

Abstract
The size and font of target words were manipulated in a masked repetition priming paradigm with ERP recordings. Repetition priming effects were found in four ERP components: the N/P150, N250, P325, and N400. Neither a change in font nor a change in size across prime and target were found to affect repetition priming in the N250, P325, and N400 components. Changing font was, however, found to affect repetition priming in the N/P150 component, while the interaction between repetition priming and size was not significant in this component. These results confirm our interpretation of the N/P150 as a component sensitive to feature-level processing, and suggest that the type of prelexical and lexical processing reflected in the N250, P325, and N400 components is performed on representations that are invariant to changes in both font and size.
Masked priming has evolved over the last two decades to become the predominant tool for behavioural scientists interested in describing the fast, highly automatised processes involved in reading printed words. The research reported in the present study describes a novel combination of the masked priming technique with the recording of event-related potentials (ERPs) in order to provide a fine-grained analysis of the time-course of component processes in word recognition. Applied in several recent studies, this methodology has enabled the observation of a series of components hypothesised to be sensitive to featural, prelexical (orthographic and phonological), lexical, and semantic information during printed word perception (Grainger, Kiyonaga, & Holcomb, 2006; Holcomb & Grainger, 2006; Kiyonaga, Midgley, Holcomb, & Grainger, 2007).
Holcomb and Grainger (2006) described four such components, the P150, N250, P325, and N400, obtained with masked repetition priming, and proposed a tentative mapping of these ERP components onto distinct levels of processing occurring during visual word recognition (see also Grainger & Holcomb, in press). The first of these, the P150 (here referred to as the N/P150), started as early as 90 ms and was over by 180 ms. This was a relatively focal component that produced priming effects that were positive-going at occipital sites (especially over the right hemisphere) and negative-going at more anterior sites. The distributional and temporal similarity of this effect across several studies using words (Holcomb & Grainger, 2006), single letters (Petit, Midgley, Holcomb, & Grainger, 2006) and pictures of objects (Eddy, Schmid, & Holcomb, 2006) suggests that it reflects an early process, possibly one that is involved in mapping visual features onto higher level representations (Grainger & Holcomb, in press). The slightly later N250, which started around 180 ms and ended close to 300 ms, produced a more widely distributed effect that was somewhat larger over more anterior scalp sites than over the most posterior sites. This component took the form of a larger negativity to target words that were unrelated to their preceding prime words (e.g., phone-TRUCK) than to target words that shared letters with their primes. In the case of prime-target letter overlap, the N250 was most attenuated for targets that overlapped the prime in every letter position (i.e., full repetitions such as truck-TRUCK) and was of intermediate amplitude for targets that overlapped their primes in all but one letter position (e.g., trock-TRUCK). Grainger and Holcomb (in press) have argued that such N250 effects, which were not seen for individual letters (Petit et al., 2006) or objects (Eddy et al., 2006), reflect the process whereby abstract orthographic representations (letters and letter clusters) are mapped onto higher-level lexical representations.
Immediately following the N250 was a positive going potential that Holcomb and Grainger (2006) labelled the P325. This component was more positive to target words that overlapped their primes in all letter positions (truck-TRUCK) and was more negative to unrelated and partially overlapping prime-target pairs (phone-TRUCK and trock-TRUCK). Grainger and Holcomb suggested that this component may be sensitive to lexical selection - the process whereby a single lexical candidate remains active during word processing. The final effect reported by Holcomb and Grainger (2006) was a component very similar to the classic N400, starting around 350 ms and ending near 550 ms and having a central-posterior scalp distribution. Like the earlier N250, it was most negative to targets that were completely unrelated to their primes, least negative to targets that were complete repetitions of their primes and intermediate to targets that partially overlapped their primes. Grainger and Holcomb proposed that these N400 repetition effects in the masked priming paradigm reflect the process of mapping word level representations onto semantic representations.
The present study is part of a research program that, building on these foundations, attempts to describe in more detail the nature of the processing reflected in the different ERP components. In particular, the present study focuses on the earliest phases of processing in visual word recognition - those involved in mapping feature information onto abstract letter identities, and the formation of a prelexical orthographic code. These processes were described by Grainger and van Heuven (2003) in their model of orthographic processing couched within the framework of a generic interactive-activation model (McClelland & Rumelhart, 1981). In this model, a bank of location-specific letter detectors receive featural information from the stimulus and send activation onto location-invariant representations that code for the relative position of letters within the stimulus (see Caramazza & Hillis, 1990, for a similar proposal). Grainger and van Heuven proposed ‘open-bigrams’, that is contiguous and non-contiguous ordered letter combinations, as a mechanism for computing a location-invariant, relative-position orthographic code (see also Whitney, 2001). It is this prelexical orthographic code that then activates whole-word orthographic representations, which in turn activate semantic representations. This general approach was recently adopted and extended by Dehaene, Cohen, Sigman, and Vinckier (2005) in their ‘neural’ model of visual word recognition. Dehaene et al. describe a more graded hierarchical approach that was only implicit in the simplified picture offered by Grainger and van Heuven.
