

Summary
Specialization of the cell-division process is a common feature of developing embryos, but most studies on vertebrate cell division have focused on cells dividing in culture. Here, we used in vivo four-dimensional confocal microscopy to explore the role of Cdc42 in governing cell division in the developing neural epithelium of Xenopus laevis. We find that Cdc42 is crucial for stable positioning of the metaphase spindle in these cells, but was not required for spindle positioning in epidermal epithelial cells. We also find that divisions in the Xenopus neural plate are planar oriented, and that rotations of mitotic spindles are essential for establishing this orientation. When Cdc42 is disrupted, spindles over-rotate and the final orientation of divisions is changed. Finally, the planar orientation of cell divisions in this tissue seems to be independent of planar cell polarity (PCP) signaling and does not require normal neural morphogenesis. Our data provide new insights into the coordination of cell division and morphogenesis in epithelial cell sheets and reveal novel, cell-type-specific roles for Cdc42 in spindle positioning and spindle orientation.
Keywords: Cdc42, Metaphase plate, Neural tube closure, Oriented cell division, Spindle
Introduction
Developmentally regulated cell division is a central facet of embryogenesis, and how cells establish orientation and asymmetries during cell division is a fundamental issue at the interface of cell and developmental biology. Precisely oriented divisions are important for controlling determination of the embryonic axes in Caenorhabditis elegans (Gonczy and Rose, 2005; Gotta et al., 2001), for axis elongation and epithelial-tube elongation in vertebrates (Concha and Adams, 1998; Fischer et al., 2006; Gong et al., 2004; Saburi et al., 2008) and for the diversification of cell types in the embryonic nervous systems of Drosophila and Xenopus (Bellaiche et al., 2001a; Bellaiche et al., 2001b; Chalmers et al., 2003; David et al., 2005).
Oriented cell divisions have been thoroughly described in more mature vertebrate nervous systems as well. In the closed neural tube, divisions that occur parallel to the plane of the neural epithelium are generally proliferative, whereas divisions that are perpendicular to the apical membrane tend to be neurogenic (Gotz and Huttner, 2005; Haydar et al., 2003; Wilcock et al., 2007). Moreover, proliferative divisions occurring within the plane of the neural epithelium can display stereotypical orientations with respect to the anteroposterior axis. Earlier in development, the hollow neural tube forms by the intricate folding of an initially flat neural plate during a process termed neural tube closure (Wallingford, 2005). During neural tube closure in chick embryos, the axis of cell division is preferentially parallel to the long axis of the embryo (Sausedo et al., 1997), whereas, in zebrafish, divisions are initially parallel to the long axis and shift to a perpendicular orientation as neurulation proceeds (Concha and Adams, 1998; Geldmacher-Voss et al., 2003; Gong et al., 2004). The Xenopus embryo represents a special situation, however. The neural plate of Xenopus has two cell layers, a deep layer giving rise to the primary neurons and a superficial layer that only differentiates much later and continues to proliferate dramatically during neural tube closure (Chalmers et al., 2002; Hartenstein, 1989).
Many oriented cell divisions have a few controlling factors in common. In particular, the planar cell polarity (PCP) pathway has been found to be a major player. PCP signaling was first shown to regulate spindle rotations that orient cell divisions in Drosophila neuroblasts, facilitating the asymmetric distribution of determination factors (Bellaiche et al., 2001a; Bellaiche et al., 2001b; David et al., 2005). PCP signaling in the zebrafish governs orientation of cell divisions along the anteroposterior axis that facilitate elongation of the embryo (Gong et al., 2004). Most recently, PCP signaling has been found to control oriented cell division in the developing mammalian kidney (Saburi et al., 2008). It should be noted, however, that this role for PCP signaling in controlling oriented cell division is not universal. For example, extension of the germ band in Drosophila also relies on oriented cell divisions, but this orientation is independent of PCP signaling (da Silva and Vincent, 2007).
Another common player in developmental regulation of oriented cell divisions is the small GTPase Cdc42. In C. elegans, Cdc42 controls spindle orientation during the first cell cycle and thereby is involved in defining the anteroposterior axis (Gonczy and Rose, 2005; Gotta et al., 2001). A clear role for Cdc42 in controlling oriented cell division in vertebrate somatic cells has not been reported as yet, but Cdc42 is essential for proper localization and organization of the meiotic spindle in mouse oocytes (Na and Zernicka-Goetz, 2006). Moreover, Cdc42 is activated asymmetrically on meiotic spindles in Xenopus oocytes, where it links spindle position to the position of the cytokinetic ring (Ma et al., 2006). These functions might reflect the role Cdc42 plays in bud-site selection in yeast (Kozminski et al., 2003).
The involvement of Cdc42 in oriented cell division in developing embryos is particularly intriguing in light of recent findings that suggest that Cdc42 plays cell-type-specific roles in many fundamental aspects of cell division. In some cell lines, Cdc42 has been reported to control microtubule attachment to kinetochores (Yasuda et al., 2004), whereas, in others, it governs assembly of the cytokinetic septin cytoskeleton, as it does in yeast (Caviston et al., 2003; Garcia et al., 2006; Joberty et al., 2001). Moreover, biosensors in live cells have revealed differing patterns of Cdc42 activity during division of different cultured cell types (Garcia et al., 2006; Yoshizaki et al., 2003). These differences might reflect the fact that even fundamental aspects of cell division are under cell-type-specific developmental control. For example, the mechanisms of central-spindle assembly differs dramatically between early neural epithelial cells and epidermal epithelia cells in Xenopus embryos (Kieserman et al., 2008). These differences, including changes in the midzone microtubule density and the extent of anaphase B, stem from cell-type-specific variation in the levels of PRC1 protein (Kieserman et al., 2008). It is interesting, then, that PRC1 can regulate Cdc42 activity during cell division (Ban et al., 2004).
To enhance our understanding of the roles of Cdc42 in vertebrate cell division, we used in vivo four-dimensional (4D) imaging to study cell divisions in the developing Xenopus embryo. Using mosaic embryos, we show here that manipulation of Cdc42 in neural epithelial cells during neural tube closure results in a failure of cells to stabilize the mitotic spindle in the cell center during metaphase. By contrast, manipulation of Cdc42 in epidermal epithelial cells had no effect on spindle positioning. Moreover, we have found that cell divisions in the early neural ectoderm of Xenopus embryos are oriented mediolaterally within the plane of the epithelium. This orientation was established during metaphase by variable rotations of mitotic spindles that were initially assembled in random orientations. The orientation of these cell divisions was not controlled by the PCP pathway, but instead was regulated by Cdc42. When Cdc42 function is altered, spindles undergo excessive rotations, and the final positioning of spindles is altered. These data demonstrate a key role for Cdc42 in spindle position and orientation in neural epithelial cells during neural tube closure. Because we find that Cdc42 plays no such role in epidermal cells, our findings highlight what may be a broad role for Cdc42 in governing the specializations of cell-division mechanisms that are essential in developing embryos.
Results
In vivo imaging of cell division and mosaic analysis of gene function during Xenopus neural tube closure
We have developed a method to examine cell divisions using three-dimensional (3D) time-lapse confocal microscopy in intact, developing vertebrate embryos during neural tube closure. mRNAs encoding fluorescent fusion proteins were injected into the early blastomeres of Xenopus embryos, typically at the four-cell stage (Fig. 1A, 1). In order to image cell behaviors during neural tube closure, neurulating embryos were immobilized upside down in embryo-sized wells that were made in agarose in a culture dish. The culture dish had a cover glass bottom and was placed on an inverted confocal microscope for imaging (Fig. 1A, 2). Horizontal confocal slices were taken of the neural plate to assemble a single stack every 30-60 seconds (Fig. 1A, 4). Stacks were then projected into an x-y projection (Fig. 1A, 5), and then exported and analyzed (Fig. 1A, 6). This technique allows for clear analysis of not only tissue-level morphogenetic events, such as the movement of the neural folds, but also subcellular behaviors (supplementary material Movie 1; Fig. 1B; Fig. 2A,A′). We exploited this method to further characterize the extensive, developmentally regulated cell divisions that occur in the closing neural tube of Xenopus (Hartenstein, 1989; Kieserman et al., 2008; Saka and Smith, 2001; Saka and Smith, 2004).
