

Abstract
Background and Aims
Current agricultural practices are based on growing monocultures or binary mixtures over large areas, with a resultant impoverishing effect on biodiversity at several trophic levels. The effects of increasing the biodiversity of a sward mixture on dry matter yield and unsown species invasion were studied.
Methods
A field experiment involving four grassland species [two grasses – perennial ryegrass (Lolium perenne) and cocksfoot (Dactylis glomerata) – and two legumes – red clover (Trifolium pratense) and white clover (Trifolium repens)], grown in monocultures and mixtures in accordance with a simplex design, was carried out. The legumes were included either as single varieties or as one of two broad genetic-base composites. The experiment was harvested three times a year over three years; dry matter yield and yield of unsown species were determined at each harvest. Yields of individual species and interactions between all species present were estimated through a statistical modelling approach.
Key Results
Species diversity produced a strong positive yield effect that resulted in transgressive over-yielding in the second and third years. Using broad genetic-base composites of the legumes had a small impact on yield and species interactions. Invasion by unsown species was strongly reduced by species diversity, but species identity was also important. Cocksfoot and white clover (with the exception of one broad genetic-base composite) reduced invasion, while red clover was the most invaded species.
Conclusions
The results show that it is possible to increase, and stabilize, the yield of a grassland crop and reduce invasion by unsown species by increasing its species diversity.
Key words: Cocksfoot, Dactylis glomerata, diversity effect, invasion, legumes, perennial ryegrass, Lolium perenne, red clover, Trifolium pratense, simplex design, statistical modelling, transgressive over-yielding, white clover, T. repens
INTRODUCTION
The loss of natural ecosystems to agriculture and other human activities is projected to have a large global impact on biodiversity in the future (Chapin et al., 2000; Tilman et al., 2002). Areas under intensive agriculture, in which applications of high doses of mineral fertilizers to monoculture crops are a prominent feature, have a negative impact on biodiversity in Europe (Reidsma et al., 2006). However, maintaining acceptable levels of agricultural productivity will continue to be a high priority, mainly because of an increasing global population that needs to be fed, but also due to the need for economic sustainability of the individual farmer. Hence, there is an urgent need to develop agricultural practices that can deliver high yields with adequate forage quality, and simultaneously promote biodiversity. Biodiversity encompasses a broad spectrum of biotic scales (Hooper et al., 2005), with components ranging from genetic diversity within populations through to the functional diversity of species present and up-scale to ecosystem diversity.
Perennial grasslands occupied around 40 % of the land used for agriculture by the European Union prior to its recent enlargement (Rath and Peel, 2005). In productive temperate grassland systems grass monocultures are often used in order to simplify management options (e.g. easier to predict the optimal harvesting time, less variation in feeding quality). Seed use data for the EU in the 1990s showed that by far the most commonly sown grassland species was perennial ryegrass (Kley, 1995) and this remains the case today. However, large inputs of artificial nitrogen fertilizer are required to maximize the productivity of such monocultures, and this is increasingly regarded by policy makers and public opinion as being environmentally unacceptable. In addition to high yields, farmers may also value a stable yield level between years. Multi-species mixtures could provide greater and more stable primary production through temporal, spatial and resource niche complementarities (Sanderson et al., 2004; van Ruijven and Berendse, 2005; Roscher et al., 2008).
Although there is considerable evidence for a strong and persistent effect of diversity on yield in natural plant systems (Hooper et al., 2005; Balvanera et al., 2006; Cardinale et al., 2007; Schmid et al., 2002), few studies have explored biodiversity effects in a realistic agronomic context (but see Tracy and Sanderson, 2004a; Sanderson et al., 2005, Picasso et al., 2008). A positive diversity effect leading to transgressive over-yielding (mixture performing better than the best monoculture) was found in the first year's results from an agronomic study (in which some of the data of the present study were included) at 28 sites in Europe in which mixtures of four species (two legumes and two grasses) were compared with the monoculture performance of these species (Kirwan et al., 2007). Genetic diversity might be expected to provide additional benefits through improved resistance to biotic (e.g. Finckh et al., 1999; Cox et al., 2004; Pilet et al., 2006) and abiotic environmental stresses (Hajjar et al., 2008), and enhanced niche complementarity (Hooper et al., 2005). Further, it has been proposed that multi-species mixtures should be more resistant to invasion by unsown species than crops composed of fewer species, and this has been confirmed in some studies, e.g. Hector et al. (2001), Tracy et al. (2004), Tracy and Sanderson (2004b) Picasso et al. (2008). Although the evidence from a meta-analysis of 44 biodiversity experiments that manipulated plant species richness is that transgressive over-yielding is not, in general, of wide occurrence and requires several years to develop (Cardinale et al., 2007), this conclusion may partly be due to methodological difficulties in its assessment (Schmid et al., 2008). Persistence and an increase in a positive diversity effect in natural plant systems with time have been noted (Hooper and Dukes, 2004; van Ruijven and Berendse, 2005; Bullock et al., 2007; Cardinale et al., 2007; Fargione et al., 2007) but whether the effect persists through time in perennial agronomic grassland systems is of considerable interest.
In this paper results are reported from a field study comprising multi-species mixtures of four grassland species [two grasses and two legumes: perennial ryegrass (Lolium perenne) and cocksfoot (Dactylis glomerata) and red clover (Trifolium pratense) and white clover (T. repens)] using the approach described in Kirwan et al. (2007). The experiment also included a treatment in which the impact of intraspecific variation in the legume species was assessed by the use of single varieties versus broad genetic-base composites, comprising mechanical mixtures of seed of many varieties (Collins et al., 2004). It was hypothesized that: (1) positive interactions between species would result in larger yields from mixtures than expected from the performance of the individual species sown as monocultures; (2) these benefits would persist over several years; (3) a broad genetic-base composite would provide a platform for further and sustained positive effects of mixing species; and (4) increased species diversity and the use of broad genetic-base composites would help to prevent invasion of unsown species into the swards.
