

Abstract
AIM: To investigate the total cytochrome P450 (CYP) content, microsomal mixed-function oxidase (MFO) activity, and expression of mRNAs for various CYP isozymes in a simple rat model of reversible obstructive jaundice.
METHODS: Obstructive jaundice was created in male rats by causing bile duct obstruction with polyester tape. In another group of rats, bile duct obstruction was followed by internal biliary drainage after releasing the tape. The expression of various CYP isozyme mRNAs was semi-quantitatively assessed by competitive RT-PCR.
RESULTS: The total CYP content and microsomal MFO activity showed a significant decrease after biliary obstruction, but returned to respective control levels after biliary drainage. A marked reduction in the expression of CYP1A2, 2B1/2, 2C11, 2E1, 3A1, and 3A2 mRNA was detected during biliary obstruction, while expression increased significantly toward the control level after biliary drainage. Although expression of CYP4A1 mRNA showed no reduction during biliary obstruction, it still increased significantly after biliary drainage.
CONCLUSION: These results suggest that not only obstructive jaundice, but also the subsequent internal biliary drainage may affect regulatory medications of the synthesis of individual CYP isozymes differently.
Keywords: Biliary obstruction, Obstructive jaundice, Biliary drainage, Mixed-function oxidase, P450 isozymes
INTRODUCTION
A large number of drugs are metabolized in the liver. The hepatic cytochrome P450-dependent microsomal mixed-function oxidase (MFO) system plays a major role in the metabolism of both exogenous and endogenous substrates. It is well known that some drug-metabolizing enzymes are inhibited in cholestatic human liver[1] while the total cytochrome P450 (CYP) content and MFO activity are reduced after bile duct ligation (BDL) in rats[2–4]. A possible explanation for these changes has been suggested to be bile acid-mediated destruction of CYP hemeproteins.
CYP has multiple isozymes with different substrate specificities and the broad range of substrates for the hepatic MFO system is based on the existence of these multiple isozymes. These isozymes have been shown to be the hemeprotein products of the CYP supergene family[5].
Previous studies have verified different susceptibility of BDL among CYP isozymes: male sex-specific CYPs (CYPs 2C11 and 3A2) than sexually undifferentiated CYPs (CYPs 1A, 2A1, 2B, 2C6, and 2E1)[6,7]. There are also different alterations of hepatic CYP isozyme expression in patients with cirrhosis depending on the presence or absence of cholestasis[1]. In patients with obstructive jaundice, biliary drainage has proved effective for both the treatment of jaundice itself and the subsequent complications. Ameliorating effect of biliary drainage on the total CYP content and MFO activities has been observed in bile duct obstructed rats[8,9]; however, effects of biliary drainage on individual CYP isozymes remain unknown. In this study, using a simple model of reversible obstructive jaundice, the total CYP content, MFO activity and levels of mRNAs for various CYP isozymes were compared before and after internal biliary drainage in rats with bile duct obstruction.
MATERIALS AND METHODS
Animals and reversible obstructive jaundice model
Male Wistar rats, weighing 200-240 g were fed standard laboratory chow and had free access to tap water. All animals were handled according to the local institutional guidelines for the care and use of laboratory animals. Animals were randomly assigned to undergo either bile duct obstruction alone for 4 d or 10 d (d 4-BO and d 10-BO, n = 5 each) or bile duct obstruction for 4 d followed by drainage for 6 d (BOD, n = 5). Another 10 rats underwent a sham operation and served as controls for the BO and BOD rats (d 4-sham, n = 5 and d 10-sham, n = 5). Each rat was anesthetized with a subcutaneous injection of pentobarbital sodium (50 mg/kg), and placed in the supine position on the operating table. Reversible bile duct obstruction was produced by a modification of the method described by Posner et al[10]. Using a sterile technique, a midline abdominal incision was made and the porta hepatis was isolated. In BO and BOD rats, a 2-mm wide polyester tape was placed posterior to the bile duct at the hepatic hilum. The two ends of the tape were brought out through bilateral incisions at sites 1 cm lateral to the xiphoid process. Then the abdomen was closed in two layers with 2-0 surgical silk. Proper tension was applied to the bile duct with the tape, and two ends of the tape were fixed to the skin with 4-0 silk sutures. In BOD rats, the tape was carefully removed on the 4th postoperative day, allowing free internal bile flow. In sham operated rats, the same operation was performed, but the tape was placed loosely in order to avoid pressure on the bile duct. A blood sample was collected from each rat and analyzed with standard laboratory techniques for serum alanine aminotransferase (ALT) and total bilirubin. After blood collection, the rats were sacrificed and their livers were immediately removed. Then the livers were frozen in liquid nitrogen and stored at -80°C until use.
