In contrast, there are many photoreceptor dystrophies and degenerations in the mouse, but it is unclear if these mutants would be sufficient to block the spontaneous signaling and trophic support that RGCs exhibit (Galli-Resta et al., 1993; Thurlow and Cooper, 1988). Instead, genetic ablations of the RGCs or the optic nerve appear to be the most relevant models to address the issue of retinal neurons instructing the central circuits. A new genetic model, where targeted mutations in both Math5 and Brn3b cause near complete ablation of RGCs and loss of the optic nerve (Moshiri et al., 2008), may be a useful tool for such studies, but the projections from LGN to visual cortex have not been examined in this strain to our knowledge. Studies have been performed on the central visual connections in the eyeless mouse strain, ZRDCT-An (Godement et al., 1979; Kaiserman-Abramof et al., 1980). Both groups found that despite the lack of an optic nerve, projections from the LGN to the visual cortex (area 17) were intact, though slightly reduced. These findings support the case for the periphery not being required for central circuit formation in the visual system.
In the olfactory system, genetic ablation of OSNs or disruption of olfactory nerve innervation dramatically disrupts olfactory bulb organization and prevents glomerular formation (Jimenez et al., 2000; Kobayakawa et al., 2007; Long et al., 2003). However, when OSN activity was largely silenced through mutation of the α subunit of the olfactory cyclic nucleotide-gated channel, olfactory nerve patterning and glomerular organization were preserved (Belluscio et al., 1998; Lin et al., 2000). Taken together, these studies suggest that while physical presence of peripheral olfactory afferents might be required for normal patterning, activity does not play an instructive role. Abolition of virtually all OSN action potentials by targeted and inducible expression of the potassium channel Kir2.1 resulted in greatly delayed glomerular formation (P12) and disrupted specificity of glomerular patterning (Yu et al., 2004). Therefore, spontaneous activity provided by OSN axons is thought to serve as a permissive cue for glomerular wiring, but instructive for innervation timing, detailed patterning, and perhaps supportive of other determinative axonal targeting mechanisms, such as OSN receptor type and level of expression (Biju et al., 2008; Wang et al., 1998; Zou et al., 2004), ephrin A (Cutforth et al., 2003), or semaphorin interaction (Lattemann et al., 2007). To date, no studies, including silenced or ablated OSNs, have addressed the dependence of MT cell input via the LOT on initial olfactory cortical development, allowing the possibility that synaptogenesis between MT axons of the LOT and their piriform targets may require peripheral direction. Future work, including efforts to prevent even spontaneous vesicle release events from before the time of birth, will provide the best assessment for the role of synaptic activity in driving peripheral-to-central circuit formation. These studies should be performed on embryonic animals, and can conveniently be performed in transgenic and knockout lines that do not survive postnatally.
In summary, with regard to the question posed in the title, this review highlights two general research topics that require further investigation. First, ultrastructural and physiological investigations should be extended in a systematic fashion to embryonic ages in the mouse. Emphasis should be placed on determining when cells can generate and transmit action potentials, which determines whether neural activity involves only local synaptic transmission or communication among neural stations. Second, better temporal resolution should be employed in experimental designs of early circuit formation, preferably using litter-mate controls. Although individual laboratories tend to focus efforts on their particular system of interest, study designs that facilitate comparisons across systems in mice will further increase the value of this species as an animal model.
Acknowledgments
Portions of this work were supported by National Institutes of Health (NIH) grants F32 DC008730 to BH, RO1 DC007695 to GS, RO1 EY012152 to PM and a NIH/NCRR COBRE grant P20 RR15574 to the Sensory Neuroscience Research Center.
