

Abstract
The deteriorating in vivo environment is thought to play a major role in reduced stem cell function with age. Stem cell capacity to support tissue maintenance depends not only on their response to cues from the surrounding niche, but also on their abundance. Here we investigate satellite cell (myogenic stem cell) pool size and its potential to participate in muscle maintenance through old age. The numbers and performance of mouse satellite cells have been analyzed using molecular markers that exclusively characterize quiescent satellite cells and their progeny as they transit through proliferation, differentiation and generation of reserve cells. The study establishes that abundance of resident satellite cells declines with age in myofibers from both fast- and slow-twitch muscles. Nevertheless, the inherent myogenic potential of satellite cells does not diminish with age. Furthermore, the aging satellite cell niche retains the capacity to support effective myogenesis upon enrichment of the mitogenic milieu with FGF. Altogether, satellite cell abundance, but not myogenic potential, deteriorates with age. This study suggests that the population of satellite cells that participate in myofiber maintenance during routine muscle utilization is not fully replenished throughout life.
Keywords: Myogenesis, satellite cell, cell renewal, stem cell, myofiber, myoblast, Pax7, MyoD, FGF, aging, sarcopenia, mouse
Introduction
Aging is associated with a significant decline in the mass, strength, and endurance of skeletal muscles. This age-related muscle deterioration (sarcopenia), has been described in both human and animal models (Cartee, 1995; Karakelides and Nair, 2005; Musaro and Rosenthal, 1999). The aging muscle consists of fewer myofibers compared to adult muscle and these myofibers show signs of atrophy and increased susceptibility to contraction-induced injury (Alnaqeeb and Goldspink, 1987; Brooks and Faulkner, 1996; Edstrom and Ulfhake, 2005; Musaro and Rosenthal, 1999; Scott et al., 2001).
Age-linked muscle atrophy is thought to result in part from the diminishing ability of muscle to repair itself (Brooks and Faulkner, 1994). Muscle repair depends on myogenic stem cells, commonly referred to as satellite cells, located underneath the basal lamina of the myofiber (Collins et al., 2005; Dhawan and Rando, 2005). During postnatal growth, satellite cells proliferate and contribute progeny that fuse with the enlarging myofibers. In mature muscles, satellite cells are typically quiescent but can be recruited as needed following subtle or massive muscle trauma. When damage is minimal, satellite cells and/or their progeny fuse with existing myofibers; upon massive damage, they fuse with each other to form new myofibers (Grounds and Yablonka-Reuveni, 1993; Hawke and Garry, 2001). Since subtle myofiber injuries routinely occur during normal muscle activity, the need for ongoing repair is essential for muscle maintenance. Hence, there is a continuous demand for functional satellite cells throughout life. A decline in satellite cell abundance and function with age may limit myofiber repair and contribute to the age-associated muscle loss.
Satellite cell performance is a result of the interplay between external cues bestowed by the surrounding environment and the inherent capability of the cells to respond to those cues and undergo myogenesis. A common approach to compare satellite cell potency in old and young animals has been the induction of muscle trauma by chemical or physical means followed by the observation of subsequent tissue recovery (Carlson et al., 2001). However, rather than mimicking the myofiber repair process that occurs during day-to-day muscle maintenance, these types of “robust” injuries elicit an overall damage of the muscle tissue and trigger an acute degeneration-regeneration episode. Impaired myofiber maintenance, which is one of the hallmarks of sarcopenia (Brooks and Faulkner, 1996), may not be adequately modeled by robust injury procedures. While the aging environment fails to support muscle regeneration effectively, injured old muscle can regenerate when placed in a young host by cross-muscle grafting or when exposed to a young systemic environment by parabiotic pairings of old and young animals (Carlson et al., 2001; Carlson and Faulkner, 1989; Conboy et al., 2005). These studies suggest that the environment, rather then the muscle stem cells themselves, deteriorates with age. Although the aforementioned studies shed light on the impact of the aging environment on myogenic stem cell performance and muscle recovery, they do not provide insight whether there are age-associated changes in the satellite cell pool within individual myofibers. The performance of the myogenic stem cell pool within each myofiber may be a critical factor for maintaining myofiber integrity throughout life. Moreover, the potency of the satellite cell pool may depend not only on the capacity of cells to respond to cues elicited by their surrounding niche, but also on their abundance.
