

Abstract
The present study used event-related potentials to examine the time-course of relative-position and absolute-position orthographic priming. Relative-position priming was examined using primes formed by a concatenated subset of the target word’s letters (e.g., cllet/COLLECT vs. dlema/COLLECT), and absolute-position priming was investigated using hyphenated versions of these primes (c-lle-t/COLLECT vs. d-lem-a/COLLECT). Both manipulations modulated the ERP waveform starting at around 100 ms post-target onset and extending into the N400 component. The first clear manifestation of priming was found in the N250 component, where hyphenated primes were found to have an earlier, more robust and more widely distributed effect than the concatenated primes. On the other hand, both prime types had similar effects on N400 amplitude. These results provide important information about the time-course of activation of location-specific and location-invariant (word-centered) orthographic representations during visual word recognition.
Keywords: orthographic processing, visual word recognition, ERPs
1. Introduction
There is a growing body of research on orthographic processing during visual word recognition that suggests that some form of approximate relative-position code for letter positions is computed and used to access whole-word orthographic representations and semantics. One key finding in this area is that masked orthographic priming effects obtained when primes and targets share letters are also observed when the shared letters do not respect their absolute, length-dependent, position in prime and target. For example, the prime SLENE shares letters with the target SILENCE, but only the relative position of these letters is respected, not their absolute position in the string.1 Several behavioral studies have found significant priming from such stimuli (Humphreys, Evett, & Quinlan, 1990; Peressotti & Grainger, 1999; Grainger, Granier, Farioli, Van Assche, & van Heuven, 2006a).
Grainger et al. (2006a) referred to this empirical finding as the relative-position priming constraint, and following Grainger and van Heuven (2003) suggested that it was one of the single most constraining pieces of evidence for models of orthographic processing. This and related findings, such as transposed-letter effects (e.g., Perea & Lupker, 2004; Schoonbaert & Grainger, 2004), have been instrumental in the development of a new class of models of orthographic processing such as the SOLAR model (Davis, 1999), the SERIOL model (Whitney, 2001; 2008) and the overlap model (Gomez, Ratcliff, & Perea, 2008; see Grainger, 2008, for review). The principal feature of these new approaches to letter position coding is that there is a certain amount of flexibility in the way in which letter identities are associated with a given position in the letter string. In Grainger and van Heuven’s (2003) model of orthographic processing this is done by coding for contiguous and non-contiguous ordered pairs of letters (open-bigrams) independently of their precise location in the string (see also Grainger & Whitney, 2004; Whitney, 2001; Whitney, 2008).
One important aspect of the behavioral evidence for relative-position priming using subset primes is the fact that adding absolute length-dependent position information by replacing the removed letters by hyphens does not increase the size of priming effects. Peressotti and Grainger (1999) and Grainger et al. (2006a) found statistically equivalent priming from concatenated (slene-silence) and hyphenated (s-len-e) subset primes. This suggests that not only can one obtain robust priming from relative-position subset primes, but that providing absolute position information (in the form of hyphens) does not add anything. Open-bigram coding can account for this result simply by assuming that the hyphens are ignored. This is not, however, a very satisfactory assumption, since hyphens likely lead to partial activation of letter representations and should therefore influence orthographic processing at some point. According to Grainger and van Heuven’s (2003) model, this influence should mainly arise at the location-specific map of letter identities postulated in their model (the alphabetic array, see Figure 1), before access to a location-invariant orthographic code.
Figure 1.
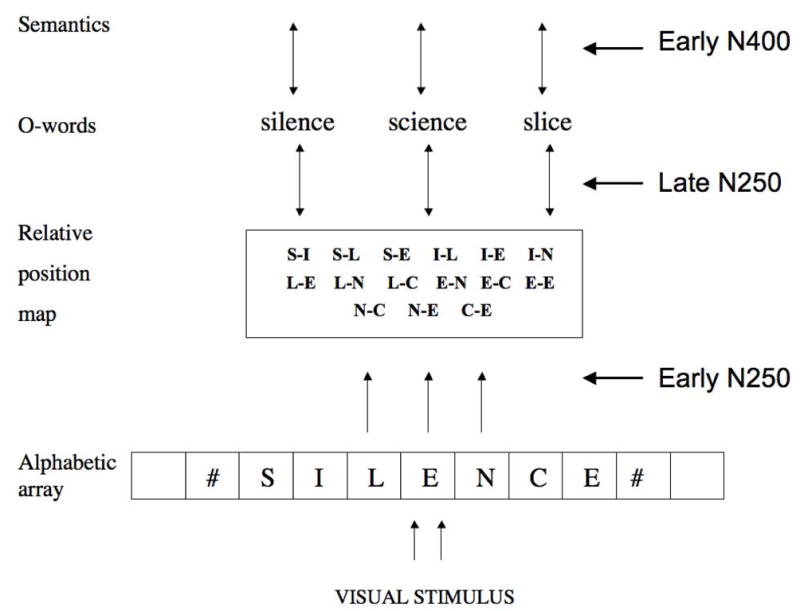
Linking ERP components to orthographic processing within the framework of Grainger and van Heuven’s (2003) model. A bank of parallel, independent, location-specific letter detectors (alphabetic array) send activation onto a location-invariant, word-centered, prelexical orthographic code (relative-position map), and from there onto whole-word orthographic representations. This corresponds to only part of the larger framework for word recognition described in the bi-modal interactive-activation model (see, Grainger & Holcomb, 2009; Holcomb & Grainger, 2007).