MATERIALS AND METHODS
Site data and plant material
The field experiment was conducted at Svalöv in southern Sweden (56 °55′N, 13 °07 E, 55 m a.s.l.). The climate is cold-temperate with an annual mean temperature of 7·7 °C and an annual precipitation averaging 700 mm. The soil at the site was a sandy clay loam with a pH of 6·9 and containing 2·12 % organic matter, 1·3 g nitrogen, 0·36 cmol potassium and 44 mg phosphorus kg−1 soil. Plots were established in late June 2003 by drilling, and initial invasion by unsown species was controlled by spraying with the herbicide bentazon (which is tolerated by the species sown) shortly after the initial emergence of the crop. The species used were two grasses and two legumes, of which one species within each functional group was relatively fast establishing but not persistent (Lolium perenne L. and Trifolium pratense L.), and the other was relatively slow establishing and persistent (Dactylis glomerata L. and Trifolium repens L.). Information on the plant material is presented in Table 1. For each legume species there was one single variety of species (denoted N) and two broad genetic-base composites, comprising ‘Broad Central European’ (BC) and ‘Broad Northern European’ (BN) populations. The single varieties of species used (N) had been bred for agricultural use within the experimental area. The red clover BC composite contained seed of 11 commercial varieties bred for use in temperate regions, plus a small amount (23 % by weight) of a ‘northern red clover composite’ population from Norway/Denmark, and encompassed all categories employed by red clover breeders (early/late flowering; diploid/tetraploid). The red clover BN composite contained seed from a population which was created by the inter-crossing for two generations of 238 accessions comprising the complete red clover collection held in the Nordic Gene Bank, plus varieties from other parts of the world (mainly from the former USSR; Á. Helgadóttir, Agricultural University of Iceland, Reykjavik, pers. comm.). The white clover BC composite contained seed of 13 commercial varieties bred for use in temperate regions, plus a small amount (5 % by weight) of ‘gene pool’ material from Central Asia. The white clover BN composite contained seed from nine varieties bred for use in northern Scandinavia, plus ten commercial varieties bred for use in southern Scandinavia (P. Marum, Graminor AS, Ilseng, Norway, pers.comm.).
Table 1.
Plant material and monoculture seeding rates
| Species | Population | Monoculture seeding rates (kg ha−1) for high sowing density |
|---|---|---|
| Perennial ryegrass (Lolium perenne) | Helmer | 30 |
| Cocksfoot (Dactylis glomerata) | Dactus | 16 |
| Red clover (Trifolium pratense) | Fanny, N | 15 |
| Red clover | Central European composite, BC | 15 |
| Red clover | Northern European composite, BN | 15 |
| White clover (Trifolium repens) | Ramona, N | 8 |
| White clover | Central European, BC | 8 |
| White clover | Northern European, BN | 8 |
Seeding rates of the respective species in monoculture in the high density treatment (see below) are found in Table 1. These corresponded to the rates used in official Swedish variety trials. Low density plots were sown at 60 % of these rates. In mixtures, the seeding rate of each species was a proportion (see below) of the monoculture seeding rates.
The experimental layout consisted of 66 communities. Thirty communities followed the simplex design described in Kirwan et al. (2007) with two grass and two legume (N) monocultures and 11 mixtures of these four species sown at two densities. The 11 mixtures consisted of four mixtures dominated in turn by each species (sown proportions were 70 % of dominant and 10 % of each other species), six mixtures dominated in turn by pairs of species (40 % of each of the two species and 10 % of the other two) and the centroid community (25 % of each species). Eighteen communities had four monocultures [two grasses (same as before) and two legumes (BC)] at two densities and five mixtures of these four species sown at two densities (sown proportions were 70 % of the dominant and 10 % of each of the other species, plus the centroid community). Another 18 communities followed the same design but with BC replaced by BN. Experimental communities were randomly assigned to 8·8 m2 plots. Data from one plot were omitted since an erroneous species mixture was sown to this plot. General information on simplex designs is available in Cornell (2002).
No harvests were taken in the establishment year. In the following three years the whole plots were harvested with an experimental plot harvester (Haldrup) 3 times a year, with a stubble height of approx. 5 cm (normal practice within this climatic zone). At each harvest biomass yield was determined and samples for analyses of dry matter (DM) and botanical composition were taken from all plots. White clover ingress into neighbouring plots was prevented by regular spraying of the borders with glyphosate. No fertilizer nitrogen was added to the experiment. In the final year of the experiment notes were taken of the unsown species identity present in all plots.
Statistical analysis
Annual sown DM yield and unsown species DM % for each year and sown DM yield for each harvest were analysed. The modelling approach of Kirwan et al. (2007) was used to relate sown yield (DMyield) for a single harvest or year to variables representing species sown proportions of monoculture seeding rates, overall density and species interaction. Sown proportions were denoted in the model as G1 (ryegrass) and G2 (cocksfoot) for the two grasses and L1N, L1BC and L1BN for the N, BC and BN genetic treatments, respectively, for the first legume species (L1, red clover). A similar notation was used for the second legume species (L2, white clover). DENS is a variable indicating high or low seeding density, coded −1 for low and 1 for high. A single variable (evenness) was used to describe interspecific interaction in a mixture. The evenness (E) of a mixture as defined in Kirwan et al. (2007) lies between 0 for a monoculture and 1 for a mixture in which the sown proportions of all species are equal (the centroid). They used E and E2 as explanatory variables to capture the effect of species interactions. The model fitted was
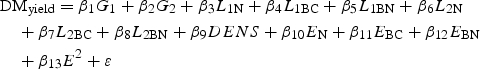 |
1 |
The coefficients β1 and β2 represent the monoculture responses of the grass species and the coefficients β3 to β8 represent the monoculture responses of the legume species, at average density. For example, 3·84, the estimate of β1 for year 1 (see Table S1 in Supplementary data available online), estimates the monoculture yield of perennial ryegrass for that year. The evenness coefficients (β10, β11 and β12) measure the effects of evenness for mixtures involving each of the legume populations. β13 is a quadratic effect of evenness that does not change with legume population. Thus, for mixtures involving N legumes, the species diversity effect (the contribution of interaction effects to yield of a mixture at evenness EN over what could be expected from a proportional mixture of monoculture yields) is β10EN + β13EN2 estimated by 5·62EN – 2·14EN2 (Table S1). For the centroid (equal sown proportion of all species), EN = 1 and the predicted diversity effect is 5·62 – 2·14 = 3·48 (Table 2). Finally, ε is a random error term assumed to be normally and independently distributed with mean of zero and constant variance. Differences were tested between coefficients for grass species (β1 vs. β2), for legume species (β3 vs. β6, β4 vs. β7, and β5 vs. β8) and legume populations (β3 vs. β4 vs. β5 and β6 vs. β7 vs. β8), and the evenness effect was compared between different legume populations (β10 vs. β11 vs. β12) and they were tested against zero.