Additional studies focusing on properties of the satellite cells themselves have not provided a clear picture as to whether or not the intrinsic propensity of aging satellite cells to undergo myogenesis is impaired. (Allen et al., 1980; Beggs et al., 2004; Chakravarthy et al., 2000; Conboy et al., 2003; Gallegly et al., 2004; Grounds, 1998; Lees et al., 2005; Mouly et al., 2005; Renault et al., 2002; Schultz and Lipton, 1982). While in some of the latter studies the reduced performance of aging satellite cells was attributed to the aging environment, other studies reported that the satellite cells themselves undergo intrinsic changes with age that result in reduced proliferation and/or reduced differentiation. Discrepancies concerning the functionality of aging satellite cells may have arisen because of the variety of muscles and measurements used to identify and quantify satellite cells and their progeny, in vivo or upon their isolation. Additionally, satellite cell progeny have been typically characterized following long-term propagation in culture, a common approach for expanding the number of available cells. The resulting cells may not always represent the initial cell pool and non-myogenic cells could have (even preferentially) been expanded as well (discussed in (Machida et al., 2004)).
In this study, we investigate the age-dependent myogenic potency of the isolated myofiber unit (i.e., parent myofiber and its associated satellite cells) as well as that of the satellite cells themselves (i.e., isolated from the muscle tissue). Here, we employed several culture systems to provide a comprehensive insight into the age-dependent pool size and performance (amplification and differentiation) of satellite cells. We studied the satellite cell pool size in individual myofibers from the fast-twitch extensor digitorum longus (EDL) muscle and the slow-twitch soleus muscle. As age-linked myofiber loss is thought to be more profound in the fastest IIB myofibers (Alnaqeeb and Goldspink, 1987; Karakelides and Nair, 2005; Lexell, 1995; Musaro and Rosenthal, 1999; Scott et al., 2001), we hypothesized that myofibers from the EDL muscle (composed predominantly of fast glycolytic IIB myofibers, Allen et al., 2001) would exhibit an earlier decline in satellite cell numbers in comparison to myofibers from the soleus muscle (composed primarily of slow oxidative type I and fast oxidative type IIA myofibers, Allen et al., 2001).
We studied the initial number of satellite cells and their performance using molecular markers that exclusively characterize quiescent satellite cells and their progeny as they proliferate, differentiate and self-renew. Specifically, in postnatal muscle quiescent satellite cells express the paired-box transcription factor Pax7 while their proliferating progeny co-express Pax7 and the muscle transcription factor MyoD. The decline in Pax7 expression, along with the onset of expression of the muscle transcription factor myogenin, marks the entry of satellite cell progeny into the differentiation phase. This differentiation commitment step is rapidly followed by permanent cell cycle withdrawal, subsequent expression of muscle structural proteins, and fusion into myotubes (Halevy et al., 2004; Seale et al., 2000; Shefer et al., 2004; Yablonka-Reuveni and Rivera, 1994). In addition, cells expressing Pax7, but not MyoD or myogenin, emerge in cultures derived from postnatal satellite cells. These cells, which we termed “reserve cells”, may reflect the self-renewal potential of satellite cells (Halevy et al., 2004; Yablonka-Reuveni, 2004; Zammit et al., 2004).
In the present study we established that resident satellite cells continue to express Pax7 throughout life; this property facilitated the quantification of satellite cells in their in-situ position on the myofiber surface. Additionally, Pax7, MyoD, myogenin, and sarcomeric myosin were used in this study as molecular markers for monitoring the capacity of satellite cell progeny to proliferate and differentiate. The direct analysis of satellite cells and their progeny in individual myofibers and primary cultures enabled us to gain insight into the intrinsic performance capacity of the myofiber unit and of isolated satellite cells throughout the lifespan. This study further identifies a decline in FGF signaling as a possible limiting factor of satellite cell function during muscle aging. Based on the data presented, we suggest that the population of satellite cells that participate in myofiber maintenance during routine muscle utilization is not fully replenished throughout life.
