

Abstract
Background and Aims
Plant functional trait responses to processes such as grassland management have been analysed frequently; however, the scaling-up from individual traits to the outcomes of vegetation dynamics has seldom been tested. In this experiment, germination success was studied with respect to the relationships between grassland management (mowing and grazing), as well as abandonment, and two traits that are relevant for seedling recruitment: seed mass and germination season. On the basis of discussions in the literature and indirect trait analyses in our previous studies, the following hypotheses are proposed: (1) with respect to seed mass, mowing and grazing favour the germination of small seeds, whereas after abandonment the germination success of larger seeds is higher; and (2) with respect to germination season, mowing and grazing favour autumn-germinating seeds, whereas succession promotes spring-germinating seeds.
Methods
The germination experiment took place in a semi-natural, dry grassland in north-east Germany. Seeds of eight herbaceous species that differ with respect to seed mass and germination season were sown in mown, grazed and abandoned plots. Germination success was documented during the following year.
Key Results and Conclusions
Contrary to the hypothesis, germination of small seeds was not promoted by mowing or grazing and they germinated relatively more often than expected in the abandoned plots. A relationship between abandonment and gaps of bare soil below the vegetation cover that favour germination of small seeds was likely, but could not be proved statistically. It is possible that the small seeds suffered less from predation. Mowing favoured autumn germination, which could be explained by the removal of biomass in late summer. Contrary to our expectation, there was relatively more spring germination after grazing than after mowing, yet vegetation height was smallest in spring. Generally, germination season was found to be related to the temporal occurrence of favourable light conditions.
Key words: Field experiment, functional analysis, germination success, seed characteristics, semi-natural grassland, assembly rules
INTRODUCTION
Plant functional traits are biological characteristics of plants that both respond to and determine the dominant processes in an ecosystem (Keddy, 1992; Kelly, 1996; Gitay and Noble, 1997; Lavorel et al., 1997). There is growing interest in plant functional trait analyses, since information on assembly rules is gained and we may scale down from the species' level to the trait level. The trait level allows comparison of sites despite species' divergence and permits prediction of responses to specific processes because of functional convergence (Kahmen and Poschlod, 2004; Fukami et al., 2005). Until now, most studies on plant functional trait responses to a specific process have been indirect analyses that examine the vegetation with respect to trait distribution (e.g. Díaz et al., 1998; Lavorel et al., 1999; McIntyre et al., 1999; Köhler, 2001; Kahmen et al., 2002; Kahmen and Poschlod, 2004; Fukami et al., 2005). However, to be sure about the effects of functional traits on the outcome of vegetation dynamics, these relations have to be validated by experiments.
This paper considers the link between individual plant traits relevant for germination and seedling recruitment. Recruitment by seeds happens rarely in dense grassland vegetation, although it is essential for the population dynamics and biodiversity in grasslands (Rusch and van der Maarel, 1992; Tilman, 1993; Kitajima and Tilman, 1996; Eriksson and Eriksson, 1997; Zobel et al., 2000). Here, the focus is on the two traits of seed mass and germination season. Seed mass is an important characteristic of a plant species with regard to seedling recruitment (Coomes and Grubb, 2003; Turnbull et al., 2004). In a variety of environmental conditions it has been shown that germination and establishment success increases with increasing seed mass (summarized in Leishman et al., 1995). A relative benefit of small-seeded species or an increased germination of small seeds has been observed repeatedly following soil disturbances (McConnaughay and Bazzaz, 1987; Reader, 1993; McIntyre et al., 1995; Thompson et al., 1996; Eriksson and Eriksson, 1997). In contrast, Lavorel et al. (1999) found seed mass was not related to disturbance. They stated that many correlated traits are important for regeneration, e.g. seed production, dispersal, seed-bank longevity, life cycle (see also Thompson and Baster, 1992; Thompson et al., 1993; Leishman et al., 1995; Westoby, 1998), and that these correlated traits may therefore condition the response of seed mass. Concluding from a germination study of calcareous grassland species, Leishman (1999) stated that seed mass is not generally a good predictor for establishment ability but is highly context dependent. Despite these controversial statements, it is widely accepted that large seeds show better germination and establishment success with decreasing light than small seeds, and that late-successional species mostly have large seeds (Salisbury, 1942; Fenner, 1987; Leishman et al., 1995; Hodkinson et al., 1998).