Fig. 1.

Analysis of cell divisions during neural tube closure. (A) Method to analyze cell divisions during neural tube closure in Xenopus laevis. Eggs are collected and undergo in vitro fertilization as described in the Materials and Methods. Solutions of mRNA, previously made by an in vitro transcription kit (Sigma), are created to label appropriate portions of cells (i.e. chromosomes via H2B-GFP or membranes via mem-RFP). Mixes of mRNA are put in small glass needles and are injected directly into the embryo at the four-cell stage by use of a picospritzer (1). For analysis of the early neural plate, the two dorsal cells are injected and, for analysis of the late tail epidermis, all four cells are injected. Embryos are then grown to an appropriate stage: stage 13 for early neural plate and stage 30 for late tail epidermis (2). Embryos are then placed on a microscope and imaged. Images of the early neural plate are obtained from horizontal confocal slices of the dorsal surface of the spinal-chord region prior to neural tube closure (3, see red box). Stacks are obtained every 30 seconds or 1 minute for 150 iterations (4). Movies are then projected into an x confocal projection in the Pascal program (5). Confocal projections are then exported to tiff files and analyzed by use of ImagePro Plus (6). (B) Use of this method allows for easy analysis of morphogenesis of the neural plate. Movies are started at early neural plate stage (t=0 minutes). As time progresses, neural folds come into view (t=80 minutes). These neural folds continue to move laterally until the neural plate is no longer well seen and the future epidermis of the embryo is easily seen (t=90 minutes and t=110 minutes) (supplementary material Movie 1). Scale bar: 50 μm. (C) Method to assess division-polarity effectors. Embryos are injected so that they have different clones for analysis of polarity effectors. Embryos are injected on their entire dorsal side with a histone-2B–GFP marker at the four-cell stage (1). Embryos are grown up one stage, to the eight-cell stage, and injected with a membrane-RFP marker and an mRNA of interest (2). Embryos are grown to the early neurula stage and cells injected with the mRNA of interest can be distinguished by the expression of GFP and RFP, whereas control cells only express the GFP marker (3). DN, dominant negative. Scale bar: 100 μm.
Fig. 2.

Mitotic chromosomes undergo drastic movements in cells expressing dominant negatives to Cdc42. (A) Still frames from a movie showing a control cell. The chromosomes can be seen to condense and remain at that location throughout metaphase. Time is shown at the top (minutes). (A′) Cartoons of the cell of interest from above. Cell outlines are shown in black and nuclei are shown in green. (B) Still frames from a movie showing a cell expressing Cdc42-F37A dominant negative during metaphase. The chromosomes can be seen to move quickly from the upper-left-hand side of the cell to the lower left, and back again (supplementary material Movie 3). Time is shown at the top (minutes). (B′) Cartoons of the cell of interest from above. Cell outlines are shown in black and nuclei are shown in green. Chromosomes move dramatically through the cell before aligning at the approximate middle of the cell and anaphase proceeds (supplementary material Movie 3). Scale bars: 10 μm.
To test the roles of specific proteins in cell division during neural tube closure, we modified this in vivo imaging approach to generate clones of manipulated cells within an otherwise normal embryo. For these experiments, embryos were injected into both dorsal blastomeres at the four-cell stage with histone-2B–GFP mRNA to allow for monitoring of division orientation throughout the neural plate (Fig. 1C, 1). Embryos were then grown to the eight-cell stage and injected again, this time into only one of the dorsal, animal blastomeres (Fig. 1C, 2). This blastomere was injected with a combination of mRNA encoding our experimental construct and mRNA encoding membrane-RFP, to distinguish the experimental cells from normal cells at neurula stages (Fig. 1C, 2 and 3). Wild-type clones and experimental clones can then be compared directly within a single embryo.
Cdc42 is required to stably position metaphase spindles in neural-plate cells during neural tube closure
Cdc42 has been shown to control spindle position and orientation during cell divisions in early C. elegans embryos, and in mouse and Xenopus oocytes (Bement et al., 2005; Gotta et al., 2001; Ma et al., 2006; Na and Zernicka-Goetz, 2006). We therefore used mosaics (see above) to examine what role Cdc42 might play in controlling spindle dynamics in the Xenopus neural plate. During cell divisions in neural epithelial cells, the metaphase plate normally remains positioned stably in the cell center (Fig. 2A,A′) (Kieserman et al., 2008). In these mosaics, cells expressing the dominant-negative Cdc42-N17 displayed chaotic movements of the condensed chromosomes during metaphase (supplementary material Movie 2). In control cells within these mosaics, metaphase plates remained in place, centered in the middle of the cell, as was observed in un-manipulated embryos (Fig. 2A,A′).
Cdc42-N17 disrupts Cdc42 function by blocking the ability of the GTPase to exchange GDP for GTP. Therefore, by binding and sequestering guanine-nucleotide exchange factors, Cdc42-N17 might disrupt the function of other GTPases that share such exchange factors (Karnoub et al., 2004; Zhang et al., 1995). To confirm the role of Cdc42 in spindle stability in the Xenopus neural plate, we repeated our experiment with another Cdc42 dominant-negative, Cdc42-F37A (Lamarche et al., 1996; Richman et al., 2002; Zhang et al., 1995). This construct is a more-specific disruptor of Cdc42 function, blocking the interaction between Cdc42 and its downstream effector proteins (Lamarche et al., 1996). In mosaics, cells expressing Cdc42-F37A displayed the aberrant movement of condensed chromosomes within the cell during metaphase. The violent movements in Cdc42-F37A-expressing cells can be observed in Fig. 2A,A′ and supplementary material Movie 3. Immediately after forming, the metaphase plate moves quickly from the upper-left-hand portion of the cell to the lower-left-hand portion. The plate then moves back, past the center of the cell, to the cortex in the upper left, and finally returns to the approximate center of the cell as anaphase begins (Fig. 2A,A′; supplementary material Movie 3)
We next sought to quantify the excessive spindle movement following manipulation of Cdc42. During neural tube closure, the entire neural plate is deforming dramatically, so it was necessary to develop a measurement scheme that isolates intracellular movement of the spindle from the overall movement of the cells in the neural plate. We therefore quantified the intracellular distance of metaphase-plate movement as a function of the length of the long axis of that cell, per unit of time (Fig. 3A). We performed these measurements over the 14 minutes preceding anaphase onset.
Fig. 3.

Cdc42 is required to stabilize metaphase plates in the early neural plate but not in the tail epidermis. (A) Diagram and equation of how chromosome stability was assessed. Cell lengths were measured each minute for the 14 minutes preceding anaphase onset in the approximate orientation of division (d1). The distance of the chromosomes from the most-posterior edge of the cell to the middle of the chromosome mass was measured for the same 14 minutes, again in the approximate angle of division (d2). The spindle instability index (SII) was assessed using the equation shown. At each time point, d2 was divided by d1. This value was then subtracted from the subsequent time point and this total was divided by the elapsed time. (B) Graph showing the mean SII value for control cells (white bar, n=4 embryos, 26 cells), cells expressing Cdc42-N17 (gray bar, n=6 embryos, 26 cells) and cells expressing Cdc42-F37A (black bar, n=2 embryos, 26 cells). Error bars show s.e.m. (C) Graph showing instantaneous SII for each time point of all of the 26 control neural cells. The maximum instantaneous SII seen for control cells is 20% and is represented by the red dashed line. (D) Graph showing instantaneous SII for each time point of all of the 26 Cdc42-N17-expressing neural cells. The maximum instantaneous SII seen for control neural cells is 20% and is represented by the red dashed line. (E) Graph showing instantaneous SII for each time point of all of the 26 Cdc42-F37A-expressing neural cells. The maximum instantaneous SII seen for control neural cells is 20% and is represented by the red dashed line. (F) Graph showing the mean SII value for control cells (white bar, n=3 embryos, 26 cells) and cells expressing Cdc42-F37A (black bar, n=2 embryos, 26 cells). Error bars show s.e.m. (G) Graph showing instantaneous SII for each time point of all of the 26 control epidermal cells. The maximum instantaneous SII seen for control neural cells is 20% and is represented by the red dashed line. (H) Graph showing instantaneous SII for each time point of all of the 26 Cdc42-N17-expressing epidermal cells. The maximum instantaneous SII seen for control neural cells is 20% and is represented by the red dashed line.
