

Abstract
Chemical inducers of dimerization (CIDs) are cell-permeable small molecules capable of dimerizing two protein targets. The most widely used CID, the natural product rapamycin and its relatives, is immunosuppressive due to interactions with endogenous targets and thus has limited utility in vivo. Here we report a new biocompatible CID, Tmp-SLF, which dimerizes E. coli DHFR and FKBP and has no endogenous mammalian targets that would lead to unwanted in vivo side effects. We employed Tmp-SLF to modulate gene expression in a yeast three-hybrid assay. Finally, we engineered the Golgi-resident glycosyltransferase FucT7 for tunable control by Tmp-SLF in mammalian cells.
The development of small molecules that modulate protein function in a tunable fashion has been a major focus in chemical biology. First described by Schreiber and co-workers,(1) chemical inducers of dimerization (CIDs) are cell-permeable, bidentate molecules capable of dimerizing two substrates. The prototype is the immunosuppressive natural product rapamycin, which binds simultaneously(2) to the FK506/rapamycin binding protein FKBP(3) and a domain of the mTOR protein termed FRB.(4) A variety of biological processes have been probed by rapamycin-induced dimerization of proteins fused to FKBP and FRB.1,5
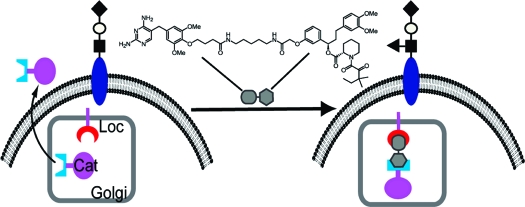
We have recently employed the CID technique in studies of glycobiology.(6) Golgi-resident glycosyltransferases and sulfotransferases comprise discrete catalytic (Cat) and localization (Loc) domains that are both required for cellular function.(7) Taking advantage of their modular nature, we separated the two domains and fused them independently to FKBP and FRB. In the absence of rapamycin, the Cat domain failed to localize to the Golgi compartment and was therefore unable to access its normal substrates (Figure 1A). The addition of rapamycin induced heterodimerization of the Loc and Cat domains, reconstituting the enzyme and restoring cellular activity.
Figure 1.

Conditional activation of a Golgi glycosyltransferase using a CID. (A) The membrane-associated Loc and soluble Cat domains are separated and fused to small molecule binding proteins. In the absence of the CID, the Cat domain has no mechanism for Golgi retention and is secreted from the cell. In the presence of the CID, the Cat domain associates with the Loc domain and is therefore retained in the Golgi compartment where it can act on substrates. In this depiction, the glycosyltransferase is fucosyltransferase 7, which adds fucose to a glycan substrate forming sialyl Lewis x. Monosaccharide symbols: (◆), sialic acid; (○), galactose; (◼), GlcNAc; (▲), fucose. (B) Tmp-SLF.
While useful for studies with cultured cells, rapamycin’s interaction with endogenous mTOR leads to undesirable in vivo side effects.(4) Thus, we sought to create a new CID that does not interfere with critical endogenous processes. Here we report the development of a trimethoprim (Tmp) conjugate (Tmp-SLF, Figure 1B) capable of dimerizing a bacterial dihydrofolate reductase (DHFR) with FKBP. The compound was employed to modulate transcription in yeast and glycosylation in mammalian cells.
The new CID replaces the mTOR-binding component of rapamycin-like analogues with a moiety, Tmp, that has no endogenous protein targets. The compound was modeled after our previously reported methotrexate (Mtx)-SLF conjugate,8,9 but unlike the promiscuous DHFR inhibitor Mtx, Tmp exhibits a 12,000-fold preference for E. coli versus human DHFR.(10) Confined within the Golgi compartment, E. coli DHFR should not perturb endogenous folate metabolism. The other CID component, SLF, is a synthetic analogue of the natural product FK50611,12 that lacks FK506’s immunosuppressive activity,1,13 is cell-permeable, and binds FKBP with nanomolar affinity.(12) The synthesis of Tmp-SLF is described in the Supporting Information (SI).
As Mtx-SLF and Tmp-SLF bind the same protein pair (DHFR/FKBP), we first tested the ability of Tmp-SLF to activate transcription in a yeast three-hybrid assay (Figure 2A).8,9 Yeast were engineered to express a B42 transcription activation domain-FKBP (B42-FKBP) protein chimera and LexA DNA binding domain-DHFR fusion (LexA-DHFR). A lacZ reporter gene under the control of four tandem LexA operators was used. The cells were grown on X-gal plates under standard conditions for 3 days in the presence of 1 μM Tmp-SLF or Mtx-SLF (Figure 2B). Both Mtx-SLF and Tmp-SLF activated lacZ transcription in yeast cells. Control strain investigations with previously described FKBP mutants fused to B42(8) indicate that Tmp-SLF is potentially more effective as a transcriptional activator than Mtx-SLF (see SI). In contrast, the previously reported Tmp-Dex CID appeared less active than Mtx-Dex in the same yeast three-hybrid system.(14)
Figure 2.