Given the potential theoretical impact of the behavioral results of Peressotti and Grainger (1999) and Grainger et al. (2006a), the present study aims to provide further data on relative-position priming effects from concatenated and hyphenated primes. More specifically, in the present study we combine the masked priming technique with the recording of event-related brain potentials (ERPs) in order to provide critical information about the time-course of location-specific and location-independent orthographic coding. Recent research applying this approach (e.g., Holcomb & Grainger, 2006; Kiyonaga, Grainger, Midgley, & Holcomb, 2007), has found that manipulations of prime-target similarity affect a cascade of ERP components starting as early as 130 ms post-target-onset and continuing through to as late as 600 ms. These recent ERP findings obtained with the masked priming paradigm offer an exciting new perspective for plotting the time-course of the component processes involved in visual word recognition.
Grainger and Holcomb (2007; 2009; Holcomb & Grainger, 2007) provided a tentative mapping of ERP components onto a functional architecture for word recognition. The N/P150 (peaking at around 150 ms post-target onset) was associated with the mapping of visual features onto letter representations involved in word recognition (Chauncey, Holcomb, & Grainger, 2008; Dufau, Grainger, & Holcomb, 2008). Other studies have shown that prime-target visual overlap also generates an N/P150 effect when targets are isolated letters or pictures of objects (Eddy, Schmid, & Holcomb, 2006; Petit, Midgley, Holcomb, & Grainger, 2006), hence suggesting that this component is sensitive to prime-target visual overlap independently of the type of stimuli. The N250 (peaking at around 250 ms post-target onset) was associated with the mapping of abstract location-specific letter identities onto higher-level location-invariant orthographic and phonological representations (Grainger, Kiyonaga, & Holcomb, 2006). This is therefore considered as the first component to be sensitive to processing specific to printed words. Following the N250, the N400 component would reflect the mapping of whole-word form representations (orthographic and phonological) onto higher-level semantic representations (Holcomb, Grainger, & O’Rourke, 2002; Holcomb & Grainger, 2006; 2007). Given the focus of the present experiment on the transition from location-specific to location-invariant orthographic representations, in what follows we introduce a division of the N250 component into an early and late phase that are expected to be differentially sensitive to location specificity.
Figure 1 summarizes our attempt to link the different ERP components revealed in our masked priming experiments with component processes of visual word recognition. Here the emphasis is on visual-orthographic processing, hence we do not show the phonological pathway that is part of the complete model (see Grainger & Holcomb, 2009, for a recent overview). Furthermore, in the present analysis we focus on the ERP components that we expect to be affected by our priming manipulations – the N250 and N400. Given the relatively low-level of orthographic overlap across primes and targets in the present study, plus the change in case between prime and target stimuli that further reduced purely visual overlap, we did not expect to see an N/P150 priming effect. According to the model presented in Figure 1, the first phase of processing involving abstract (shape-invariant) letter representations is the mapping of location-specific letter identities in the alphabetic array onto location-invariant, word-centered orthographic representations. This processing is associated with the early phase of the N250 ERP component. Location-invariant prelexical orthographic representations are then mapped onto whole-word orthographic representations. This phase of processing is associated with the later part of the N250. Therefore, according to the above analysis we expect to see stronger priming effects for hyphenated primes compared with relative-position primes in the early phase of the N250 component which should dissipate in the later phase of the N250 when the shift to a location-invariant orthographic code is complete.
In order to put this account to test, the present experiment manipulates the positional overlap of letters shared by prime and target stimuli in a masked priming experiment with ERP recordings. Orthographically related prime stimuli were formed of the 1st, 3rd, 4th, 5th, and 7th letters of 7-letter word targets (i.e., subset primes), and the prime letters could either be concatenated to form a 5-letter string (13457) or were presented with hyphen marks replacing the 2nd and 6th letters of the target (1-345-7). Unrelated primes were related primes from the same category (concatenated or hyphenated) associated with a different target. We expect to see stronger priming effects in the hyphenated prime condition than the concatenated prime condition in the early phase of the N250, reflecting the influence of location-specific letter representations in this time window. This advantage for hyphenated primes should diminish as location-invariant orthographic representations gradually take over and dominate processing in the later phase of the N250 and the subsequent N400.