In control cells, metaphase plates moved on average only 5% of the cell length per minute (Fig. 3B), and metaphase plates almost never moved more than 20% of cell length in a given minute of time, with the maximum movement being 26% (Fig. 3C). By contrast, cells in embryos expressing either of the dominant-negative forms of Cdc42 frequently moved greater than 20% of cell length per minute, and the maximum movement for both manipulations was over 46% (Fig. 3D,E). Likewise, these nuclei on average moved greater than 8% of the cell length per minute (Fig. 3B). This difference in the average distance that was moved between control and experimental groups was extremely significant (P<0.001, Kruskal-Wallis multiple-comparisons test).
Cdc42 does not control spindle positioning in epithelial cells of the developing Xenopus epidermis
Given that core cytokinesis mechanisms differ surprisingly between neural epithelial cells and epidermal epithelial cells examined in vivo (Kieserman et al., 2008), we next investigated whether epidermal cell division was affected by disruption of Cdc42. We made in vivo 4D movies of Xenopus tail epidermis. In control epidermal cells, chromosomes separated to the geometric center of the new daughter cells and cytokinesis initiated at about 4 minutes after anaphase onset (supplementary material Movie 4) (Kieserman et al., 2008). In contrast to the effect on neural cells, metaphase, anaphase and cytokinesis in epidermal cells expressing Cdc42-N17 were indistinguishable from that in control cells (supplementary material Movie 5).
We quantified the intracellular movement of metaphase plates in epidermal cells and found that they moved on average only 3.9% of the cell length per minute, and movements of 20% or more of the cell length were never observed (Fig. 3F-H). Thus, intracellular movement of metaphase plates in non-manipulated neural cells was significantly greater than that of non-manipulated epidermal cells (P<0.001). The jostling of metaphase plates of neural cells might reflect the fact that the neural plate is actively moving, or it might be related to the spindle rotations that establish the planar orientation of these cells divisions (see below). Regardless, expression of Cdc42-N17 had no effect whatsoever on the stable position of metaphase plates in dividing epidermal cells in vivo (Fig. 3F; compare Fig. 3H with 3G). These data demonstrate that Cdc42 plays a cell-type-specific role in governing cell division in vertebrate embryonic epithelia, revealing a neural-specific role for Cdc42 in the stable positioning of the metaphase spindle.
Planar orientation of cell divisions in the closing neural tube of Xenopus
In C. elegans, Cdc42 governs not only spindle positioning but also the orientation of cell divisions (Gotta et al., 2001). Because the orientation of cell divisions is developmentally regulated in the closing neural tube of other vertebrates, we investigated whether the same might be true for Xenopus and whether Cdc42 has any role in controlling the process. Previous results from fish and chick suggest that the planar cell divisions during neural tube closure would be aligned along the anteroposterior or mediolateral axes (see Introduction, above). We quantified the orientation of neural-plate cell divisions by plotting the position of the mitotic spindles on a rose diagram, and this analysis clearly demonstrates that the vast majority of cell divisions in the neural plate during neural tube closure are aligned roughly perpendicular to the anteroposterior axis.
However, the division angles of Xenopus neural-plate cells varied more widely than those observed in zebrafish (Concha and Adams, 1998); many cells in the Xenopus superficial neural plate divided at an angle that was oblique with respect to the mediolateral axis of the embryo (supplementary material Movie 6; Fig. 4A-B). Indeed, in our movies, more cells were observed dividing obliquely with respect to the mediolateral axis than were observed dividing directly parallel to the mediolateral axis (Fig. 4C). Curiously, the `diagonal' or `slanted' orientation of a cell-division angle was irrespective of the mediolateral position of a cell in the neural plate. Fig. 4A provides an example: within a small area on the right side of a neural plate (Fig. 4A,A′) some cells were observed to divide with their anterior-most daughter cells facing towards the midline (Fig. 4A′, pink arrows), whereas other cells in the same region divided with their anterior-most daughter cells away from the midline (Fig. 4A′, blue arrows). In many cases, we observed adjacent cells in the neural plate dividing simultaneously, one oriented obliquely towards the midline and the other oriented obliquely away from the midline (supplementary material Movie 7; Fig. 4B).
Fig. 4.

Cell divisions in the early neural plate are polarized at an oblique angle with respect to the midline. (A) Still frames from a section of the movie in Fig. 1. Anterior is to the top and the midline is to the left. A few cells undergoing division are highlighted (transparent squares). Cells with their anterior daughter cell oriented away from the midline are denoted by a blue arrow (t=0 minutes and 24 minutes) (supplementary material Movie 6). Cells that divide with their anterior daughter cell towards the midline are denoted by a pink arrow (t=10 minutes and 39 minutes). Scale bar: 50 μm. (A′) Blow up of cells highlighted in A. The angle of division, with respect to the midline, is shown in white text. Scale bar: 10 μm. (B) Still frames from a movie of cells dividing in the early neural plate. Cells that are adjacent in the anteroposterior area divide with opposite oblique polarities. One cell is seen to divide with its anterior daughter cell facing away from the midline (blue arrow). The posterior neighbor of this cell instead divides with its anterior daughter cell facing towards the midline (pink arrow). The angles of these divisions are noted and correspond to the diagram in Fig. 2D (supplementary material Movie 7). Scale bar: 10 μm. (C) Rose diagram of division orientation of 323 cells in six embryos. Division angles are binned from 0° to 360° in bins of 10°. Pink bars represent cells dividing with their anterior daughter cell facing towards the midline. Blue bars represent cells dividing with their anterior daughter cell facing away from the midline.
To quantify the orientation of cell divisions, each division was assigned an angle on the basis of the position of the anterior-most daughter cell relative to the midline. To simplify this quantification, all division angles were converted to represent divisions on the right side of the embryo. Thus, a positive value indicates an orientation obliquely towards the midline (Fig. 4, pink arrows) and a negative value indicates a division obliquely away from the midline (Fig. 4, blue arrows). A cell dividing parallel to the mediolateral axis is given an angle of 0°. Representative division angles are provided in Fig. 4A′,B. This data can also be gleaned from a color-coded rose diagram (Fig. 4C). The circles in the rose diagram refer to the number of data points for each bin (the outer ring in Fig. 4C represents n=40).
Rotations of the mitotic spindle establish division orientation
We next sought to characterize the cellular basis of oriented cell division in the closing Xenopus neural tube (supplementary material Movies 8, 9). Because our in vivo imaging approach allows for in-depth analysis of the dynamics of cell division, we were able to observe that metaphase plates often rotated just prior to anaphase (supplementary material Movie 9). Such rotation was reminiscent of mitotic-spindle rotations observed in a variety of developmentally controlled cell divisions (Bellaiche et al., 2001a; Geldmacher-Voss et al., 2003; Haydar et al., 2003; Kaltschmidt et al., 2000). We therefore examined microtubule dynamics in the neural plate by expression of tau-GFP (Kwan and Kirschner, 2005), and we observed dramatic rotations of the mitotic spindle (supplementary material Movie 8; Fig. 5B).
