

Abstract
Coral bleaching, during which corals lose their symbiotic dinoflagellates, typically corresponds with periods of intense heat stress, and appears to be increasing in frequency and geographic extent as the climate warms. A fundamental question in coral reef ecology is whether chronic local stress reduces coral resistance and resilience from episodic stress such as bleaching, or alternatively promotes acclimatization, potentially increasing resistance and resilience. Here we show that following a major bleaching event, Montastraea faveolata coral growth rates at sites with higher local anthropogenic stressors remained suppressed for at least 8 years, while coral growth rates at sites with lower stress recovered in 2–3 years. Instead of promoting acclimatization, our data indicate that background stress reduces coral fitness and resilience to episodic events. We also suggest that reducing chronic stress through local coral reef management efforts may increase coral resilience to global climate change.
Introduction
Ecological studies have demonstrated that stressors currently affecting coral reefs include, among others, coral diseases [1], over-fishing [2], and a combination of pollution and sedimentation from coastal development [3]. These chronic stressors are often associated with the gradual loss of coral cover and overgrowth by fleshy algae. However, abrupt and severe episodic events, such as coral bleaching, may also be responsible for coral reef degradation [4]. An outstanding issue is whether the combination of multiple stressors reduces coral resistance or resilience to episodic events such as bleaching [5]–[7], or alternatively whether acclimatization to stressful conditions can increase coral resistance—the ability of corals to withstand future stress [8], [9]. Bleaching is a generalized term for the loss of symbiotic dinoflagellate zooxanthellae or their pigments in scleractinian corals and is typically associated with sustained, unusually warm water temperatures [10]. Several studies have found that bleaching reduces skeletal growth in corals [11]–[13]. Ocean acidification may also reduce the ability of corals to calcify as normal; a recent study of corals from the Great Barrier Reef attributes a 14.2% decrease in calcification since 1990 to a combination of acidification and warming [14]. Here we define resistance as an individual coral's ability to continue normal skeletal growth even under stress (whether chronic or episodic), and resilience as a coral's ability to recover to normal growth rates after a stressful event (Fig. 1a–c). To test our hypothesis that chronic local stress reduces coral resistance and resilience to bleaching, we focus on coral growth before and after the 1998 mass-bleaching event [15] from four sites on the Mesoamerican Reef (Fig. 2, Table 1) with relatively high and low chronic local stress.
Figure 1. X-radiographs of various coral cores showing the different types of growth behavior discussed.

(A) Coral without the 1998 growth suppression, indicating resistance to bleaching in 1998. (B) Coral with the 1998 growth suppression, recognized by the bright high-density band, but with a quick return to pre-1998 extension rates, indicating resilience after bleaching. (C) Coral with the 1998 growth suppression and continuing depressed extension rates after 1998, indicating a lack of both resistance and resilience to bleaching. (D) A coral with relatively high average extension rate. (E) A coral with relatively low average extension rate. (F) A coral with a partial mortality scar on the left (noted by white arrow), coincident with the 1998 growth anomaly.
Figure 2. Map of the Mesoamerican Reef with locations of coral collections as black circles.

Dark grey denotes coral, light grey denotes land areas. T1, T2 = Turneffe Atoll (4 cores from T1, 13 from T2), S = Sapodilla Cayes (44 cores), U = Utila (17 cores), C = Cayos Cochinos (14 cores).
Table 1. Coral core collection site locations.
| Site | Dive Site Name | Coordinates | Total Cores | 1998 Stress Band | Partial Mortality in 1998 | Previous Stress Bands |
| Turneffe 1 | Dog Flea Caye | 17°29′59″N, 87°45′30″W | 17 | 71% (12) | 6% (1) | 2 |
| Turneffe 2 | Harry Jones | 17°18′25″N, 87°48′04″W | ||||
| Sapodilla | Frank's Caye, NE buoy | 16°07′45″N, 88°14′59″W | 44 | 100% (44) | 16% (7) | 0 |
| Utila | Diamond Caye | 16°03′52″N, 86°57′30″W | 17 | 100% (17) | 12% (2) | 1 |
| Cayos Cochinos | Pelican Point, Peli 2 | 15°58′41″N, 86°29′06″W | 14 | 100% (14) | 21% (3) | 0 |
| Total | 92 | 95% (87) | 14% (13) | 3 |
Location names with dive site name or nearby caye, coordinates, number of cores from each site, along with growth anomalies in 1998 and earlier. Table lists the total number of cores which were drilled and slabbed along the growth axis, the percentage and number of these that have dense stress bands associated with the 1998 event, the percentage and number that contained a partial mortality scar, and any previous individual stress bands.