2. Results
Plotted in Figure 2 are the waveforms for each experimental condition at the nine electrode sites used in the statistical analyses (see Methods). Plotted in Figure 3 are the voltage maps formed (using EEGlab) from all 29 scalp sites contrasting the two types of priming effects.
Figure 2.
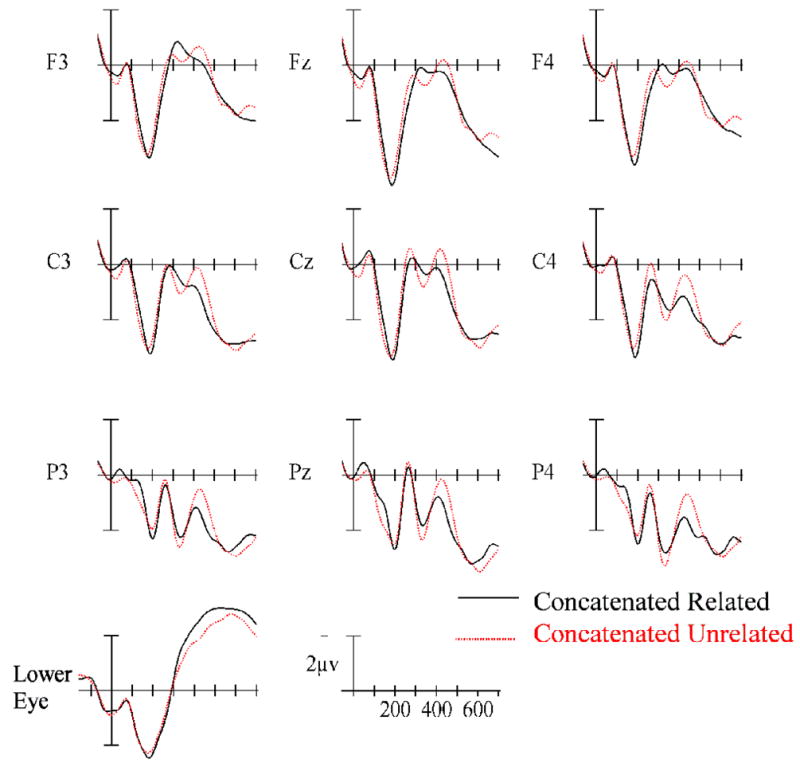
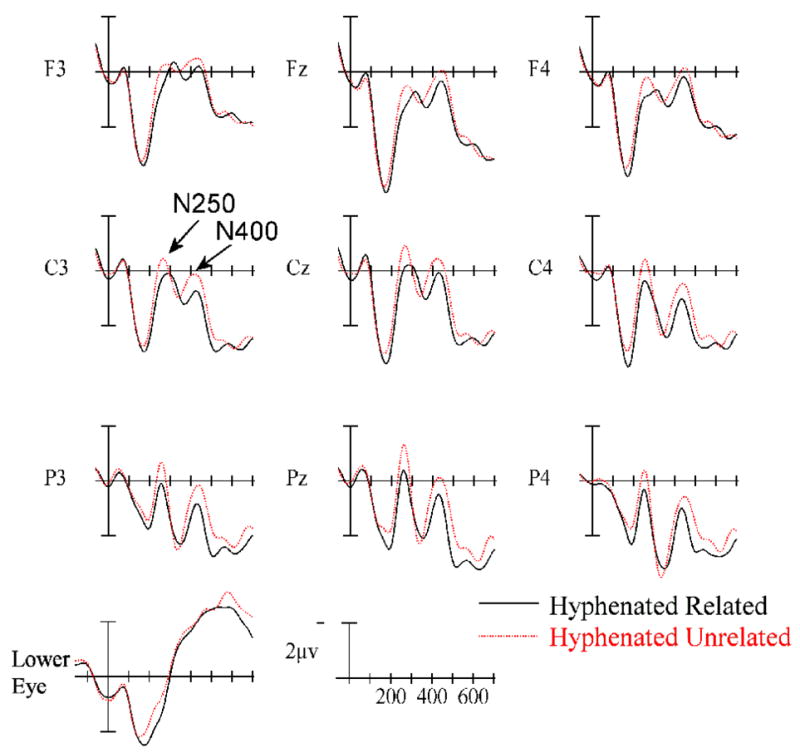
ERPs time locked to target onset in the concatenated (top) and hyphenated (bottom) priming conditions over-plotted with their respective unrelated control conditions. Target onset is marked by the vertical calibration bar and each tic mark represents 100 ms. Negative values plotted up.
Figure 3.

Voltage maps calculated by subtracting target ERPs from the concatenated related condition from their control ERPs (top) and the hyphenated related ERPs from their controls (bottom).
