

Abstract
AIM: To determine the anti-inflammatory activity of probiotic Bifidobacteria in Bifidobacteria-fermented milk (BFM) which is effective against active ulcerative colitis (UC) and exacerbations of UC, and to explore the immunoregulatory mechanisms.
METHODS: Peripheral blood mononuclear cells (PBMNC) from UC patients or HT-29 cells were co-cultured with heat-killed probiotic bacteria or culture supernatant of Bifidobacterium breve strain Yakult (BbrY) or Bifidobacterium bifidum strain Yakult (BbiY) to estimate the amount of IL-10 or IL-8 secreted.
RESULTS: Both strains of probiotic Bifidobacteria contained in the BFM induced IL-10 production in PBMNC from UC patients, though BbrY was more effective than BbiY. Conditioned medium (CM) and DNA of both strains inhibited IL-8 secretion in HT-29 cells stimulated with TNF-α, whereas no such effect was observed with heat-killed bacteria. The inhibitory effect of CM derived from BbiY was greater than that of CM derived from BbrY. DNAs of the two strains had a comparable inhibitory activity against the secretion of IL-8. CM of BbiY induced a repression of IL-8 gene expression with a higher expression of IκB-ζ mRNA 4 h after culture of HT-29 cells compared to that in the absence of CM.
CONCLUSION: Probiotic Bifidobacterium strains in BFM enhance IL-10 production in PBMNC and inhibit IL-8 secretion in intestinal epithelial cells, suggesting that BFM has anti-inflammatory effects against ulcerative colitis.
Keywords: Bifidobacterium, Ulcerative colitis, Anti-inflammatory, Interleukin-10, Interleukin-8
INTRODUCTION
Ulcerative colitis (UC) is assumed to be a result of an impaired intestinal immune response to intestinal environmental antigens such as ubiquitous microflora[1,2]. There is evidence that probiotic treatment is effective against UC[3–6]. We have previously shown that treatment with bifidobacteria-fermented milk (BFM) containing live bifidobacteria is more effective than the usual treatment in inducing[7] and maintaining remission[8] of ulcerative colitis. An improvement in the composition of intestinal flora by BFM may prevent the overgrowth of potentially pathogenic bacteria such as Bacteroides vulgatus, but we hypothesized that probiotic strains of bifidobacteria in BFM may directly interfere with the host signaling events that drive the intestinal inflammatory response. In inflammatory bowel disease, IL-10 is a cytokine of particular therapeutic interest since it has been shown in animal models that interleukin (IL)-10(-/-) mice spontaneously develop intestinal inflammation. IL-10 plays a key role in the control of inflammatory responses to enteric organisms[9–12]. More recently, it has been shown in animal models that probiotic strains displaying an in vitro potential to induce higher levels of the anti-inflammatory cytokine IL-10 and lower levels of the inflammatory cytokine IL-12, offer the best protection against in vivo colitis in the model[13]. A genetically engineered Lactococcus lactis thy12 producing IL-10 ameliorated colitis in two models of experimental colitis, providing proof of principal that topically delivers IL-10, can be therapeutically efficacious[14] and a recent proof-or-principle experiment using this transgenic bacterium expressing IL-10 in 10 patients with Crohn’s disease showed efficacy[15]. In addition, there is an increasing amount of evidence to suggest that the potent neutrophil chemoattractant, IL-8, has an important role in the pathogenesis of inflammatory bowel disease (IBD)[16–19]. Recently, a higher concentration of IL-8 was found in more histologically inflamed tissue segments from pediatric IBD patients, suggesting that IL-8-specific therapies may universally modify the inflammatory activity in IBD patients[20]. In this study, we focused on the effect of probiotic strains on the secretion of IL-10 by peripheral blood mononuclear cells and also the production of IL-8 by intestinal epithelial cells.