Methods
The level of local stress at each site was calculated using the methodology of Halpern et al. (2008) [16] to estimate a cumulative impact index based on weighted, log-transformed, re-scaled data representing (1) sedimentation (2) nutrient input (3) local human population size adjacent to our sites and (4) a relative measure of fishing pressure based on fish abundance surveys [17, supplementary information Text S1] (Table 2). Our calculated local impact scores for the Mesoamerican reef sites have the same ranking as the “Halpern Index” that we calculated as the average of 4 cells representing our sites from the Halpern et al. (2008) [16] global impact map. We also compared our results with the integrated reef health index at our sites from the Healthy Reefs Report Card [17] (Table 2). According to all three indices of local stressors and reef health, our sites in the Sapodilla Cayes and Utila experience the highest levels of local stress, while sites in Cayos Cochinos and Turneffe Atoll experience lower levels.
Table 2. Anthropogenic stress scores.
| Site | Sedimentation | Nutrients | Human Population | Fishing Pressure | Stress Index | Halpern Index | Healthy Reefs Index* |
| Sapodilla | 1.60 | 1.80 | 0.00 | 0.00 | 3.40 | 13.35 | 2.10 |
| Utila | 0.91 | 0.56 | 2.30 | −2.08 | 1.70 | 13.27 | 2.49 |
| Cayos Cochinos | 1.37 | 0.89 | 1.00 | −2.41 | 0.84 | 11.06 | 2.98 |
| Turneffe | 0.00 | 0.00 | 0.31 | −0.11 | 0.20 | 9.80 | 2.59 |
Four measures of local stress at each site: sedimentation, nutrients, human population and fishing pressure used to calculate our cumulative “Stress Index” (sources in supplemental information). A comparison with the Halpern et al. (2008) [15] and Healthy Reefs [16] indices is also shown, though note that only the relative rankings are comparable. Sapodilla and Utila experience higher local impacts and lower reef health. *Note that while low Stress Index and Halpern Index scores indicate fewer local stressors, a low Healthy Reefs Index score indicates worse reef health.
We collected a total of 92 coral cores from Montastraea faveolata, the dominant reef-builder on the fore reef [18] between 2.5–13 m depth in spur and groove habitat (Table 1). Permission to collect and export coral samples was granted by the Belize Fisheries Department and the Secretaria de Agricultura y Ganadería, Honduras. Coral skeletal growth rates were measured based on annual density bands in the skeleton [19], supplementary information Text S1. Coral extension, average density, and calcification (the product of extension and density) were analyzed from digital x-rays using the program CoralXDS [20] for each high-density, low-density annual couplet, though in this paper we focus on extension rates (cm/year). To ensure all growth-increment chronologies were annually resolved, we made novel use of tree-ring techniques, including “crossdating” of annual extension rate (growth-increment width). Average extension rates and 95% confidence intervals were calculated using 10,000 bootstrapped samples (Fig. 3). Differences between sites were compared using pairwise permutation tests on extension rates between 1950–1997. Edinger et al. (2000) [21] found that comparing absolute extension rates between sites is not necessarily representative of overall reef health. Indeed, Turneffe Atoll, one of the less impacted sites, has significantly (p<0.001) higher extension rates than the other three sites before 1998, but this site also lost significant coral cover due to the 1998 bleaching event and hurricane Mitch (Fig. S1). Therefore, here we focus on resistance to and recovery after bleaching in 1998 instead of absolute differences between sites. Recovery to normal mean extension rates after 1998 were estimated, while controlling for the year, by fitting the generalized linear model (GLM): (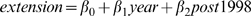 ) (Table S1). Extension rates from the years 1950 to 1997 (post1998 = 0) were compared to rates from 2002 to 2006 (post1998 = 1) to avoid the years 1998–2001 during which all sites had severely depressed growth.
) (Table S1). Extension rates from the years 1950 to 1997 (post1998 = 0) were compared to rates from 2002 to 2006 (post1998 = 1) to avoid the years 1998–2001 during which all sites had severely depressed growth.