Table 2.
Mean estimated species diversity effects for sown dry matter yield at the centroid (t ha−1) of the three legume population treatments for each of the three harvest years with tests of significance; also shown for each year are yields (t ha−1) at the centroid predicted both from monoculture performance only, and including the species diversity effect
| Harvest year 1 |
Harvest year 2 |
Harvest year 3 |
|||||||
|---|---|---|---|---|---|---|---|---|---|
| Estimate | s.e. | P | Estimate | s.e. | P | Estimate | s.e. | P | |
| Estimated species diversity effect at the centroid | |||||||||
| N | 3·48 | 0·277 | <0·001 | 4·82 | 0·231 | <0·001 | 3·7 | 0·241 | <0·001 |
| BC | 3·96 | 0·339 | <0·001 | 5·3 | 0·303 | <0·001 | 4·18 | 0·311 | <0·001 |
| BN | 3·21 | 0·349 | <0·001 | 4·55 | 0·313 | <0·001 | 3·44 | 0·321 | <0·001 |
| Estimated yield at the centroid based on monoculture performances | |||||||||
| N | 7·14 | 0·173 | 3·81 | 0·142 | 1·99 | 0·149 | |||
| BC | 7·04 | 0·175 | 4·05 | 0·143 | 2·16 | 0·150 | |||
| BN | 7·34 | 0·193 | 4·04 | 0·160 | 1·78 | 0·168 | |||
| Estimated yield at the centroid including the species diversity effect | |||||||||
| N | 10·62 | 0·220 | 8·63 | 0·180 | 5·70 | 0·189 | |||
| BC | 11·00 | 0·306 | 9·35 | 0·266 | 6·34 | 0·275 | |||
| BN | 10·56 | 0·308 | 8·59 | 0·267 | 5·22 | 0·276 | |||
Comparisons among estimates within and across years are presented in the text. N denotes a legume variety, while BC and BN denote Broad Central European and Broad Northern European genetic-base composites, respectively.
Repeated measures analyses were carried out using the MIXED procedure and univariate analyses using the GLM Procedure in the SAS/STAT software, Version 9·1 of the SAS System for Windows.
Repeated measures analyses of sown species biomass were performed across harvests within each year and for annual sown yields across the 3 years of the experiment based on model (1) augmented with terms for interactions with time, which were significant. An unstructured covariance matrix was used to describe the time-dependence among harvests or years (Verbeke and Molenbergh, 2000). The effects of year and legume population were tested on the estimated species diversity effect at the centroid and the average monoculture yield across years. Transgressive over-yielding was tested for each legume population using a non-parametric test similar to that described in Kirwan et al. (2007).
The percentage of total annual unsown species biomass in a repeated measures analysis across years was analysed using model (1) and including year effects as just described. Responses were log-transformed before analysis to reduce heteroscedasticity. Many mixed plots were free from unsown species in the two first harvest years (14 in each year), and the recorded zero values from these plots were replaced by 0·0014, the lowest recorded non-zero value, before transformation.
There was no interaction between density and the other model variables. Model (1) (and its extensions for repeated measures analysis across years) generally emerged as either the dominant or the best of a number of alternative models for overall sown yield, for most harvests and years and for log of unsown species yield (by dominant it is meant that the explanatory power of the evenness-based species diversity effects greatly exceeds any refinement introduced by partitioning the evenness effect into more coefficients as described below). Alternative models included a model with separate interaction coefficients for each pairwise interaction, and a model that included only two pairwise interaction coefficients, one for an interaction between any grass and any legume and another for grass–grass or legume–legume interaction (Kirwan et al., 2007). Other models allowed the species diversity effect to vary with legume species. There was some evidence in favour of separate pairwise coefficients for each interaction for sown yield at some harvests and for a separate coefficient × year interaction for the analysis of sown annual yield. In the latter analysis there was no clear pattern among the six interaction coefficients across years. Mixtures were not replicated and the variation around the regression model was used to estimate the residual mean square and the standard errors of coefficient estimates (Cornell, 2002).
RESULTS
Species-specific effects
Estimates of coefficients of model (1) fitted to annual sown yield as a repeated measure over 3 years show that there was a considerable and significant decline in monoculture yield for all species across years (P < 0·05 for each test; Fig. 1, and Table S1 in Supplementary data, available online). The estimated DM yield of the cocksfoot monoculture was consistently greater than that of perennial ryegrass (P < 0·01 in each harvest year; Table S1 in Supplementary data, available online). White clover consistently outyielded the two grasses over the 3 years (P < 0·01 for each test except L2BN vs. G2 in year 3), while red clover did so in the first 2 years (P < 0·001 for each test). There was generally no strong or consistent effect of legume populations in the monoculture performances of either legume species for any year. Red clover outyielded white clover in the first year (P < 0·001 for N and BC populations, P = 0·06 for BN) but underyielded it in the third year (P < 0·01 for all populations). Density never affected DM yield.
Fig. 1.

Total yield predicted for each of the four monocultures and for three mixtures divided into sown and unsown species for each year. The four monocultures are Dactylis glomerata, Lolium perenne, Trifolium pratense (averaged over N, BC and BN) and Trifolium repens (averaged over N, BC and BN). The three mixture communities are the Centroid (0·25, 0·25, 0·25, 0·25), Mixture 1 (the average of the mixtures with two species dominant, e.g. 0·4, 0·4, 0·1, 0·1.) and Mixture 2 (the average of the mixtures with one species dominant, e.g. 0·7·01 0·1 0·1). Standard error bars are given for the predicted total yield.
While the cumulative yield for each year was little affected by the broad genetic-base composite treatment, the different legume populations nonetheless showed a different yielding pattern within each year (see Table S2 in Supplementary data, available online). Monocultures of the BC red clover populations had higher estimated yield (β4 vs. β3 and β4 vs. β5 red clover) than the N and BN red clover populations in the final harvest of each year, significantly so (P < 0·01) in the first 2 years. Further, monocultures of the BC white clover populations had higher estimated yield than the BN white clover populations in the final harvest of each year (β7 vs. β8 white clover), significantly so in 2 of the 3 years. In the other harvests differences were rarely significant.