175–240 ms Target Epoch (early N250)
The ANOVA in this epoch revealed a main effect of Relatedness (F(1,24) = 4.36, p = .048), as well as a higher order interaction between Relatedness, Prime Type and the two scalp distribution variables (Relatedness × Prime Type × AntPost × Laterality: F(4,96) = 3.24, p = .027). To better understand this interaction, ANOVAs were run separately on the two Prime Type conditions. For the concatenated primes (e.g., cllet/COLLECT) there was no main effect of Relatedness (p < .2) and only a trend for unrelated targets to be more negative than related targets at central and anterior sites (marginal Relatedness × AntPos interaction: F(2,48) = 2.95, p = .086). However, for the hyphenated primes (e.g., c-lle-t/COLLECT) there was a clear main effect of Relatedness (F(1,24) = 5.17, p = .032 – see Figure 3b at 190 ms) indicating a more robust and widely distributed effect of priming compared with the concatenated primes.
240–305 ms Target Epoch (late N250)
As can be seen in Figures 3 and 4, between 240 and 305 ms ERPs to targets following related primes were more positive-going than to targets preceded by unrelated primes. The ANOVA revealed a main effect of Relatedness: F(1,24) = 4.77, p = .039), and no indication that the effects of Relatedness differed as a function of Prime Type (all interactions involving Prime Type × Relatedness, Fs < 2.0).
Figure 4.
Electrode montage. The nine sites used for ANOVA are interconnected with grey lines.
375–550 ms Target Epoch (N400)
Examination of Figures 3 and 4 reveals once again that ERPs to targets following related primes were more positive-going than to targets preceded by unrelated primes (main effect of Relatedness: F(1,24) = 7.26, p = .013). These priming effects were more pronounced at posterior scalp sites (Relatedness × AntPost: F(2,48) = 4.58, p = .037 – see Figure 3), with significant effects of Relatedness at electrode sites P3, Pz, P4, (F(1,24) = 15.4, p = .0006), and non-significant effects at electrode sites F3, Fz, F4, (F=1.73). As in the previous epoch, there was no indication of a difference in the Relatedness factor for the two Prime Types (all interactions involving Prime Type × Relatedness, Fs < 1).
Time-course Analysis
To better quantify the time-course of Priming effects, we also compared the repeated and unrelated ERPs in successive 50 ms intervals for the concatenated and hyphenated targets at three midline sites Fz, Cz and Pz) sites starting at 150 ms and continuing through 550 ms post-target onset. The results of these analyses are reported in Table 1.
Table 1.
Time-course of the priming effects at electrodes Fz, Cz and Pz.
| 150–200ms | 200–250ms | 250–300ms | 300–350ms | 350–400ms | 400–450ms | 450–500ms | 500–550ms | |
|---|---|---|---|---|---|---|---|---|
| Concatenated-Fz | - | ~ | * | - | - | - | - | - |
| Concatenated-Cz | - | - | - | - | - | * | * | - |
| Concatenated-Pz | - | - | - | - | - | ~ | ~ | - |
| Hyphenated-Fz | - | * | ~ | - | - | - | ~ | - |
| Hyphenated-Cz | - | ** | ~ | - | ** | ~ | * | - |
| Hyphenated-Pz | - | ** | * | - | * | ~ | ** | * |
- = NS,
< 0.10,
< 0.05,
< 0.01
Behavioral Data
Participants detected 94% (SD = .08) of animal probes in the target position and 8% (SD = .15) in the prime position within a window of 200 to 1200ms post-probe onset.
3. Discussion
The present study examined relative-position priming effects in a masked priming paradigm combined with ERP recordings. Effects of concatenated subset primes (e.g., dlema-dilemma) were compared with effects of hyphenated subset primes (e.g., d-lem-a) both measured relative to appropriate unrelated control primes. Significant priming effects were found in two ERP components highlighted in our prior work on masked repetition priming–the N250 and the N400 (Holcomb & Grainger, 2006; 2007; Kiyonaga et al., 2007).
Priming effects on the N250 component were examined separately for early (175–240 ms) and late (240–305 ms) phases of this component. The early N250 was found to be sensitive to priming from hyphenated primes but not concatenated primes, while the late N250 showed priming effects that were independent of the type of priming (see Table 1 and Figures 2 & 3).2 Priming effects were also present in the N400 component, and did not differ as a function of the type of priming. Indeed, the N400 scalp maps revealed a widely distributed priming effect with a posterior focus for both types of prime. This is in line with the pattern found in prior behavioral research (Peressotti & Grainger, 1999; Grainger, Granier, et al., 2006) where statistically equivalent priming effects were found for concatenated and hyphenated primes.