Species-specific seasonal germination is supposed to be a response to temporal occurrence of regeneration niches (summarized in Schütz, 2000; Grime, 2001). Eriksson and Eriksson (1997) found germination in autumn being promoted through soil disturbances, whereas Lavorel et al. (1994, 1998, 1999) observed germination promotion in spring after soil disturbances. This difference may be due to the different climates of the study sites, these being northern Europe in the first and southern Europe in the latter case. Generally, the seasonal timing of germination determines the degree of competition for light experienced by the seedlings and is thus supposed to be related to the time of disturbance (McIntyre et al., 1999). Consequently, the removal of biomass by grazing or mowing in late summer should promote germination in autumn (see Olff et al., 1994).
The present paper aims to test experimentally the functional trait responses to management and abandonment of grasslands. In a previous study, we analysed trait responses to grazing, mowing and abandonment by indirect analyses of vegetation data of the ‘Fallow Experiments in Baden Württemberg’ (Schreiber, 1977; Kahmen et al., 2002; Kahmen and Poschlod, 2004). Surprisingly, for the trait seed mass there was a relative promotion of small-seeded species (<0·5 mg) at the abandoned sites. With respect to germination season, mowing and grazing favoured germination of seeds germinating in autumn, and abandonment favoured germination of seeds germinating in spring. A field germination experiment was therefore conducted to prove these trait effects on vegetation dynamics, considering mowing and grazing as management strategies as well as abandonment. Seeds of eight species that differed in seed mass and germination season were sown and germination success was analysed with respect to traits and management.
The following hypotheses were tested: (1) for seed mass that (a) mowing and grazing favour germination of small seeds (<0·5 mg) and (b) abandonment favours germination of larger seeds (0·5–2 mg); and (2) for germination season that (a) mowing and grazing favour germination of seeds germinating in autumn and (b) abandonment favours germination of seeds germinating in spring.
METHODS
Study site
The study was conducted on a pasture of 300 ha in the Müritz National Park, Germany, with a high plant species' richness (Martin, 1997). The pasture is managed by the Müritzhof in charge of the National Park, with free-ranging cattle, sheep and ponies at low intensities during the whole year, and without fertilizer application. Since the animals are free-ranging over a large area, there are intensively grazed as well as barely grazed or abandoned sites within the area. The pasture contains diverse vegetation types, from nutrient poor to rich, and dry to wet grasslands, wetlands and forests. The germination experiment was carried out on dry grassland with clusters of the shrub Juniperus communis (‘Wacholderheide’). Frequently, the clusters accreted to Juniperus shrubs where no grazing took place any more.
In 2000, 30 localities were randomly selected in open parts of the grassland and here we installed exclosure fences of 1·5 × 1·5 m and 1-m high for each locality. The grassland within the exclosures was cut once per year in August (meadow treatment). A permanent plot of 1 × 1 m was set up in the centre of the exclosures to avoid edge effects. An unfenced permanent plot was installed adjacent to each exclosure on the pasture, representing the grazing treatment. A further 30 localities close to Juniperus shrubs with no or very low grazing (see Kühner, 2004) were selected randomly. These were categorized as being abandoned despite, in principle, being part of the pasture. Again, a permanent plot of 1 × 1 m was set up in each locality, which resulted in 90 permanent plots in total.
In any experiment there is a trade-off between the number of species and the number of replicates that are used, given the usual constraints of available labour and time. The high variability in germination success led to the decision to increase the number of replicates at the expense of the number of species in this experiment. Consequently, the results for trait response are based on only eight species; thus, the data on germination success are reliable for this study site.
Germination experiment
The species for the germination experiment were selected following three criteria: (1) the considered trait categories of seed mass and germination season should be represented equally; (2) seedlings should be unambiguous to identify; and (3) the species should occur at the study site (Table 1). Following Kahmen et al. (2002), seed mass was considered with two categories, small (<0·5 mg) and large (0·5–2 mg). The categories of germination season selected contained species that germinate mainly in autumn (autumn germination) and those that germinate mainly in spring (spring germination). The seeds were collected in late August 2001 at the pasture and sown after 1 week, in the beginning of September. The number of seeds per species and plot is given in Table 1.
Table 1.
Species included in the experiment
| Thymus pulegioides | Achillea millefolium | Leontodon hispidus | Armeria maritima | Linum catharticum | Euphrasia officinalis | Rhinanthus serotinus | Centaurea jacea | |
|---|---|---|---|---|---|---|---|---|
| Germination season | Autumn | Autumn | Autumn | Autumn | Spring | Spring | Spring | Spring |
| Seed mass category* | Small | Small | Large | Large | Small | Small | Large | Large |
| Seed mass (mg)† | 0·12 | 0·17 | 0·90 | 1·42 | 0·11 | 0·12 | 1·60 | 1·94 |
| No. of seeds sown per plot | 100 | 100 | 50 | 100 | 100 | 100 | 50 | 100 |
*Seed mass categories: small (<0·5 mg) and large (0·5–2 mg). The nomenclature follows Wisskirchen and Haeupler (1998).