Diversity effects
Evidence of transgressive over-yielding was found in 3-year accumulated yield in the mixtures using N and BC legumes (P < 0·05; Table 3). Transgressive over-yielding was observed in the second and the third years for mixtures using all legume populations (P < 0·05).
Table 3.
Results of tests for transgressive over-yielding
| Year | Harvest | N | BC | BN |
|---|---|---|---|---|
| 1 | 1 | 4 | 2 | 3 |
| 2 | 0 | 2 | 0 | |
| 3 | 7 | 1 | 2 | |
| 2 | 1 | 11 | 5 | 5 |
| 2 | 10 | 4 | 0 | |
| 3 | 5 | 2 | 4 | |
| 3 | 1 | 11 | 5 | 5 |
| 2 | 9 | 2 | 1 | |
| 3 | 11 | 4 | 2 | |
| 1 | 6 | 1 | 2 | |
| 2 | 11 | 5 | 5 | |
| 3 | 11 | 5 | 5 | |
| 1 | 11 | 5 | 5 | |
| 2 | 8 | 3 | 1 | |
| 3 | 8 | 3 | 4 | |
| Total for 3 years | 10 | 5 | 3 |
The test is carried out for each of the three populations (N, BC and BN), for each harvest by year combination, for yield accumulated each year and for yield accumulated over all 3 years. N denotes a legume variety, while BC and BN denote Broad Central European and Broad Northern European genetic-base composites, respectively. The value given in the table is the test statistic*, d, which is the absolute difference between the number of mixture means better than the best monoculture and worse than the worst monoculture. Bold indicates significance of the test (P < 0·05). The test is based on the non-parametric test described in Kirwan et al. (2007).
* Transgressive over-yielding occurs when the yield of a mixture community exceeds that of the highest-yielding monoculture of its component species. Here, a permutation test is used to determine the significance of transgressive over-yielding. There were 11 (4), 5 (4) and 5 (4) mixture communities (monoculture communities) in the three legume populations N, BC and BN, respectively. The average across replicates for each monoculture and mixture community was calculated within each legume population except for grass monocultures which were averaged across the legume populations (there was no difference between the grass monocultures for each legume population), giving 15, 9 and 9 community values for N, BC and BN respectively. The test statistic, d, is calculated from these community values as the absolute difference between the number of mixtures greater than the best monoculture and lower than the worst monoculture. Significance was tested using a permutation test that compares the result against that which would be expected by selecting 11 (or 5) values at random from 15 (or 9) and comparing them with the best and worst of the remaining 4 (or 4) values. The test is a two-sided test as the mixtures could be better than the best monoculture or worse than the worst monoculture (under-yielding) and the direction cannot be prejudged in advance. For more details see supplementary material in Kirwan et al. (2007).
There was a strong and persistent species diversity effect irrespective of the legume population used. The linear evenness effects were always positive and greater in magnitude than the quadratic evenness effects (Table S1 in Supplementary data available online), giving a positive species diversity effect that remained strong over the 3 years (Table 2). The linear evenness effect varied with both legume population used (P < 0·001) and year (P < 0·001), and the quadratic evenness effect varied with year (P < 0·001; Table S1 in Supplementary data, available online). The maximum species diversity effect occurred at a level of evenness at or close to 1, the centroid. The species diversity effect was calculated at the centroid for each genetic diversity level × year combination and also the mean yield calculated at the centroid predicted from the monoculture yields of species, i.e. with no diversity effect added, and the mean yield including the species diversity effect (Table 2). The species diversity effect at the centroid increased between the first and second years (P < 0·001; Table 2) and in the third year declined to levels similar to those in the first year (P < 0·001; Table 2) for each of them. Yield at the centroid predicted from an equal mixture of monoculture yields declined strongly over time, irrespective of the legume population used (P < 0·001 for each test; Table 2). Predicted yield at the centroid including the species diversity effect also declined over time (P < 0·001 for each test; Table 2) but far less rapidly than when based solely on monoculture performance. Thus, the species diversity effect on yield relative to yield predicted from the respective monoculture performances was much higher in the second and third years than in the first year. Mixtures with the BC legumes produced a generally stronger species diversity effect than those with BN legumes (P < 0·05), while the species diversity effect of the mixtures including the N legume was not significantly different from that of either of the two broad genetic-base composites (Table 2).
Invasion by unsown species
The effect of species diversity was very marked and persisted through time (significant species diversity effect each year P < 0·001), and the predicted level of unsown species at the centroid was <2 % even in the third year (Table 4).
Table 4.
Estimated biomass percentage of unsown species in the respective monocultures and at the centroid for each of three harvest years (HY)
| Estimated weed content ( %) |
|||
|---|---|---|---|
| Parameter | HY 1 | HY 2 | HY 3 |
| β1(G1 – ryegrass) | 1·5 | 3·4 | 41·3 |
| β2(G2 – cocksfoot) | 0·2 | 0·7 | 5·9 |
| β3(L1N – red clover) | 1·2 | 12·6 | 68·2 |
| β4(L1BC – red clover) | 1·2 | 12·6 | 68·2 |
| β5(L1BN – red clover) | 1·0 | 10·6 | 57·2 |
| β6(L2N – white clover) | 1·4 | 2·3 | 6·0 |
| β7(L2BC – white clover) | 1·3 | 2·2 | 5·8 |
| β8(L2BN – white clover) | 5·9 | 10·1 | 26·2 |
| Centroid | |||
| N | 0·1 | 0·1 | 1·1 |
| BC | 0·1 | 0·1 | 1·1 |
| BN | 0·1 | 0·1 | 1·5 |
N denotes legume varieties, while BC and BN denote Broad Central European and Broad Northern European genetic-base composites, respectively. The centroid is the mixture sown with equal proportions of the four species.
The most striking monoculture effects observed were the ability of cocksfoot and white clover (N and BC) to resist invasions by unsown species (Table 4, and Table S3 in Supplementary data available online). Even in the third year levels of unsown species were <6 % in these monocultures, whereas they increased over years to high levels for monocultures of both ryegrass (41 %) and red clover (around 60 %). Using broad genetic-base composites appeared to have little effect on the amount of unsown species in red clover swards, but the BN white clover population contained a larger amount than the N or the BC populations (P < 0·01; Table 4).