One key element in Grainger and van Heuven’s model is the alphabetic array, a hypothesised bank of letter detectors that perform parallel, independent letter identification. These letter detectors are assumed to be invariant to the physical characteristics of letters (both their size and their shape), but not invariant to position. According to this model, featural information at a given location along the horizontal meridian is mapped onto abstract letter representations that code for the presence of a given letter identity at that particular location. These abstract letter representations are thought to be activated equally well by the same letter written in a different case, in a different font, or a different size (within a given level of tolerance for such variations). Processing upstream of the alphabetic array must therefore also be invariant to size and shape. More specifically, the next stage of processing, referred to as the relative-position map, is thought to code for the within-stimulus position of letter identities independently of their shape and their size, and independently of the location of the stimulus word.
In the present study we focus on size and shape invariance1 in visual word recognition, by seeking evidence in the EEG signal concerning the precise moment in time that such invariance is achieved during the word recognition process. As noted above, according to Grainger and van Heuven’s (2003) model of orthographic processing, size and shape invariance are achieved early in the processing stream, at the level of location-specific letter detectors. On the basis of our prior work on isolated letter perception (Petit et al., 2006), it is estimated that shape invariance is achieved between 220 and 300ms post-target onset. In that study, masked repetition priming effects for single letter stimuli were found to be independent of whether or not primes and targets were presented in the same case or in different case after approximately 250 ms of target processing. These results obtained with isolated letters would roughly correspond to the time window of the N250 ERP component found with strings of letters in other ERP studies (Grainger et al., 2006; Holcomb & Grainger, 2006; Kiyonaga et al., 2007). Processing occurring before this point in time should be sensitive to changes in shape, with different representations involved in the processing of the same letter identity presented in a physically different format. Indeed, in the Petit et al. study, case-specific repetition priming for single letters was evident between 180 and 220 ms post-target onset. This provides us with an upper limit for observing shape invariance in letter perception, since it is possible that some form of case-specific shape invariance is achieved before case-independent letter representations have been reached (i.e., mapping different physical forms of the same lower-case letter printed in different fonts onto a case-specific letter representation). Thus, our prior work on isolated letter perception (Petit et al., 2006) suggests that shape-specific letter processing is performed before approximately 220 ms post-target onset. Such processing might involve the mapping of size invariant features (letter parts) onto case-specific letter representations. According to the results of Petit et al., feature-level processing of isolated letters occurs between 100 and 180 ms post-target onset - the N/P150 ERP component.
In the present study we manipulated the size and font of target stimuli in a masked repetition priming experiment, such that prime and target could be the same size or not, and printed in the same font or not. Our font manipulation provides a manipulation of visual features that is stronger than the standard manipulation of letter case, given the large amount of feature overlap across many upper and lower case letters (but see Dehaene et al. (2004) and Petit et al. (2006) for case manipulations involving large changes in feature overlap). The lower-case version of the ‘Gigi’ font provides a very distinct visual format compared with the more standard ‘Arial’ font (see Figure 2). Furthermore, our font manipulation allows us to vary visual similarity across prime and target without a change in case. This is important given that a case change could involve mechanisms specific to the processing of lower- and upper-case letters, independently of visual feature overlap (i.e., case-specific letter representations).2
Figure 2.

Schematic of a typical trial with example stimuli.
The present experiment was designed to test the following predictions:
As shown in our prior work (Holcomb & Grainger, 2006; Kiyonaga et al., 2007), repetition priming should modulate the amplitude of N/P150, N250, P325, and N400 ERP components.
Manipulations of target font should modulate the amount of repetition priming found in the N/P150 ERP component, with a reduction in priming effects when targets are presented in a different font compared with prime stimuli.
If size invariance is achieved before shape invariance in printed word perception, then a change in size across prime and target should not modulate repetition priming effects on N/P150 amplitude.
Repetition priming effects in later components (N250, P325, N400) should not be modulated by a change in size or font between prime and target.
METHODS
Participants
Twenty-four participants (11 females, age range 18-23, average age 20.4 years) were recruited from Tufts University and compensated for their time. All were right-handed native speakers of English with normal or corrected-to-normal visual acuity with no history of neurological insult or language disability. Participants who had learned a second language before puberty were excluded.
Apparatus
Electroencephalograms were collected using 32-channel caps (Electro-cap International). The tin electrodes were arranged according to International 10-20 system (see Figure 1). In addition, an electrode below the left eye (LE) was used to monitor for blinks and vertical eye movements and the bipolar montage of F7/F8 was used to monitor for horizontal eye movements. Two electrodes were placed behind the ears on the mastoid bone; the left mastoid site (A1) was used as a reference for all electrodes, and the right mastoid site (A2) was also recorded to evaluate differential mastoid activity. Impedance was measured at less than 2 kΩ for all electrode sites except the lower eye channel which was below 10 kΩ. The EEG was amplified using an SA Bioamplifier (SA Instruments, San Diego, CA) operating on a bandpass of .01 and 40 Hz. The digitising computer continuously sampled the EEG at a rate of 200 Hz, while a stimulus computer simultaneously presented stimuli to a 19-inch CRT monitor located 54 inches in front of the participant (all stimuli subtended less than 2 degrees of horizontal visual angle).
Figure 1.