List of Abberviations
- CNS
central nervous system
- EM
electron microscopy
- LGN
lateral geniculate nucleus
- LOT
lateral olfactory tract
- LSO
lateral superior olive
- MNTB
medial nucleus of the trapezoid body
- MT
mitral/tufted
- OSN
olfactory sensory neuron
- RGC
retinal ganglion cell
- SOC
superior olivary complex
- VCN
ventral cochlear nucleus
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Angevine JB., Jr Time of neuron origin in the diencephalon of the mouse. An autoradiographic study. J Comp Neurol. 1970;139:129–87. doi: 10.1002/cne.901390202. [DOI] [PubMed] [Google Scholar]
- Bansal A, Singer JH, Hwang BJ, Xu W, Beaudet A, Feller MB. Mice Lacking Specific Nicotinic Acetylcholine Receptor Subunits Exhibit Dramatically Altered Spontaneous Activity Patterns and Reveal a Limited Role for Retinal Waves in Forming ON and OFF Circuits in the Inner Retina. J Neurosci. 2000;20:7672–7681. doi: 10.1523/JNEUROSCI.20-20-07672.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belluscio L, Gold GH, Nemes A, Axel R. Mice deficient in G(olf) are anosmic. Neuron. 1998;20:69–81. doi: 10.1016/s0896-6273(00)80435-3. [DOI] [PubMed] [Google Scholar]
- Benson TE, Ryugo DK, Hinds JW. Effects of sensory deprivation on the developing mouse olfactory system: a light and electron microscopic, morphometric analysis. J Neurosci. 1984;4:638–53. doi: 10.1523/JNEUROSCI.04-03-00638.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermingham NA, Hassan BA, Price SD, Vollrath MA, Ben-Arie N, Eatock RA, Bellen HJ, Lysakowski A, Zoghbi HY. Math1: an essential gene for the generation of inner ear hair cells. Science. 1999;284:1837–41. doi: 10.1126/science.284.5421.1837. [DOI] [PubMed] [Google Scholar]
- Beurg M, Safieddine S, Roux I, Bouleau Y, Petit C, Dulon D. Calcium- and otoferlin-dependent exocytosis by immature outer hair cells. J Neurosci. 2008;28:1798–803. doi: 10.1523/JNEUROSCI.4653-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beutner D, Voets T, Neher E, Moser T. Calcium dependence of exocytosis and endocytosis at the cochlear inner hair cell afferent synapse. Neuron. 2001;29:681–90. doi: 10.1016/s0896-6273(01)00243-4. [DOI] [PubMed] [Google Scholar]
- Biju KC, Marks DR, Mast TG, Fadool DA. Deletion of voltage-gated channel affects glomerular refinement and odorant receptor expression in the mouse olfactory system. J Comp Neurol. 2008;506:161–79. doi: 10.1002/cne.21540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanchart A, De Carlos JA, Lopez-Mascaraque L. Time frame of mitral cell development in the mice olfactory bulb. J Comp Neurol. 2006;496:529–43. doi: 10.1002/cne.20941. [DOI] [PubMed] [Google Scholar]
- Blanchart A, Romaguera M, Garcia-Verdugo JM, de Carlos JA, Lopez-Mascaraque L. Synaptogenesis in the mouse olfactory bulb during glomerulus development. Eur J Neurosci. 2008;27:2838–46. doi: 10.1111/j.1460-9568.2008.06283.x. [DOI] [PubMed] [Google Scholar]
- Blanks JC, Adinolfi AM, Lolley RN. Synaptogenesis in the photoreceptor terminal of the mouse retina. J Comp Neurol. 1974;156:81–93. doi: 10.1002/cne.901560107. [DOI] [PubMed] [Google Scholar]
- Bozza T, Feinstein P, Zheng C, Mombaerts P. Odorant receptor expression defines functional units in the mouse olfactory system. J Neurosci. 2002;22:3033–43. doi: 10.1523/JNEUROSCI.22-08-03033.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandt A, Striessnig J, Moser T. CaV1.3 Channels Are Essential for Development and Presynaptic Activity of Cochlear Inner Hair Cells. J Neurosci. 2003;23:10832–10840. doi: 10.1523/JNEUROSCI.23-34-10832.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunjes PC. Unilateral naris closure and olfactory system development. Brain Res Brain Res Rev. 1994;19:146–60. doi: 10.1016/0165-0173(94)90007-8. [DOI] [PubMed] [Google Scholar]
- Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991;65:175–87. doi: 10.1016/0092-8674(91)90418-x. [DOI] [PubMed] [Google Scholar]
- Cang J, Renteria RC, Kaneko M, Liu X, Copenhagen DR, Stryker MP. Development of precise maps in visual cortex requires patterned spontaneous activity in the retina. Neuron. 2005;48:797–809. doi: 10.1016/j.neuron.2005.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cant NB. Structural Development of the Mammalian Auditory Pathways. In: Rubel EW, Popper AN, Fay RR, editors. Development of the auditory system. Springer handbook of auditory research. Vol. 9. Springer; New York: 1998. pp. 315–413. [Google Scholar]