†Mean seed mass of 16 × 50 seeds per species is given.
The permanent plots were divided into 20 × 20 cm subplots and each species was sown in an individual subplot to prevent strong interspecific competition between the seedlings. Germination and establishment success were determined by counting seedlings in October 2001 (autumn), April 2002 and June 2002 (spring). Control of the seedlings of ‘naturally’ sown seeds took place in one control-subplot per permanent plot.
Plot characteristics
On all 90 plots the following site factors were measured: (1) height of the vegetation in September 2001, and (2) April 2002; (3) photosynthetically active radiation (PAR); (4) temperature at the soil surface; (5) percentage of bare soil; (6) percentage cover of the moss layer; (7) height of the moss layer; (8) percentage cover of the litter layer; (9) height of the litter layer; and (10) pH value. The height of the vegetation, PAR, percentage of bare soil, percentage cover of moss layer, height of the moss layer, percentage cover of litter layer and height of the litter layer were all measured in September 2001. These were determined per subplot (eight subplots per plot), whereas the remaining factors pertained to the plots.
PAR was measured using a SunScan (UP GmbH, Cottbus, Germany) at the soil surface in combination with a Beam Fraction Sensor above the vegetation. Using both values the percentage of PAR that reached the soil surface in relation to the total PAR could be calculated. pH values were determined for a mixed soil sample consisting of three separate samples each of 4 cm diameter and 2 cm depth located next to the plots. Sampling took place in September 2001. pH measurements were made using 10 g soil in 25 mL distilled water with a pH meter CG 837 (Schott, Mainz, Germany). The temperature was measured using one Thermochrom iButton (Dallas, Semiconductor, Dallas, Texas) per plot. The iButtons were fixed at the soil surface and measured temperature every 4 h from 20 April to 2 June 2002. The mean temperature for this period was calculated.
Data analysis
During the experiment, one locality designated as abandoned was destroyed by shrub-clearing management. To keep the design balanced, the adjacent mowing and grazing treatment localities were also excluded from the analyses.
Germination success was determined as the number of seedlings in relation to the number of seeds (percentage of seedlings, in short). The germination success was corrected by the number of seedlings found in the control subplots, although germination was low in these controls.
First, the germination success recorded on the three observation dates was analysed for each species separately. Due to large variation in the percentage of seedlings, a non-parametric Kruskal–Wallis H-test examined for differences among the observation dates followed by Mann–Whitney U-tests for pair-wise comparisons. The maximal percentage of seedlings for the three observation dates of each plot and species was chosen for all further calculations.
The percentage of seedlings was determined for each trait category, seed mass (small, large) and germination season (autumn, spring) per plot. Germination success was analysed for relationships between management and traits using two-factor ANOVA for both traits separately.
One-factor ANOVA tested for differences for the site factors among the management treatments. Correlations among the site factors were calculated using Spearman correlation coefficients. Finally, the variation of all grazed plots from open to abandoned localities was considered using the site factors. The germination success was analysed for relationships among the site factors and trait categories using linear regressions with groups (GLM) for both traits separately. The site factors were the continuous variables and trait categories were the grouping variables. Percentage of bare soil was transformed by arcsin prior to analysis to optimize to normal distribution (Sokal and Rohlf, 2000). All analyses except GLM were run in SPSS 10·0. GLM was done in Genstat 5.
RESULTS
The germination success is the number of seedlings produced in relation to the number of seeds sown. All four species that favour germination in the autumn demonstrated a higher percentage of seedlings in October than in April, whereas the opposite was true for species that favour germination in spring (Fig. 1). Centaurea, which was classified for germination in spring, had clearly already germinated in the autumn. Even so, the percentage of seedlings increased significantly in April, so that Centaurea remained in its initial category of spring germination despite its germination activity during the whole observation period.
Fig. 1.

Percentage of seedlings per species with respect to the three sampling dates. Percentages refer to all plots per date. Testing for differences among the sampling dates and management treatments with the data of each plot to consider plot variances (H-test; ***P < 0·001, n.s., not significant). Different letters indicate significant differences among sampling dates, resulting from pair-wise comparisons (U-test, P < 0·05). n = 87.
Figure 2 presents the maximal percentage of seedlings produced at the three sampling dates with respect to management and both traits. The ANOVAs revealed significant relationships between management and traits. With respect to seed mass (Fig. 2A), the germination success was generally higher for large than for small seeds. On the abandoned localities the germination success of small seeds was higher than expected. With respect to germination season (Fig. 2B), seeds that favour germination in autumn produced markedly more seedlings in the mown plots, whereas spring germination was relatively higher at the grazed and abandoned plots.