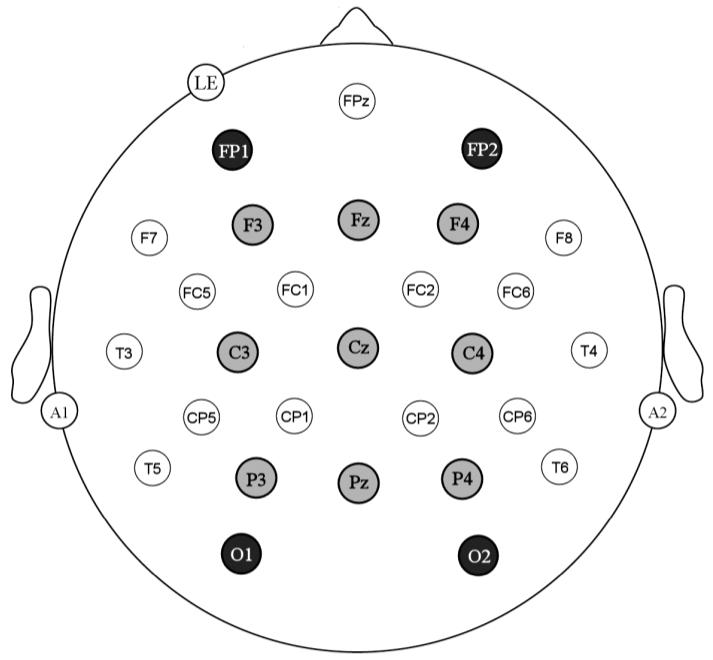
Electrode montage used in this experiment. Light grey sites were those used in the main analyses of the 180-550 ms epochs and the four darker grey sites are those used to analyse the earliest epoch (90-180 ms).
Stimuli
All critical stimuli were four to six letter words which were presented in two different sizes and two different fonts, and were primed either by the same word or a different word. A total of 480 words were selected for the experiment. The factors Repetition (prime is the same word as the target or a different word), Font (targets were in Arial or Gigi font - primes were always in Arial font), and Size (targets were in 30 point or in 44 point - primes were always in 30 point) were crossed in a 2 × 2 × 2 repeated measures factorial design, giving 80 items per condition. The eight trial types were organised into two blocks - one in which all targets were in the Arial font and one in which all targets were in the Gigi font. The order of blocks was counter-balanced across participants. Both primes and targets were displayed in lower-case. Standard counterbalancing was used with six different lists such that each target word was tested in each of the six experimental conditions but with different participants.
Procedure
Stimuli were presented on a 19-inch CRT monitor at a viewing distance of 60 inches resulting in all stimuli subtending less than 2 degrees of horizontal visual angle. Each trial consisted of a 200 ms forward patterned mask, a 50 ms prime word and a 20 ms backward mask. A 300 ms target word immediately followed the backward mask, and the trial ended with a 3400 ms interval divided into a 900 ms black screen, a 2000 ms stimulus indicating that blinking was allowed and finally a 500 ms blank screen warning that the next trial was about to begin (see Figure 2 for a schematic of a typical trial and example stimuli). Participants were asked to silently read all words displayed on the screen and to press a single button on a game pad resting in their lap as quickly as possible whenever they detected a ‘probe word’ in the designated semantic category of animal names (15% of trials). Half of all animal probes occurred in the target position and half in the masked prime position (the latter as a check on prime visibility).
After completing informed consent and handedness forms, participants were fitted with an electrode cap and seated in a comfortable armchair in a sound-attenuated room. Before each experimental block, participants were presented with a practice block to familiarise them with the task. Before the Gigi experimental block, participants were asked to indicate when they were approximately as comfortable reading words in Gigi as in Arial. Practice was continued until they reached this level.
Design and analysis
A multi-factor repeated measures design was used to analyse the ERP data. This included factors of target Font (same font Arial trials versus trials where the prime was in Arial and the target was in Gigi), target Size (target size the same as prime versus target size larger than prime), and target Repetition (prime and target are the same word or different words). The dependent measure was the mean amplitude of the target ERPs in four consecutive time windows used in our previous masked repetition priming studies (90-180 ms, 180-300 ms, 300-375 ms, and 375-550 ms). The main analyses were applied to the three later time windows (180-300 ms, 300-375 ms, and 375-550 ms) and contrasted nine electrode sites arranged in three columns and three rows across the scalp (see highlighted sites in Figure 1). This resulted in two separate three level factors: Laterality (left vs. centre vs. right) and Electrode (frontal vs. central vs. posterior). A second set of analyses contrasted two additional sets of lateral electrodes at the most frontal and most posterior sites (FP1/FP2 vs. O1/O2) in the earliest time window (90-180 ms). These sites were selected because previous masked priming ERP studies have indicated that the earliest effects of priming are largely restricted to these sites (e.g., Eddy et al., 2006; Holcomb & Grainger, 2006; Petit et al., 2006). The Geisser-Greenhouse (Geisser & Greenhouse, 1959) correction was applied to all repeated measures with more than one degree of freedom.
We only report results concerning the main effect of Repetition and the interaction of this factor with Size and Font, as well as higher-order interactions including electrode site. Simple effects of Size and Font (and interactions of these variables with electrode site) cannot be unambiguously interpreted in the present design, since changes in target size or font are confounded with whether or not primes and targets are presented in the same size/font.