MATERIALS AND METHODS
Bacteria and related preparations
Bifidobacterium bifidum strain Yakult (BbiY) and Bifidobacterium breve strain Yakult (BbrY) were grown in MRS broth (Becton, Dickinson and Company, Sparks, MD). Heat-killed BbiY or BbrY was prepared by heating bacteria resuspended in distilled water at 100°C for 30 min, and then lyophilized[21]. For the preparation of conditioned medium (CM), bacteria grown in MRS broth were collected by centrifugation and cultured over 16 h in RPMI-1640 medium (Sigma-Aldrich, St Louis, MO) containing 10% fetal calf serum (FCS) and 2% lactose, then centrifuged[22]. The supernatant was filtrated on a 0.22 μm membrane and neutralized with sodium hydroxide. To characterize the active component in CM, it was separated into fractions of more than and less than 3 kDa through Centricon YM-3 (Millipore, Bedford, MA), adjusted to the initial volume, and then to heat treatment at 100°C for 15 min[23]. DNA was isolated using the method of Yuki[24] with a slight modification. Briefly, bacterial cells were suspended in Tris-EDTA buffer (pH 8.0) containing 0.5 mol/L sucrose and treated with N-acetylmuramidase SG (Seikagaku Corp., Tokyo, Japan) and lysozyme (Sigma-Aldrich) at 37°C for 1 h. The cells were lysed by addition of sodium dodecyl sulfate and proteinase K (Sigma-Aldrich) followed by a 60-min incubation at 65°C. Deproteinization was done by extraction with Tris-saturated phenol and phenol/chloroform/isoamyl alchol (25:24:1). Finally, DNA was precipitated by ethanol.
Peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMNC) were isolated from peripheral blood of UC patients by Ficoll-Conray (Lymphosepar I; Immuno-Biological Laboratories, Takasaki, Japan) density gradient centrifugation. Table 1 summarizes the patient characteristics. All 9 patients (outpatient) had active UC which was moderate or mild (1 mild, 8 moderate) according to the criteria of Truelove & Witts[25]. All patients received a standard therapeutic regimen consisting of oral 5-ASA (mesalazine) and five of the 9 patients with active UC took a low dose of oral predonine. Cells (2 × 105) were cultured with heat-killed bacteria (10 μg/mL) in 200 μL of AIM-V medium (Invitrogen Corp., Carlsbad, CA) in a flat-bottomed 96-well culture plate (Nunc, Roskilde, Denmark) for 48 h. Supernatant was collected and frozen until cytokine levels were quantified. In each assay, a positive control with lipopolysaccharide (LPS, 10 μg/mL) added to PBMNC and a negative control with no stimuli were included.
Table 1.
Characteristics of the patients with ulcerative colitis
| Patient No. | Age | Sex | Disease extent | Clinical pattern | Disease duration (yr) | Disease severity |
Treatment |
||
| 5-ASA (mesalazine) | (SASP) | Steroid (predonine) | |||||||
| 1 | 23 | Male | Total | Relapsing | 6 | Moderate | + | ||
| 2 | 38 | Female | Total | Relapsing | 2 | Moderate | + | + | |
| 3 | 32 | Male | Total | Relapsing | 2 | Moderate | + | + | |
| 4 | 36 | Female | Proctitis | Relapsing | 15 | Mild | + | ||
| 5 | 23 | Male | Total | First | 3 | Moderate | + | ||
| 6 | 20 | Male | Total | Relapsing | 2 | Moderate | + | + | |
| 7 | 25 | Male | Total | First | 0.5 | Moderate | + | ||
| 8 | 42 | Male | Left-sided | Chronic | 7 | Moderate | + | + | |
| 9 | 36 | Male | Left-sided | Relapsing | 14 | Moderate | + | + | |
SASP: Salazosulphapyridine.
Quantification of cytokine levels in culture supernatant
IL-10 and IL-8 levels in the culture supernatant were determined by sandwich enzyme-linked immunosorbent assay (ELISA). IL-10 was detected using anti-human IL-10 antibody (51-26171E, BD Biosciences PharMingen, Franklin Lakes, NJ) and biotinylated antibody (51-26172E, BD Biosciences PharMingen). IL-8 was detected using anti-human IL-8 antibody (AHC0932, Invitrogen BioSource) and biotinylated anti-human IL-8 antibody (AHC0789, Invitrogen BioSource).
HT-29 cell culture
HT-29 cells were cultured at 37°C in an atmosphere containing 50 mL/L CO2 in RPMI-1640 medium containing 10% FCS, 1 mmol/L sodium pyruvate (Invitrogen) and 10 mmol/L 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES). Confluent monolayers were incubated in 96-well or 24-well tissue culture plates with human TNF-α (10 ng/mL, PeproTech, London, UK).
Quantitative RT-PCR
Total RNA was isolated from HT-29 cells using the TRIzol reagent (Invitrogen) according to the manufacturer’s instructions. The RNA was reverse transcribed into cDNA using a SuperScript First-Strand Synthesis System kit (Invitrogen). Real-time quantitative PCR was performed in an ABI prism 7500 Real Time PCR System (Applied Biosystems, Foster City, CA) with the SYBR Premix Ex Taq (Takara Bio, Shiga, Japan). The following primers were used to amplify cDNA fragments: IL-8: forward: 5’-ACACTGCGCCAACACAGAAATTA-3’, reverse: 5’-TTTGCTTGAAGTTTCACTGGCATC-3’; GAPDH: forward: 5’-GCACCGTCAAGGCTGAGAAC-3’, reverse: 5’-ATGGTGGTGAAGACGCCAGT-3’; IκB-ζ: forward: 5’-ACGCGCAAACATGAGTCCAG-3’, reverse: 5’-CTCAGCAGCAGCAACAGCATC-3’. All results were finally determined after correction with GAPDH expression.