- Cepko CL, Austin CP, Yang X, Alexiades M, Ezzeddine D. Cell fate determination in the vertebrate retina. Proc Natl Acad Sci U S A. 1996;93:589–95. doi: 10.1073/pnas.93.2.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen P, Segil N. p27(Kip1) links cell proliferation to morphogenesis in the developing organ of Corti. Development. 1999;126:1581–90. doi: 10.1242/dev.126.8.1581. [DOI] [PubMed] [Google Scholar]
- Chen P, Johnson JE, Zoghbi HY, Segil N. The role of Math1 in inner ear development: Uncoupling the establishment of the sensory primordium from hair cell fate determination. Development. 2002;129:2495–2505. doi: 10.1242/dev.129.10.2495. [DOI] [PubMed] [Google Scholar]
- Chess A, Simon I, Cedar H, Axel R. Allelic inactivation regulates olfactory receptor gene expression. Cell. 1994;78:823–34. doi: 10.1016/s0092-8674(94)90562-2. [DOI] [PubMed] [Google Scholar]
- Cutforth T, Moring L, Mendelsohn M, Nemes A, Shah NM, Kim MM, Frisen J, Axel R. Axonal ephrin-As and odorant receptors: coordinate determination of the olfactory sensory map. Cell. 2003;114:311–22. doi: 10.1016/s0092-8674(03)00568-3. [DOI] [PubMed] [Google Scholar]
- Del Rio JA, Martinez A, Auladell C, Soriano E. Developmental History of the Subplate and Developing White Matter in the Murine Neocortex. Neuronal Organization and Relationship with the Main Afferent Systems at Embryonic and Perinatal Stages. Cereb Cortex. 2000;10:784–801. doi: 10.1093/cercor/10.8.784. [DOI] [PubMed] [Google Scholar]
- Derer P, Caviness VS, Jr, Sidman RL. Early cortical histogenesis in the primary olfactory cortex of the mouse. Brain Res. 1977;123:27–40. doi: 10.1016/0006-8993(77)90641-2. [DOI] [PubMed] [Google Scholar]
- Ene FA, Kullmann PHM, Gillespie DC, Kandler K. Glutamatergic Calcium Responses in the Developing Lateral Superior Olive: Receptor Types and Their Specific Activation by Synaptic Activity Patterns. J Neurophysiol. 2003;90:2581–2591. doi: 10.1152/jn.00238.2003. [DOI] [PubMed] [Google Scholar]
- Erazo-Fischer E, Striessnig J, Taschenberger H. The Role of Physiological Afferent Nerve Activity during In Vivo Maturation of the Calyx of Held Synapse. J Neurosci. 2007;27:1725–1737. doi: 10.1523/JNEUROSCI.4116-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feller MB, Wellis DP, Stellwagen D, Werblin FS, Shatz CJ. Requirement for cholinergic synaptic transmission in the propagation of spontaneous retinal waves. Science. 1996;272:1182–7. doi: 10.1126/science.272.5265.1182. [DOI] [PubMed] [Google Scholar]
- Feller MB, Butts DA, Aaron HL, Rokhsar DS, Shatz CJ. Dynamic processes shape spatiotemporal properties of retinal waves. Neuron. 1997;19:293–306. doi: 10.1016/s0896-6273(00)80940-x. [DOI] [PubMed] [Google Scholar]
- Fisher LJ. Development of synaptic arrays in the inner plexiform layer of neonatal mouse retina. J Comp Neurol. 1979;187:359–72. doi: 10.1002/cne.901870207. [DOI] [PubMed] [Google Scholar]
- Galli-Resta L, Ensini M, Fusco E, Gravina A, Margheritti B. Afferent spontaneous electrical activity promotes the survival of target cells in the developing retinotectal system of the rat. J Neurosci. 1993;13:243–50. doi: 10.1523/JNEUROSCI.13-01-00243.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardette R, Courtois M, Bisconte JC. Prenatal development of mouse central nervous structures: time of neuron origin and gradients of neuronal production. A radioautographic study. J Hirnforsch. 1982;23:415–31. [PubMed] [Google Scholar]
- Gesteland RC, Yancey RA, Farbman AI. Development of olfactory receptor neuron selectivity in the rat fetus. Neuroscience. 1982;7:3127–36. doi: 10.1016/0306-4522(82)90235-4. [DOI] [PubMed] [Google Scholar]
- Getchell ML, Getchell TV. Fine structural aspects of secretion and extrinsic innervation in the olfactory mucosa. Microsc Res Tech. 1992;23:111–27. doi: 10.1002/jemt.1070230203. [DOI] [PubMed] [Google Scholar]
- Glowatzki E, Fuchs PA. Transmitter release at the hair cell ribbon synapse. Nat Neurosci. 2002;5:147–54. doi: 10.1038/nn796. [DOI] [PubMed] [Google Scholar]
- Glueckert R, Wietzorrek G, Kammen-Jolly K, Scholtz A, Stephan K, Striessnig J, Schrott-Fischer A. Role of class D L-type Ca2+ channels for cochlear morphology. Hearing Research. 2003;178:95–105. doi: 10.1016/s0378-5955(03)00054-6. [DOI] [PubMed] [Google Scholar]
- Godement P, Saillour P, Imbert M. Thalamic afferents to the visual cortex in congenitally anophthalmic mice. Neurosci Lett. 1979;13:271–8. doi: 10.1016/0304-3940(79)91506-4. [DOI] [PubMed] [Google Scholar]