RESULTS
Electrophysiological data
90-180ms post-target (N/P150)
Repetition priming modulated the amplitude of an early posterior negativity and anterior positivity peaking near 150 ms post-target onset. Having the same words in the prime and target position significantly enhanced the size of these components (i.e., the posterior negativity was more negative and the anterior positivity was more positive) as revealed by a significant interaction with the AntPost variable, F(1, 23)=6.93, p=.01. There was also an interaction between Font, Repetition and AntPos, F(1, 23)=6.09, p=.021, and between Font, Repetition, AntPos, and Laterality, F(1, 23)=8.03, p=.009. These interactions were due to the Repetition by AntPos effect being larger for the Arial targets (with both primes and targets in Arial font) than for the Gigi targets (with primes in Arial font) - especially over the right hemisphere (see Figure 3a). In the same font condition (Arial targets) there was a significant Repetition by AntPos interaction, F(1, 23)=15.78, p=.001, but this effect was not significant in the different font condition, F(1, 23)=0.267, p=.610. As can be seen in Figure 3b and Table 1 there was no interaction between Size and Repetition in this time window, F(1, 23)=0.378, p=.545.
Figure 3.

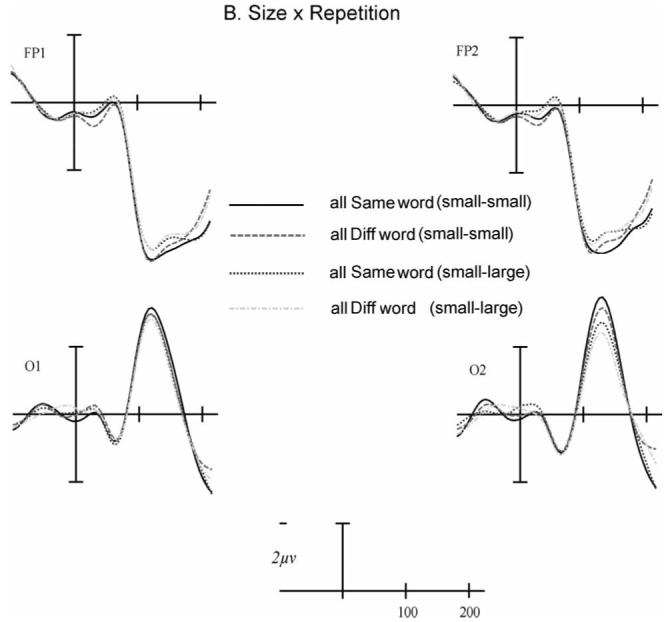
Event-related potentials (ERPs) at two frontal and posterior sites showing an interaction between Font and Repetition (panel A) and no interaction between Size and Repetition (panel B). The abscissa shows time (in milliseconds) from target onset.
TABLE 1.
Time-course analysis of ERP effects
| Repetition | Repetition × Font | Repetition × Size | |
|---|---|---|---|
| 50-100 | ns | ns | ns |
| 100-150 | * | * | ns |
| 150-200 | ** | * | ns |
| 200-250 | ** | * | ns |
| 250-300 | ** | ns | ns |
| 300-350 | ** | ns | ns |
| 350-400 | ** | ns | ns |
| 400-450 | ** | ns | ** |
| 450-500 | ** | ns | ns |
p<.05
p<.01
180-300ms post-target (N250)
The most notable effect in this window was the larger negativity to unrelated compared with repeated words: main effect of Repetition, F(1, 23)=29.54, p<.001 - see Figure 4. This replicates our prior work showing repetition priming in the N250 ERP component (Holcomb & Grainger, 2006; Kiyonaga et al., 2007). As can be seen in Figure 5 and Table 1, neither Size nor Font interacted with the effects of Repetition in this time window: Size × Repetition: F(1, 23)=0.003, p=.956; Font × Repetition: F(1, 23)=0.156, p=.696.
Figure 4.
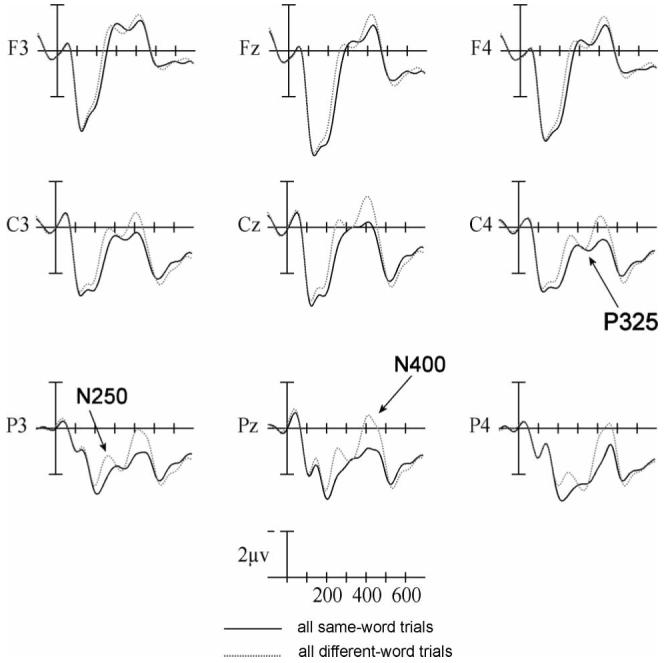
Event-related potentials (ERPs) from nine electrode sites showing the effects of Repetition Priming. The abscissa shows time (in milliseconds) from target onset.