Preparation of hepatic microsomes
Livers were homogenized in a 4-fold volume of 0.15 mol/L KCl solution containing 10 mmol/L EDTA. The homogenate was centrifuged at 10 000 g for 15 min. The supernatant was centrifuged at 105 000 g for 60 min, the pellet was suspended, and centrifugation was done again. These preparations were performed at 0-4°C. The resulting pellet was suspended in 20 mmol/L potassium phosphate buffer (pH 7.4) containing 15% glycerol and used as hepatic microsomes.
Determination of the total hepatic microsomal CYP content and MFO activity
The concentration of hepatic microsomal protein was determined according to the method of Lowry et al[11]. The total CYP content was determined according to the method of Omura and Sato[12]. Three types of MFO activity (i.e., aniline 4-hydroxylation, aminophylline N-demethylation and 7-ethoxycoumarin O-demethylation) were determined according to the methods described by Imai et al[13], Nash[14], and Ullrich and Weber[15], respectively. Each type of MFO activity was assayed by using NADPH as the sole electron source.
Determination of CYP mRNA levels by competitive RT-PCR
Total RNA was extracted from frozen hepatic tissue according to the method of Chomczynski and Sacchi[16]. The resulting RNA was reconstituted in diethylpyrocarbonate-treated water, quantitated by spectrophotometry at 260 nm, and adjusted to 500 ng/μL. The levels of mRNA for CYP1A2, 2B1/2, 2C11, 2E1, 3A1, 3A2, and 4A1 were determined with the rat cytochrome P450 competitive reverse transcriptase (RT) polymerase chain reaction (PCR) Set (Takara, Kyoto, Japan) and an RNA LA PCR™ Kit (Takara, Kyoto, Japan). Reverse transcription was performed in a final volume of 100 μL containing RT-buffer, 500 ng of total liver RNA, 5 μL of various concentrations of RNA competitor (1 × 107, 4 × 107, 1.6 × 108 and 6.4 × 108 copies/μL), 5 mol/L MgCl2, 1 mol/L of each dNTP, 1 U/μL of RNase inhibitor, 0.125 pmol/μL of oligo dT-adaptor primer, and 0.25 U/μL of AMV reverse transcriptase. Samples were incubated at 30°C for 10 min, 55°C for 20 min, 95°C for 5 min, and 5°C for 5 min. PCR amplification was performed in a final volume of 50 μL, containing PCR buffer, 10 μL of RT products, 5 mol/L MgCl2, 0.025 U/μL of Taq DNA polymerase, and 0.2 μmol/L each of the sense and antisense PCR primers. The primers were designed to specifically amplify the mRNA for CYP1A2, CYP2B1/2, CYP2C11, CYP2E1, CYP3A1, CYP3A2, CYP4A1, and cyclophilin. Cyclophilin is a housekeeping gene to correct for differences in RNA amounts between samples. Samples were incubated at 94°C for 2 min, followed by 24 cycles consisting of denaturation at 94°C for 30 s, annealing at 56°C for 30 s, and an elongation at 72°C for 30 s. Incubation of the samples was performed in a Gene Amp PCR system 9600 (Perkin-Elmer, USA). RT-PCR products were electrophoresed in 2% agarose gel. Then the gels were stained with ethidium bromide, and RT-PCR signals were visualized with UV light. Quantitation was performed by comparing RT-PCR signals generated from each specific CYP mRNA with the RT-PCR signals generated from varying concentrations of each RNA competitor.
Statistical analyses
Statistical analysis was done with the Mann-Whitney U test, and P < 0.05 was defined as indicating a significant difference between groups.
RESULTS
Characteristics of reversible obstructive jaundice model
No significant differences of the serum ALT and bilirubin levels were observed between the controls, d 4-sham rats, and d 10-sham rats. BO rats became jaundiced 4 d after bile duct obstruction, and their serum ALT and total bilirubin levels were significantly elevated compared with those of d 4-sham rats. At 6 d after the subsequent internal biliary drainage (BOD rats), these values showed a significant decrease compared with those in d 10-BO rats and were not significantly different from those in d 10-sham rats (Table 1).