- Godement P, Salaun J, Imbert M. Prenatal and postnatal development of retinogeniculate and retinocollicular projections in the mouse. J Comp Neurol. 1984;230:552–75. doi: 10.1002/cne.902300406. [DOI] [PubMed] [Google Scholar]
- Guthrie KM, Gall C. Anatomic mapping of neuronal odor responses in the developing rat olfactory bulb. J Comp Neurol. 2003;455:56–71. doi: 10.1002/cne.10452. [DOI] [PubMed] [Google Scholar]
- Haberly LB. Parallel-distributed processing in olfactory cortex: new insights from morphological and physiological analysis of neuronal circuitry. Chem Senses. 2001;26:551–76. doi: 10.1093/chemse/26.5.551. [DOI] [PubMed] [Google Scholar]
- Hanganu IL, Ben-Ari Y, Khazipov R. Retinal Waves Trigger Spindle Bursts in the Neonatal Rat Visual Cortex. J Neurosci. 2006;26:6728–6736. doi: 10.1523/JNEUROSCI.0752-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison JM, Warr WB. A study of the cochlear nuclei and ascending auditory pathways of the medulla. J Comp Neurol. 1962;119:341–79. doi: 10.1002/cne.901190306. [DOI] [PubMed] [Google Scholar]
- Hegg CC, Irwin M, Lucero MT. Calcium store-mediated signaling in sustentacular cells of the mouse olfactory epithelium. Glia. 2008 doi: 10.1002/glia.20792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinds JW, Angevine JB. Autoradiographic study of histogenesis in the area pyriformis and claustrum of the mouse. Anatomical Record. 1965;151:456–457. [Google Scholar]
- Hinds JW. Autoradiographic study of histogenesis in the mouse olfactory bulb. I. Time of origin of neurons and neuroglia. J Comp Neurol. 1968;134:287–304. doi: 10.1002/cne.901340304. [DOI] [PubMed] [Google Scholar]
- Hinds JW, Hinds PL. Synapse formation in the mouse olfactory bulb. I. Quantitative studies. J Comp Neurol. 1976;169:15–40. doi: 10.1002/cne.901690103. [DOI] [PubMed] [Google Scholar]
- Hoffpauir BK, Grimes JL, Mathers PH, Spirou GA. Synaptogenesis of the calyx of Held: rapid onset of function and one-to-one morphological innervation. J Neurosci. 2006;26:5511–23. doi: 10.1523/JNEUROSCI.5525-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holt CE, Bertsch TW, Ellis HM, Harris WA. Cellular determination in the Xenopus retina is independent of lineage and birth date. Neuron. 1988;1:15–26. doi: 10.1016/0896-6273(88)90205-x. [DOI] [PubMed] [Google Scholar]
- Hooks BM, Chen C. Critical periods in the visual system: changing views for a model of experience-dependent plasticity. Neuron. 2007;56:312–26. doi: 10.1016/j.neuron.2007.10.003. [DOI] [PubMed] [Google Scholar]
- Howell DM, Morgan WJ, Jarjour AA, Spirou GA, Berrebi AS, Kennedy TE, Mathers PH. Molecular guidance cues necessary for axon pathfinding from the ventral cochlear nucleus. J Comp Neurol. 2007;504:533–49. doi: 10.1002/cne.21443. [DOI] [PubMed] [Google Scholar]
- Huang L-C, Thorne PR, Housley GD, Montgomery JM. Spatiotemporal definition of neurite outgrowth, refinement and retraction in the developing mouse cochlea. Development. 2007;134:2925–2933. doi: 10.1242/dev.001925. [DOI] [PubMed] [Google Scholar]
- Huang LC, Ryan AF, Cockayne DA, Housley GD. Developmentally regulated expression of the P2X3 receptor in the mouse cochlea. Histochem Cell Biol. 2006;125:681–92. doi: 10.1007/s00418-005-0119-4. [DOI] [PubMed] [Google Scholar]
- Huberman AD, Feller MB, Chapman B. Mechanisms underlying development of visual maps and receptive fields. Annu Rev Neurosci. 2008;31:479–509. doi: 10.1146/annurev.neuro.31.060407.125533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Illig KR. Developmental changes in odor-evoked activity in rat piriform cortex. Neuroscience. 2007;145:370–6. doi: 10.1016/j.neuroscience.2006.11.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimenez D, Garcia C, de Castro F, Chedotal A, Sotelo C, de Carlos JA, Valverde F, Lopez-Mascaraque L. Evidence for intrinsic development of olfactory structures in Pax-6 mutant mice. J Comp Neurol. 2000;428:511–26. [PubMed] [Google Scholar]
- Johnson SL, Marcotti W, Kros CJ. Increase in efficiency and reduction in Ca2+ dependence of exocytosis during development of mouse inner hair cells. J Physiol. 2005;563:177–91. doi: 10.1113/jphysiol.2004.074740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaiserman-Abramof IR, Graybiel AM, Nauta WJH. The thalamic projection to cortical area 17 in a congenitally anophthalmic mouse strain. Neuroscience. 1980;5:41–52. doi: 10.1016/0306-4522(80)90069-x. [DOI] [PubMed] [Google Scholar]