Figure 5.
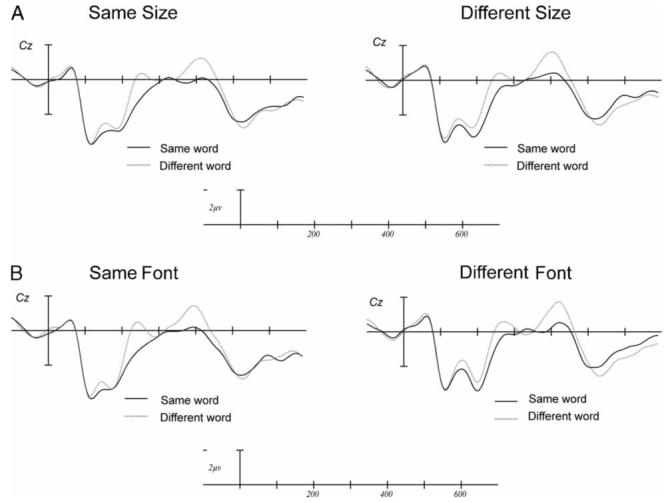
Repetition priming broken down as a function of target size (panel A) and font (panel B). The abscissa shows time (in milliseconds) from target onset.
300-375 ms post-target (P325)
In this epoch target words that were unrelated to their primes were significantly more negative than targets that were repetitions of their primes but only at central and more posterior sites, Repetition × Electrode Interaction, F(2, 46)=10.03, p=.001 - see Figure 4. Again, as can be seen in Figure 5, the interaction effects with Size and Repetition and Font and Repetition were not significant in this epoch: Size × Repetition: F(1, 23)=0.055, p=.817; Font × Repetition: F(1, 23)=0.922, p=.347.
375-550 ms post-target (N400)
In the final epoch, target words that were unrelated to their primes were significantly more negative-going than targets that were repetitions of their primes: main effect of Repetition: F(1, 23)=9.93, p=.007, especially at central and more posterior sites (Repetition × Electrode Interaction, F(2, 46)=12.79, p=0.001 - see Figure 4. As can be seen in Figure 5, neither Size nor Font interacted with Repetition priming in this time window: Size × Repetition: F(1, 23)=0.007, p=.936; Font × Repetition: F(1, 23)=0.137, p=.715.
Time-course analysis
In order to provide a more fine-grained analysis of the evolution of priming effects over time we also performed a time-course analysis on successive 50 ms epochs starting at 50 ms post-target onset and continuing through the N400 epoch. Reported in Table 1 are the main effects of REPETITION and its interaction with FONT and SIZE across nine electrode sites (F3, Fz, F4, C3, Cz, C4, P3, Pz, P4).
The time-course analysis confirms the results of the ANOVAs based on data centred on pre-identified ERP components. There are strong effects of repetition priming arising as early as 100 ms post-target onset. These repetition priming effects interact with prime-target font compatibility in a time-window extending from 100 ms to 250 ms post-target onset, and reappear at the latest epoch (450-500 ms post-target onset). There is no sign of an interaction between repetition priming and prime-target size compatibility in any of the epochs. The re-emergence of the Repetition × Font interaction at the final epoch suggests that a change in font across prime and target (Arial primes, Gigi targets) is affecting processing at the very end of the classic N400 component, possibly related to some form of explicit recognition that prime and target stimuli were the same word or not.
Behavioural data
Probe words (animal names) were correctly identified on 86% of trials when they appeared at the target location and on 23.7% of trials when they appeared at the prime location.
DISCUSSION
A masked priming experiment with ERP recordings examined the influence of changes in size and shape across prime and target on repetition priming effects. Effects of repetition priming were found in the N/P150, N250, P325, and N400 ERP components, thus replicating prior observations (Holcomb & Grainger, 2006; Kiyonaga et al., 2007). The repetition priming effects in the N250, P325, and N400 components were not affected by a change in size or a change in font across prime and target stimuli. On the other hand, a significant interaction between font and repetition was found in the N/P150 ERP component.
Is the processing reflected in the N/P150 component operating on representations that are invariant to size and font? The critical results with respect to this question are the interactions between size and repetition and between font and repetition. The former was not significant, while the latter was. These results therefore suggest that the N/P150 reflects processing at a point in time when size invariance has been achieved,3 but invariance to a change in font is not yet complete. In line with the proposal of Grainger and Holcomb (in press), this could well correspond to the process by which visual features are mapped onto abstract letter identities. These abstract letter identities would correspond to the letter representations of the alphabetic array in Grainger and van Heuven’s model. We therefore suggest that the N/P150 reflects processing occurring immediately prior to the activation of such case and font invariant letter representations. This processing would involve the mapping of size invariant letter features onto letter representations, possibly involving neurons in posterior fusiform gyrus. The work of Dehaene et al. (2004) suggests that left posterior fusiform contains neurons that code for the presence of abstract letter identities at a particular retinal location (as in the alphabetic array of Grainger & van Heuven’s (2003) model). Furthermore, the MEG study of Cornelissen, Tarkiainen, Helenius, and Salmelin (2003) suggests that such location-specific coding of letter identities in left fusiform is performed before 200 ms post-stimulus onset.