Table 1.
Serum total bilirubin concentration, serum ALT concentration, the total cytochrome P450 content in hepatic microsomes and enzyme activities in hepatic microsomes from sham, bile duct obstruction alone and bile duct obstruction plus drainage groups (mean ± SD, n = 5)
| d 4-sham | d 4-BO | d 10-sham | BOD | d 10-BO | |
| Serum total bilirubin (mg/dL) | 0.12 ± 0.04 | 10.54 ± 3.54a | 0.12 ± 0.04 | 0.16 ± 0.05e | 11.18 ± 1.03 |
| Serum ALT (U/L) | 36.6 ± 5.7 | 119.6 ± 50.1a | 37.4 ± 3.9 | 2.5 ± 7.3e | 88.8 ± 11.78 |
| Total CYP content (nmol/mg MS protein) | 0.577 ± 0.029 | 0.378 ± 0.012a | 0.668 ± 0.080 | 0.603 ± 0.080e | 0.329 ± 0.05 |
| Aniline 4-hydroxylation (nmol/mg MS protein/min) | 0.527 ± 0.054 | 0.261 ± 0.026a | 0.568 ± 0.068 | 0.570 ± 0.091e | 0.252 ± 0.03 |
| Aminophylline N-demethylation (nmol/mg MS protein/min) | 7.13 ± 0.61 | 2.83 ± 0.50a | 7.88 ± 1.81 | 5.82 ± 1.00c | 2.510 ± 0.23 |
| 7-ethoxycoumarin O- demethylation (pmol/mg MS protein/min) | 104.4 ± 14.0 | 68.9 ± 20.5a | 123.0 ± 23.8 | 113.7 ± 22.9e | 64.1 ± 4.23 |
P < 0.05 vs d 4-sham;
P < 0.05 vs d 4-BO;
P < 0.05 vs d 4-BO and d 10-BO; d 10-BO and d 10-sham; BO: Bile duct obstruction alone; BOD: Bile duct obstruction plus drainage; CYP: Cytochrome P450.
Total CYP content and MFO activity
The total CYP content and MFO activity showed no significant differences between d 4-sham rats and d 10-sham rats. The total CYP content, aniline 4-hydroxylation activity, and 7-ethoxycoumarin O-demethylation activity were significantly decreased at 4 d after bile flow blockade and returned to the respective control levels after 6 d of internal biliary drainage. Aminophylline N-demethylation activity was also significantly decreased at 4 d after bile flow blockade and showed a significant increase after 6 d of internal biliary drainage, but did not return to the control level (Table 1).
CYP mRNA levels
CYP mRNA levels demonstrated no significant differences between d 4-sham rats and d 10-sham rats. The levels of CYP2C11, CYP3A2, CYP2B1/2, CYP2E1 and CYP3A1 mRNAs were significantly decreased at 4 d after bile flow blockade, and returned to the respective control levels after 6 d of internal biliary drainage (Figures 1 and 2). CYP1A2 mRNA also showed a significant decrease at 4 d after bile flow blockade and increased significantly after 6 d of internal biliary drainage, but did not return to the control level (Figure 3). CYP4A1 mRNA did not demonstrate significant change after 4 d of bile flow blockade, but showed a significant increase after 6 d of internal biliary drainage (Figure 3).
Figure 1.
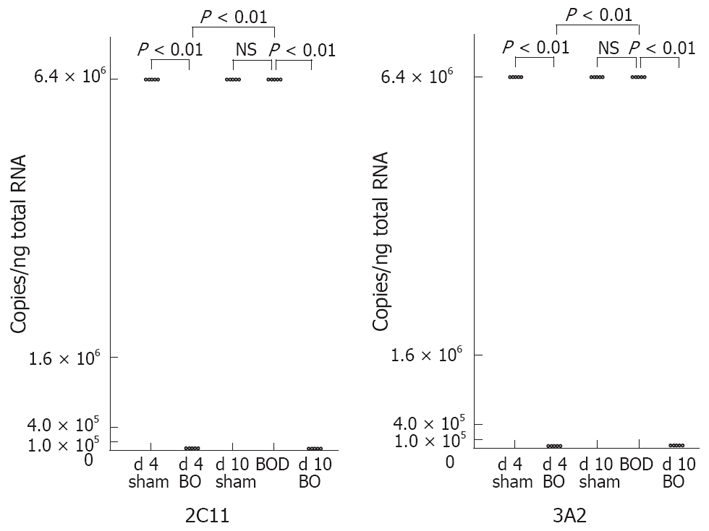
Effect of bile duct ligation and following internal biliary drainage on the mRNA expression of male-specific CYP isozymes. The mRNA expression was measured semi-quantitatively by competitive RT-PCR.