Fig. 5.

Mitotic-spindle rotations establish division polarity. (A) Still frames from a movie of cells expressing membrane-RFP (red) and tau-GFP (green). Mitotic spindles are easily seen and a rotation of 15° is observed before anaphase onset. White line represents the orientation of spindle set up and corresponds to the purple line. Yellow line represents the orientation of spindle-pole separation. (B) Still frames from a different sector of the same movie as in A. This spindle rotates a total of 86° before anaphase onset and occurs in a time span of 1.5 minutes. White line represents the orientation of spindle set up and corresponds to the purple line. Yellow line represents the orientation of spindle-pole separation (supplementary material Movie 8). Scale bars: 10 μm. (C) Rose diagram of the angle of 144 cells in six embryos each expressing membrane-RFP and tau-GFP. Angles of spindle set up are shown from 0° to 360° in bins of 10°. (D) Rose diagram of the angle of cell division in the same 144 cells shown in C. Angles of spindle set up are shown from 0° to 360° in bins of 10°.
We quantified the orientation of the mitotic spindles upon their initial formation during metaphase and found that spindles were initially oriented more or less randomly with respect to the midline (Fig. 5A,B, white lines; Fig. 5C). However, when the angle of the spindle in these cells was assessed again at anaphase, the pattern reflected the mediolateral orientation of telophase cell divisions described above (Fig. 5A,B, yellow lines; Fig. 5D).
Disruption of Cdc42 function results in over-rotation of mitotic spindles and shifts the angle of cell division
Because Cdc42 controls spindle positioning, we investigated whether Cdc42 also controlled spindle rotation. Indeed, we found that disruption of Cdc42 function resulted in spindles rotating to a far greater extent than those in control cells (compare supplementary material Movie 10 with Movie 9 and Fig. 6C with Fig. 6A). Control cells in the early neural plate had a restricted field of movement: the average rotation was 45° (Fig. 6E), the majority of control cells rotated between 0° and 60°, and we rarely observed control cells that rotated beyond 120° (Fig. 6B). By contrast, metaphase plates in cells expressing Cdc42-F37A rotated farther, with an average of 78° (Fig. 6E). Cdc42-F37A-expressing cells were frequently observed to rotate more than 90° (compare Fig. 6D with 6B) and, in many cases, chromosome rotation exceeded 200° (Fig. 6D). The mean degree of rotation in Cdc42-F37A-expressing cells was significantly higher than that of controls (P=0.0005 using the Mann-Whitney U-test).
Fig. 6.

Alteration of Cdc42 function causes over-rotations of mitotic spindles. (A) Still frames from a movie of a control embryo expressing histone-2B–GFP and membrane-RFP. Chromosomes are seen to condense and rotate approximately 90° prior to anaphase. The blue arrow represents the major axis of the chromosomes during the rotation and the white arrow represents the direction of rotation of the chromosomes (supplementary material Movie 9). Scale bar: 10 μm. (A′) Cartoons of the cell of interest from A. Cell outlines are shown in black and nuclei are shown in green. Chromosomes remain stable in the approximate center of the cell while rotating. (B) Histogram representing the rotation of 114 chromosomes from four control (wild-type, WT) embryos. The angle of chromosome condensation was assessed, followed by the angle of chromosome segregation. The degree of rotation was determined by the difference between these two values. These values are shown in bins of 5° from 0° to 290°. (C) Still frames from a movie of cells expressing Cdc42-F37A, histone-2B–GFP and membrane-RFP. Chromosomes are seen to condense and rotate approximately 180° prior to anaphase. The blue arrow represents the major axis of the chromosomes during the rotation and the white arrow represents the direction of the chromosomal rotation (supplementary material Movie 10). Scale bar: 10 μm. (C′) Cartoons of the cell of interest from C. Cell outlines are shown in black and nuclei are shown in green. Chromosomes do not remain stable in the cell and instead move from one side of the cell to the other during rotation. (D) Histogram representing the rotation of 82 chromosomes from four embryos expressing Cdc42-F37A. The angle of chromosome condensation was assessed, followed by the angle of chromosome segregation. The degree of rotation was determined by the difference between these two values. These values are shown in bins of 5° from 0° to 290°. (E) Graph of the average rotation of chromosomes in control cells (light-grey bar) and cells expressing Cdc42-F37A (dark-grey bar). Error bars represent standard error for each (2.9 for control and 7.0 for Cdc42-F37A). A Mann-Whitney test, with a U-statistic of 3302.5 and a U′ of 6045.5, was performed and showed a two-tailed P-value of 0.0005.
We next examined the final orientation of cell divisions following manipulation of Cdc42. Expression of Cdc42-N17 resulted in a shift in the final angle of cell divisions such that the number of cells with anterior daughters facing away from the midline increased at the expense of the number of cells with anterior daughters facing towards the midline (Fig. 7C,C′). Thus, the average angle of cell division shifted roughly 18° following expression of Cdc42-N17.
Fig. 7.

Cdc42 controls orientation of cell divisions in the early neural plate. (A) Rose diagram of division polarity of a control clone. In total, 99 cells from five embryos were binned from 0° to 360° in bins of 10°. (A′) Diagram representing the polarity of division seen in control clones of Cdc42-N17- and Cdc42-F37A-injected embryos. Double-headed pink arrows represent cells dividing in a medial manner and double-headed blue arrows represent cells dividing in a lateral manner. There is an equal number of each arrow set, indicating that an equal number of cells divide in each orientation. (B) Rose diagram of the division polarity in a clone expressing a dominant negative to Cdc42, Cdc42-F37A. In total, 73 cells from three embryos were binned from 0° to 360° in bins of 10°. (B′) Diagram representing the polarity of division seen in Cdc42-F37A clones. Pink double-headed arrows represent cells dividing in a medial manner and blue double-headed arrows represent cells dividing in a lateral manner. There are more blue double-headed arrows, indicating that more cells in the clone divide in the lateral orientation. (C) Rose diagram of the division polarity in a clone expressing a dominant negative to Cdc42, Cdc42-N17. In total, 87 cells from four embryos were binned from –90° to +90° in bins of 10°. (C′) Diagram representing the polarity of division seen in Cdc42-N17 clones. Pink double-headed arrows represent cells dividing in a medial manner and blue double-headed arrows represent cells dividing in a lateral manner. There are more blue double-headed arrows, indicating that more cells in the clone divide in the lateral orientation.
Injection of mRNA encoding Cdc42-F37A elicited the same specific alteration to the orientation of neural-cell division. In Cdc42-F37A-expressing clones, the number of cells dividing with their anterior-most daughter cell oriented laterally increased at the expense of those oriented medially, resulting in a 12° shift in the average orientation of division (Fig. 7B,B′). The Kolmogorov-Smirnov (K-S) test confirmed a significant shift in the distribution of cell-division orientations in these cells, as compared with controls (P=0.001 and 0.039, respectively). These data suggest that Cdc42 contributes to the distribution of oriented cell divisions in the Xenopus neural plate.
Spindle rotations align divisions with the cellular long axis, but cellular axis alignment is not affected by disruption of Cdc42
`Hertwig's rule' states that cells generally divide along their cellular long axis and, in mammalian cells in culture, variable rotations align the mitotic spindle with the long axis of the cell (Hertwig, 1893; O'Connell and Wang, 2000; Thery and Bornens, 2006). We therefore examined cell shape in the Xenopus neural plate and found that the long axes of cells in the neural plate were oriented in an angle comparable to the angle of observed divisions (Fig. 8A, compare to Fig. 3C). Thus, cell shape is initially polarized in these neural-plate cells, although spindles form in random orientations; spindle rotations then align the spindles such that division occurs parallel to the long axis of the cell (supplementary material Movie 8). When we assessed the alignment of cellular long axes in embryos expressing Cdc42-F37A or Cdc42-N17, we found no difference from controls (Fig. 8B,C) (K-S test, P=0.431 and 0.083, respectively). We also found that manipulation of Cdc42 did not affect the length-to-width ratio of these cells (not shown). These data suggest that Cdc42 does not govern cell polarization in this tissue, but rather acts directly to govern the positioning of the spindle within the cells.