- Kandler K, Friauf E. Pre- and postnatal development of efferent connections of the cochlear nucleus in the rat. J Comp Neurol. 1993;328:161–84. doi: 10.1002/cne.903280202. [DOI] [PubMed] [Google Scholar]
- Kandler K, Friauf E. Development of glycinergic and glutamatergic synaptic transmission in the auditory brainstem of perinatal rats. J Neurosci. 1995;15:6890–6904. doi: 10.1523/JNEUROSCI.15-10-06890.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kandler K. Activity-dependent organization of inhibitory circuits: lessons from the auditory system. Curr Opin Neurobiol. 2004;14:96–104. doi: 10.1016/j.conb.2004.01.017. [DOI] [PubMed] [Google Scholar]
- Kobayakawa K, Kobayakawa R, Matsumoto H, Oka Y, Imai T, Ikawa M, Okabe M, Ikeda T, Itohara S, Kikusui T, Mori K, Sakano H. Innate versus learned odour processing in the mouse olfactory bulb. Nature. 2007;450:503–8. doi: 10.1038/nature06281. [DOI] [PubMed] [Google Scholar]
- Koundakjian EJ, Appler JL, Goodrich LV. Auditory Neurons Make Stereotyped Wiring Decisions before Maturation of Their Targets. J Neurosci. 2007;27:14078–14088. doi: 10.1523/JNEUROSCI.3765-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kros CJ, Ruppersberg JP, Rusch A. Expression of a potassium current in inner hair cells during development of hearing in mice. Nature. 1998;394:281–4. doi: 10.1038/28401. [DOI] [PubMed] [Google Scholar]
- Lattemann M, Zierau A, Schulte C, Seidl S, Kuhlmann B, Hummel T. Semaphorin-1a controls receptor neuron-specific axonal convergence in the primary olfactory center of Drosophila. Neuron. 2007;53:169–84. doi: 10.1016/j.neuron.2006.12.024. [DOI] [PubMed] [Google Scholar]
- Leon M. Neuroethology of olfactory preference development. J Neurobiol. 1992;23:1557–73. doi: 10.1002/neu.480231012. [DOI] [PubMed] [Google Scholar]
- Limb CJ, Ryugo DK. Development of primary axosomatic endings in the anteroventral cochlear nucleus of mice. J Assoc Res Otolaryngol. 2000;1:103–19. doi: 10.1007/s101620010032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin DM, Wang F, Lowe G, Gold GH, Axel R, Ngai J, Brunet L. Formation of precise connections in the olfactory bulb occurs in the absence of odorant-evoked neuronal activity. Neuron. 2000;26:69–80. doi: 10.1016/s0896-6273(00)81139-3. [DOI] [PubMed] [Google Scholar]
- Long JE, Garel S, Depew MJ, Tobet S, Rubenstein JL. DLX5 regulates development of peripheral and central components of the olfactory system. J Neurosci. 2003;23:568–78. doi: 10.1523/JNEUROSCI.23-02-00568.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mair RG, Gesteland RC. Response properties of mitral cells in the olfactory bulb of the neonatal rat. Neuroscience. 1982;7:3117–25. doi: 10.1016/0306-4522(82)90234-2. [DOI] [PubMed] [Google Scholar]
- Marcotti W, Johnson SL, Holley MC, Kros CJ. Developmental changes in the expression of potassium currents of embryonic, neonatal and mature mouse inner hair cells. J Physiol. 2003a;548:383–400. doi: 10.1113/jphysiol.2002.034801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcotti W, Johnson SL, Rusch A, Kros CJ. Sodium and calcium currents shape action potentials in immature mouse inner hair cells. J Physiol. 2003b;552:743–61. doi: 10.1113/jphysiol.2003.043612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin MR, Rickets C. Histogenesis of the cochlear nucleus of the mouse. J Comp Neurol. 1981;197:169–84. doi: 10.1002/cne.901970113. [DOI] [PubMed] [Google Scholar]
- Meister M, Wong RO, Baylor DA, Shatz CJ. Synchronous bursts of action potentials in ganglion cells of the developing mammalian retina. Science. 1991;252:939–43. doi: 10.1126/science.2035024. [DOI] [PubMed] [Google Scholar]
- Mikaelian D, Ruben R. Development of hearing in the normal CBA-J Mouse. Acta Otolaryngol. 1965;59:451–461. [Google Scholar]
- Miyamichi K, Serizawa S, Kimura HM, Sakano H. Continuous and overlapping expression domains of odorant receptor genes in the olfactory epithelium determine the dorsal/ventral positioning of glomeruli in the olfactory bulb. J Neurosci. 2005;25:3586–92. doi: 10.1523/JNEUROSCI.0324-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mo ZL, Davis RL. Endogenous firing patterns of murine spiral ganglion neurons. J Neurophysiol. 1997;77:1294–305. doi: 10.1152/jn.1997.77.3.1294. [DOI] [PubMed] [Google Scholar]