Fig. 2.

Relationships between germination success and management treatment for (A) seed mass and (B) germination season, tested by two-factor ANOVA. Means and s.e. are presented, n = 29; ***P < 0·001, **P < 0·01.
The permanent plots were characterized by several site factors, as illustrated in Fig. 3 for each management treatment. The pH value was the only factor that did not differ significantly among the treatments. Vegetation height at the grazed plots was high in autumn but low in spring. The PAR was measured in September and was thus highly correlated with the respective vegetation height (r = –0·76, P < 0·001, n = 696). The height of the litter layer was also significantly correlated with vegetation height in September (r = 0·56, P < 0·001, n = 696). Furthermore, percentage cover and height of the moss layer showed a strong correlation (r = 0·83, P < 0·001, n = 696).
Fig. 3.
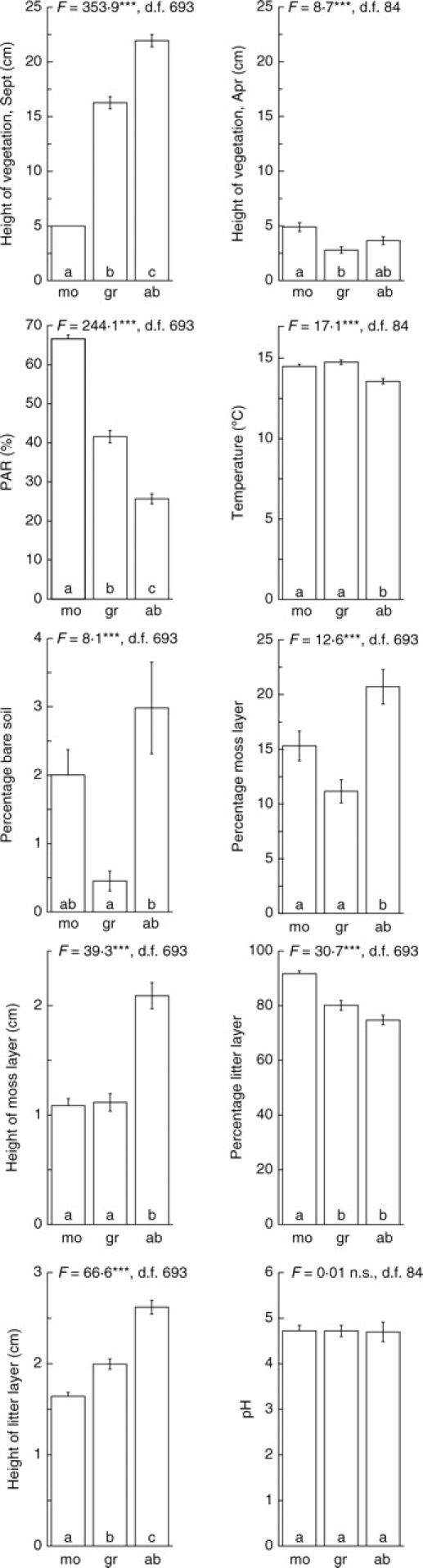
Site factors of the mown (mo), grazed (gr) and abandoned (ab) plots. Mean and s.e. are indicated. One-factor ANOVA tested for differences among the management treatments (***P < 0·001, n.s., not significant). Different letters indicate significant differences of the treatments resulting from pair-wise comparisons (Sheffé test, P < 0·05).
In order to describe the variation of the grazed plots from open to abandoned localities, all site factors except pH value were considered. Table 2 lists the regression analyses for the influence of the site factors and trait categories on germination success; these relationships are illustrated in Fig. 4. Four site factors showed a significant interaction with seed mass: the height of the vegetation in September, PAR, temperature and height of the litter layer (Table 2). PAR also interacted with germination season, although none of the other factors did. There was a stronger decrease in germination success of large than of small seeds with increasing height of the vegetation (Sept.) and height of the litter layer, as well as a stronger increase with increasing PAR and temperature (Fig. 4A). Increasing PAR promoted the germination success of seeds favouring germination in autumn more than of those favouring germination in spring (Fig. 4B).
Table 2.