Statistical analysis
Results were expressed as mean ± SD. Differences in mean values between groups were analyzed with Student’s t-test.
RESULTS
Effect of probiotic bacteria on IL-10 secretion in PBMNC
The secretion of IL-10 increased in all the cultures of PBMNC isolated from the nine UC patients with a historical record of the treatment as shown in Table 1 in the presence of heat-killed BbrY or BbiY, compared to the basal secretion in the absence of probiotic bacteria (Figure 1). BbrY induced significantly higher levels of IL-10 than BbiY in 8 out of the 9 PBMNC preparations. Incubation with BbrY also elicited significantly higher levels of IL-10 than that with LPS (10 μg/mL) in 6 out of the 9 PBMNC preparations.
Figure 1.
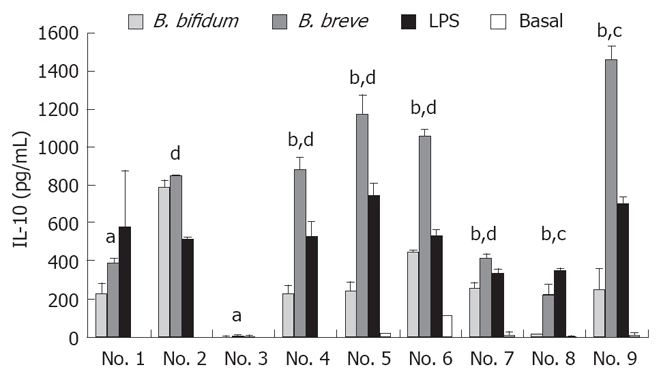
Effects of probiotic bifidobacteria on IL-10 production in PBMNC. PBMNC were isolated from 9 ulcerative-colitis patients and incubated with heat-killed probiotic BbiY or BbrY (10 μg/mL), or LPS. At forty-eight hours after incubation, the IL-10 concentration was determined by ELISA (mean ± SD, n = 3). aP < 0.05, bP < 0.01 vs BbiY; cP < 0.05, dP < 0.01 LPS.
Effect of probiotic Bifidobacterium on IL-8 secretion in TNF-α-stimulated HT-29 cells
HT-29 cells were incubated with TNF-α for six hours in the presence or absence of heat-killed BbiY and BbrY. Neither of the heat-killed probiotic bacteria had an effect on the secretion of IL-8 at the concentration ranging from 1 μg/mL-100 μg/mL (Figure 2).
Figure 2.
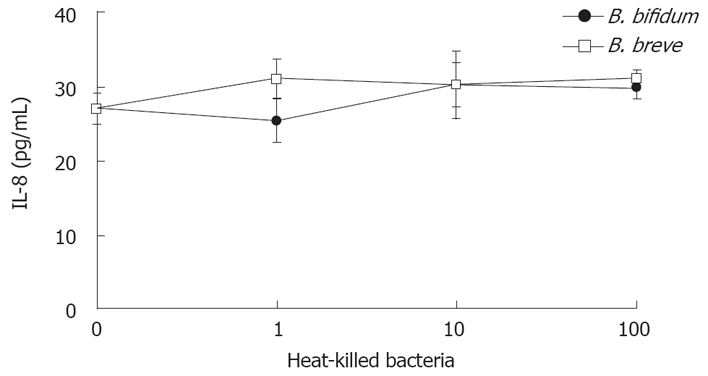
Effects of probiotic bifidobacteria on IL-8 production in HT-29 cells. HT-29 cells were stimulated with TNF-α (10 ng/mL) in the absence or presence of various concentrations of probiotic bacteria. Six hours after incubation, the IL-8 concentration was determined by ELISA (mean ± SD, n = 4).
Next, we examined the effect of conditioned medium (CM) on IL-8 secretion by HT-29 cells. Both CMs of BbiY and BbrY inhibited TNF-α-induced secretion of IL-8 in a dose-dependent manner (Figure 3). Concentrations of BbiY and BbrY cultured over 16 h in RPMI-1640 medium were 3.6 × 108 CFU/mL and 2.2 × 108 CFU/mL, respectively.