Yeast three-hybrid assay of Tmp-SLF activity. (A) Tmp-SLF (or Mtx-SLF) heterodimerizes the DNA-binding LexA domain-DHFR fusion protein and a B42-FKBP transcriptional activator fusion protein, effectively reconstituting a transcriptional activator and activating transcription of the lacZ reporter gene. (B) Yeast treated with 1 μM Tmp-SLF or Mtx-SLF grown on X-gal plates. The extent of X-gal hydrolysis (indigo color) provides a measure of the level of transcriptional activation.
Next, we evaluated the ability of Tmp-SLF to activate a Golgi- resident glycosyltransferase in mammalian cells. Fucosyltransferase VII (FucT7) adds an α1,3-linked fucose residue to sialyl N-acetyllactosamine to generate the sialyl Lexis x (sLex) epitope.(15) The enzyme participates in the assembly of selectin ligands, thereby playing a critical role in the immune system. Accordingly, immunosuppressive CIDs based on rapamycin are not suitable for in vivo studies of FucT7 function. A more biocompatible CID such as Tmp-SLF would potentially enable conditional activation of FucT7 in vivo.
We fused DNA encoding the FucT7 Loc and Cat domains(6a) separately to one, two, or three tandem copies of the FKBP or DHFR gene. Matched pairs of the corresponding chimeric proteins were transiently expressed in Chinese hamster ovary (CHO) cells, which lack endogenous FucT7 activity, in the presence or absence of Tmp-SLF. Cell-surface sLex was detected with a biotin-conjugated anti-sLex antibody (Mab HECA-452) followed by tricolor-labeled streptavidin and quantified by flow cytometry. Of the 18 protein pairs tested, 6 were active with the Loc-DHFR and 3xFKBP-Cat combination giving the best response (see SI). Cells transfected with these two constructs expressed sLex in the presence of Tmp-SLF, but not in its absence (Figure 3). The CID had no effect on the activity of full-length FucT7, and the 3xFKBP-Cat construct alone produced no detectable sLex in the presence or absence of Tmp-SLF. Furthermore, cells transfected with Loc-DHFR and 3xDHFR-Cat, a mismatched pair, expressed no detectable sLex and were unresponsive to Tmp-SLF.
Figure 3.

Effect of Tmp-SLF on FucT7 activity in CHO cells. Cells were transfected with plasmids encoding the indicated FucT7 constructs. Open bars: vehicle-treated cells; filled bars: cells treated with 1 μM Tmp-SLF for 24 h. Cells were then incubated with biotinylated Mab HECA-452 followed by tricolor-streptavidin and analyzed by flow cytometry. MFI = mean fluorescence intensities of all live cells. Error bars = SD of triplicates.
Importantly, modulation of sLex expression by Tmp-SLF was tunable and inhibitable by free Tmp. Tmp-SLF-dependent FucT7 activity was dose-dependent with an EC50 value of 43 nM (Figure 4A) and was inhibited by simultaneous treatment with Tmp (IC50 = 1.5 nM) (Figure 4B). Finally, we observed no cytotoxicity at the various Tmp-SLF concentrations tested, even with doses 200-fold higher than the EC50. By contrast, Mtx has an LD50 of 17.3 nM with CHO cells.(16)
Figure 4.