Figure 2.

Effect of bile duct ligation and following internal biliary drainage on the mRNA expression of sexually undifferentiated CYP isozymes.The mRNA expression was measured semi-quantitatively by competitive RT-PCR.
Figure 3.
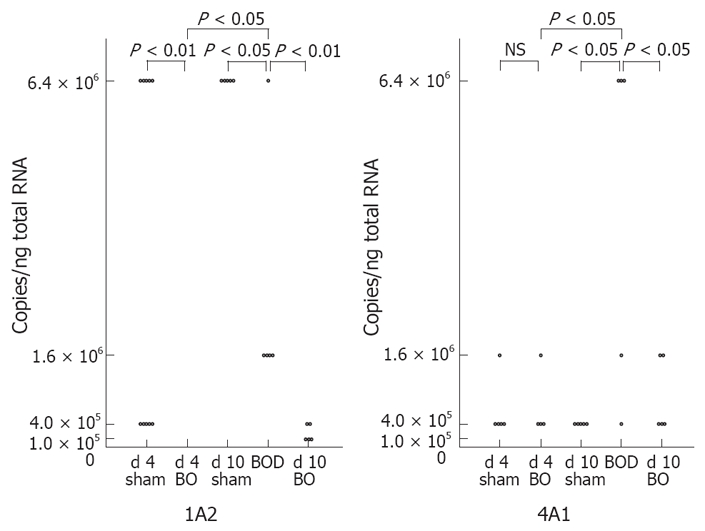
Effect of bile duct ligation and following internal biliary drainage on the mRNA expression of sexually undifferentiated CYP isozymes. The mRNA expression was measured semi-quantitatively by competitive RT-PCR.
DISCUSSION
When patients suffer from obstructive jaundice caused by conditions such as choledocholithiasis or bile duct cancer, various drugs are administered to prevent subsequent complications (e.g. acute obstructive suppurative cholangitis or sepsis). Endoscopic or percutaneous transhepatic biliary drainage has also proved effective for the treatment of obstructive jaundice and its complications[17–19]. Several studies have been conducted to ascertain the effects of bile duct obstruction on drug metabolism in the liver using animal[2–4] models and human[20], but the changes of hepatic drug metabolism following biliary drainage have not been clarified. Therefore, we investigated the total hepatic cytochrome P450 content, MFO activity, and expression of mRNA for various CYP isozymes prior to and following internal biliary drainage in rats with obstructive jaundice. The experimental model used in the present study was the internal biliary drainage model proposed by Posner et al[10]. External biliary drainage models are unphysiologic because bile is excreted out of the body after the relief of obstructive jaundice. With other internal biliary drainage models such as choledochojejunostomy, the effects of surgical invasion and anesthesia during internal biliary drainage must be taken into account. These events could affect CYP after following the relief of obstructive jaundice. On the other hand, the internal biliary drainage model used in the present study is more physiologic, and the impact of anesthesia and surgical invasion are relatively small. In rats with bile duct obstruction, irreversible fibrosis of the liver occurs after more than 14 d of bile duct obstruction[21]. Therefore, in order to observe the changes of CYP during internal biliary drainage after acute obstructive jaundice, we performed internal biliary drainage after four days of bile duct obstruction.
In jaundiced patients, proinflammatory cytokines such as TNF or IL-6 are known to be released[22], and an increase of TNF and IL-6 has been reported in murine obstructive jaundice models[23]. On the other hand, it has been reported that both TNF and IL-6 decrease the expression of mRNA for CYP isozymes[24–26]. These findings suggest that one of the mechanisms of reduced mRNA expression of CYP isozymes in obstructive jaundice may an increase in the level of cytokines such as TNF and IL-6.