On the basis of our prior work combining masked priming and ERP recordings, interpreted within the general framework of Grainger and van Heuven’s (2003) model of orthographic processing (see Figure 1), we expected to find evidence for a shift from a location-specific orthographic code to a word-centered, location-invariant orthographic code sometime between about 150 and 250 ms post-target onset. The observed advantage for hyphenated primes, starting around 200 ms post-target onset, is in line with this prediction. Hyphenated primes had a greater impact on the early phase of the N250 component compared with concatenated primes. This interaction between priming and type of prime was no longer significant in the later phase of the N250 and the N400. These results suggests that location-specific letter identities are influencing the processing that is reflected in the early phase of the N250 (200 – 250 ms post-target onset), and that the transition to a location-invariant orthographic code is already achieved by about 250 ms post-target onset. This is in line with the estimate from the MEG study of Cornelissen, Tarkiainen, Helenius, and Salmelin (2003) who found sensitivity to shifts of letter position before 200 ms post-stimulus onset.
In the introduction we presented a tentative linkage of ERP components to the different phases of orthographic processing that occur during visual word recognition (Figure 1). Rather than associating ERP components with a particular level of representation (i.e., location-specific vs. location-invariant), in Figure 1 we linked ERP components to the processing involved in mapping from one type of representation to another. The early phase of the N250 is associated with the processes involved in mapping from location-specific letter detectors to a location-invariant prelexical orthographic code. Processing in this component is still influenced by absolute position information (i.e., location along the horizontal meridian), hence the advantage for hyphenated primes compared with concatenated primes found in the present study. The later phase of the N250 is associated with the mapping of a location-invariant prelexical orthographic code on to whole-word orthographic representations. At this level of processing, the shift to a location-invariant code is complete, hence the statistically equivalent priming found for hyphenated and concatenated primes. Finally, the early phase of the N400 is associated with the mapping of whole-word orthographic representations onto semantics.
It is likely that the shift to a location-independent, word-centered orthographic code coincides temporally with the computation of a location-independent, word-centered prelexical phonological code (see Grainger & Holcomb, 2009, for a more detailed description of these processes). The timing of this shift as estimated from the pattern of priming effects in the present study, is in agreement with the estimated onset of phonological priming effects in Grainger et al. (2006b). In that study, an orthographic priming manipulation (transposed-letter primes – e.g., talbe-table vs. tarpe-table) was already present in the early phase of the N250, whereas a phonological priming manipulation (pseudohomophone primes – e.g., bakon-bacon vs. bafon-bacon) only emerged in the later part of the N250. It should be noted that there is evidence from behavioral studies that priming manipulations using subset relative-position primes and transposed-letter primes principally affect orthographic processing (e.g., Grainger et al., 2006a; Perea & Carreiras, 2006). Other recent ERP studies further corroborate this time-course pattern of early orthographic effects (in the early phase of the N250) following by phonological priming (Carreiras, Vergara, & Perea, 2009; Carreiras, Vergara, Perea, & Pollatsek, 2009). Given this evidence, we think that it is unlikely that the stronger priming effect found for hyphenated primes compared with concatenated primes in the early phase of the N250 is due to improved conversion of orthography to phonology with these primes.
A number of issues remain, however, concerning possible interpretations of priming effects in the N250 ERP component. In the present study we have argued that the early phase of the N250 is still sensitive to location-specific orthographic coding (see Figure 1). At the same time there is evidence for effects of transposed-letter priming in this time window (Carreiras et al., 2009a; Grainger et al., 2006b), and according to Grainger and van Heuven’s (2003) model, such priming effects should reflect processing at the level of location-invariant orthographic representations. The way out of this apparent contradiction is to interpret the N250 as reflecting the progressive mapping of location-specific orthographic representations onto a location-invariant orthographic code in a cascaded interactive processing network. The precise timing of priming effects will therefore depend on the strength of the priming manipulation. In this respect, the behavioral evidence at present shows that transposed-letter primes present a stronger priming manipulation than the subset primes used in the present study (see Grainger, 2008, for review). This would also explain why repetition priming (the strongest possible priming manipulation) produced effects in the N250 that were not sensitive to shifts in prime location (Dufau et al., 2008). Finally, the hypothesized cascaded interactive nature of processing during visual word recognition allows us to capture lexical and semantic influences on N250 priming effects (Morris, Franck, Grainger, & Holcomb, 2007; Dunabeitia, Molinaro, Laka, Estevez, & Carreiras, 2009). While the bulk of processing involved in activating a location-invariant orthographic code is still in progress, top-down feedback from the fastest feedforward processes could modulate on-going feedforward processing.
In conclusion, the present study has provided a further demonstration of sensitivity to location-specificity in the early phases of orthographic processing that gradually dissipates as processing continues to higher-level lexical and semantic levels. The timing estimate of the location-specific influence provided by ERPs locates the effect at the level of orthographic processing rather than at the feature level. This is therefore further evidence in favor of the location-specific letter detectors postulated in Grainger and van Heuven’s (2003) model. The word-centered location-invariant code of this model also provides a natural account of why there is no longer a difference in the size of priming effects produced by concatenated primes and hyphenated primes at later stages of processing.