Figure 3.
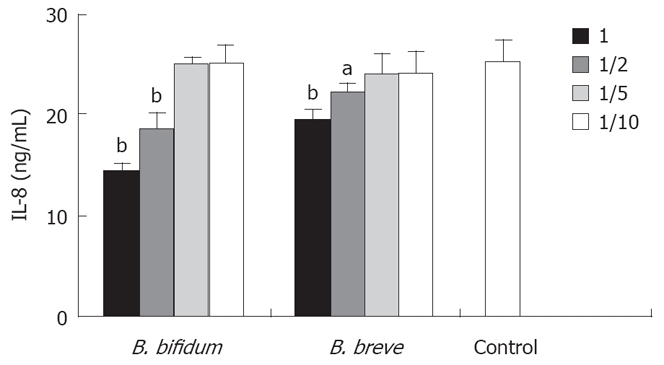
Effects of probiotic bifidobacteria-derived CMs on IL-8 production in HT-29 cells. HT-29 cells were stimulated with TNF-α in the absence or presence of graded dilutions of CMs. CMs were prepared as described in the Methods. Other experimental conditions are shown in Figure 2. aP < 0.05, bP < 0.01 vs control.
Nature of the inhibitory effect on IL-8 secretion in probiotic-derived CM
To investigate the nature of this soluble factor, BbiY-derived CM was subjected to molecular sieve and heat treatment. The fractions of less than and more than 3 kDa both retained the inhibitory activity toward the secretion of IL-8 in HT-29 cells, though the former fraction was greater than the latter one (Figure 4). The inhibitory effect of the latter fraction but not the former one was diminished by heat-treatment (data not shown). We checked the effect of acetic and lactic acid with their major constituents in less than 3 kDa fraction. Acetic acid but not lactic acid inhibited the IL-8 secretion in HT-29 cells, when added at the same concentration as in the CM, 11 mmol/L and 4 mmol/L, respectively (Figure 5).
Figure 4.
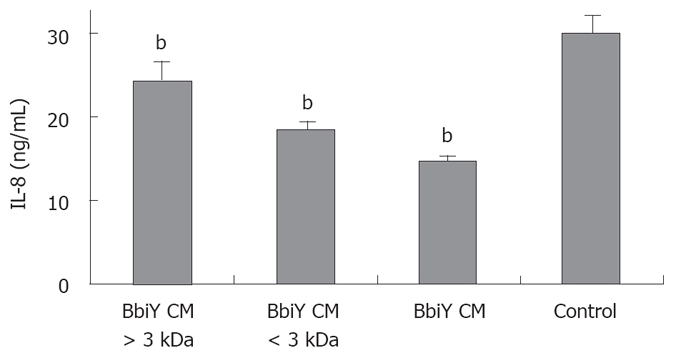
Assessment of the molecular weight of active component in probiotic-derived CMs. CM was separated into fractions of more than and less than 3 kDa, adjusted to the initial volume. The inhibitory effect on the secretion of IL-8 was determined as described in Figure 2. bP < 0.01 vs control.
Figure 5.
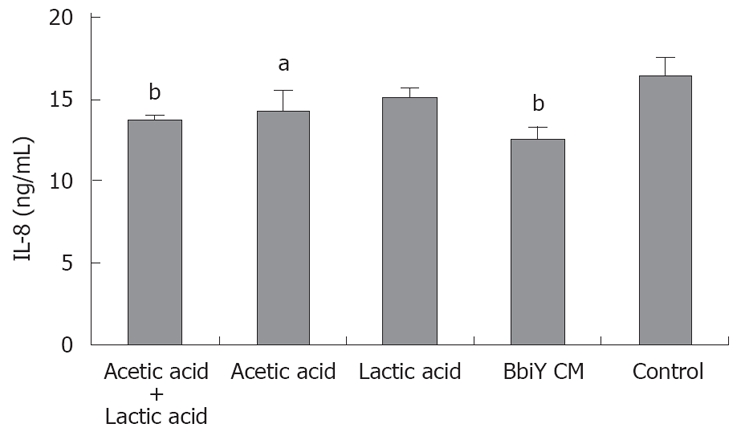
Effects of acetic acid and/or lactic acid on IL-8 secretion in HT-29 cells. HT-29 cells were incubated with approximately the same concentration of acetic acid (11 mmol/L) and/or lactic acid (4 mmol/L) to that of CMs. The inhibitory effect on the secretion of IL-8 was determined as described in Figure 2. aP < 0.05, bP < 0.01 vs control.