The changes of the total CYP content and MFO activity following internal biliary drainage were comparable to those reported previously[8]. In the present study, although the expression of mRNA for CYP isozymes decreased after bile duct obstruction, it returned to the control level after 6 d of internal biliary drainage, except for CYP1A2. The expression of CYP1A2 mRNA was also increased after 6 d of internal biliary drainage, but not to the control level. However, this does not exclude the possibility that the expression of CYP1A2 mRNA could return to the control level after more than 6 d of internal biliary drainage. The recovery of CYP mRNA expression can be explained as follows: factors that reduced expression of the various CYP isozymes after bile duct obstruction, such as changes of sex steroids and cytokines, were eliminated or reduced by internal biliary drainage. In addition, bile duct obstruction did not alter the expression of CYP4A1 mRNA, while internal biliary drainage increased its expression. Peroxisome proliferators such as clofibrate have been known to induce CYP4A1[27,28]. It has been reported that LPS administration increases the expression of CYP4A1 mRNA in Fischer 344 rats[29]. On the other hand, it has been reported that bile duct obstruction increases the serum estradiol concentration, and that estradiol suppresses the induction of CYP4A1 by clofibrate[30]. The results of the present study showed that bile duct obstruction did not affect CYP4A1 mRNA expression although internal biliary drainage promoted of factors that increased its mRNA expression. However, it is also possible that bile duct obstruction produced multiple factors that both induced and suppressed CYP4A1 mRNA expression, so that there was no apparent change. Internal biliary drainage may then have selectively eliminated the suppressive factors and led to increased expression of CYP4A1 mRNA.
Clinically, patients with obstructive jaundice resulting from biliary disease such as choledocholithiasis or bile duct cancer are generally treated by biliary drainage, and the effectiveness of this treatment has been well documented[17–19]. In the present study, bile duct obstruction increased the serum ALT and total bilirubin levels, but both returned to their control levels after 6 d of internal biliary drainage, and obstructive jaundice dissipated. Similarly, the reduced total CYP content returned to the control level. Nonetheless, the bile duct obstruction and internal biliary drainage did not affect the expression of mRNA for the various CYP isozymes in the same manner. Internal biliary drainage did not result in sufficient recovery of the expression of mRNA for some CYP isozymes, and while bile duct obstruction had no effects, internal biliary drainage increased the expression of mRNA for some CYP isozymes. Therefore, even after the recovery of common indicators of obstructive jaundice, such as serum ATL and total bilirubin, hepatic drug metabolism could remain compromised. Caution should therefore be exercised when administering drugs following apparent recovery from obstructive jaundice.
Changes of CYP isozymes following obstructive jaundice and internal biliary drainage differed among the various isozymes, suggesting the existence of not simply a single mechanism, but multiple mechanisms. Further studies, such as the measurement of various cytokines, are therefore warranted in the future.
COMMENTS
Background
The hepatic cytochrome P450-dependent microsomal mixed-function oxidase (MFO) system plays a major role in the metabolism of exogenous and endogenous substrates. Several authors have observed a decrease of the total hepatic cytochrome P450 (CYP) content and MFO activity in cholestatic bile duct ligation models. Since CYP consists of multiple isozymes with different substrate specificities, liver damage would probably affect various isozymes differently.
Research frontiers
This study focuses on the effect of biliary drainage on individual CYP isozymes in rats with bile duct obstruction. We used a model of reversible obstructive jaundice due to bile duct obstruction, and mRNA levels of different CYP isozymes were compared before and after internal biliary drainage.
Innovations and breakthroughs
There have been previous reports about the effects of biliary drainage on the total CYP content and MFO activity in rats with bile duct obstruction, but the effects of biliary drainage on individual CYP isozymes remain unknown. We demonstrated that bile duct obstruction and subsequent internal biliary drainage had various effects on the activity and mRNA expression of different CYP isozymes.
Applications
The effects of obstructive jaundice and internal biliary drainage differed among the various CYP isozymes. These results suggest the existence of multiple mechanisms that modify CYP activity, so further studies are warranted in the future.
Peer reviewer
This study used a simple model of reversible obstructive jaundice in rats to demonstrate that biliary obstruction and subsequent biliary drainage had a differential effect on the expression of mRNA for several CYP isozymes. The experiments were well designed and conducted.