Fig. 8.

Long axis of cells is unaltered in cells expressing dominant negatives to Cdc42. (A) Rose diagram representing the angle of the long axis of 84 dividing control neural cells in six embryos at initiation of metaphase. Angles of cellular long axis are shown from 0° to 360° in bins of 10°. Pink bars represent cells dividing with their anterior daughter cell facing towards the midline. Blue bars represent cells dividing with their anterior daughter cell facing away from the midline. (B) Rose diagram representing the angle of the long axis of 99 dividing Cdc42-F37A-expressing neural cells in nine embryos at initiation of metaphase. Angles of cellular long axis are shown from 0° to 360° in bins of 10°. (C) Rose diagram representing the angle of the long axis of 79 dividing Cdc42-N17-expressing neural cells in eight embryos at initiation of metaphase. Angles of cellular long axis are shown from 0° to 360° in bins of 10°.
Cell-division orientation is not controlled by PCP signaling and does not require normal neural tube morphogenesis
Given the very specific role for Cdc42 in governing the distribution of oriented neural-cell divisions (above), we next examined PCP signaling, which might play a broader role than Cdc42 in orienting these divisions in Xenopus, because it controls oriented divisions in the fish and the fly (Bellaiche et al., 2001a; Gho and Schweisguth, 1998; Gong et al., 2004; Kaltschmidt et al., 2000). To disrupt PCP signaling, we expressed Xdd1, a dominant-negative deletion of the PCP protein Dishevelled that effectively disrupts planar polarization of cell behavior in several contexts, including polarized cell division in zebrafish (Gong et al., 2004; Park et al., 2008; Wallingford et al., 2000). Surprisingly, overexpression of Xdd1 did not alter the predominant mediolateral orientation of these divisions and did not affect the distribution of oblique cell divisions in the neural tube (Fig. 9B, compare to 9A). The K-S test for comparison of histograms was used to confirm that there was no difference in the distribution of cell-division orientations in control and Xdd1-expressing cells (P=0.892).
Fig. 9.
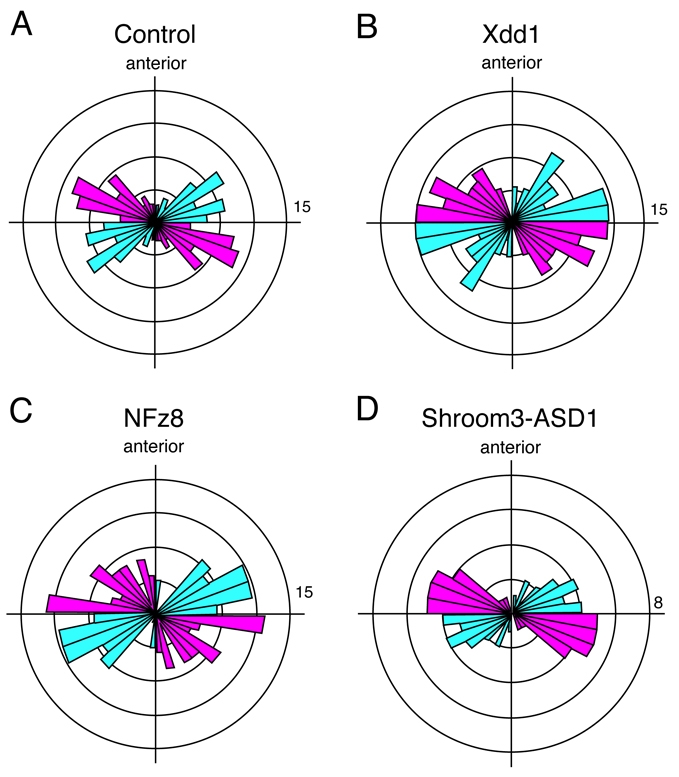
PCP signaling does not control cell-division polarity. (A) Representative rose diagram of the division polarity in a control clone. In total, 85 cells from three embryos were binned from 0° to 360° in bins of 10°. (B) Rose diagram of the division polarity in clones expressing Xdd1, a dominant negative to Dishevelled. In total, 110 cells from four embryos were binned from 0° to 360° in bins of 10°. (C) Rose diagram of the division polarity in clones expressing a dominant negative to Frizzled Xfz8N. In total, 100 cells from five embryos are binned from 0° to 360° in bins of 10°. (D) Rose diagram of the division polarity in clones expressing a dominant negative to Shroom3, the ASD1 domain. In total, 41 cells from two embryos are binned from 0° to 360° in bins of 10°. Pink bars represent cells dividing with their anterior daughter cell facing towards the midline. Blue bars represent cells dividing with their anterior daughter cell facing away from the midline.
To confirm this result, we also tested the effect of Nxfz-8, a dominant-negative version of Frizzled-8 that disrupts PCP-mediated convergent extension (Deardorff et al., 1998). Similar to Xdd1, Nxfz-8 did not alter the orientation of neural-cell divisions (Fig. 9C) (K-S test, P=0.49). Finally, we tested overexpression of wild-type Frizzled-8, which also disrupts PCP-mediated convergent extension (Deardorff et al., 1998; Wallingford et al., 2001). Overexpression of wild-type Frizzled-8 also failed to alter cell-division orientation (not shown, K-S test, P=0.296). Our data thus suggest that the PCP signaling system is dispensable for oriented cell divisions in the Xenopus neural plate, similar to PCP-independent oriented cell divisions in the extending Drosophila germ band (da Silva and Vincent, 2007).
Although dispensable for oriented cell divisions, PCP signaling is required for normal neural tube closure (Wallingford and Harland, 2002), and we noted that expression of either Xdd1 or Nxfz-8 resulted in severely disrupted neural morphogenesis (data not shown) (Wallingford and Harland, 2001; Wallingford and Harland, 2002). This phenotype serves as a useful indicator for the efficacy of our reagents, and this result also suggests that cell-division orientation is independent of neural morphogenesis. To further test the independence of cell-division orientation and neural morphogenesis, we examined cell divisions in clones of cells expressing Shroom3-ASD1, a dominant-negative form of the Shroom3 protein that potently disrupts neural tube closure (Haigo et al., 2003). Expression of Shroom3-ASD1 in clones disrupted neural tube closure (not shown) but did not change the pattern of oriented cell divisions (Fig. 9D) (K-S test, P=0.127).
Discussion
The specialization of cell-division mechanisms is a hallmark of developing embryos. Perhaps the most notable specialization is the absence of G1 and G0 phases of the cell cycle in the first several divisions of most embryos (Etkin, 1988; Masui and Wang, 1998; Philpott and Yew, 2008). Orientation of cell-division angle is another important specialization that is essential for normal development in many animals, and a small number of conserved mechanisms are typically associated with setting up these division planes. Here, we describe a novel mode of oriented cell division that occurs in the closing neural tube of Xenopus embryos.
Previous studies have shown that cells tend to divide in one particular orientation within a given tissue and rotations of the mitotic spindle are a general feature of oriented cell divisions. Typically, spindles rotate to a precisely defined degree to achieve this orientation for a given tissue (Bellaiche et al., 2001a; Bellaiche et al., 2001b; Concha and Adams, 1998; Gong et al., 2004). For example, during oriented cell divisions in Drosophila neuroblasts and in the neural plate of zebrafish embryos, mitotic spindles consistently rotate precisely 90° (Geldmacher-Voss et al., 2003; Kaltschmidt et al., 2000). In C. elegans embryos, some cells undergo rotations of 90° whereas others rotate 45° (Hyman and White, 1987). In all of these cases, however, spindles in a given cell type consistently rotate to a precisely set degree.