- Mombaerts P, Wang F, Dulac C, Chao SK, Nemes A, Mendelsohn M, Edmondson J, Axel R. Visualizing an olfactory sensory map. Cell. 1996;87:675–86. doi: 10.1016/s0092-8674(00)81387-2. [DOI] [PubMed] [Google Scholar]
- Mooney R, Penn AA, Gallego R, Shatz CJ. Thalamic Relay of Spontaneous Retinal Activity Prior to Vision. Neuron. 1996;17:863–874. doi: 10.1016/s0896-6273(00)80218-4. [DOI] [PubMed] [Google Scholar]
- Moriizumi T, Sakashita H, Furukawa M, Kawano J, Okoyama S, Kitao Y, Kudo M. Electron microscopic study of synaptogenesis and myelination of the olfactory centers in developing rats. Exp Brain Res. 1995;103:385–92. doi: 10.1007/BF00241497. [DOI] [PubMed] [Google Scholar]
- Moshiri A, Gonzalez E, Tagawa K, Maeda H, Wang M, Frishman LJ, Wang SW. Near complete loss of retinal ganglion cells in the math5/brn3b double knockout elicits severe reductions of other cell types during retinal development. Developmental Biology. 2008;316:214–227. doi: 10.1016/j.ydbio.2008.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neher E, Marty A. Discrete changes of cell membrane capacitance observed under conditions of enhanced secretion in bovine adrenal chromaffin cells. Proc Natl Acad Sci U S A. 1982;79:6712–6. doi: 10.1073/pnas.79.21.6712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nemzou RMN, Bulankina AV, Khimich D, Giese A, Moser T. Synaptic organization in cochlear inner hair cells deficient for the CaV1.3 ([alpha]1D) subunit of L-type Ca2+ channels. Neuroscience. 2006;141:1849–1860. doi: 10.1016/j.neuroscience.2006.05.057. [DOI] [PubMed] [Google Scholar]
- Ollo C, Schwartz IR. The superior olivary complex in C57BL/6 mice. Am J Anat. 1979;155:349–73. doi: 10.1002/aja.1001550306. [DOI] [PubMed] [Google Scholar]
- Olney JW. An electron microscopic study of synapse formation, receptor outer segment development, and other aspects of developing mouse retina. Invest Ophthalmol. 1968;7:250–68. [PubMed] [Google Scholar]
- Pierce ET. Histogenesis of the dorsal and ventral cochlear nuclei in the mouse. An autoradiographic study. J Comp Neurol. 1967;131:27–54. doi: 10.1002/cne.901310104. [DOI] [PubMed] [Google Scholar]
- Pierce ET. Time of origin of neurons in the brain stem of the mouse. Prog Brain Res. 1973;40:53–65. doi: 10.1016/S0079-6123(08)60679-2. [DOI] [PubMed] [Google Scholar]
- Polleux F, Dehay C, Kennedy H. The timetable of laminar neurogenesis contributes to the specification of cortical areas in mouse isocortex. J Comp Neurol. 1997;385:95–116. [PubMed] [Google Scholar]
- Polley EH, Zimmerman RP, Fortney RL. Neurogenesis and maturation of cell morphology in the development of the mammalian retina. In: Finlay BL, Sengelaub DR, editors. Development of the Vertebrate Retina. Plenum Press; New York: 1989. pp. 3–42. [Google Scholar]
- Poole T, editor. The UFAW Handbook on the Care and Management of Laboratory Animals. Churchill Livingstone, Inc.; New York: 1987. [Google Scholar]
- Price JL. An autoradiographic study of complementary laminar patterns of termination of afferent fibers to the olfactory cortex. J Comp Neurol. 1973;150:87–108. doi: 10.1002/cne.901500105. [DOI] [PubMed] [Google Scholar]
- Pujol R, Lavigne-Rebillard M, Lenoir M. Development of Sensory and Neural Structures in the Mammalian Cochlea. In: Rubel EW, Popper AN, Fay RR, editors. Development of the auditory system. Springer handbook of auditory research. Vol. 9. Springer; New York: 1998. pp. 146–192. [Google Scholar]
- Puyal J, Sage C, Demêmes D, Dechesne CJ. Distribution of [alpha]-amino-3-hydroxy-5-methyl-4 isoazolepropionic acid and N-methyl--aspartate receptor subunits in the vestibular and spiral ganglia of the mouse during early development. Developmental Brain Research. 2002;139:51–57. doi: 10.1016/s0165-3806(02)00535-7. [DOI] [PubMed] [Google Scholar]
- Rapaport DH, Wong LL, Wood ED, Yasumura D, LaVail MM. Timing and topography of cell genesis in the rat retina. J Comp Neurol. 2004;474:304–24. doi: 10.1002/cne.20134. [DOI] [PubMed] [Google Scholar]
- Ressler KJ, Sullivan SL, Buck LB. A zonal organization of odorant receptor gene expression in the olfactory epithelium. Cell. 1993;73:597–609. doi: 10.1016/0092-8674(93)90145-g. [DOI] [PubMed] [Google Scholar]
- Rich KA, Zhan Y, Blanks JC. Migration and synaptogenesis of cone photoreceptors in the developing mouse retina. J Comp Neurol. 1997;388:47–63. [PubMed] [Google Scholar]