Figure 3. Bootstrapped means (solid lines) and 95% confidence intervals (shading) for extension rates after 1955.

Extension rates at Sapodilla and Utila remain suppressed after the 1998 bleaching event (p<0.001).
Results and Discussion
At all four sites, nearly every coral displays a visually prominent stress band in 1998 [11] (Fig. 1b), indicated by an increase in skeletal density and decrease in extension and calcification rates more than four standard deviations outside the mean chronology. Indeed, 95% of coral cores show a marked reduction in extension rates that persists for two years or more following the 1998 event (Fig. 3, Table 1). Additional rare stress bands (denser than the long-term mean by at least 1.5 standard deviations) are seen in three individual coral cores in 1950, 1965 and 1995. However, no other coral cores displayed significant changes in growth during these years, nor did we observe any partial mortality scars before 1998.
Although the 1998 bleaching event affected the entire Mesoamerican reef system, there are spatial differences in its intensity revealed in our coral cores. We compared the number of coral cores with partial mortality scars (Fig. 1f) and 1998 stress banding between sites using a permutation test. Turneffe Atoll, the site with the lowest level of local stress, is also the only site without ubiquitous stress banding (p<0.05) (Fig. 1a), and has the lowest, but non-significant, frequency of partial mortality scars (Table 2). The lack of stress banding in some coral cores at Turneffe Atoll suggests that at least some of those corals may have resisted bleaching. By comparison, the universal occurrence of stress banding at all other sites supports the hypothesis that high chronic stress decreases coral resistance to bleaching.
In contrast to the modest variability in resistance to the 1998 bleaching event, there are large differences in the resilience of corals between sites. At Sapodilla and Utila where stress indices are high, coral growth still did not recover completely by the time of collection more than eight years following the 1998 bleaching event, even controlling for long-term decreasing trends at these sites (GLM: Sapodilla, F(2,50) = 20.70, p<0.001, R2 = 0.45; Utila, F(2,50) = 18.26, p<0.001, R2 = 0.42, Table S1). Such a long recovery period is unprecedented in the literature, with most studies reporting growth suppression due to bleaching on the order of one year [11]–[13] and the longest growth suppression reported for four years [22]. In comparison, corals from lower-stress sites (Turneffe and Cayos Cochinos) recovered to pre-bleaching extension rates in about three years (Fig. 3, Table S1). However, at sites with high local stress, corals have been unable to recover to pre-disturbance growth rates.
The between-site differences in both the level of impact of the 1998 bleaching event and the subsequent recovery time are not easily explained by between-site differences in the strength of the 1998 bleaching event. We tested the hypothesis that our sites experienced differences in heat stress in 1998 by calculating the degree-heating-weeks (DHW) [23] for our four study areas from 7-day composite night time sea surface temperature data [supplementary information Table S1]. Our findings indicate that during 1998, heat stress was higher at Cayos Cochinos and the Sapodilla Cayes (6.84 and 5.54 maximum DHW, respectively), compared to Utila and Turneffe Atoll (3.34 and 2.27 maximum DHW, respectively). However, while lower temperature stress may help explain the lower stress banding at Turneffe it cannot explain the lack of resilience at Utila or the higher resilience in Cayos Cochinos (which experienced the highest heat stress).
We also examined the possibility that a hurricane strike may have had different effects across the Mesoamerican Reef. Category 5 hurricane Mitch (October 21–29, 1998) produced extreme runoff over most of the southern portion of the Mesoamerican Reef and reduced water clarity for several weeks [24]. However, we have found no geochemical signature associated with runoff from Mitch, even in coral cores analyzed at extremely high resolution using laser ablation [25]. The lack of signal indicates the corals stopped calcifying due to the bleaching event (August 1998) prior to the hurricane. While poor water quality could explain the subsequent low resiliency of corals at our southern sites, it is notable that corals at the most southerly site, Cayos Cochinos, recover to pre-1998 growth rates just as rapidly as corals at Turneffe where the runoff impact of the hurricane was low. In addition, earlier hurricane strikes have left almost no record in our coral growth rate data suggesting that their overall impact has been low on Montastraea faveolata.