Interactions between site factors of the grazed plots and the traits seed mass and germination season, tested by linear regression with groups (GLM). Site factors were the continuous and trait categories the grouping variables, df = 112
| Height of vegetation, Sept (cm) | Height of vegetation, Apr (cm) | PAR (% of incident) | Temperature (°C) | Percentage of bare soil† | Percentage of moss layer | Height of moss layer (cm) | Percentage of litter layer | Height of litter layer (cm) | |
|---|---|---|---|---|---|---|---|---|---|
| Seed mass | |||||||||
| R2 | 60·3 | 51·2 | 66·5 | 58·8 | 42·5 | 47·0 | 49·9 | 41·9 | 55·2 |
| F | 59·1*** | 41·3*** | 77·2*** | 54·8*** | 29·3*** | 35·1*** | 39·3*** | 28·6*** | 48·3*** |
| Constant | 11·3*** | 10·2*** | 3·3* | −15·1n.s. | 2·9*** | 8·4*** | 8·7*** | 8·8* | 11·9*** |
| Factor | −0·3* | −1·2* | 9·9* | 1·5n.s. | 7·7n.s. | −0·1n.s. | −1·4n.s. | 0·0n.s. | −2·3* |
| Trait | 23·0*** | 15·7*** | 5·4*** | −42·8* | 13·9*** | 13·8*** | 15·1*** | 10·9* | 21·5*** |
| Factor × Trait | −0·5** | −0·9n.s. | 23·3*** | 4·0** | −14·9n.s. | −0·1n.s. | −1·3n.s. | 0·0n.s. | −3·7* |
| Germination season | |||||||||
| R2 | 26·1 | 16·6 | 33·5 | 20·7 | 0·8 | 11·5 | 13·4 | 2·6 | 21·1 |
| F | 14·5*** | 8·7*** | 20·3*** | 10·8*** | 1·3n.s. | 6·0*** | 7·0*** | 2·0n.s. | 10·7*** |
| Constant | 22·6*** | 14·5*** | 2·4n.s. | −31·3* | 10·5*** | 12·0*** | 13·4*** | 15·2*** | 22·0*** |
| Factor | −0·6*** | −1·2* | 25·8*** | 3·0*** | 3·2n.s. | −0·1n.s. | −1·7* | −0·1n.s. | −4·9*** |
| Trait | −2·9n.s. | 4·9n.s. | 6·5** | −4·3n.s. | 2·9* | 4·6* | 3·5n.s. | −3·5n.s. | −1·8n.s. |
| Factor × Trait | 0·3n.s. | −0·7n.s. | −12·0* | 0·5n.s. | −3·8n.s. | −0·1n.s. | −0·5n.s. | 0·1n.s. | 1·9n.s. |
†Percentage of bare soil was arcsin-transformed prior to analysis.
***P < 0·001, **P < 0·01, *P < 0·05, n.s., not significant.
Fig. 4.
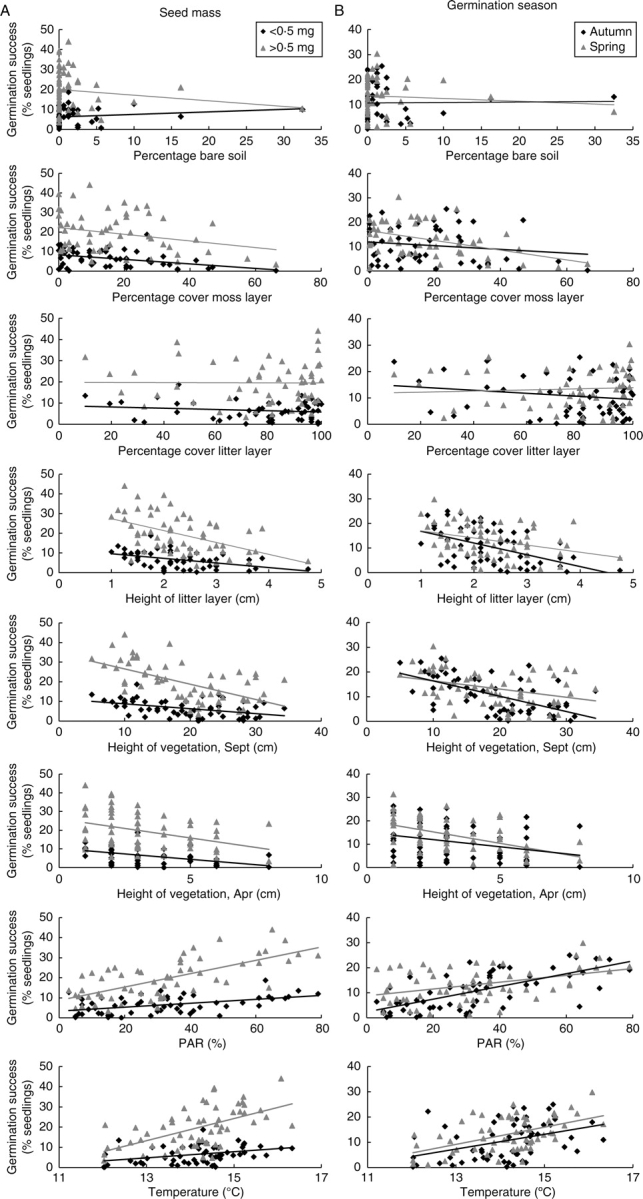
Relationships between germination success and site factors at the grazed plots for (A) seed mass and (B) germination season, visualized by regression lines for each trait category. For regression analyses see Table 2. The site factor ‘height of moss layer’ is not presented because of a high correlation with ‘percentage cover moss layer’. n = 58.