Activation of FucT7 by Tmp-SLF is tunable and can be inhibited with Tmp. (A) Cells transfected with either Loc-DHFR/3xFKBP-Cat (●) or Loc-DHFR/3xDHFR-Cat (○) construct pairs were treated with Tmp-SLF at various concentrations. sLex was probed as in Figure 3. (B) Free Tmp inhibits the Tmp-SLF-dependent response. MFI = mean fluorescence intensities of all live cells. Error bars = SD of three replicates.
In summary, Tmp-SLF can modulate Golgi proteins in mammalian systems without off-target interactions and in vivo immunosuppression. Its application in vivo is a future goal.
Acknowledgments
We thank Zeljka Cabrilo for E. coli DH5α genomic DNA and Steven Rosen for the FucT7 cDNA. This work was supported by grants from the National Institutes of Health to C.R.B. (GM59907) and V.W.C. (GM071754). J.L.C. was supported by an NIH postdoctoral fellowship (GM69156), and J.J.K. was supported by an American Cancer Society postdoctoral fellowship (PF TBE-101932).
Supporting Information Available
Synthetic procedures and spectral data for Tmp-SLF, general biological procedures, yeast three-hybrid assays with FKBP mutants, and plasmid maps for FucT7 fusions. This material is available free of charge via the Internet at http://pubs.acs.org.
Supplementary Material
References
- Spencer D. M.; Wandless T. J.; Schreiber S. L.; Crabtree G. R. Science 1993, 262, 1019. [DOI] [PubMed] [Google Scholar]
- Choi J.; Chen J.; Schreiber S. L.; Clardy J. Science 1996, 273, 239. [DOI] [PubMed] [Google Scholar]
- Standaert R. F.; Galat A.; Verdine G. L.; Schreiber S. L. Nature 1990, 346, 671. [DOI] [PubMed] [Google Scholar]
- Brown E. J.; Albers M. W.; Shin T. B.; Ichikawa K.; Keith C. T.; Lane W. S.; Schreiber S. L. Nature 1994, 369, 756. [DOI] [PubMed] [Google Scholar]
- a Klemm J. D.; Schreiber S. L.; Crabtree G. R. Annu. Rev. Immunol. 1998, 16, 569. [DOI] [PubMed] [Google Scholar]; b Pratt M. R.; Schwartz E. C.; Muir T. W. Proc. Natl. Acad. Sci. U.S.A. 2007, 104, 11209. [DOI] [PMC free article] [PubMed] [Google Scholar]; c Clackson T. Curr. Opin. Chem. Biol. 1997, 1, 210. [DOI] [PubMed] [Google Scholar]
- a Kohler J. J.; Bertozzi C. R. Chem. Biol. 2003, 10, 1303. [DOI] [PubMed] [Google Scholar]; b Kohler J. J.; Czlapinski J. L.; Laughlin S. T.; Schelle M. W.; de Graffenried C. L.; Bertozzi C. R. ChemBioChem 2004, 5, 1455. [DOI] [PubMed] [Google Scholar]; c de Graffenried C. L.; Laughlin S. T.; Kohler J. J.; Bertozzi C. R. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 16715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- a Colley K. J. Glycobiology 1997, 7, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]; b Munro S. Trends Cell Biol. 1998, 8, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Felipe K. S.; Carter B. T.; Althoff E. A.; Cornish V. W. Biochemistry 2004, 43, 10353. [DOI] [PubMed] [Google Scholar]
- Althoff E. A.; Cornish V. W. Angew. Chem., Int. Ed. 2002, 41, 2327. [DOI] [PubMed] [Google Scholar]
- Appleman J. R.; Prendergast N.; Delcamp T. J.; Freisheim J. H.; Blakley R. L. J. Biol. Chem. 1988, 263, 10304. [PubMed] [Google Scholar]
- Amara J. F.; Clackson T.; Rivera V. M.; Guo T.; Keenan T.; Natesan S.; Pollock R.; Yang W.; Courage N. L.; Holt D. A.; Gilman M. Proc. Natl. Acad. Sci. U.S.A. 1997, 94, 10618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keenan T.; Yaeger D. R.; Courage N. L.; Rollins C. T.; Pavone M. E.; Rivera V. M.; Yang W.; Guo T.; Amara J. F.; Clackson T.; Gilman M.; Holt D. A. Bioorg. Med. Chem. 1998, 6, 1309. [DOI] [PubMed] [Google Scholar]
- Braun P. D.; Barglow K. T.; Lin Y. M.; Akompong T.; Briesewitz R.; Ray G. T.; Haldar K.; Wandless T. J. J. Am. Chem. Soc. 2003, 125, 7575. [DOI] [PubMed] [Google Scholar]
- Gallagher S. S.; Miller L. W.; Cornish V. W. Anal. Biochem. 2007, 363, 160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Natsuka S.; Gersten K. M.; Zenita K.; Kannagi R.; Lowe J. B. J. Biol. Chem. 1994, 269, 16789. [PubMed] [Google Scholar]
- Assaraf Y. G.; Molina A.; Schimke R. T. J. Natl. Cancer Inst. 1989, 81, 290. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.