4. Experimental Procedure
Participants
Twenty-five volunteers from Tufts University (14 females and 11 males, mean age = 19.3, SD = 1.4) were paid $25 to participate in this experiment. All participants reported being right-handed native speakers of English with normal or corrected to normal visual acuity.
Stimuli
The critical stimuli for this experiment were formed from the 400 7-letter words (targets) and their respective 5-letter subset primes. Word frequency was on average 49 per million and ranged between 7 and 515 per million (Kuçera & Francis, 1967), and words were nouns, adjectives, and verbs in infinitive form. Stimulus lists consisted of 500 trials each containing a prime-target pair of items, with the prime presented in lower case letters and the target in upper case. In each of four lists, there were 100 concatenated prime trials in which the second and sixth letter of the target word was removed and the remaining five letters were juxtaposed (e.g. cllet/COLLECT), 100 unrelated concatenated trials in which a concatenated prime was formed from a different target word (e.g. dlema/COLLECT, where “dlema” is formed from DILEMMA), 100 hyphenated trials in which the removed letters from primes were replaced with hyphens in order to preserve the relative spacing between letters (e.g. c-lle-t/COLLECT) and 100 unrelated hyphenated trials (e.g. d-lem-a/COLLECT). The same set of target words was tested in all four priming conditions across lists and participants, but in each list (and participant) each item was presented only once. There were also 100 non-critical trials randomly mixed in with the above 400 critical trials. Fifty contained animal names in the prime position and a filler word in the target position, while another 50 trials contained an animal word in the target position and an unrelated concatenated or hyphenated item (50% each) in the prime position. The animal names served as probe items in a go/no-go semantic categorization task in which participants were instructed to press a single button as soon as they detected an animal name. Participants were told to read all other words passively (i.e., critical stimuli did not require an overt response). Having animal names in the prime position provided a measure of prime awareness during the course of the experiment. A practice session was administered before the main experiment to familiarize the participant with the procedure. The practice session included 22 trials of the same structure as the main experiment with the same proportion of animal names to which participants responded.
Procedure
Visual stimuli were presented on a 19″ monitor set to a refresh rate of 100 Hz (which allows 10 ms resolution of stimulus control) and located 143 cm in front of the participant. Stimuli were displayed at high contrast as white letters on a black background in the Arial font (letter matrix 32 pixels tall × 16 pixels wide). Each trial began with the termination of a fixation stimulus in the middle of the screen. Five hundred milliseconds later, a forward mask of nine hash marks (#########) was presented for a duration of 500ms. The forward mask was replaced at the same location on the screen by the lower case prime stimulus for 50ms. The prime was immediately replaced by a backward mask of hash marks. The backward mask remained on the screen for one retrace interval (10 ms) and was immediately replaced by the visual target in upper case letters for a duration of 500 ms. All target words were followed by a 1000ms black screen which was replaced by a 2000ms fixation stimulus marking the end of the trial. Subjects were asked to blink and move their eyes only when the fixation stimulus appeared on the screen. The next trial followed a 500 ms blank screen warning interval.
EEG recording procedure
Participants were seated in a comfortable chair in a sound attenuated darkened room. The electroencephalogram (EEG) was recorded from 29 active tin electrodes held in place on the scalp by an elastic cap (Electrode-Cap International – see Figure 4). In addition to the 29 scalp sites, additional electrodes were attached to below the left eye (to monitor for vertical eye movement/blinks), to the right of the right eye (to monitor for horizontal eye movements), over the left mastoid bone (reference) and over the right mastoid bone (recorded actively to monitor for differential mastoid activity). All EEG electrode impedances were maintained below 5 k (impedance for eye electrodes was less than 10 k). The EEG was amplified by an SA Bioamplifier with a bandpass of 0.01 and 40 Hz and the EEG was continuously sampled at a rate of 200 Hz throughout the experiment.