DISCUSSION
The focus of this study was on the relationships between traits and grassland management with respect to germination. Regarding the trait seed mass, there was a clear trend to more germination of large than of small seeds for all management treatments (Fig. 2A). Contrary to the general hypothesis, but in agreement with the findings in Kahmen and Poschlod (2004), germination of small seeds was not generally favoured by mowing or grazing relative to abandonment. In contrast, small seeds were relatively less badly affected by abandonment than large seeds (Fig. 2A) as well as by related site factors (increasing vegetation height, decreasing PAR and temperature; Table 2 and Fig. 4A). Grazing, and sometimes mowing, is characterized by the creation of gaps of bare soil through trampling or the action of mowing machinery. The positive effects of gaps in enhancing seedling recruitment is widely agreed (summarized in Schütz, 2000; Turnbull et al., 2000; Zobel et al., 2000). However, the absolute importance of gaps for germination and establishment of small seeds is controversial (see Introduction). In this study, the percentage of bare soil was low, particularly at the grazed plots (Fig. 3), which may be due to low grazing intensities; indeed, grazing did not promote germination of small seeds. The percentage of bare soil below the vegetation cover was slightly higher at the abandoned plots and there was a relative promotion of germination of small seeds (Fig. 2A). The interaction between percentage of bare soil and seed mass, however, could not be verified statistically, which may be due to the low variation in the percentage of bare soil among the plots (Table 2, Fig. 3).
In a study of seedling recruitment in calcareous grassland, Ryser (1990) stressed the negative impact of physical hazards such as drought or frost. He claimed that the shelter of neighbouring plants counteracts the positive effect of gaps on seedling recruitment. In this germination experiment, all seeds showed reduced germination with increasing height of the vegetation (Sept.) or, conversely, increasing germination with increasing PAR and temperature (Fig. 4A). There is thus a general disadvantage of abandonment, and also larger seeds suffer from shading. Germination success of large seeds, however, decreased more with shade than did the germination success of small seeds. According to Ryser (1990), the higher vegetation after abandonment may be a shelter against physical hazards as it effectively helps retain moisture and lowers fluctuations in temperature. This may be especially important for the germination of small seeds since they do not possess reserves to outlast bad conditions (Leishman et al., 1995). Furthermore, seedling recruitment is largely impaired by seed predation. Reader (1993) and Bonser and Reader (1998) demonstrated with cage experiments that seed mass determines the attractiveness for predators (vertebrates and invertebrates), with small seeds being less attractive. In the present study, small seeds may have suffered less from predators than larger seeds, but no study was made to see if predation was higher in the abandoned plots.
The present study revealed that small seeds were less affected than larger ones by litter (Table 2, height of the litter layer). This does not mean that litter promoted germination of small seeds, but probably the litter layer affected germination success less than other factors. This contrasts with previous observations on 35 grassland species (Jensen and Gutekunst, 2003) that germination of large seeds was less affected by litter than germination of small seeds (see also summary in Schütz, 2000).
Both hypotheses relating to the trait of germination season could be partly supported by this experiment. Mowing significantly favoured seeds that germinated mainly in autumn, whereas spring germination was relatively enhanced by grazing and abandonment (Fig. 2B). It seems reasonable to suggest that the short vegetation in autumn after the mowing treatment enhanced instant germination. The same reason may explain the relative increase of spring germination by grazing, since the vegetation of the grazed plots was shortest in spring (Fig. 3). Because of the free movement afforded to them over a large area throughout the year, the livestock favoured the dry grassland where the study took place during winter. The negative relation between vegetation height in April and spring germination further stresses the importance of short vegetation height for germination (Fig. 4B). Furthermore, decreasing PAR in September influenced germination in autumn negatively, whereas germination in spring was less affected. When seeds germinate in spring, seedlings do not risk winter mortality. Thus, if the height of the vegetation is too high to promote germination in autumn, spring germination may be the better strategy for successful seedling recruitment (Schütz, 2000). There was no relation between germination season and bare soil as discussed in the Introduction, which may again be due to low variance in the percentage of bare soil among the plots.