Peer reviewer: Shannon S Glaser, Dr, Department of Internal Medicine, Scott & White Hospital, 702 SW HK Dodgen Loop, Medical Research Building, Temple 76504, United States
S- Editor Zhu WL L- Editor Rippe RA E- Editor Lu W
References
- 1.George J, Murray M, Byth K, Farrell GC. Differential alterations of cytochrome P450 proteins in livers from patients with severe chronic liver disease. Hepatology. 1995;21:120–128. [PubMed] [Google Scholar]
- 2.Chen J, Farrell GC. Bile acids produce a generalized reduction of the catalytic activity of cytochromes P450 and other hepatic microsomal enzymes in vitro: relevance to drug metabolism in experimental cholestasis. J Gastroenterol Hepatol. 1996;11:870–877. doi: 10.1111/j.1440-1746.1996.tb00095.x. [DOI] [PubMed] [Google Scholar]
- 3.Schaffner F, Bacchin PG, Hutterer F, Scharnbeck HH, Sarkozi LL, Denk H, Popper H. Mechanism of cholestasis. 4. Structural and biochemical changes in the liver and serum in rats after bile duct ligation. Gastroenterology. 1971;60:888–897. [PubMed] [Google Scholar]
- 4.Mackinnon AM, Simon FR. Reduced synthesis of hepatic microsomal cytochroma P450 in the bile duct ligated rat. Biochem Biophys Res Commun. 1974;56:437–443. doi: 10.1016/0006-291x(74)90861-4. [DOI] [PubMed] [Google Scholar]
- 5.Nelson DR, Koymans L, Kamataki T, Stegeman JJ, Feyereisen R, Waxman DJ, Waterman MR, Gotoh O, Coon MJ, Estabrook RW, et al. P450 superfamily: update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics. 1996;6:1–42. doi: 10.1097/00008571-199602000-00002. [DOI] [PubMed] [Google Scholar]
- 6.Chen J, Murray M, Liddle C, Jiang XM, Farrell GC. Downregulation of male-specific cytochrome P450s 2C11 and 3A2 in bile duct-ligated male rats: importance to reduced hepatic content of cytochrome P450 in cholestasis. Hepatology. 1995;22:580–587. [PubMed] [Google Scholar]
- 7.Tateishi T, Watanabe M, Nakura H, Tanaka M, Kumai T, Kobayashi S. Liver damage induced by bile duct ligation affects CYP isoenzymes differently in rats. Pharmacol Toxicol. 1998;82:89–92. doi: 10.1111/j.1600-0773.1998.tb01403.x. [DOI] [PubMed] [Google Scholar]
- 8.Nishiura S, Koga A, Yanagisawa J. Effects of bile duct obstruction and decompression on hepatic microsomal mixed function oxidase system in rats. Exp Mol Pathol. 1988;49:62–74. doi: 10.1016/0014-4800(88)90021-4. [DOI] [PubMed] [Google Scholar]
- 9.Zimmermann H, Reichen J, Zimmermann A, Sagesser H, Thenisch B, Hoflin F. Reversibility of secondary biliary fibrosis by biliodigestive anastomosis in the rat. Gastroenterology. 1992;103:579–589. doi: 10.1016/0016-5085(92)90850-x. [DOI] [PubMed] [Google Scholar]
- 10.Posner MC, Burt ME, Stone MD, Han BL, Warren RS, Vydelingum NA, Brennan MF. A model of reversible obstructive jaundice in the rat. J Surg Res. 1990;48:204–210. doi: 10.1016/0022-4804(90)90214-m. [DOI] [PubMed] [Google Scholar]
- 11.Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 12.Omura T, Sato R. The carbon monoxide-binding pigment of liver microsomes. I. evidence for its hemoprotein nature. J Biol Chem. 1964;239:2370–2378. [PubMed] [Google Scholar]
- 13.Imai Y, Ito A, Sato R. Evidence for biochemically different types of vesicles in the hepatic microsomal fraction. J Biochem. 1966;60:417–428. doi: 10.1093/oxfordjournals.jbchem.a128453. [DOI] [PubMed] [Google Scholar]
- 14.NASH T. The colorimetric estimation of formaldehyde by means of the Hantzsch reaction. Biochem J. 1953;55:416–421. doi: 10.1042/bj0550416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ullrich V, Weber P. The O-dealkylation of 7-ethoxycoumarin by liver microsomes. A direct fluorometric test. Hoppe Seylers Z Physiol Chem. 1972;353:1171–1177. doi: 10.1515/bchm2.1972.353.2.1171. [DOI] [PubMed] [Google Scholar]