DISCUSSION
The present study differs from many earlier biodiversity studies (e.g. Hector et al., 1999; Tilman et al., 2001; Roscher et al., 2003) with respect to the species used (all of major agronomic importance in temperate regions) and the management carried out (aimed at maximizing biomass harvested; see Hooper et al., 2005). The design of the experiment also differed from the above-mentioned studies in establishing a gradient of evenness with all species present in all mixed communities, instead of just increasing species richness as a treatment.
With respect to the four main hypotheses addressed in this study it was found that species diversity had a strong, persistent and positive effect on yield and on the ability of the sward to resist invasion by unsown species. However, using a broad genetic-base composite in two of the four species in the experiment had little impact on these ecosystem properties.
Diversity effects on yield
It was hypothesized that the mixing of species would result in larger yields from the mixtures than would be expected from the individual species monoculture yields. That mixing of grasses with legumes, with a resulting niche complementarity for N (Palmborg et al., 2005; Kahmen et al., 2006), results in a positive yield effect has been shown in other studies (e.g. Spehn et al., 2002). However, when a model that partitioned the diversity effect into a term representing legume × grass interaction was fitted, and a term jointly representing grass × grass and legume × legume interactions (Kirwan et al., 2007) the two coefficients did not differ significantly, suggesting that the evenness model was a more parsimonious description of these data. This suggests that the effects of mixing grasses or mixing legumes had the same impact on increasing yield as did functional group mixing between grasses and legumes. Similar results have been reported by van Ruijven and Berendse (2003, 2005) and Hector et al. (2007). Thus, it appears that the characteristics of the species within each functional group were either of a complementary or a facilitating nature (Hooper et al., 2005). The species diversity effect based on between-species interactions was always positive. In the centroid mixture the average contribution to yield of the species diversity effect were 3·55, 4·89 and 3·77 tons DM ha−1 (+33 %, +55 % and +65 % yield increase over mean monoculture performances) in the first, second and third years respectively. This diversity effect is stronger than the mean effect observed for the first harvest year across 28 European sites (Kirwan et al., 2007).
The sustained species diversity effect and the declining monoculture effects suggest that the importance of this diversity contribution to yield increased with time. This was reflected in the increasing evidence of transgressive over-yielding for all legume populations as time advanced. Reports of transgressive over-yielding in biodiversity experiments carried out in the field with grassland species have been rare (e.g. Hooper and Dukes, 2004; Cardinale et al., 2007; Lanta and Lepš, 2007; Schmid et al., 2008; but see Schmid et al., 2002). It is argued (Schmid et al., 2008) that a wide range in monoculture yields makes it difficult to observe transgressive over-yielding. However, in some studies where transgressive over-yielding has been observed, the species involved have been high-yielding (Jolliffe and Wanjau, 1999; Roscher et al., 2005), the soil has been nutrient-rich (Roscher et al., 2008) and species richness has been low (Roscher et al., 2005), which was also the case in the present experiment. This suggests that transgressive over-yielding should be more common in an agricultural context, where soil fertility usually is good and the species used are selected on the basis of their yielding properties, than it is in natural communities in which low-yielding species frequently occur. Indeed, in an analysis of 44 plant experiments (39 grassland) with species number ranging from 6 to 32 (Cardinale et al., 2007) the yield of the highest yielding monoculture was 93 % above the yield of the average monoculture and transgressive over-yielding was only observed in 12 % of cases. However, in the present data, the monoculture variation was lower than this but still of a substantial magnitude (the highest yielding monoculture exceeded the average monoculture by 51, 51 and 77 % in harvest years 1, 2 and 3, respectively). So, the emergence of transgressive over-yielding over time here is not due to a relatively small species diversity effect being compared with rather similar yielding species in monoculture, but rather to the decline in monoculture yields over time and the persistence of a relatively large species diversity effect.
The persistent and increasing effect of species diversity on yield has important practical implications. In farming practice, the application of fertilizer nitrogen by European grassland farmers is a common way to increase and/or maintain yield over time. Given the need to reduce the load of nitrogen into the European environment (e.g. Rougoor and van der Weijden, 2001) the practical benefit of this diversity effect is obvious, and the use of mixtures could therefore be recommended as a way of reducing inputs. It should be noted that no fertilizer nitrogen was added to this experiment.
Using broad genetic-base composite populations of the two legumes had a relatively small impact on yield in the current experiment, contrary to hypothesis (3). This could be due to the fact that traits are more likely to be similar within species than between different species. Similar results have been obtained in studies with white clover by Annicchiarico and Piano (1997) and Williams et al. (2003). Legumes, and red clover in particular, are susceptible to a vast range of pathogens that destroy sensitive genotypes (Frame et al., 1998), and the loss of red clover plants due to diseases was very high in this experiment. Although there are no data on disease susceptibility of the red clover populations or pathogen load in the plots, the fact that yields of all red clover populations were similar in the final year suggest no difference in susceptibility to pest and diseases among them. Resistance of perennial plant species to abiotic stress during winter is often gained at the expense of late-season growth (e.g. white clover; Eagles and Othman, 1989). However, in the environment of southern Sweden the observed differences in end-of-season growth pattern of the legumes did not appear to have any impact on the overwintering abilities of the populations (as judged by monoculture spring yields). Attempts in the USA to improve the yield of a lucerne (Medicago sativa) sward by mixing varieties with a different seasonal growth pattern were short-lived, and this strategy was not recommended as a way to improve yield (Brummer et al., 2002). Another explanation for the weak effect of the broad genetic-base composites might be the occurrence of selection of the most adapted individuals for the site and the management over time (e.g. Frankow-Lindberg, 1999, 2001). Joshi et al. (2001) found that red clover ecotypes performed best in their original sites compared with more distant sites, and their performance was poorer with increased transplanting distance. The broad genetic-base composites might therefore not have been well adapted to Swedish conditions. Hypothesis (3) was thus not confirmed, but it is possible that the use of a broad genetic-base composites of varieties specifically adapted to southern Sweden might have resulted in a more long-lasting complementary effect within the plant community.