Data analysis
Averaged ERPs were formed off-line from trials free of ocular and muscular artifact (less than 7% of trials) using an in-house suite of software.3 Separate waveforms were calculated by averaging compound stimulus ERPs (i.e., the EEG time locked to a point 100 ms pre-prime onset and lasting until 900 ms post-target onset) based on the four types of critical trials: targets preceded by concatenated primes (e.g., cllet/COLLECT), targets preceded by concatenated unrelated primes (e.g., dlema/COLLECT), targets preceded by hyphenated primes (e.g., c-lle-t/COLLECT) and targets preceded by unrelated hyphenated primes (e.g., d-lem-a/COLLECT). All waves were baselined to the average activity in the 50 ms epoch immediately before target onset. In order to carefully quantify the time-course of the ERP effects, mean amplitudes in three post-target onset latency windows were measured: 175–240 ms (early N250), 240–305 ms (late N250), and 375–550 ms (N400). Repeated measures analyses of variance (ANOVAs) with within-subject factors of Prime Type (concatenated vs. hyphenated) and Relatedness (related vs. unrelated) were used to analyze the ERP data. The Huynh-Feldt correction was applied to all repeated measures with more than one degree of freedom. Three anterior (F3, Fz, F4), three middle (C3, Cz, C4) and three posterior (P3, Pz, P4) electrode sites were chosen for analysis which provided ANOVA factors of laterality (left vs. center vs. right) and AntPost (anterior vs. middle vs. posterior). We used this more restricted montage for two reasons. First to simplify the exposition of results (utilizing the entire montage requires multiple ANOVAs per epoch) and second, because the major effects reported were most clearly seen in these sites. We have successfully used this approach in several previous studies (e.g., Grainger et al., 2006b) and feel that when augmented by the use of voltage maps (which are computed across all sites and therefore give a picture of the entire data set), that it offers the best combination of thorough yet easily comprehensible analyses.
Acknowledgments
This research was supported by NICHD grants HD25889 and HD043251.
Footnotes
This type of relative-position priming is referred to as subset priming, since primes are formed of a subset of the target’s letters, to be distinguished from another form of relative-position priming using superset primes (e.g., sipflence-silence: Van Assche & Grainger, 2006; Welvaert, Farioli, & Grainger, 2008).
It is possible that what we are calling the early N250 effect is actually, in part, the exogenous P2 component. However, in the fast masked priming paradigm used here it is very difficult to know exactly which component(s) are contributing to such an effect – especially at relatively early points after target onset. This is because the ERP time locked to targets in this paradigm reflects not only components generated by the target itself but also those generated by the forward mask, prime and backward mask events - all of which occur within 80 ms of target onset. In other words, what looks like a target P2 in Figure 2 is actually an amalgamation of overlapping ERPs generated by all four stimuli, which makes it difficult to attribute differences to well-known exogenous ERP components such as the P2. Our approach for describing these early differences is to refer to them with labels that refer to effects (i.e., differences between conditions) rather than typical ERP components. We have discussed this approach in several previous papers (e.g., Holcomb & Grainger, 2006).
On average fewer than 8% of trials were rejected due to eye blinks/movements or EEG artifact, resulting in, on average per participant, the following numbers of trials per condition: 93 trials in the related concatenated, 93 trials in the related hyphenated, 94 trials in the unrelated concatenated and 92 trials in the unrelated hyphenated.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Carreiras M, Vergara M, Perea M. ERP correlates of transposed-letter priming effects: The role of vowels vs. consonants. Psychophysiology. 2009a doi: 10.1111/j.1469-8986.2008.00725.x. in press. [DOI] [PubMed] [Google Scholar]
- Carreiras M, Perea M, Vergara M, Pollatsek A. The time course of orthography and phonology: ERP correlates of masked priming effects in Spanish. Cerebral Cortex. 2009b doi: 10.1111/j.1469-8986.2009.00844.x. in press. [DOI] [PubMed] [Google Scholar]
- Chauncey K, Holcomb PJ, Grainger J. Effects of stimulus font and size on masked repetition priming: An ERP investigation. Language and Cognitive Processes. 2008;23:183–200. doi: 10.1080/01690960701579839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelissen P, Tarkiainen A, Helenius P, Salmelin R. Cortical effects of shifting letter position in letter strings of varying length. Journal of Cognitive Neuroscience. 2003;15:731–746. doi: 10.1162/089892903322307447. [DOI] [PubMed] [Google Scholar]
- Davis CJ. Unpublished doctoral dissertation. University of New South Wales; Australia: 1999. The self-organizing lexical acquisition and recognition (SOLAR) model of visual word recognition. [Google Scholar]
- Dufau S, Grainger J, Holcomb PJ. An ERP investigation of location invariance in masked repetition priming. Cognitive Affective and Behavioral Neuroscience. 2008 doi: 10.3758/cabn.8.2.222. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunabeitia JA, Molinaro N, Laka I, Estevez A, Carreiras M. N250 effects for letter transpositions depend on lexicality: Casual or causal? . Neuroreport. 2009 doi: 10.1097/wnr.0b013e3283249b1c. in press. [DOI] [PubMed] [Google Scholar]