We conclude that the large differences in chronic stress between our sites are responsible for differences in coral resilience following exposure to the 1998 bleaching event. To date the 1998 bleaching event remains the most significant bleaching event recorded on the Mesoamerican reef, as the 2005 event was significantly less severe than in other parts of the Caribbean [26]. Our data do not support the hypothesis that exposure to stress might help coral colonies acclimatize and therefore resist bleaching. Instead, it is clear that coral colonies experiencing higher local stress before 1998 were more severely affected by bleaching and recovered more slowly than those exposed to lower chronic stress. Possibly, the acclimatization hypothesis is only applicable for the same stressor or for lower levels of stress than Sapodilla and Utila experience, and the multi-species coral community may exhibit acclimatization patterns different from the individual coral colony response measured in this study. For example, repetitive bleaching may increase a coral's ability to withstand future heat stress [8], [9], but other local stressors such as increased sedimentation may depress a coral's energy reserves [27], making it less likely to survive or recover from a bleaching event [28]. Even if acclimatization can occur in some cases, the differential responses of M. faveolata across various stress regimes indicate that local conservation efforts that reduce stress, such as reducing runoff by replanting mangroves at the coast or protecting an area from overfishing, could have significant impacts on the ability of corals to withstand the effects of climate change. Future research could investigate whether this interaction between local and global stressors extends to other coral species.
Supporting Information
Detailed Materials and Methods as well as a supplementary table of the outcome of our statistical test for growth rate recovery after bleaching, and a supplemental figure showing coral cover before and after 1998.
(0.06 MB DOC)
Supplemental figure of coral cover before and after 1998.
(3.79 MB TIF)
Recovery of extension rates after 1998.
(0.03 MB DOC)
Acknowledgments
We thank M. Roldan for x-raying the cores repeatedly, and M. Carilli, B. Erisman, K. Hughen, E. Kisfaludy, N. Prouty, J. Shrives, A. Stumpf, and A. Young for help in the field. We thank the Belizean and Honduran governments for supplying permits and World Wildlife Fund Belize, The University of Belize, Reef Conservation International, The Honduran Coral Reef Foundation, and Operation Wallacea for logistical support.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: This research was supported by the PADI Foundation, B. Katz, and two anonymous donors. The funders (including the anonymous donors) had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Harvell CD, Kim K, Burkholder JM, Colwell RR, Epstein PR, et al. Emerging marine diseases: Climate links and anthropogenic factors. Science. 1999;285:1505–1510. doi: 10.1126/science.285.5433.1505. [DOI] [PubMed] [Google Scholar]
- 2.Hughes TP. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science. 1994;265:1547–1551. doi: 10.1126/science.265.5178.1547. [DOI] [PubMed] [Google Scholar]
- 3.Maragos JE. Impact of coastal construction on coral reefs in the US-affiliated Pacific islands. Coast Manage. 1993;21:235–269. [Google Scholar]
- 4.Knowlton N. The future of coral reefs. Proc Natl Acad Sci. 2001;98:5419–5425. doi: 10.1073/pnas.091092998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hughes TP, Baird AH, Bellwood DR, Card M, Connolly SR, et al. Climate change, human impacts, and the resilience of coral reefs. Science. 2003;301:929–933. doi: 10.1126/science.1085046. [DOI] [PubMed] [Google Scholar]
- 6.Knowlton N, Jackson JBC. Shifting baselines, local impacts, and global change on coral reefs. PLoS Biol. 2008;6:215–220. doi: 10.1371/journal.pbio.0060054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sandin SA, Smith JE, DeMartini EE, Dinsdale EA, Donner SD, et al. Baselines and degradation of coral reefs in the northern Line Islands. PLoS ONE. 2008;3:e1548. doi: 10.1371/journal.pone.0001548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brown BE, Dunne RP, Goodson MS, Douglas AE. Experience shapes the susceptibility of a reef coral to bleaching. Coral Reefs. 2002;21:119–126. [Google Scholar]