Diversity effects on unsown species invasion
The species diversity effect on unsown species was very strong, confirming hypothesis (4). The nature of the relationship between species diversity and resistance to invasion by unsown species is a hotly debated topic among ecologists (e.g. Fridley et al., 2007). In this experiment, where no weeding was carried out, both species diversity and species identity were important in controlling invasion. The main unsown species recorded was Taraxacum officinale, which is a wind-dispersed species that does not form a persistent soil seed bank. This shows that the soil seed bank was a factor of little importance for invasion by unsown species at this site, and that any effect of the herbicide application at the very beginning of the experiment had little (if any) carry-over effect on this ecosystem property.
An increased invasion resistance in species-diverse grassland communities compared with less diverse communities has been found in studies where initially unsown species have been deliberately introduced (Fargione et al., 2003), where unsown species have been removed over time (Knops et al., 1999; Hector et al., 2001; Tracy and Sanderson, 2004b), when ingress by unsown species in the harvested biomass was determined (Picasso et al., 2008), and in surveys of pastures across a wide geographic area (Tracy and Sanderson, 2004b). Similar results have also been obtained for annual crops (Hauggaard-Nielsen et al., 2008). However, plant functional diversity effects on invasion resistance have not been extensively studied. In this context, it is of interest to note that the most common of the unsown species observed in the experiment belonged to a functional type (deep-rooted forb) that was not present in the seeding mixtures (Fargione et al., 2003; Hooper et al., 2005). Species with competitive traits may, however, play a key role (see below). The causal mechanism for the increased invasion resistance by species-diverse communities is likely to be a more complete utilization of environmental resources (Hooper et al., 2005). Differences in temporal growth patterns between the grasses and legumes currently used is, for example, a generally accepted agronomic fact (see Table S2 in Supplementary data available online).
Of the monocultures, the cocksfoot and two of the white clover (N and BC) communities were least susceptible to invasion, while all red clover monocultures were the most invaded. The reason why the BN white clover monoculture did not conform to the pattern might be that it had a lower growth rate at the end of the growing season, leaving more ‘space’ for unsown species to enter the sward at this time of the year. Since an efficient herbicide treatment was applied to this experiment shortly after sowing, the ingress of unsown species reflected characteristics of the mature community. Mwangi et al. (2007) found that grasses exerted a strong negative effect on invaders, whilst legumes had a positive effect, which to some extent agrees with the results from this study, since red clover resisted invasion poorly. Crawley et al. (1999), reporting a 7-year experiment with four levels of species richness where unsown species were allowed to accumulate in the plots, concluded that species identity mattered more than species richness in determining both the number of invading species and the total biomass of the invaders. Legumes play a vital role in the nitrogen economy of grassland communities (Spehn et al., 2002; Temperton et al., 2007), and it has been found that community invasibility is positively correlated with nitrate availability in the soil (Dukes, 2001). However, since all white clover monocultures contained substantially lower amounts of unsown species than the red clover monocultures, this cannot be the only mechanism that facilitated invasion of unsown species into the red clover plots. The fact that red clover plants died throughout the experimental period, leaving large gaps that could provide entry points for invading species, was probably just as important (see, for example, Milbau et al., 2003, 2005). One trait common to cocksfoot and white clover is the ability to spread laterally, cocksfoot both above- (Lorentzen et al., 2008) and below ground (Personeni and Loiseau, 2004), and white clover above ground (Frame et al., 1998). This trait would result in a more complete resource utilization, thus preventing the establishment of unsown species. The present monoculture results (Table 4) are in accordance with the suggestion made by Richardson and Pyšek (2006) that, in future, more emphasis should be placed on species identities and their characteristics in the search for an increased understanding of community invasibility, but in this study the species mixture results showed an even stronger effect.
It is concluded that increased species diversity within a forage crop provided clear and persistent yield benefits. This biodiversity effect could be profitably exploited by grassland farmers, and has the potential to reduce requirements for fertilizer inputs. Using broad genetic-base composites of the legumes was of minor importance in terms of yield at this site, although the use of broad genetic-base populations containing site-adapted germplasm may have produced a different result. Invasion by unsown species was generally reduced by increased species diversity of the sward. However, the strong effect of individual species identity suggests that traits conferring a more complete resource utilization also contributed to control invasibility.
SUPPLEMENTARY DATA
ACKNOWLEDGEMENTS
We thank SW Seed for providing access to the field site and excellent technical field assistance, and Mrs Annika Hansson for doing the botanical analyses. Thanks to An Gehesquiere, Vibeke Meyer, Petter Marum and Loek van Soest for providing seed for the broad genetic-base clover populations. This work was supported by a grant from The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (22·9/2003-0823) and the EU Commission through COST Action 852. The Faculty of Natural Resources and Agricultural Sciences at Swedish University of Agricultural Sciences provided support to B.E.F.-L. for a stay at University College Dublin.