- Eddy MD, Schmid A, Holcomb PJ. Masked repetition priming and event-related brain potentials: A new approach for tracking the time-course of object perception. Psychophysiology. 2006;43:564–568. doi: 10.1111/j.1469-8986.2006.00455.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez P, Ratcliff R, Perea M. The overlap model: A model of letter position coding. Psychological Review. 2008;115:577–601. doi: 10.1037/a0012667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J. Cracking the orthographic code: An introduction. Language and Cognitive Processes. 2008;23:1–35. [Google Scholar]
- Grainger J, Granier JP, Farioli F, Van Assche E, van Heuven W. Letter position information and printed word perception: The relative-position priming constraint. Journal of Experimental Psychology: Human Perception and Performance. 2006a;32:865–884. doi: 10.1037/0096-1523.32.4.865. [DOI] [PubMed] [Google Scholar]
- Grainger J, Holcomb PJ. Contraintes neurales pour une architecture fonctionnelle de la reconnaissance des mots. L’Année Psychologique. 2007;107:623–658. [Google Scholar]
- Grainger J, Holcomb PJ. Watching the word go by: On the time-course of component processes in visual word recognition. Language and Linguistic Compass. 2009;3:128–156. doi: 10.1111/j.1749-818X.2008.00121.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J, Kiyonaga K, Holcomb PJ. The time-course of orthographic and phonological code activation. Psychological Science. 2006b;17:1021–1026. doi: 10.1111/j.1467-9280.2006.01821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J, Van Heuven W. Modeling Letter Position Coding in Printed Word Perception. In: Bonin P, editor. The Mental Lexicon. New York: Nova Science Publishers; 2003. pp. 1–24. [Google Scholar]
- Grainger J, Whitney C. Does the huamn mnid raed wrods as a wlohe? Trends in Cognitive Sciences. 2004;8:58–59. doi: 10.1016/j.tics.2003.11.006. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. On the time-course of visual word recognition: En ERP investigation using masked repetition priming. Journal of Cognitive Neuroscience. 2006;18:1631–1643. doi: 10.1162/jocn.2006.18.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. Exploring the temporal dynamics of visual word recognition in the masked repetition priming paradigm using event-related potentials. Brain Research. 2007;1180:39–58. doi: 10.1016/j.brainres.2007.06.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J, O’Rourke T. An electrophysiological study of the effects of orthographic neighborhood size on printed word perception. Journal of Cognitive Neuroscience. 2002;14:938–950. doi: 10.1162/089892902760191153. [DOI] [PubMed] [Google Scholar]
- Humphreys GW, Evett LJ, Quinlan PT. Orthographic processing in visual word identification. Cognitive Psychology. 1990;22:517–560. doi: 10.1016/0010-0285(90)90012-s. [DOI] [PubMed] [Google Scholar]
- Kiyonaga K, Grainger J, Midgley KJ, Holcomb PJ. Masked cross-modal repetition priming: An event-related potential investigation. Language & Cognitive Processes. 2007;22:337–376. doi: 10.1080/01690960600652471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris J, Franck T, Grainger J, Holcomb PJ. Semantic transparency and masked morphological priming: AN ERP investigation. Psychophysiology. 2007;44:506–521. doi: 10.1111/j.1469-8986.2007.00538.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perea M, Carreiras M. Do transposed-letter similarity effects occur at a prelexical phonological level? Quarterly Journal of Experimental Psychology. 2006;59:1600–1613. doi: 10.1080/17470210500298880. [DOI] [PubMed] [Google Scholar]
- Perea M, Lupker SJ. Can Caniso activate Casino? Transposed-letter similarity effects with nonadjacent letter positions. Journal of Memory and Language. 2004;51:231–246. [Google Scholar]
- Peressotti F, Grainger J. The role of letter identity and letter position in orthographic priming. Perception & Psychophysics. 1999;61:691–706. doi: 10.3758/bf03205539. [DOI] [PubMed] [Google Scholar]
- Petit JP, Midgley KJ, Holcomb PJ, Grainger J. On the time-course of letter perception: A masked priming ERP investigation. Psychonomic Bulletin & Review. 2006;13:674–681. doi: 10.3758/bf03193980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoonbaert S, Grainger J. Letter position coding in printed word perception: Effects of repeated and transposed letters. Language and Cognitive Processes. 2004;19:333–367. [Google Scholar]
- Van Assche E, Grainger J. A study of relative-position priming with superset primes. Journal of Experimental Psychology: Learning, Memory and Cognition. 2006;32:399–415. doi: 10.1037/0278-7393.32.2.399. [DOI] [PubMed] [Google Scholar]
- Welvaert M, Farioli F, Grainger J. Graded effects of number of inserted letters in superset priming. Experimental Psychology. 2008;55:54–63. doi: 10.1027/1618-3169.55.1.54. [DOI] [PubMed] [Google Scholar]
- Whitney C. How the brain codes the order of letters in a printed word: The SERIOL model and selective literature review. Psychonomic Bulletin & Review. 2001;8:221–243. doi: 10.3758/bf03196158. [DOI] [PubMed] [Google Scholar]
- Whitney C. A comparison of the SERIOL and SOLAR theories of letter-position encoding. Brain and Language. 2008 doi: 10.1016/j.bandl.2007.08.002. in press. [DOI] [PubMed] [Google Scholar]