In conclusion, we were able to demonstrate experimentally that the functional traits seed mass and germination season significantly determine germination success with respect to grassland management and abandonment. The experiment supports the finding of Kahmen and Poschlod (2004) that the germination of small seeds is favoured by abandonment in comparison to seeds of medium weight. This is in contrast to the general idea that small seeds suffer from successional vegetation. Observing a promotion of autumn-germinating species after mowing but not after grazing indicates that management performance and intensity largely determine trait responses. However, to understand all functional trait responses to the different grassland management types is a challenge for the future. Generally, this study shows that experimental testing of functional trait responses might give deeper insights for the functional understanding of ecosystems and assembly rules.
ACKNOWLEDGEMENTS
We are grateful to the students who helped during fieldwork. We thank the Müritzhof for help in construction and removal of the exclosures. Special thanks to O. Tackenberg for valuable comments on the manuscript. The research study was funded by the German Federal Ministry of Education and Research under FKZ 01 LN 0007. The experiment complies with the current laws of Germany.
LITERATURE CITED
- Bonser SP, Reader RJ. Species and biomass dependence of an indirect effect of vegetation on seedling recruitment. Ecoscience. 1998;5:207–212. [Google Scholar]
- Coomes DA, Grubb PJ. Colonization, tolerance, competition and seed-size variation within functional groups. Trends in Ecology and Evolution. 2003;18:283–291. [Google Scholar]
- Díaz S, Marcelo C, Casanoves F. Plant functional traits and environmental filters at regional scale. Journal of Vegetation Science. 1998;9:113–122. [Google Scholar]
- Eriksson A, Eriksson O. Seedling recruitment in semi-natural pastures: the effects of disturbance, seed size, phenology and seed bank. Nordic Journal of Botany. 1997;17:469–482. [Google Scholar]
- Fenner M. Seed characteristics in relation to succession. In: Gray AJ, Crawley MJ, Edwards PJ, editors. Colonization, succession and stability. Oxford: Blackwell Scientific Publications; 1987. pp. 103–114. [Google Scholar]
- Fukami T, Bezemer TM, Mortimer SR, van der Putten WH. Species divergence and trait convergence in experimental plant community assembly. Ecology Letters. 2005;8:1283–1290. [Google Scholar]
- Gitay H, Noble IR. What are functional types and how should we seek them? In. In: Smith TM, Shugart HH, Woodward FI, editors. Plant functional types – their relevance to ecosystem properties and global change. Cambridge: Cambridge University Press; 1997. pp. 3–19. [Google Scholar]
- Grime JP. Plant strategies, vegetation processes, and ecosystem properties. 2nd. edn. Chichester, UK: John Wiley & Sons, Ltd; 2001. [Google Scholar]
- Hodkinson D, Askew A, Thompson K, Hodgson J, Bakker J, Bekker R. Ecological correlates of seed size in the British flora. Functional Ecology. 1998;12:762–766. [Google Scholar]
- Jensen K, Gutekunst K. Effects of litter on establishment of grassland plant species: the role of seed size and successional status. Basic and Applied Ecology. 2003;4:579–587. [Google Scholar]
- Kahmen S, Poschlod P. Plant functional trait repsonse to grassland succession over 25 years. Journal of Vegetation Science. 2004;15:21–32. [Google Scholar]
- Kahmen S, Poschlod P, Schreiber KF. Conservation management of calcareous grasslands. Changes in plant species composition and response of functional traits during 25 years. Biological Conservation. 2002;104:319–328. [Google Scholar]
- Keddy PA. A pragmatic approach to functional ecology. Functional Ecology. 1992;6:621–626. [Google Scholar]
- Kelly CK. Identifying plant functional types using floristic data bases: ecological correlates of plant size. Journal of Vegetation Science. 1996;7:417–424. [Google Scholar]
- Kitajima K, Tilman D. Seed banks and seedling establishment on an experimental productivity gradient. Oikos. 1996;76:381–391. [Google Scholar]
- Köhler B. Mechanisms and extent of vegetation changes in differently managed limestone grasslands. Dissertationes ETH, Zürich. 2001 http://e-collection.ethbib.ethz.ch/cgibin/show.pl?type=diss&nr=14227 . [Google Scholar]
- Kühner A. Habitat models for plant functional groups with respect to soil parameters and management. Germany: University of Oldenburg; 2004. PhD thesis. [Google Scholar]