LITERATURE CITED
- Annicchiarico P, Piano E. Response of white clover genotypes to intergenotypic and interspecific interference. Journal of Agricultural Science. 1997;128:431–437. [Google Scholar]
- Balvanera P, Pfisterer AB, Buchmann N, et al. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters. 2006;9:1146–1156. doi: 10.1111/j.1461-0248.2006.00963.x. [DOI] [PubMed] [Google Scholar]
- Brummer EC, Moore K, Bjork NC. Agronomic consequences of dormant-nondormant alfalfa mixtures. Agronomy Journal. 2002;94:782–785. [Google Scholar]
- Bullock JM, Pywell RF, Burke MJW, Walker KJ. Restoration of biodiversity enhances agricultural production. Ecology Letters. 2007;4::185–189. [Google Scholar]
- Cardinale BJ, Wright JP, Cadotte MW, et al. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences of the USA. 2007;104:18123–18128. doi: 10.1073/pnas.0709069104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapin FS, III, Zavaleta ES, Eviner VT, et al. Consequences of changing biodiversity. Nature. 2000;405:234–242. doi: 10.1038/35012241. [DOI] [PubMed] [Google Scholar]
- Collins RP, Connolly J, Porqueddu C. Effects of legume genetic diversity on the productivity of legume/grass mixtures – COST Action 852. Grassland Science in Europe. 2004;9:486–488. [Google Scholar]
- Cornell JA. Experiments with mixtures: designs, models and the analysis of mixture data. 3rd edn. New York, NY: John Wiley and Sons; 2002. [Google Scholar]
- Cox CM, Garrett KA, Bowden RL, Fritz AK, Dendy SP. Cultivar mixtures for the simultaneous management of multiple diseases: tan spot band leaf rust of wheat. Phytopathology. 2004;94:961–969. doi: 10.1094/PHYTO.2004.94.9.961. [DOI] [PubMed] [Google Scholar]
- Crawley MJ, Brown SL, Heard MS, Edwards GR. Invasion-resistance in experimental grassland communities: species richness or species identity? Ecology Letters. 1999;2:140–148. [Google Scholar]
- Dukes JS. Biodiversity and invasibility in grassland microcosms. Oecologia. 2001;126:563–568. doi: 10.1007/s004420000549. [DOI] [PubMed] [Google Scholar]
- Eagles CF, Othman OB. Yield and persistency of contrasting white clover populations grown in pure swards and in mixed swards with S.23 perennial ryegrass. Annals of Applied Biology. 1989;114:545–557. [Google Scholar]
- Fargione J, Brown CS, Tilman D. Community assembly and invasion: an experimental test of neutral versus niche processes. Proceedings of the National Academy of Sciences of the USA. 2003;100:8916–8920. doi: 10.1073/pnas.1033107100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fargione J, Tilman D, Dybzinski R, et al. From selection to complementarity: shifts in the causes of biodiversity-productivity relationships in a long-term biodiversity experiment. Proceedings of the Royal Society. 2007;274:871–876. doi: 10.1098/rspb.2006.0351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finckh MR, Gacek ES, Czembor HJ, Wolfe MS. Host frequency and density effects of powdery mildew and yield in mixtures of barley cultivars. Plant Pathology. 1999;48:807–816. [Google Scholar]
- Frame J, Charlton JFL, Laidlaw AS. Temperate forage legumes. Wallingford: CAB International; 1998. [Google Scholar]
- Frankow-Lindberg BE. Effects of adaptation to winter stress on biomass production, growth and morphology of three contrasting white clover cultivars. Physiologia Plantarum. 1999;106:196–202. [Google Scholar]
- Frankow-Lindberg BE. Adaptation to winter stress in nine white clover populations: changes in non-structural carbohydrates during exposure to simulated winter conditions, and ‘spring’ regrowth potential. Annals of Botany. 2001;88:745–751. [Google Scholar]
- Friedley JD, Stachowicz JJ, Naeem S, et al. The invasion paradox: reconciling pattern and processes in species invasions. Ecology. 2007;88:3–17. doi: 10.1890/0012-9658(2007)88[3:tiprpa]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Hajjar R, Jarvis DI, Gemmill-Herren B. The utility of crop genetic diversity in maintaining ecosystem services. Agriculture, Ecosystem and Environment. 2008;123:261–270. [Google Scholar]
- Hauggaard-Nielsen H, Jørnsgard B, Kinane J, Steen Jensen E. Grain legume – cereal intercropping: the practical application of diversity, competition and facilitation in arable organic cropping systems. Renewable Agriculture and Food Systems. 2008;23:3–12. [Google Scholar]
- Hector A, Schmid B, Beierkuhnlein C, et al. Plant diversity and productivity experiments in European grasslands. Science. 1999;286:1123–1127. doi: 10.1126/science.286.5442.1123. [DOI] [PubMed] [Google Scholar]
- Hector A, Dobson K, Minns A, Bazeley-White E, Lawton JH. Community diversity and invasion resistance: an experimental test in a grassland ecosystem and a review of comparable studies. Ecological Research. 2001;16:819–831. [Google Scholar]
- Hector A, Joshi J, Scherer-Lorenzen M, et al. Biodiversity and ecosystem functioning: reconciling the results of experimental and observational studies. Functional Ecology. 2007;21:998–1002. [Google Scholar]
- Hooper DU, Dukes JS. Overyielding among plant functional groups in a long-term experiment. Ecology Letters. 2004;7:95–105. [Google Scholar]
- Hooper DU, Chapin FS, III, Ewel JJ, et al. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs. 2005;75:3–35. [Google Scholar]
- Jolliffe PA, Wanjau FM. Competition and productivity in crop mixtures: some properties of productive crops. Journal of Agricultural Science, Cambridge. 1999;132:425–435. [Google Scholar]
- Joshi J, Schmid B, Caldeira MC, et al. Local adaptation enhances performance of common plant species. Ecology Letters. 2001;4:536–544. [Google Scholar]
- Kahmen A, Renker C, Unsicker SB, Buchmann N. Niche complementarity for nitrogen: an explanation for the biodiversity and ecosystem functioning relationship? Ecology. 2006;87:1244–1255. doi: 10.1890/0012-9658(2006)87[1244:ncfnae]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Kirwan L, Sebastià MT, Finn JA, et al. Evenness drives consistent diversity effects in an intensive grassland system across 28 European sites. Journal of Ecology. 2007;95:530–539. [Google Scholar]
- Kley G. Seed production in grass and clover species in Europe. In: Schöberlein W, Förster K, editors. Proceedings of the Third International Herbage Seed Conference. 1995. p. 12. [Google Scholar]
- Knops JMH, Tilman D, Haddad NM, et al. Effects of plant species richness on invasion dynamics, disease outbreaks, insect abundance and diversity. Ecology Letters. 1999;2:286–293. doi: 10.1046/j.1461-0248.1999.00083.x. [DOI] [PubMed] [Google Scholar]