IL-8 and IκB-ζ expression during incubation with probiotic bacteia in HT-29 cells
MRNA levels of IL-8 and IκB-ζ were determined in TNF-α-stimulated HT-29 cells in the presence or absence of BbiY-derived CM (Figure 6). A repression of IL-8 expression occurred with an up-regulation of IκB-ζ expression 4 h after incubation of HT-29 cells with probiotic-CM.
Figure 6.

IL-8 and IκB-ζ mRNA expression during the culture of HT-29 cells in the presence of probitic-derived CMs. The mRNA expression was quantified as described in Materials and Methods. Results are representative of three separate experiments.
Probiotic DNA inhibited epithelial secretion of IL-8
HT-29 cells were stimulated with TNF-α in the presence or absence of bacterial DNA isolated from BbiY or BbrY. DNA from both BbiY and BbrY significantly inhibited the IL-8 secretion in HT-29 cells, at the concentration ranging from 1 μg/mL-100 μg/mL, while DNA from L. acidophilus (YIT 0168) had no effect (Figure 7A). Treatment of probiotic DNAs with DNase abolished their inhibitory effects on the secretion of IL-8 (Figure 7B).
Figure 7.
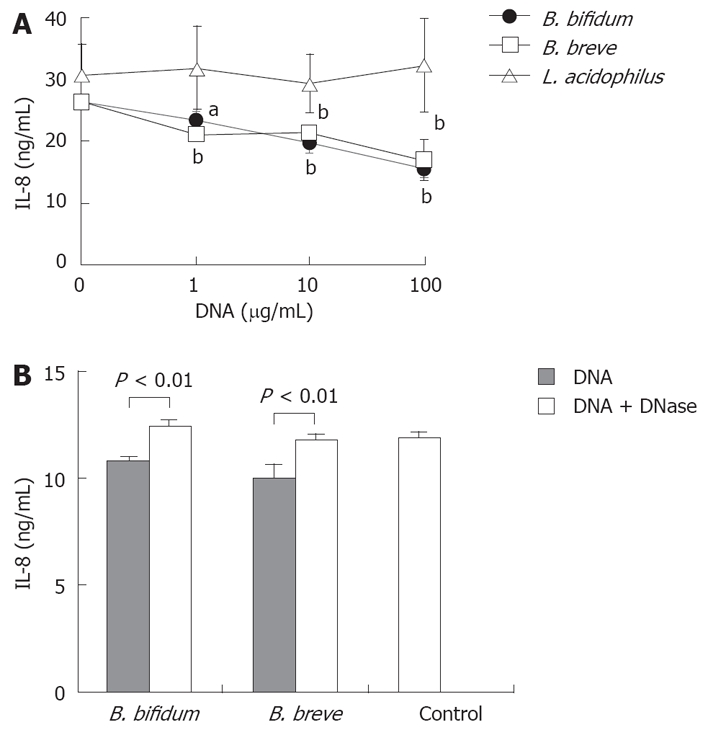
Effects of probiotic DNAs on IL-8 production in HT-29 cells. HT-29 cells were stimulated with TNF-α in the absence or presence of various concentrations of probiotic DNA without (A) or with DNase treatment (B). The inhibitory effect on IL-8 secretion was determined as described in Figure 2. aP < 0.05, bP < 0.01 vs control.
DISCUSSION
The efficacy of bifidobacteria-fermented milk (BFM) in the treatment of ulcerative colitis has been reported elsewhere[7,8]. To determine the anti-inflammatory activity of probiotic Bifidobacterium strains in the BFM, we firstly investigated the effects of these strains on the production of anti-inflammatory cytokine IL-10 by PBMNC isolated from UC patients in vitro. IL-10 down regulates the TNF-α secretion by macrophages[26]. IL-10 knockout mice develop IBD under conventional conditions, indicating the importance of IL-10 in the control of intestinal inflammation[27]. It was reported that intragastric administration of IL-10-secreting Lactococcus lactis causes a 50% reduction in colitis in mice treated with dextran sulfate sodium[14] and BFM containing BbiY and BbrY ameliorates IBD in SAMP1/Yit mice with an up-regulation of IL-10 synthesis and down-regulation of IFN-γ production in mesenteric lymph node cells[28]. In this study, the two heat-killed probiotic bacterial strains in BFM induced the secretion of IL-10 in PBMNC isolated from UC patients.