- 9.Castillo KD, Helmuth BST. Influence of thermal history on the response of Montrastrea annularis to short-term temperature exposure. Mar Biol. 2005;148:261–270. [Google Scholar]
- 10.Hoegh-Guldberg O. Climate change, coral bleaching, and the future of the world's coral reefs. Mar Freshw Res. 1999;50:839–866. [Google Scholar]
- 11.Leder JJ, Szmant AM, Swart PK. The effect of prolonged “bleaching” on skeletal banding and stable isotopic composition in Montastrea annularis. Coral Reefs. 1991;10:19–27. [Google Scholar]
- 12.Suzuki A, Gagan MK, Fabricius K, Isdale PJ, Yukino I, et al. Skeletal isotope microprofiles of growth perturbations in Porites corals during the 1997–1998 mass bleaching event. Coral Reefs. 2003;22:357–369. [Google Scholar]
- 13.Goreau T, Macfarlane AH. Reduced growth rate of Montastrea annularis following the 1987–88 coral-bleaching event. Coral Reefs. 1990;8:211–215. [Google Scholar]
- 14.De'ath G, Lough JM, Fabricius KF. Declining coral calcification on the Great Barrier Reef. Science. 2009;323:116–120. doi: 10.1126/science.1165283. [DOI] [PubMed] [Google Scholar]
- 15.Kramer P, Kramer PR, Arias-Gonzalez E, McField M. Status of coral reefs of northern Central America: Mexico, Belize, Guatemala, Honduras, Nicaragua and El Salvador. In: Wilkinson C, editor. Status of Coral Reefs of the World: 2000. Townsville: Australian Institute for Marine Science; 2000. [Google Scholar]
- 16.Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, et al. A Global map of human impact on marine ecosystems. Science. 2008;319:948–952. doi: 10.1126/science.1149345. [DOI] [PubMed] [Google Scholar]
- 17.Healthy Reefs Initiative. Eco-health Report Card for the Mesoamerican Reef. An Evaluation of Ecosystem Health. 2008 http://www.healthyreefs.org. [Google Scholar]
- 18.McField MD. Influence of disturbance on coral reef community structure in Belize. Proc 9th Int Coral Reef Symp. 2000;1:62–68. [Google Scholar]
- 19.Knutson RA, Buddemeier RW, Smith SV. Coral chronometers: seasonal growth bands in reef corals. Science. 1972;177:270–212. doi: 10.1126/science.177.4045.270. [DOI] [PubMed] [Google Scholar]
- 20.Helmle KP, Kohler KE, Dodge RE. Coral X-radiography Densitometry System. 2002. http://www.nova.edu/ocean/coralxds/index.html.
- 21.Edinger E, Limmon GV, Jompa J, Widjatmoko W, Heikoop JM, et al. Normal coral growth rates of dying reefs: Are coral growth rates good indicators of reef health? Mar Poll Bull. 2000;40:404–425. [Google Scholar]
- 22.Omata T, Suzuki A, Kawahata H, Nojima S, Minoshima K, et al. Oxygen and carbon stable isotope systematics in Porites coral near its latitudinal limit: The coral response to low-thermal temperature stress. Global Planet Change. 2006;53:137–146. [Google Scholar]
- 23.Strong A, Barrientos CS, Duda C, Sapper J. Improved satellite techniques for monitoring coral reef bleaching. Proc 8th Int Coral Reef Symp. 1997;2:1495–1498. [Google Scholar]
- 24.Andréfouët S, Mumby PJ, McField M, Hu C, Muller-Karger FE. Revisiting coral reef connectivity. Coral Reefs. 2002;21:43–48. [Google Scholar]
- 25.Koenig A, Carilli J, Buster N. A comparison of chemical records in Montastraea faveolata and Siderastrea siderea as examined by laser ablation ICP-MS: A look at methodology for Caribbean coral for seasonal and storm events. in prep.
- 26.McField M, Bood N, Fonseca A, Arrivillaga A, Franquesa Rinos A, Loreto Viruel RM. Status of the Mesoamerican Reef after the 2005 Coral Bleaching Event. In: Wilkinson C, Souter D, editors. Status of Caribbean Coral Reefs after Bleaching and Hurricanes in 2005. Townsville, Australia: Global Coral Reef Monitoring Network; 2005. [Google Scholar]
- 27.Rogers C. Responses of coral reefs and reef organisms to sedimentation. Mar Ecol Prog Ser. 1990;62:185–202. [Google Scholar]
- 28.Rodrigues L, Grottoli AG. Energy reserves and metabolism as indicators of coral recovery from bleaching. Limnol Oceanog. 2007;52:1874–1882. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Detailed Materials and Methods as well as a supplementary table of the outcome of our statistical test for growth rate recovery after bleaching, and a supplemental figure showing coral cover before and after 1998.
(0.06 MB DOC)
Supplemental figure of coral cover before and after 1998.
(3.79 MB TIF)
Recovery of extension rates after 1998.
(0.03 MB DOC)