- Lanta V, Lepš J. Effects of species and functional group richness on production in two fertility environments: an experiment with communities of perennial plants. Acta Oecologia. 2007;32:93–103. [Google Scholar]
- Lorentzen S, Roscher C, Schumacher J, Schultze E-D, Schmid B. Species richness and identity affect the use of aboveground space in experimental grasslands. Perspectives in Plant Ecology, Evolution and Systematics. 2008;10:73–87. [Google Scholar]
- Milbau A, Nijs I, Van Peer L, Reheul D, De Cauwer B. Disentangling invasiveness and invasibility during invasion in synthesized grassland communities. New Phytologist. 2003;159:657–667. doi: 10.1046/j.1469-8137.2003.00833.x. [DOI] [PubMed] [Google Scholar]
- Milbau A, Nijs I, De Raedemaecker F, Reheul D, De Cauwer B. Invasion in grassland gaps: the role of neighbourhood richness, light availability and species complementarity during two successive years. Functional Ecology. 2005;19:27–37. [Google Scholar]
- Mwangi PN, Schmitz M, Scherber C, et al. Niche pre-emption increases with species richness in experimental communities. Journal of Ecology. 2007;95:65–78. [Google Scholar]
- Palmborg C, Scherer-Lorentzen M, Jumpponen A, Carlsson G, Huss-Danell K, Högberg P. Inorganic soil nitrogen under grassland plant communities of different species composition and diversity. Oikos. 2005;110:271–282. [Google Scholar]
- Personeni E, Loiseau P. Species strategy and N fluxes in grassland soil: a question of root litter quality or rhisosphere activity? European Journal of Agronomy. 2004;22:217–229. [Google Scholar]
- Picasso VD, Brummer EC, Liebman M, Dixon PM, Wilsey BJ. Crop species diversity affects productivity and weed suppression in perennial polycultures under two management strategies. Crop Science. 2008;48:331–342. [Google Scholar]
- Pilet F, Chacon G, Forbes GA, Andrivon D. Protection of potato cultivars against late blight in mixtures increases with decreasing disease pressure. Phytopathology. 2006;96:777–783. doi: 10.1094/PHYTO-96-0777. [DOI] [PubMed] [Google Scholar]
- Rath M, Peel S. Grassland in Ireland and the UK. In: McGilloway DA, editor. Grassland: a global resource. The Netherlands: Wageningen Academic Publishers; 2005. pp. 13–27. [Google Scholar]
- Reidsma P, Tekelenburg T, van den Berg M, Alkemade R. Impacts of land-use change on biodiversity: an assessment of agricultural biodiversity in the European Union. Agriculture, Ecosystems and Environment. 2006;114:86–102. [Google Scholar]
- Richardson DM, Pyšek P. Plant invasions: merging the concepts of species invasiveness and community invasibility. Progress in Physical Geography. 2006;30:409–431. [Google Scholar]
- Roscher C, Schumacher J, Baade J, et al. The role of biodiversity for element cycling and trophic interactions: an experimental approach in a grassland community. Basic and Applied Ecology. 2003;5:107–121. [Google Scholar]
- Roscher C, Temperton VM, Scherer-Lorenzen M, et al. Overyielding in experimental grassland communities – irrespective of species pool and scale. Ecology Letters. 2005;8:419–429. [Google Scholar]
- Roscher C, Thein S, Schmid B, Scherer-Lorentzen M. Complementary nitrogen use among potentially dominant species in a biodiversity experiment varies between two years. Journal of Ecology. 2008;96:477–488. [Google Scholar]
- Rougoor C, van der Weijden W. Towards a European levy on nitrogen: a new policy tool for reducing eutrophication, acidification and climate change. Utrecht: Centre for Agriculture and Environment; 2001. [Google Scholar]
- van Ruijven J, Berendse F. Positive effects of plant species diversity on productivity in the absence of legumes. Ecology Letters. 2003;6:170–175. [Google Scholar]
- van Ruijven J, Berendse F. Diversity-productivity relationships: Initial effects, long-term patterns, and underlying mechanisms. Proceedings of the National Academy of Science of the USA. 2005;102:695–700. doi: 10.1073/pnas.0407524102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderson MA, Skinner RH, Barker DJ, Edwards GR, Tracy BF, Wedin DA. Plant species diversity and management of temperate forage and grazing land ecosystem. Crop Science. 2004;44:1132–1144. [Google Scholar]
- Sanderson MA, Soder KJ, Muller LD, Klement KD, Skinner RH, Goslee SC. Forage mixture productivity and botanical composition in pastures grazed by dairy cattle. Agronomy Journal. 2005;97:1465–1471. [Google Scholar]
- Schmid B, Joshi J, Schläpfer F. Empirical evidence for biodiversity – ecosystem functioning relationships. In: Kinzig A, Tilman D, Pacala S, editors. Functional consequences of biodiversity: experimental progress and theoretical extensions. Princeton, NJ: Princeton University Press; 2002. pp. 120–150. [Google Scholar]
- Schmid B, Hector A, Saha P, Loreau M. Biodiversity effects and transgressive overyielding. Journal of Plant Ecology. 2008;1:95–102. [Google Scholar]
- Spehn EM, Scherer-Lorenzen M, Schmid B, et al. The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen. Oikos. 2002;98:205–218. [Google Scholar]
- Temperton VM, Mwangi PN, Scherer-Lorenzen M, Schmid B, Buchmann N. Positive interactions between nitrogen-fixing legumes and four neighbouring species in a biodiversity experiment. Oecologia. 2007;151:190–205. doi: 10.1007/s00442-006-0576-z. [DOI] [PubMed] [Google Scholar]
- Tilman D, Reich PB, Knops J, Wedin D, Mielke T, Lehman C. Diversity and productivity in a long-term grassland experiment. Science. 2001;294:843–845. doi: 10.1126/science.1060391. [DOI] [PubMed] [Google Scholar]
- Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S. Agricultural sustainability and intensive production practices. Nature. 2002;418:671–677. doi: 10.1038/nature01014. [DOI] [PubMed] [Google Scholar]
- Tracy BF, Sanderson MA. Productivity and stability relationships in mowed pasture communities of varying species composition. Crop Science. 2004;a 44:2180–2186. [Google Scholar]
- Tracy BF, Sanderson MA. Forage productivity, species evenness and weed invasion in pasture communities. Agriculture, Ecosystems and Environment. 2004;b 102:175–183. [Google Scholar]
- Tracy BF, Renne IJ, Gerrish J, Sanderson MA. Effects of plant diversity on invasion of weed species in experimental pasture communities. Basic and Applied Ecology. 2004;5:543–550. [Google Scholar]
- Verbeke G, Molenbergh G. Linear mixed models for longitudinal data. New York, NY: Springer-Verlag; 2000. Springer Series in Statistics. [Google Scholar]
- Williams TA, Abberton MT, Rhodes I. Performance of white clover varieties combined in blends and alone when grown with perennial ryegrass under sheep and cattle grazing. Grass and Forage Science. 2003;58:90–93. [Google Scholar]