- Rodriguez-Contreras A, John SSvH, Ron LPH, Heiko LJGGB. Dynamic development of the calyx of Held synapse. Proceedings of the National Academy of Sciences. 2008;105:5603–5608. doi: 10.1073/pnas.0801395105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roux I, Safieddine S, Nouvian R, Grati M, Simmler MC, Bahloul A, Perfettini I, Le Gall M, Rostaing P, Hamard G, Triller A, Avan P, Moser T, Petit C. Otoferlin, defective in a human deafness form, is essential for exocytosis at the auditory ribbon synapse. Cell. 2006;127:277–89. doi: 10.1016/j.cell.2006.08.040. [DOI] [PubMed] [Google Scholar]
- Rubel EW, Popper AN, Fay RR. Development of the auditory system. Vol. 9. Springer; New York: 1998. [Google Scholar]
- Ruben RJ. Development of the inner ear of the mouse: a radioautographic study of terminal mitoses. Acta Otolaryngol Suppl. 1967;220:1–44. [PubMed] [Google Scholar]
- Schwob JE, Haberly LB, Price JL. The development of physiological responses of the piriform cortex in rats to stimulation of the lateral olfactory tract. J Comp Neurol. 1984;223:223–37. doi: 10.1002/cne.902230206. [DOI] [PubMed] [Google Scholar]
- Schwob JE, Price JL. The development of axonal connections in the central olfactory system of rats. J Comp Neurol. 1984;223:177–202. doi: 10.1002/cne.902230204. [DOI] [PubMed] [Google Scholar]
- Seal RP, Akil O, Yi E, Weber CM, Grant L, Yoo J, Clause A, Kandler K, Noebels JL, Glowatzki E, Lustig LR, Edwards RH. Sensorineural deafness and seizures in mice lacking vesicular glutamate transporter 3. Neuron. 2008;57:263–75. doi: 10.1016/j.neuron.2007.11.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma RK, O’Leary TE, Fields CM, Johnson DA. Development of the outer retina in the mouse. Developmental Brain Research. 2003;145:93–105. doi: 10.1016/s0165-3806(03)00217-7. [DOI] [PubMed] [Google Scholar]
- Sherry DM, Wang MM, Bates J, Frishman LJ. Expression of vesicular glutamate transporter 1 in the mouse retina reveals temporal ordering in development of rod vs. cone and ON vs. OFF circuits. The Journal of Comparative Neurology. 2003;465:480–498. doi: 10.1002/cne.10838. [DOI] [PubMed] [Google Scholar]
- Smotherman WP, Robinson SR, La Vallee PA, Hennessy MB. Influences of the early olfactory environment on the survival, behavior and pituitary-adrenal activity of caesarean delivered preterm rat pups. Dev Psychobiol. 1987;20:415–23. doi: 10.1002/dev.420200406. [DOI] [PubMed] [Google Scholar]
- Smotherman WP, Robinson SR. Rat fetuses respond to chemical stimuli in gas phase. Physiol Behav. 1990;47:863–8. doi: 10.1016/0031-9384(90)90010-2. [DOI] [PubMed] [Google Scholar]
- Sobkowicz HM, Slapnick SM, August BK. Reciprocal synapses between inner hair cell spines and afferent dendrites in the organ of corti of the mouse. Synapse. 2003;50:53–66. doi: 10.1002/syn.10241. [DOI] [PubMed] [Google Scholar]
- Spoendlin H. The innervation of the organ of Corti. J Laryngol Otol. 1967;81:717–38. doi: 10.1017/s0022215100067669. [DOI] [PubMed] [Google Scholar]
- Sullivan RM. Developing a sense of safety: the neurobiology of neonatal attachment. Ann N Y Acad Sci. 2003;1008:122–31. doi: 10.1196/annals.1301.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Syed MM, Lee S, Zheng J, Zhou ZJ. Stage-dependent dynamics and modulation of spontaneous waves in the developing rabbit retina. J Physiol. 2004;560:533–49. doi: 10.1113/jphysiol.2004.066597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tessier-Lavigne M, Goodman CS. The molecular biology of axon guidance. Science. 1996;274:1123–33. doi: 10.1126/science.274.5290.1123. [DOI] [PubMed] [Google Scholar]
- Thurlow GA, Cooper RM. Metabolic changes in the superior colliculus after retinal receptor loss--neurotrophic interactions in the inactive visual system. Exp Neurol. 1988;100:563–77. doi: 10.1016/0014-4886(88)90040-4. [DOI] [PubMed] [Google Scholar]
- Tian N, Copenhagen DR. Visual stimulation is required for refinement of ON and OFF pathways in postnatal retina. Neuron. 2003;39:85–96. doi: 10.1016/s0896-6273(03)00389-1. [DOI] [PubMed] [Google Scholar]
- Torborg CL, Feller MB. Spontaneous patterned retinal activity and the refinement of retinal projections. Progress in Neurobiology. 2005;76:213–235. doi: 10.1016/j.pneurobio.2005.09.002. [DOI] [PubMed] [Google Scholar]
- Tritsch NX, Yi E, Gale JE, Glowatzki E, Bergles DE. The origin of spontaneous activity in the developing auditory system. Nature. 2007;450:50–55. doi: 10.1038/nature06233. [DOI] [PubMed] [Google Scholar]