It has been found that the degree of local inflammation and tissue damage in patients with IBD is dependent on the local expression of specific chemokines within affected tissues[16]. IL-8 protein[17] and mRNA[18] expression are associated with inflammation in UC. The fact that conditioned media of BbiY and BbrY reduced the TNF-α-induced secretion of IL-8 by HT-29 cells suggests that both probiotic strains in the BFM could produce soluble anti-inflammatory factors that inhibit IL-8 secretion in intestinal epithelial cells. No single factor produced by the probiotic strain seems to be responsible, because the fractions of less than and more than 3 kDa, differing in heat-sensitivity, were found to have inhibitory effects on IL-8 secretion in this study. Acetic acid but not lactic acid, a major component of CM, was likely to largely contribute to the inhibitory activity of the less than 3 kDa fraction. Components responsible for the inhibition of IL-8 in the more than 3 kDa fraction remain to be investigated.
IκB-ζ is an inducible nuclear factor-κB (NF-κB) regulator, which is associated with the p65 and p50 subunits of NF-κB and inhibits transcription as well as the DNA-binding of the transcription factor[29,30]. Increased expression of IκB-ζ mRNA in the presence of probiotic-derived CM was observed with repression of IL-8 gene expression in HT-29 cells compared to that in the absence of CM, indicating that NF-κB-mediated IL-8 gene expression in HT-29 cells is inhibited through the induction of IκB-ζ expression by the CM of the Bifidobacterium strain.
The present study has shown that the Bifidobacterium strains proved to be beneficial in inducing and maintaining remission of ulcerative colitis exhibit regulatory activities that contribute to the control of intestinal inflammation, thus unraveling the biological basis of the beneficial effects of probiotics in human disease.
COMMENTS
Background
We have previously shown that treatment with bifidobacteria-fermented milk (BFM) containing live bifidobacteria is more effective than usual treatment in inducing and maintaining remission of ulcerative colitis (UC). We hypothesized that probiotic strains of Bifidobacteria in BFM may interfere with the intestinal inflammatory response as well as prevention of the overgrowth of potentially pathogenic bacteria such as Bacteroides vulgatus. In this study, we focused on the effect of probiotic strains on the secretion of IL-10 by peripheral blood mononuclear cells (PBMNC) and also the production of IL-8 by intestinal epithelial cells.
Research frontiers
There is evidence that probiotic treatment is effective in patients with UC. The precise mechanisms by which probiotic microorganisms used in clinical trials exert their beneficial effect have not been well defined yet.
Innovations and breakthroughs
This study has shown that Bifidobacterium strains that proved to be beneficial in inducing and maintaining remission of ulcerative colitis exhibit regulatory activities that contribute to the control of intestinal inflammation, thus unraveling the biological basis of the beneficial effects of probiotics in human disease.
Applications
Anti-inflammatory activity of probiotic bifidobacteria shown in this study supports the beneficial effects of BFM on in clinical trials. Therapeutic applications of BFM in the treatment of UC are promising.
Peer review
Probiotic Bifidobacterium strains in BFM that proved to be beneficial in inducing and maintaining remission of UC enhanced IL-10 production in PBMNC and inhibited IL-8 secretion in intestinal epithelial cells, suggesting that BFM has anti-inflammatory effects against UC. The mechanism of action regarding the immunosuppressive effect of probiotics has wider therapeutic implications for the treatment of inflammatory bowel disease in general.
Acknowledgments
The authors are grateful to Dr. Pena AS (VU University Medical Center Amsterdam) for critical reviewing of the manuscript and to Dr. H Ishikawa H (Kyoto Prefectural University of Medicine) for helpful discussion and advice regarding the manuscript.