- Lavorel S, Lepart J, Debussche M, Lebreton JD, Beffy JL. Small scale disturbance and the maintenance of species diversity in Mediterranean old fields. Oikos. 1994;70:455–473. [Google Scholar]
- Lavorel S, McIntyre S, Landsberg J, Forbes TDA. Plant functional classifications: from general groups to specific groups based on response to disturbance. Trends in Ecology and Evolution. 1997;12:474–478. doi: 10.1016/s0169-5347(97)01219-6. [DOI] [PubMed] [Google Scholar]
- Lavorel S, Touzard B, Lebreton JD, Clément B. Identifying functional groups for response to disturbance in an abandoned pasture. Acta Oecologica. 1998;19:227–240. [Google Scholar]
- Lavorel S, Rochette C, Lebreton JD. Functional groups for response to disturbance in Mediterranean old fields. Oikos. 1999;84:480–498. [Google Scholar]
- Leishman MR. How well do plant traits correlate with establishment ability? Evidence from a study of 16 calcareous grassland species. New Phytologist. 1999;141:487–496. [Google Scholar]
- Leishman MR, Westoby M, Jurado E. Correlates of seed size variation: a comparison among five temperate floras. Journal of Ecology. 1995;83:517–530. [Google Scholar]
- Martin D. Erfahrungen mit der Extensiv-Haltung von Fjällrindern im Müritz-Nationalpark. Schriftenreihe für Landschaftspflege und Naturschutz. 1997;54:161–175. [Google Scholar]
- McConnaughay K, Bazzaz F. The relationship between gap size and performance of several colonizing annuals. Ecology. 1987;68:411–416. [Google Scholar]
- McIntyre S, Lavorel S, Tremont RM. Plant life-history attributes: their relationship to disturbance response in herbaceous vegetation. Journal of Ecology. 1995;83:31–44. [Google Scholar]
- McIntyre S, Lavorel S, Landsberg J, Forbes TDA. Disturbance response in vegetation – towards a global perspective on functional traits. Journal of Vegetation Science. 1999;100:621–630. [Google Scholar]
- Olff H, Pegtel DM, van Groenendael JM, Bakker JP. Germination strategies during grassland succession. Journal of Ecology. 1994;82:69–77. [Google Scholar]
- Reader RJ. Control of seedling emergence by ground cover and seed predation in relation to seed size for some old-field species. Journal of Ecology. 1993;81:169–175. [Google Scholar]
- Rusch G, van der Maarel E. Species turnover and seedling recruitment in limestone grasslands. Oikos. 1992;63:139–146. [Google Scholar]
- Ryser P. Influence of gaps and neighbouring plants on seedling establishment in limestone grassland. Experimental field studies in northern Switzerland. Veröffentlichungen des Geobotanischen Institutes der ETH, Stiftung Rübel. 1990;104:1–71. [Google Scholar]
- Salisbury EJ. The reproductive capacity of plants. London: Bell; 1942. [Google Scholar]
- Schreiber KF. Zur Sukzession und Flächenfreihaltung auf Brachland in Baden-Württemberg. Verhandlungen der Gesellschaft für Ökologie. 1977:251–263.. Göttingen 1976. [Google Scholar]
- Schütz W. The importance of seed regeneration strategies for the persistence of species in the changing landscape of Central Europe. Zeitschrift für Ökologie und Naturschutz. 2000;9:73–83. [Google Scholar]
- Sokal R, Rohlf FJ. Biometry. 3. edn. New York: W.H. Freeman and Company; 2000. [Google Scholar]
- Thompson K, Baster K. Establishment from seed of selected Umbelliferae in unmanaged grassland. Functional Ecology. 1992;6:346–352. [Google Scholar]
- Thompson K, Band SR, Hodgson JG. Seed size and shape predict persistence in soil. Functional Ecology. 1993;7:236–241. [Google Scholar]
- Thompson K, Hillier SH, Grime JP, Bossard CC, Brand SR. A functional analysis of a limestone grassland community. Journal of Vegetation Science. 1996;7:371–380. [Google Scholar]
- Tilman D. Species richness of experimental productivity gradients: how important is colonization limitation? Ecology. 1993;74:2179–2191. [Google Scholar]
- Turnbull LA, Crawley MJ, Rees M. Are plant populations seed-limited? A review of seed sowing experiments. Oikos. 2000;88:225–238. [Google Scholar]
- Turnbull LA, Coomes D, Hector A, Rees M. Seed mass and the competition/colonization trade-off: competitive interactions and spatial patterns in a guild of annual plants. Journal of Ecology. 2004;92:97–109. [Google Scholar]
- Westoby M. A leaf–height–seed (LHS) plant ecology strategy scheme. Plant and Soil. 1998;199:213–227. [Google Scholar]
- Wisskirchen R, Haeupler H. Standardliste der Farn- und Blütenpflanzn Deutschlands. Stuttgart: Ulmer; 1998. [Google Scholar]
- Zobel M, Otsus M, Liira J, Moora M, Möls T. Is small-scale species richness limited by seed availability or microsite availability? Ecology. 2000;81:3274–3282. [Google Scholar]