- Turner DL, Cepko CL. A common progenitor for neurons and glia persists in rat retina late in development. Nature. 1987;328:131–6. doi: 10.1038/328131a0. [DOI] [PubMed] [Google Scholar]
- Varoqueaux F, Sigler A, Rhee JS, Brose N, Enk C, Reim K, Rosenmund C. Total arrest of spontaneous and evoked synaptic transmission but normal synaptogenesis in the absence of Munc13-mediated vesicle priming. Proc Natl Acad Sci U S A. 2002;99:9037–42. doi: 10.1073/pnas.122623799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verhage M, Maia AS, Plomp JJ, Brussaard AB, Heeroma JH, Vermeer H, Toonen RF, Hammer RE, van den Berg TK, Missler M, Geuze HJ, Sudhof TC. Synaptic assembly of the brain in the absence of neurotransmitter secretion. Science. 2000;287:864–9. doi: 10.1126/science.287.5454.864. [DOI] [PubMed] [Google Scholar]
- Vogalis F, Hegg CC, Lucero MT. Electrical coupling in sustentacular cells of the mouse olfactory epithelium. J Neurophysiol. 2005;94:1001–12. doi: 10.1152/jn.01299.2004. [DOI] [PubMed] [Google Scholar]
- Walz A, Omura M, Mombaerts P. Development and topography of the lateral olfactory tract in the mouse: imaging by genetically encoded and injected fluorescent markers. J Neurobiol. 2006;66:835–46. doi: 10.1002/neu.20266. [DOI] [PubMed] [Google Scholar]
- Wang CT, Blankenship AG, Anishchenko A, Elstrott J, Fikhman M, Nakanishi S, Feller MB. GABA(A) receptor-mediated signaling alters the structure of spontaneous activity in the developing retina. J Neurosci. 2007;27:9130–40. doi: 10.1523/JNEUROSCI.1293-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang F, Nemes A, Mendelsohn M, Axel R. Odorant receptors govern the formation of a precise topographic map. Cell. 1998;93:47–60. doi: 10.1016/s0092-8674(00)81145-9. [DOI] [PubMed] [Google Scholar]
- Wang VY, Rose MF, Zoghbi HY. Math1 Expression Redefines the Rhombic Lip Derivatives and Reveals Novel Lineages within the Brainstem and Cerebellum. Neuron. 2005;48:31–43. doi: 10.1016/j.neuron.2005.08.024. [DOI] [PubMed] [Google Scholar]
- Wetts R, Fraser SE. Multipotent precursors can give rise to all major cell types of the frog retina. Science. 1988;239:1142–5. doi: 10.1126/science.2449732. [DOI] [PubMed] [Google Scholar]
- Wong RO, Meister M, Shatz CJ. Transient period of correlated bursting activity during development of the mammalian retina. Neuron. 1993;11:923–38. doi: 10.1016/0896-6273(93)90122-8. [DOI] [PubMed] [Google Scholar]
- Wong RO, Chernjavsky A, Smith SJ, Shatz CJ. Early functional neural networks in the developing retina. Nature. 1995;374:716–8. doi: 10.1038/374716a0. [DOI] [PubMed] [Google Scholar]
- Wong WT, Myhr KL, Miller ED, Wong RO. Developmental changes in the neurotransmitter regulation of correlated spontaneous retinal activity. J Neurosci. 2000;20:351–60. doi: 10.1523/JNEUROSCI.20-01-00351.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods C, Montcouquiol M, Kelley MW. Math1 regulates development of the sensory epithelium in the mammalian cochlea. Nat Neurosci. 2004;7:1310–8. doi: 10.1038/nn1349. [DOI] [PubMed] [Google Scholar]
- Wu SH, Oertel D. Maturation of synapses and electrical properties of cells in the cochlear nuclei. Hear Res. 1987;30:99–110. doi: 10.1016/0378-5955(87)90187-0. [DOI] [PubMed] [Google Scholar]
- Yee KT, Simon HH, Tessier-Lavigne M, O’Leary DDM. Extension of Long Leading Processes and Neuronal Migration in the Mammalian Brain Directed by the Chemoattractant Netrin-1. Neuron. 1999;24:607–622. doi: 10.1016/s0896-6273(00)81116-2. [DOI] [PubMed] [Google Scholar]
- Young RW. Cell differentiation in the retina of the mouse. Anat Rec. 1985;212:199–205. doi: 10.1002/ar.1092120215. [DOI] [PubMed] [Google Scholar]
- Yu CR, Power J, Barnea G, O’Donnell S, Brown HE, Osborne J, Axel R, Gogos JA. Spontaneous neural activity is required for the establishment and maintenance of the olfactory sensory map. Neuron. 2004;42:553–66. doi: 10.1016/s0896-6273(04)00224-7. [DOI] [PubMed] [Google Scholar]
- Zheng J, Lee S, Zhou ZJ. A transient network of intrinsically bursting starburst cells underlies the generation of retinal waves. Nat Neurosci. 2006;9:363–71. doi: 10.1038/nn1644. [DOI] [PubMed] [Google Scholar]
- Zou DJ, Feinstein P, Rivers AL, Mathews GA, Kim A, Greer CA, Mombaerts P, Firestein S. Postnatal refinement of peripheral olfactory projections. Science. 2004;304:1976–9. doi: 10.1126/science.1093468. [DOI] [PubMed] [Google Scholar]