Peer reviewers: Jay Pravda, MD, Inflammatory Disease Research Center, Gainesville, Florida, 32614-2181, United States; Francesco Costa, Dr, Dipartimento di Medicina Interna-U.O. di Gastroenterologia Università di Pisa-Via Roma, 67-56122-Pisa, Italy
S- Editor Zhong XY L- Editor Wang XL E- Editor Liu Y
References
- 1.Sartor RB. Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: antibiotics, probiotics, and prebiotics. Gastroenterology. 2004;126:1620–1633. doi: 10.1053/j.gastro.2004.03.024. [DOI] [PubMed] [Google Scholar]
- 2.Campieri M, Gionchetti P. Probiotics in inflammatory bowel disease: new insight to pathogenesis or a possible therapeutic alternative? Gastroenterology. 1999;116:1246–1249. doi: 10.1016/s0016-5085(99)70029-6. [DOI] [PubMed] [Google Scholar]
- 3.Kruis W, Fric P, Pokrotnieks J, Lukas M, Fixa B, Kascak M, Kamm MA, Weismueller J, Beglinger C, Stolte M, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut. 2004;53:1617–1623. doi: 10.1136/gut.2003.037747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bibiloni R, Fedorak RN, Tannock GW, Madsen KL, Gionchetti P, Campieri M, De Simone C, Sartor RB. VSL#3 probiotic-mixture induces remission in patients with active ulcerative colitis. Am J Gastroenterol. 2005;100:1539–1546. doi: 10.1111/j.1572-0241.2005.41794.x. [DOI] [PubMed] [Google Scholar]
- 5.Zocco MA, dal Verme LZ, Cremonini F, Piscaglia AC, Nista EC, Candelli M, Novi M, Rigante D, Cazzato IA, Ojetti V, et al. Efficacy of Lactobacillus GG in maintaining remission of ulcerative colitis. Aliment Pharmacol Ther. 2006;23:1567–1574. doi: 10.1111/j.1365-2036.2006.02927.x. [DOI] [PubMed] [Google Scholar]
- 6.Furrie E, Macfarlane S, Kennedy A, Cummings JH, Walsh SV, O'neil DA, Macfarlane GT. Synbiotic therapy (Bifidobacterium longum/Synergy 1) initiates resolution of inflammation in patients with active ulcerative colitis: a randomised controlled pilot trial. Gut. 2005;54:242–249. doi: 10.1136/gut.2004.044834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kato K, Mizuno S, Umesaki Y, Ishii Y, Sugitani M, Imaoka A, Otsuka M, Hasunuma O, Kurihara R, Iwasaki A, et al. Randomized placebo-controlled trial assessing the effect of bifidobacteria-fermented milk on active ulcerative colitis. Aliment Pharmacol Ther. 2004;20:1133–1141. doi: 10.1111/j.1365-2036.2004.02268.x. [DOI] [PubMed] [Google Scholar]
- 8.Ishikawa H, Akedo I, Umesaki Y, Tanaka R, Imaoka A, Otani T. Randomized controlled trial of the effect of bifidobacteria-fermented milk on ulcerative colitis. J Am Coll Nutr. 2003;22:56–63. doi: 10.1080/07315724.2003.10719276. [DOI] [PubMed] [Google Scholar]
- 9.Rennick DM, Fort MM. Lessons from genetically engineered animal models. XII. IL-10-deficient (IL-10(-/-) mice and intestinal inflammation. Am J Physiol Gastrointest Liver Physiol. 2000;278:G829–G833. doi: 10.1152/ajpgi.2000.278.6.G829. [DOI] [PubMed] [Google Scholar]
- 10.Asseman C, Read S, Powrie F. Colitogenic Th1 cells are present in the antigen-experienced T cell pool in normal mice: control by CD4+ regulatory T cells and IL-10. J Immunol. 2003;171:971–978. doi: 10.4049/jimmunol.171.2.971. [DOI] [PubMed] [Google Scholar]
- 11.Takahashi I, Matsuda J, Gapin L, DeWinter H, Kai Y, Tamagawa H, Kronenberg M, Kiyono H. Colitis-related public T cells are selected in the colonic lamina propria of IL-10-deficient mice. Clin Immunol. 2002;102:237–248. doi: 10.1006/clim.2001.5166. [DOI] [PubMed] [Google Scholar]
- 12.Sydora BC, Tavernini MM, Wessler A, Jewell LD, Fedorak RN. Lack of interleukin-10 leads to intestinal inflammation, independent of the time at which luminal microbial colonization occurs. Inflamm Bowel Dis. 2003;9:87–97. doi: 10.1097/00054725-200303000-00002. [DOI] [PubMed] [Google Scholar]
- 13.Foligne B, Nutten S, Grangette C, Dennin V, Goudercourt D, Poiret S, Dewulf J, Brassart D, Mercenier A, Pot B. Correlation between in vitro and in vivo immunomodulatory properties of lactic acid bacteria. World J Gastroenterol. 2007;13:236–243. doi: 10.3748/wjg.v13.i2.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Steidler L, Hans W, Schotte L, Neirynck S, Obermeier F, Falk W, Fiers W, Remaut E. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science. 2000;289:1352–1355. doi: 10.1126/science.289.5483.1352. [DOI] [PubMed] [Google Scholar]