Concerning the relative influence of stimulus size and stimulus shape, there is converging evidence from neurophysiological studies with monkeys and fMRI studies with humans that suggests that size invariance is achieved before shape invariance. Human studies using fMRI (Grill-Spector et al., 1999; Sawamura, Georgieva, Vogels, Vanduffel, & Orban, 2005) show a large amount of size invariance in the most anterior regions of lateral occipital complex (LOC) that is already present to a lesser extent in the more posterior regions. These results fit with the results of monkey studies (Logothetis, Pauls, & Poggio, 1995; Sawamura et al., 2005) showing size invariance in monkey anterior inferotemporal cortex (IT, thought to be the monkey homologue of human LOC). Logothetis et al. (1995) found IT neurons that responded to the same view of an object presented at different sizes, but not to different views of the same object. Furthermore, there is recent evidence that the most anterior region of human LOC does not show priming across different exemplars of the same object, such as an upright piano and a grand piano (Hayworth & Biederman, 2006). Most important, Hayworth and Biederman also failed to find priming when the same object was presented with non-overlapping subsets of its component parts. These authors therefore concluded that neurons in anterior LOC encode for parts of an object. This might also be the region that codes for letter features, prior to the mapping of such features onto letter representations possibly located in posterior fusiform (Dehaene et al., 2004). Orthographic processing would then proceed in a posterior-anterior sweep through left fusiform, known to be sensitive to orthographic structure (e.g., Binder, Medler, Westbury, Liebenthal, & Buchanan, 2006; Cohen et al., 2000; Dehaene et al., 2005). Also, the fact that an N/P150 component is found in repetition priming studies with picture stimuli (Eddy et al., 2006), suggests that it reflects a general process of mapping visual features onto higher-level representations (object parts) during visual object recognition.
The second important finding of the present study is that the effects of repetition priming on N250, P325, and N400 amplitude, were not significantly modulated by our size and font manipulation. This confirms that the N250 (and a fortiori, the P325 and N400) reflects processing that occurs once size and shape invariance has been achieved. If this were not the case, we ought to have observed stronger repetition priming effects with primes and targets in the same size and/or same font compared with the condition where there was a change in size or font across prime and target. Therefore, we have further evidence that the N250 ERP component observed in our prior work on masked priming and visual word recognition, reflects processing involving orthographic representations that are insensitive to changes in size and shape. It is also possible that part of the activity reflected in the N250 ERP component is generated by prelexical phonological representations. Grainger et al. (2006) showed that pseudohomophone primes affected N250 amplitude compared with orthographic control primes. However, they also demonstrated orthographic priming (from transposed-letter primes) in the N250, and this orthographic priming effect had a slightly earlier onset and a more posterior distribution than the phonological priming effect. This suggests that the N250 ERP component reflects prelexical orthographic and phonological processing. Furthermore, given the effects of transposed-letter primes in the Grainger et al. study, it would appear that the type of orthographic processing reflected in the N250 codes for within-word relative-position, rather than a location-specific coding of letter identities.
Therefore, in terms of Grainger and van Heuven’s (2003) model of orthographic processing, the results obtained so far fit with the proposal that the N/P150 ERP component reflects the mapping of feature information onto location-specific, abstract letter representations (the alphabetic array), whereas the N250 reflects processing performed by location-invariant orthographic and phonological representations (see Grainger & Holcomb, in press, for a detailed discussion of this proposal). However, these estimates of the time-course of visual word recognition would appear to conflict with other ERP studies showing effects of letter cluster (Ngram) frequency as early as 100 ms post-stimulus onset (Hauk et al., 2006), effects of word frequency and lexicality as early as 150 ms post-stimulus onset (Hauk & Pulvermüller, 2004; Sereno, Rayner, & Posner, 1998), and semantic effects as early as 200 ms post-stimulus onset (Dell’Acqua, Pesciarelli, Jolicoeur, Eimer, & Peressotti, 2007; Morris, Frank, Grainger, & Holcomb, 2007). More in line with our estimates, on the other hand, are the studies of Bentin et al. (1999), Tarkiainen, Helenius, Hansen, Cornelissen, and Salmelin (1999), and Schendan, Giorgio, and Kutas (1998) showing differences between orthographic and non-orthographic stimuli (symbols) emerging at around 150 ms post-stimulus onset. We would argue that such apparent discrepancies might be due to the cascaded, interactive nature of lexical processing (McClelland & Rumelhart, 1981). The components described in our work arguably reflect the peak of neural activity associated with a given type of processing, but earlier effects of the same type of processing could result from feedback from the fastest of the feedforward processes affecting the bulk of feedforward processing (Morris et al., 2007). Such feedback processes might well be influenced by attentional factors and task demands, hence giving rise to some of the above-mentioned discrepancies.
Finally, in future work it will be important to compare effects of stimulus font with effects of case. As noted in the Introduction, although it is reasonable to assume the existence of case-specific letter representations, at least for letters with highly distinct upper- and lower-case versions (e.g., a-A, see Petit et al., 2006), it seems unreasonable to postulate the existence of font-specific representations. Changes in font could be handled by a font tuning mechanism (Gauthier, Wong, Hayward, & Cheung, 2006; Sanocki, 1988) that would work for non-arbitrary changes in letter shape. Such a font tuning mechanism would operate on case-specific letter representations, thus allowing various forms of lower-case ‘a’ (a, a) to be mapped onto a canonical lowercase ‘a’, and idem for uppercase ‘A’. We would therefore expect to see an interaction between repetition priming and stimulus case in the ERP signal arriving later than the interaction with stimulus font found in the present study - sometime between the fall of the N/P150 and the rise of the N250. This would correspond to the effect of case-specific letter representations found between 180 and 220 ms post-target onset for isolated letters in the Petit et al. (2006) study.