We found that these themes are not universal, and we describe here a system of oriented cell divisions in which variable rotations align spindles in the Xenopus neural plate. Some spindles divided in approximately the orientation that they were set up in, whereas others underwent large, rapid rotations to divide nearly perpendicular to the orientation in which they set up (Fig. 5). We also found that, although the extent of spindle rotation varied from cell to cell, all spindles rotated to the extent necessary to align them with the cellular long axis (Fig. 8). Similar orienting rotations have been observed in cultured cells when cell shape is experimentally altered (O'Connell and Wang, 2000).
It is noteworthy that the orientations of cells on opposite sides of the midline are mirror symmetric (Fig. 7, diagrams). Signals emanating from the midline have been shown to direct the cellular behavior of deep neural cells engaged in convergent extension (Ezin et al., 2003), and it is possible that similar signals guide the orientation of cellular long axes in the superficial neural plate, which appear to impose the orientation of divisions. Regardless, the orientation of cell division is independent of normal neural morphogenesis, as disruption of neural tube closure by a variety of different molecular manipulations failed to alter cell-division orientation (Fig. 9B-D). This result might be important because the neural plate is subject to defined mechanical strains, and such strains are likely to influence cell shape and cell division in epithelial sheets (Benko and Brodland, 2007; Brodland and Veldhuis, 2002; Thery and Bornens, 2006).
The oriented cell divisions characterized here are also notable for their independence from PCP signaling, which plays a major role in controlling orientated cell divisions in a wide variety of vertebrate and invertebrate animals. The divisions reported here thus bear some resemblance to the PCP-independent oriented cell divisions in the elongating Drosophila germ band (da Silva and Vincent, 2007). Instead of PCP signaling, we found that Cdc42, which has not previously been shown to influence oriented cell division in vertebrate embryos, plays a role in establishing division orientation in the closing Xenopus neural tube. Cells expressing two different dominant negatives to Cdc42, Cdc42-N17 or Cdc42-F37A shifted their average angle of these divisions by roughly 15° (Fig. 7B,C). This shift was modest, but very consistent. This modest role for Cdc42 in orienting these divisions suggests that there are multiple signaling inputs controlling spindle orientation in the Xenopus neural plate. Such a mechanism is known to act in Drosophila neuroblasts, in which disruption of Ric-8 results in consistent misalignment of spindle by only fewer than 10° (David et al., 2005).
We also found that alteration of Cdc42 function has a dramatic effect on the extent of mitotic-spindle rotation in the Xenopus neural plate (Fig. 6C-F). Whereas control cells have a limited range of rotation, with very few spindles rotating more than 120°, cells with altered Cdc42 function had spindles that rotated in excess of 200° (Fig. 6B,D). These data suggest a model for the orientation of cell divisions in the neural plate. Spindle positioning in general involves interactions between astral microtubules and the cell cortex (Gundersen et al., 2004; O'Connell and Wang, 2000; Siller and Doe, 2008), and we propose that spindle rotations are initiated essentially at random, and that astral microtubules identify `catch points' on the cell cortex that stop rotation prior to anaphase and in accordance with the long axis of a cell. One such catch point that is necessary for stopping spindle rotation so that the final division angle is aligned with the cellular long axis is Cdc42-dependent. In this model, if the Cdc42-dependent catch point is disrupted, spindles rotate excessively, and eventually are able to stop by responding to other, Cdc42-independent mechanisms. This general mechanism, by which multiple inputs `compete' to control spindle orientation, has been demonstrated in Drosophila neuroblasts (Bellaiche et al., 2001a; Bellaiche et al., 2001b; David et al., 2005).
We also found that, during cell divisions in the Xenopus neural plate, observed in vivo, spindles in cells expressing Cdc42-N17 or Cdc42-F37A were not stably positioned during metaphase. Chromosomes `bounce' from one side of the cell to the other during and after spindle rotation (Fig. 2A and Fig. 6C). This phenotype might be related to the chaotic interactions between meiotic spindles and the cell cortex of mouse oocytes expressing mutants of Cdc42 (Na and Zernicka-Goetz, 2006). Moreover, such hyperactive nuclear movement is linked to spindle positioning in C. elegans (Couwenbergs et al., 2004). Cdc42 has been shown to control the reorientation of the microtubule-organizing center (MTOC) in migrating cells by governing association of the microtubules with the cell cortex (Etienne-Manneville and Hall, 2001; Palazzo et al., 2001). Our results suggest that a similar mechanism functions to position the spindle in Xenopus neural-plate cells. Curiously, Cdc42 had no such effect on spindle positioning in Xenopus epidermal cells observed in vivo.
Finally, this work provides a rare glimpse into the process of cell division in an intact epithelial cell sheet that is simultaneously engaged in both massive proliferation and a massive morphogenetic event. Our data reveal cell-type-specific roles for Cdc42 in spindle positioning and spindle orientation, suggesting that additional in vivo studies will be important.
Materials and Methods
Collection and manipulation of embryos
Female Xenopus laevis were injected with 700 ml of human chorionic gonadotropin hormone (HCG) and stored at 18°C overnight. The next day, eggs were isolated and fertilized. Embryos were de-jellied using a 3% cysteine solution in 1/3× Marc's modified Ringers solution (MMR).
Time-lapse analysis of cell divisions in the neural plate and tail epidermis
Embryos were injected dorsally, at the four-cell stage, with 200 pg histone-eGFP, 200 pg membrane-RFP, 50 pg tau-eGFP, 200 pg histone-RFP or any combination of these to examine cell divisions in the neural plate. To examine the epidermis, all cells were injected at the four-cell stage (Fig. 1). To examine the neural plate, embryos were grown to stage 13 and then placed upside down in a culture dish. This dish contained 2% agarose with wells made from a hair comb. To image the tail epidermis, embryos were grown to stage 30 and covered in agarose in a culture dish. Both of these sets of embryos were imaged on a Zeiss Pascal LSM5, at 10× (NA, 0.3), 20× (NA, 0.5), 40× (oil; NA, 1.3) or 63× (oil; NA, 1.4) and cell divisions were analyzed using Image-Pro Plus.
Determination of division polarity
Stacks were projected and exported from the Zeiss LSM5 Pascal program and analyzed using Image-Pro Plus. The polarity of cell division was assessed at the onset of telophase for all cells. Lines were drawn bisecting the center of each set of daughter chromosomes in a posterior-to-anterior manner. The features of these lines were then exported to Microsoft Excel. The angles of divisions were converted placed into a –90° to +90° range and histograms were made using Delta-Graph. K-S tests were performed using an online tool available at http://www.physics.csbsju.edu/stats/KS-test.html.
Effect of molecular effectors on cell-division polarity
To examine the effect of PCP signaling and Cdc42 function, embryos were first injected with 200 pg of histone-eGFP along the entire dorsal side at the four-cell stage. These embryos were grown to the eight-cell stage and 200 pg of membrane-RFP along with either 500 pg Xdd1, 500 pg Xfz8N, 1 ng Shroom3-ASD1, 750 pg Cdc42-F37A or 750 pg Cdc42-N17 was injected into one of the dorsal animal blastomeres (Fig. 1C). To examine the effect of Cdc42 on the tail epidermis, embryos were injected into one of the ventral cells with 150 pg of histone-eGFP, 150 pg of membrane-RFP and 750 pg of Cdc42-N17 or 750 pg of Cdc42-F37A. Embryos were then imaged and analyzed as above.
Quantifying spindle rotations
This longer axis of the metaphase spindle was assessed upon initial formation, and again 1 minute before anaphase onset. This was done for control cells and cells expressing Cdc42-F37A. The difference of these angles were determined and plotted in Delta-Graph. The Mann-Whitney comparison test was conducted on the two sets of data using GraphPad Instat software.