- 15.Braat H, Rottiers P, Hommes DW, Huyghebaert N, Remaut E, Remon JP, van Deventer SJ, Neirynck S, Peppelenbosch MP, Steidler L. A phase I trial with transgenic bacteria expressing interleukin-10 in Crohn's disease. Clin Gastroenterol Hepatol. 2006;4:754–759. doi: 10.1016/j.cgh.2006.03.028. [DOI] [PubMed] [Google Scholar]
- 16.Banks C, Bateman A, Payne R, Johnson P, Sheron N. Chemokine expression in IBD. Mucosal chemokine expression is unselectively increased in both ulcerative colitis and Crohn's disease. J Pathol. 2003;199:28–35. doi: 10.1002/path.1245. [DOI] [PubMed] [Google Scholar]
- 17.Izzo RS, Witkon K, Chen AI, Hadjiyane C, Weinstein MI, Pellecchia C. Interleukin-8 and neutrophil markers in colonic mucosa from patients with ulcerative colitis. Am J Gastroenterol. 1992;87:1447–1452. [PubMed] [Google Scholar]
- 18.Katsuta T, Lim C, Shimoda K, Shibuta K, Mitra P, Banner BF, Mori M, Barnard GF. Interleukin-8 and SDF1-alpha mRNA expression in colonic biopsies from patients with inflammatory bowel disease. Am J Gastroenterol. 2000;95:3157–3164. doi: 10.1111/j.1572-0241.2000.03289.x. [DOI] [PubMed] [Google Scholar]
- 19.Umehara Y, Kudo M, Nakaoka R, Kawasaki T, Shiomi M. Serum proinflammatory cytokines and adhesion molecules in ulcerative colitis. Hepatogastroenterology. 2006;53:879–882. [PubMed] [Google Scholar]
- 20.Reddy KP, Markowitz JE, Ruchelli ED, Baldassano RN, Brown KA. Lamina propria and circulating interleukin-8 in newly and previously diagnosed pediatric inflammatory bowel disease patients. Dig Dis Sci. 2007;52:365–372. doi: 10.1007/s10620-006-9322-y. [DOI] [PubMed] [Google Scholar]
- 21.Yasui H, Nagaoka N, Hayakawa K. Augmentation of anti-influenza virus hemagglutinin antibody production by Peyer's patch cells with Bifidobacterium breve YIT4064. Clin Diagn Lab Immunol. 1994;1:244–246. doi: 10.1128/cdli.1.2.244-246.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Menard S, Candalh C, Bambou JC, Terpend K, Cerf-Bensussan N, Heyman M. Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut. 2004;53:821–828. doi: 10.1136/gut.2003.026252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, Yachimec C, Doyle J, Jewell L, De Simone C. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology. 2001;121:580–591. doi: 10.1053/gast.2001.27224. [DOI] [PubMed] [Google Scholar]
- 24.Yuki N, Shimazaki T, Kushiro A, Watanabe K, Uchida K, Yuyama T, Morotomi M. Colonization of the stratified squamous epithelium of the nonsecreting area of horse stomach by lactobacilli. Appl Environ Microbiol. 2000;66:5030–5034. doi: 10.1128/aem.66.11.5030-5034.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Truelove SC, Witts LJ. Cortisone in ulcerative colitis; final report on a therapeutic trial. Br Med J. 1955;2:1041–1048. doi: 10.1136/bmj.2.4947.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schreiber S, Heinig T, Thiele HG, Raedler A. Immunoregulatory role of interleukin 10 in patients with inflammatory bowel disease. Gastroenterology. 1995;108:1434–1444. doi: 10.1016/0016-5085(95)90692-4. [DOI] [PubMed] [Google Scholar]
- 27.Kuhn R, Lohler J, Rennick D, Rajewsky K, Muller W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell. 1993;75:263–274. doi: 10.1016/0092-8674(93)80068-p. [DOI] [PubMed] [Google Scholar]
- 28.Matsumoto S, Watanabe N, Imaoka A, Okabe Y. Preventive effects of Bifidobacterium- and Lactobacillus-fermented milk on the development of inflammatory bowel disease in senescence-accelerated mouse P1/Yit strain mice. Digestion. 2001;64:92–99. doi: 10.1159/000048846. [DOI] [PubMed] [Google Scholar]
- 29.Yamazaki S, Muta T, Takeshige K. A novel IkappaB protein, IkappaB-zeta, induced by proinflammatory stimuli, negatively regulates nuclear factor-kappaB in the nuclei. J Biol Chem. 2001;276:27657–27662. doi: 10.1074/jbc.M103426200. [DOI] [PubMed] [Google Scholar]
- 30.Totzke G, Essmann F, Pohlmann S, Lindenblatt C, Janicke RU, Schulze-Osthoff K. A novel member of the IkappaB family, human IkappaB-zeta, inhibits transactivation of p65 and its DNA binding. J Biol Chem. 2006;281:12645–12654. doi: 10.1074/jbc.M511956200. [DOI] [PubMed] [Google Scholar]