In conclusion, the present study manipulated the size and font of target words in a masked repetition priming study with ERP recordings. Repetition priming effects were found in a cascade of ERP components starting at 150 ms post-target onset (N/P150) and up to the N400 ERP component. Stimulus font, but not stimulus size, interacted with the effects of repetition priming in the N/P150 component. This was taken as evidence that the N/P150 reflects the mapping of size invariant feature representations onto abstract, shape-invariant, letter representations. The higher-level orthographic processing that is performed once this mapping is complete is no longer sensitive to changes in stimulus font across prime and target.
Acknowledgments
This research was supported by grants HD25889 and HD043251.
Thanks to Irving Biederman for helpful discussion of size and shape invariance in visual object recognition, and to two anonymous reviewers for helpful comments on an earlier version of this work.
Footnotes
Here we use the term ‘shape invariance’, applied to the specific case of perceiving printed words, to refer to the ability to recognise words presented in different formats (different fonts and/or different case). Vision scientists typically use the term to refer to our ability to perceive objects from different viewpoints, with different orientations and different sizes.
Although there is a reasonable case to be made in favour of case-specific letter representations, at least for visually dissimilar pairs of upper- and lower-case letters (e.g., a-A), it would not appear reasonable to assume the existence of font-specific letter representations. Difficulties in processing an unusual font could be achieved by a tuning process that adjusts the recognition process in order to optimise processing of atypical stimuli (e.g., Gauthier, Wong, Hayward, & Cheung, 2006; Sanocki, 1988).
We do acknowledge that our size manipulation may not have been severe enough to observe an interaction with repetition priming in the present study. Majaj, Pelli, Kurshan, and Palomares (2002) have shown that the critical spatial frequency channel for letter identification varies with more extreme manipulations of letter size. Future research should therefore involve a parametric manipulation of size differences across prime and target stimuli with more extreme end points.
Contributor Information
Krysta Chauncey, Tufts University, Melford, MA, USA.
Phillip J. Holcomb, Tufts University, Melford, MA, USA
Jonathan Grainger, Centre National de la Recherche Scientifique (CNRS), and University of Provence, Aix-Marseille, France.
REFERENCES
- Bentin S, Mouchetant-Rostaing Y, Giard MH, Echallier JF, Pernier J. ERP manifestations of processing printed words at different psycholinguistic levels: Time course and scalp distribution. Journal of Cognitive Neuroscience. 1999;11:235–260. doi: 10.1162/089892999563373. [DOI] [PubMed] [Google Scholar]
- Binder JR, Medler DA, Westbury CF, Liebenthal E, Buchanan L. Tuning of the human left fusiform gyrus to sublexical orthographic structure. NeuroImage. 2006;33:739–748. doi: 10.1016/j.neuroimage.2006.06.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caramazza A, Hillis AE. Spatial representation of words in the brain implied by studies of a unilateral neglect patient. Nature. 1990;346:267–269. doi: 10.1038/346267a0. [DOI] [PubMed] [Google Scholar]
- Cohen L, Dehaene S, Naccache L, Lehéricy S, Dehaene-Lambertz G, Hénaff MA, Michel F. The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain. 2000;123:291–307. doi: 10.1093/brain/123.2.291. [DOI] [PubMed] [Google Scholar]
- Cornelissen P, Tarkiainen A, Helenius P, Salmelin R. Cortical effects of shifting letter position in letter strings of varying length. Journal of Cognitive Neuroscience. 2003;15:731–746. doi: 10.1162/089892903322307447. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Jobert A, Naccache L, Ciuciu P, Poline J, Le Bihan D, Cohen L. Letter binding and invariant recognition of masked words: behavioral and neuroimaging evidence. Psychological Science. 2004;15:307–313. doi: 10.1111/j.0956-7976.2004.00674.x. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Cohen L, Sigman M, Vinckier F. The neural code for written words: a proposal. Trends in Cognitive Sciences. 2005;9:335–341. doi: 10.1016/j.tics.2005.05.004. [DOI] [PubMed] [Google Scholar]
- Dell’Acqua R, Pesciarelli F, Jolicoeur P, Eimer M, Peressotti F. The interdependence of spatial attention and lexical access as revealed by early asymmetries in occipito-parietal ERP activity. Psychophysiology. 2007;44:436–443. doi: 10.1111/j.1469-8986.2007.00514.x. [DOI] [PubMed] [Google Scholar]
- Eddy M, Schmid A, Holcomb PJ. A new approach to tracking the time-course of object perception: Masked repetition priming and event-related brain potentials. Psychophysiology. 2006;43:564–568. doi: 10.1111/j.1469-8986.2006.00455.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gauthier I, Wong AC-N, Hayward WG, Cheung OS. Font tuning. associated with expertise in letter perception. Perception. 2006;35:541–559. doi: 10.1068/p5313. [DOI] [PubMed] [Google Scholar]