Supplementary Material
Supplementary material available online at http://jcs.biologists.org/cgi/content/full/122/14/2481/DC1
We thank Chanjae Lee for technical assistance and Andy Ewald for critical reading. This work was funded by the NIH/NIGMS (R01-GM74104). Deposited in PMC for release after 12 months.
References
- Ban, R., Irino, Y., Fukami, K. and Tanaka, H. (2004). Human mitotic spindle-associated protein PRC1 inhibits MgcRacGAP activity toward Cdc42 during the metaphase. J. Biol. Chem. 279, 16394-16402. [DOI] [PubMed] [Google Scholar]
- Bellaiche, Y., Gho, M., Kaltschmidt, J. A., Brand, A. H. and Schweisguth, F. (2001a). Frizzled regulates localization of cell-fate determinants and mitotic spindle rotation during asymmetric cell division. Nat. Cell Biol. 3, 50-57. [DOI] [PubMed] [Google Scholar]
- Bellaiche, Y., Radovic, A., Woods, D. F., Hough, C. D., Parmentier, M. L., O'Kane, C. J., Bryant, P. J. and Schweisguth, F. (2001b). The Partner of Inscuteable/Discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell 106, 355-366. [DOI] [PubMed] [Google Scholar]
- Bement, W. M., Benink, H. A. and von Dassow, G. (2005). A microtubule-dependent zone of active RhoA during cleavage plane specification. J. Cell Biol. 170, 91-101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benko, R. and Brodland, G. W. (2007). Measurement of in vivo stress resultants in neurulation-stage amphibian embryos. Ann. Biomed. Eng. 35, 672-681. [DOI] [PubMed] [Google Scholar]
- Brodland, G. W. and Veldhuis, J. H. (2002). Computer simulations of mitosis and interdependencies between mitosis orientation, cell shape and epithelia reshaping. J. Biomech. 35, 673-681. [DOI] [PubMed] [Google Scholar]
- Caviston, J. P., Longtine, M., Pringle, J. R. and Bi, E. (2003). The role of Cdc42p GTPase-activating proteins in assembly of the septin ring in yeast. Mol. Biol. Cell 14, 4051-4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chalmers, A. D., Welchman, D. and Papalopulu, N. (2002). Intrinsic differences between the superficial and deep layers of the Xenopus ectoderm control primary neuronal differentiation. Dev. Cell 2, 171-182. [DOI] [PubMed] [Google Scholar]
- Chalmers, A. D., Strauss, B. and Papalopulu, N. (2003). Oriented cell divisions asymmetrically segregate aPKC and generate cell fate diversity in the early Xenopus embryo. Development 130, 2657-2668. [DOI] [PubMed] [Google Scholar]
- Concha, M. L. and Adams, R. J. (1998). Oriented cell divisions and cellular morphogenesis in the zebrafish gastrula and neurula: a time-lapse analysis. Development 125, 983-994. [DOI] [PubMed] [Google Scholar]
- Couwenbergs, C., Spilker, A. C. and Gotta, M. (2004). Control of embryonic spindle positioning and Galpha activity by C. elegans RIC-8. Curr. Biol. 14, 1871-1876. [DOI] [PubMed] [Google Scholar]
- da Silva, S. M. and Vincent, J. P. (2007). Oriented cell divisions in the extending germband of Drosophila. Development 134, 3049-3054. [DOI] [PubMed] [Google Scholar]
- David, N. B., Martin, C. A., Segalen, M., Rosenfeld, F., Schweisguth, F. and Bellaiche, Y. (2005). Drosophila Ric-8 regulates Galphai cortical localization to promote Galphai-dependent planar orientation of the mitotic spindle during asymmetric cell division. Nat. Cell Biol. 7, 1083-1090. [DOI] [PubMed] [Google Scholar]
- Deardorff, M. A., Tan, C., Conrad, L. J. and Klein, P. S. (1998). Frizzled-8 is expressed in the Spemann organizer and plays a role in early morphogenesis. Development 125, 2687-2700. [DOI] [PubMed] [Google Scholar]
- Etienne-Manneville, S. and Hall, A. (2001). Integrin-mediated activation of Cdc42 controls cell polarity in migrating astrocytes through PKCzeta. Cell 106, 489-498. [DOI] [PubMed] [Google Scholar]
- Etkin, L. D. (1988). Regulation of the mid-blastula transition in amphibians. Dev. Biol. 5, 209-225. [DOI] [PubMed] [Google Scholar]
- Ezin, A. M., Skoglund, P. and Keller, R. (2003). The midline (notochord and notoplate) patterns the cell motility underlying convergence and extension of the Xenopus neural plate. Dev. Biol. 256, 100-114. [DOI] [PubMed] [Google Scholar]
- Fischer, E., Legue, E., Doyen, A., Nato, F., Nicolas, J. F., Torres, V., Yaniv, M. and Pontoglio, M. (2006). Defective planar cell polarity in polycystic kidney disease. Nat. Genet. 38, 21-23. [DOI] [PubMed] [Google Scholar]
- Garcia, Z., Silio, V., Marques, M., Cortes, I., Kumar, A., Hernandez, C., Checa, A. I., Serrano, A. and Carrera, A. C. (2006). A PI3K activity-independent function of p85 regulatory subunit in control of mammalian cytokinesis. EMBO J. 25, 4740-4751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geldmacher-Voss, B., Reugels, A. M., Pauls, S. and Campos-Ortega, J. A. (2003). A 90-degree rotation of the mitotic spindle changes the orientation of mitoses of zebrafish neuroepithelial cells. Development 130, 3767-3780. [DOI] [PubMed] [Google Scholar]
- Gho, M. and Schweisguth, F. (1998). Frizzled signalling controls orientation of asymmetric sense organ precursor cell divisions in Drosophila. Nature 393, 178-181. [DOI] [PubMed] [Google Scholar]
- Gonczy, P. and Rose, L. S. (2005). Asymmetric cell division and axis formation in the embryo. WormBook 1-20. [DOI] [PMC free article] [PubMed]
- Gong, Y., Mo, C. and Fraser, S. E. (2004). Planar cell polarity signalling controls cell division orientation during zebrafish gastrulation. Nature 430, 689-693. [DOI] [PubMed] [Google Scholar]
- Gotta, M., Abraham, M. C. and Ahringer, J. (2001). CDC-42 controls early cell polarity and spindle orientation in C. elegans. Curr. Biol. 11, 482-488. [DOI] [PubMed] [Google Scholar]
- Gotz, M. and Huttner, W. B. (2005). The cell biology of neurogenesis. Nat. Rev. Mol. Cell. Biol. 6, 777-788. [DOI] [PubMed] [Google Scholar]
- Gundersen, G. G., Gomes, E. R. and Wen, Y. (2004). Cortical control of microtubule stability and polarization. Curr. Opin. Cell Biol. 16, 106-112. [DOI] [PubMed] [Google Scholar]
- Haigo, S. L., Hildebrand, J. D., Harland, R. M. and Wallingford, J. B. (2003). Shroom induces apical constriction and is required for hingepoint formation during neural tube closure. Curr. Biol. 13, 2125-2137. [DOI] [PubMed] [Google Scholar]
- Hartenstein, V. (1989). Early neurogenesis in Xenopus: the spatio-temporal pattern of proliferation and cell lineages in the embryonic spinal cord. Neuron 3, 399-411. [DOI] [PubMed] [Google Scholar]
- Haydar, T. F., Ang, E., Jr and Rakic, P. (2003). Mitotic spindle rotation and mode of cell division in the developing telencephalon. Proc. Natl. Acad. Sci. USA 100, 2890-2895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hertwig, O. (1893). Ueber den Werth der ersten Furchungszellen fur die Organbildung des Embryo: Experimentelle Studien am Froschund Tritonei. Arch. Mikrosk. Anat. 42, 662-804. [Google Scholar]