- 16.Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 17.Kiil J, Kruse A, Rokkjaer M. Endoscopic biliary drainage. Br J Surg. 1987;74:1087–1090. doi: 10.1002/bjs.1800741206. [DOI] [PubMed] [Google Scholar]
- 18.Lai EC, Mok FP, Tan ES, Lo CM, Fan ST, You KT, Wong J. Endoscopic biliary drainage for severe acute cholangitis. N Engl J Med. 1992;326:1582–1586. doi: 10.1056/NEJM199206113262401. [DOI] [PubMed] [Google Scholar]
- 19.Nakayama T, Ikeda A, Okuda K. Percutaneous transhepatic drainage of the biliary tract: technique and results in 104 cases. Gastroenterology. 1978;74:554–559. [PubMed] [Google Scholar]
- 20.McPherson GA, Benjamin IS, Boobis AR, Brodie MJ, Hampden C, Blumgart LH. Antipyrine elimination as a dynamic test of hepatic functional integrity in obstructive jaundice. Gut. 1982;23:734–738. doi: 10.1136/gut.23.9.734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kountouras J, Billing BH, Scheuer PJ. Prolonged bile duct obstruction: a new experimental model for cirrhosis in the rat. Br J Exp Pathol. 1984;65:305–311. [PMC free article] [PubMed] [Google Scholar]
- 22.Lechner AJ, Velasquez A, Knudsen KR, Johanns CA, Tracy TF Jr, Matuschak GM. Cholestatic liver injury increases circulating TNF-alpha and IL-6 and mortality after Escherichia coli endotoxemia. Am J Respir Crit Care Med. 1998;157:1550–1558. doi: 10.1164/ajrccm.157.5.9709067. [DOI] [PubMed] [Google Scholar]
- 23.Bemelmans MH, Gouma DJ, Greve JW, Buurman WA. Cytokines tumor necrosis factor and interleukin-6 in experimental biliary obstruction in mice. Hepatology. 1992;15:1132–1136. doi: 10.1002/hep.1840150626. [DOI] [PubMed] [Google Scholar]
- 24.Calleja C, Eeckhoutte C, Larrieu G, Dupuy J, Pineau T, Galtier P. Differential effects of interleukin-1 beta, interleukin-2, and interferon-gamma on the inducible expression of CYP 1A1 and CYP 1A2 in cultured rabbit hepatocytes. Biochem Biophys Res Commun. 1997;239:273–278. doi: 10.1006/bbrc.1997.7468. [DOI] [PubMed] [Google Scholar]
- 25.Barker CW, Fagan JB, Pasco DS. Interleukin-1 beta suppresses the induction of P4501A1 and P4501A2 mRNAs in isolated hepatocytes. J Biol Chem. 1992;267:8050–8055. [PubMed] [Google Scholar]
- 26.Abdel-Razzak Z, Loyer P, Fautrel A, Gautier JC, Corcos L, Turlin B, Beaune P, Guillouzo A. Cytokines down-regulate expression of major cytochrome P-450 enzymes in adult human hepatocytes in primary culture. Mol Pharmacol. 1993;44:707–715. [PubMed] [Google Scholar]
- 27.Milton MN, Elcombe CR, Gibson GG. On the mechanism of induction of microsomal cytochrome P450IVA1 and peroxisome proliferation in rat liver by clofibrate. Biochem Pharmacol. 1990;40:2727–2732. doi: 10.1016/0006-2952(90)90594-b. [DOI] [PubMed] [Google Scholar]
- 28.Bars RG, Bell DR, Elcombe CR. Induction of cytochrome P450 and peroxisomal enzymes by clofibric acid in vivo and in vitro. Biochem Pharmacol. 1993;45:2045–2053. doi: 10.1016/0006-2952(93)90015-o. [DOI] [PubMed] [Google Scholar]
- 29.Sewer MB, Koop DR, Morgan ET. Endotoxemia in rats is associated with induction of the P4504A subfamily and suppression of several other forms of cytochrome P450. Drug Metab Dispos. 1996;24:401–407. [PubMed] [Google Scholar]
- 30.Hiratsuka M, Matsuura T, Sato M, Suzuki Y. Effects of gonadectomy and sex hormones on the induction of hepatic CYP4A by clofibrate in rats. Biol Pharm Bull. 1996;19:34–38. doi: 10.1248/bpb.19.34. [DOI] [PubMed] [Google Scholar]