- Geisser S, Greenhouse S. On methods in the analysis of profile data. Psychometrica. 1959;24:95–112. [Google Scholar]
- Grainger J, Holcomb PJ. Neural constraints on a functional architecture for word recognition. In: Cornelissen P, Hansen P, Kringelbach M, Pugh K, editors. The neural basis of reading. Oxford University Press; Oxford: in press. [Google Scholar]
- Grainger J, Kiyonaga K, Holcomb PJ. The time-course of orthographic and phonological code activation. Psychological Science. 2006;17:1021–1026. doi: 10.1111/j.1467-9280.2006.01821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J, Van Heuven W. Modeling letter position coding in printed word perception. In: Bonin P, editor. The mental lexicon. Nova Science Publishers; New York: 2003. pp. 1–24. [Google Scholar]
- Grill-Spector K, Kushnir T, Edelman S, Avidan G, Itzchak Y, Malach R. Differential processing of objects under various viewing conditions in the human lateral occipital complex. Neuron. 1999;24:187–203. doi: 10.1016/s0896-6273(00)80832-6. [DOI] [PubMed] [Google Scholar]
- Hauk O, Patterson K, Woollams A, Watling L, Pulvermüller F, Rogers TT. Q: When would you prefer a SOSSAGE to a SAUSAGE? A: At about 100 msec. ERP correlates of orthographic typicality and lexicality in written word recognition. Journal of Cognitive Neuroscience. 2006;18:818–832. doi: 10.1162/jocn.2006.18.5.818. [DOI] [PubMed] [Google Scholar]
- Hauk O, Pulvermüller F. Effects of word length and frequency on the human event-related potential. Clinical Neurophysiology. 2004;115:1090–1103. doi: 10.1016/j.clinph.2003.12.020. [DOI] [PubMed] [Google Scholar]
- Hayworth K, Biederman I. Neural evidence for intermediate representations in object recognition. Vision Research. 2006;46:4024–4031. doi: 10.1016/j.visres.2006.07.015. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. On the time course of visual word recognition: An event-related potential investigation using masked repetition priming. Journal of Cognitive Neuroscience. 2006;18:1631–1643. doi: 10.1162/jocn.2006.18.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiyonaga K, Midgley KJ, Holcomb PJ, Grainger J. Masked cross-modal repetition priming: An ERP investigation. Language and Cognitive Processes. 2007;22:337–376. doi: 10.1080/01690960600652471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logothetis NK, Pauls J, Poggio T. Shape representation in the inferior temporal cortex of monkeys. Current Opinion in Biology. 1995;5:552–563. doi: 10.1016/s0960-9822(95)00108-4. [DOI] [PubMed] [Google Scholar]
- Majaj NJ, Pelli DG, Kurshan P, Palomares M. The role of spatial frequency channels in letter identification. Vision Research. 2002;42:1165–1184. doi: 10.1016/s0042-6989(02)00045-7. [DOI] [PubMed] [Google Scholar]
- McClelland JL, Rumelhart DE. An interactive activation model of context effects in letter perception: Part 1, An account of basic findings. Psychological Review. 1981;88:375–407. [PubMed] [Google Scholar]
- Morris J, Frank T, Grainger J, Holcomb PJ. Semantic transparency and masked morphological priming: An ERP investigation. Psychophysiology. 2007;44:506–521. doi: 10.1111/j.1469-8986.2007.00538.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petit JP, Midgley KJ, Holcomb PJ, Grainger J. On the time-course of letter perception: A masked priming ERP investigation. Psychonomic Bulletin and Review. 2006;13:674–681. doi: 10.3758/bf03193980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanocki T. Font regularity constraints on letter recognition. Journal of Experimental Psychology: Human Perception and Performance. 1988;14:472–480. doi: 10.1037//0096-1523.14.3.472. [DOI] [PubMed] [Google Scholar]
- Sawamura H, Georgieva S, Vogels R, Vanduffel W, Orban GA. Using functional magnetic resonance imaging to assess adaptation and size invariance of shape processing by humans and monkeys. Journal of Neuroscience. 2005;25:4294–4306. doi: 10.1523/JNEUROSCI.0377-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schendan HE, Giorgio G, Kutas M. Neurophysiological evidence for visual perceptual categorization of words and faces within 150ms. Psychophysiology. 1998;35:240–251. [PubMed] [Google Scholar]
- Sereno SC, Rayner K, Posner MI. Establishing a time-line of word recognition: evidence from eye movements and event-related potentials. NeuroReport. 1998;9:2195–2200. doi: 10.1097/00001756-199807130-00009. [DOI] [PubMed] [Google Scholar]
- Tarkiainen A, Helenius P, Hansen PC, Cornelissen PL, Salmelin R. Dynamics of letter string perception in the human occipito-temporal cortex. Brain. 1999;122:2119–2132. doi: 10.1093/brain/122.11.2119. [DOI] [PubMed] [Google Scholar]
- Whitney C. How the brain codes the order of letters in a printed word: The SERIOL model and selective literature review. Psychonomic Bulletin and Review. 2001;8:221–243. doi: 10.3758/bf03196158. [DOI] [PubMed] [Google Scholar]