- Hyman, A. A. and White, J. G. (1987). Determination of cell division axes in the early embryogenesis of Caenorhabditis elegans. J. Cell Biol. 105, 2123-2135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joberty, G., Perlungher, R. R., Sheffield, P. J., Kinoshita, M., Noda, M., Haystead, T. and Macara, I. G. (2001). Borg proteins control septin organization and are negatively regulated by Cdc42. Nat. Cell Biol. 3, 861-866. [DOI] [PubMed] [Google Scholar]
- Kaltschmidt, J. A., Davidson, C. M., Brown, N. H. and Brand, A. H. (2000). Rotation and asymmetry of the mitotic spindle direct asymmetric cell division in the developing central nervous system. Nat. Cell Biol. 2, 7-12. [DOI] [PubMed] [Google Scholar]
- Karnoub, A. E., Symons, M., Campbell, S. L. and Der, C. J. (2004). Molecular basis for Rho GTPase signaling specificity. Breast Cancer Res. Treat. 84, 61-71. [DOI] [PubMed] [Google Scholar]
- Kieserman, E. K., Glotzer, M. and Wallingford, J. B. (2008). Developmental regulation of central spindle assembly and cytokinesis during vertebrate embryogenesis. Curr. Biol. 18, 116-123. [DOI] [PubMed] [Google Scholar]
- Kozminski, K. G., Beven, L., Angerman, E., Tong, A. H., Boone, C. and Park, H. O. (2003). Interaction between a Ras and a Rho GTPase couples selection of a growth site to the development of cell polarity in yeast. Mol. Biol. Cell 14, 4958-4970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwan, K. M. and Kirschner, M. W. (2005). A microtubule-binding Rho-GEF controls cell morphology during convergent extension of Xenopus laevis. Development 132, 4599-4610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamarche, N., Tapon, N., Stowers, L., Burbelo, P. D., Aspenstrom, P., Bridges, T., Chant, J. and Hall, A. (1996). Rac and Cdc42 induce actin polymerization and G1 cell cycle progression independently of p65PAK and the JNK/SAPK MAP kinase cascade. Cell 87, 519-529. [DOI] [PubMed] [Google Scholar]
- Ma, C., Benink, H. A., Cheng, D., Montplaisir, V., Wang, L., Xi, Y., Zheng, P. P., Bement, W. M. and Liu, X. J. (2006). Cdc42 activation couples spindle positioning to first polar body formation in oocyte maturation. Curr. Biol. 16, 214-220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masui, Y. and Wang, P. (1998). Cell cycle transition in early embryonic development of Xenopus laevis. Biol. Cell 90, 537-548. [PubMed] [Google Scholar]
- Na, J. and Zernicka-Goetz, M. (2006). Asymmetric positioning and organization of the meiotic spindle of mouse oocytes requires CDC42 function. Curr. Biol. 16, 1249-1254. [DOI] [PubMed] [Google Scholar]
- O'Connell, C. B. and Wang, Y. L. (2000). Mammalian spindle orientation and position respond to changes in cell shape in a dynein-dependent fashion. Mol. Biol. Cell 11, 1765-1774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palazzo, A. F., Joseph, H. L., Chen, Y. J., Dujardin, D. L., Alberts, A. S., Pfister, K. K., Vallee, R. B. and Gundersen, G. G. (2001). Cdc42, dynein, and dynactin regulate MTOC reorientation independent of Rho-regulated microtubule stabilization. Curr. Biol. 11, 1536-1541. [DOI] [PubMed] [Google Scholar]
- Park, T. J., Mitchell, B. J., Abitua, P. B., Kintner, C. and Wallingford, J. B. (2008). Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells. Nat. Genet. 40, 871-879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Philpott, A. and Yew, P. R. (2008). The Xenopus cell cycle: an overview. Mol. Biotechnol. 39, 9-19. [DOI] [PubMed] [Google Scholar]
- Richman, T. J., Sawyer, M. M. and Johnson, D. I. (2002). Saccharomyces cerevisiae Cdc42p localizes to cellular membranes and clusters at sites of polarized growth. Eukaryot. Cell 1, 458-468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saburi, S., Hester, I., Fischer, E., Pontoglio, M., Eremina, V., Gessler, M., Quaggin, S. E., Harrison, R., Mount, R. and McNeill, H. (2008). Loss of Fat4 disrupts PCP signaling and oriented cell division and leads to cystic kidney disease. Nat. Genet. 40, 1010-1015. [DOI] [PubMed] [Google Scholar]
- Saka, Y. and Smith, J. C. (2001). Spatial and temporal patterns of cell division during early Xenopus embryogenesis. Dev. Biol. 229, 307-318. [DOI] [PubMed] [Google Scholar]
- Saka, Y. and Smith, J. C. (2004). A Xenopus tribbles orthologue is required for the progression of mitosis and for development of the nervous system. Dev. Biol. 273, 210-225. [DOI] [PubMed] [Google Scholar]
- Sausedo, R. A., Smith, J. L. and Schoenwolf, G. C. (1997). Role of nonrandomly oriented cell division in shaping and bending of the neural plate. J. Comp. Neurol. 381, 473-488. [PubMed] [Google Scholar]
- Siller, K. H. and Doe, C. Q. (2008). Lis1/dynactin regulates metaphase spindle orientation in Drosophila neuroblasts. Dev. Biol. 319, 1-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thery, M. and Bornens, M. (2006). Cell shape and cell division. Curr. Opin. Cell Biol. 18, 648-657. [DOI] [PubMed] [Google Scholar]
- Wallingford, J. B. (2005). Neural tube closure and neural tube defects: studies in animal models reveal known knowns and known unknowns. Am. J. Med. Genet. C Semin. Med. Genet. 135C, 59-68. [DOI] [PubMed] [Google Scholar]
- Wallingford, J. B. and Harland, R. M. (2001). Xenopus dishevelled signaling regulates both neural and mesodermal convergent extension: parallel forces elongating the body axis. Development 128, 2581-2592. [DOI] [PubMed] [Google Scholar]
- Wallingford, J. B. and Harland, R. M. (2002). Neural tube closure requires Dishevelled-dependent convergent extension of the midline. Development 129, 5815-5825. [DOI] [PubMed] [Google Scholar]
- Wallingford, J. B., Rowning, B. A., Vogeli, K. M., Rothbächer, U., Fraser, S. E. and Harland, R. M. (2000). Dishevelled controls cell polarity during Xenopus gastrulation. Nature 405, 81-85. [DOI] [PubMed] [Google Scholar]
- Wallingford, J. B., Vogeli, K. M. and Harland, R. M. (2001). Regulation of convergent extension in Xenopus by Wnt5a and Frizzled-8 is independent of the canonical Wnt pathway. Int. J. Dev. Biol. 45, 225-227. [PubMed] [Google Scholar]
- Wilcock, A. C., Swedlow, J. R. and Storey, K. G. (2007). Mitotic spindle orientation distinguishes stem cell and terminal modes of neuron production in the early spinal cord. Development 134, 1943-1954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuda, S., Oceguera-Yanez, F., Kato, T., Okamoto, M., Yonemura, S., Terada, Y., Ishizaki, T. and Narumiya, S. (2004). Cdc42 and mDia3 regulate microtubule attachment to kinetochores. Nature 428, 767-771. [DOI] [PubMed] [Google Scholar]
- Yoshizaki, H., Ohba, Y., Kurokawa, K., Itoh, R. E., Nakamura, T., Mochizuki, N., Nagashima, K. and Matsuda, M. (2003). Activity of Rho-family GTPases during cell division as visualized with FRET-based probes. J. Cell Biol. 162, 223-232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, S., Han, J., Sells, M. A., Chernoff, J., Knaus, U. G., Ulevitch, R. J. and Bokoch, G. M. (1995). Rho family GTPases regulate p38 mitogen-activated protein kinase through the downstream mediator Pak1. J. Biol. Chem. 270, 23934-23936. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.