Materials and Methods
Animals
Muscles were isolated from the hindlimbs of mice of different ages. C57BL/6 male mice were generously provided by our colleagues at the University of Washington Nathan Shock Center for Excellence in the Basic Biology of Aging program. Mice were classified into four age groups: young (3–6 month old), adult (7–10 and 11–13 months), old (19–25 months old) and senile (29–33 months old). The Myf5nlacZ/+ mouse strain (Beauchamp et al., 2000), developed by Drs. Shahragim Tajbakhsh and Margaret Buckingham (Pasteur Institute), was kindly provided by Dr. Michael Rudnicki (Ottawa Health Research Institute). Animal care and experimental procedures were approved by the Institutional Animal Care and Use Committee at the University of Washington.
Experimental approach
Throughout this investigation, satellite cells and their progeny were characterized in a variety of culture systems. Typically, each donor mouse was used for several culture systems. Muscles from young or adult mice were always processed in parallel with muscles from old or senile mice.
The C57BL/6 mouse strain was selected due to its long lifespan (Storer, 1966), a prerequisite for measuring gradual age-dependent changes. The age range chosen for this study corresponds with changes in muscle fiber maturation and life span in this strain. Specifically, the youngest age group (3–6 months) aligns with the time of postnatal EDL muscle maturation (Agbulut et al., 2003); the oldest age group (29–33 months) represents the upper limit of this strain’s life span (Goodrick, 1975; Kunstyr and Leuenberger, 1975; Mewissen, 1971). Since age-related attrition can depend on muscle activity, the high level of spontaneous activity exhibited by C57BL/6 mice (Nikulina et al., 1991) is particularly advantageous in studying the consequences of aging on muscle deterioration.
Growth media
Myofibers from EDL and soleus muscles, as well as primary myogenic cultures, were maintained in rich medium that supports both proliferation and differentiation of satellite cell progeny (Yablonka-Reuveni, 2004). This medium consisted of DMEM (Dulbeco’s Modified Eagle Medium [high glucose, with L-glutamine, 110 mg/L sodium pyruvate, and piridoxine hydrochloride] supplemented with 50U/ml penicillin and 50 mg/ml streptomycin; GIBCO-Invitrogen) containing 20% fetal bovine serum (Sigma-Aldrich), 10% horse serum (HyClone), 1% chicken embryo extract prepared from whole 10-day old embryos (Shefer and Yablonka-Reuveni, 2005; Yablonka-Reuveni, 2004).
Myofibers from flexor digitorum brevis (FDB) muscle were maintained in a basal medium (Shefer and Yablonka-Reuveni, 2005; Yablonka-Reuveni and Rivera, 1994; Yablonka-Reuveni et al., 1999a). The basal medium consisted of DMEM, 20% Controlled Process Serum Replacement (CPSR-2, bovine plasma treated with charcoal followed by dialysis, a process which depletes growth promoting factors; Sigma-Aldrich) and 1% horse serum with or without 2 ng/ml FGF2 (human recombinant; provided by Dr. S. Hauschka, University of Washington).
Isolation and culturing of intact myofibers from EDL and soleus muscles
EDL and soleus muscles were digested with collagenase for 60 and 90 minutes, respectively, and released myofibers were cultured individually in Matrigel-coated wells using rich growth medium as previously described (Shefer and Yablonka-Reuveni, 2005; Shefer et al., 2004). Typically, a preparation from a single mouse yielded four to six parallel 24-well tissue culture dishes. Due to the presence of atrophying myofibers in muscles from older mice, more myofibers were released from older animals to achieve the same number of intact myofibers as in younger animals. To detect satellite cells in their in-situ position, EDL and soleus myofibers were allowed to adhere for 3 hours followed by fixation (i.e., freshly isolated myofibers). EDL myofibers were also cultured for either 1 or 2 weeks to study long-term myogenesis. Viable myofibers may influence the performance of satellite cells during early days in culture; because of this, only cultures that contained intact myofibers by culture day 3 were further followed. Clonal cultures were derived from ten to fifteen EDL myofibers per mouse (Shefer et al., 2004). Clonal size was determined as previously described based on the number of microscopic fields occupied by the clones using a 10x objective (Yablonka-Reuveni and Rivera, 1997a). Clones were categorized as small (about 0.75 to 1 field), medium (about 1.5 to 2 fields) and large (more then 2.5 fields).
Isolation and culturing of intact FDB myofibers
To follow the initial steps of satellite cell myogenesis within the myofiber unit, myofiber cultures were prepared from the FDB muscles as previously described (Shefer and Yablonka-Reuveni, 2005; Yablonka-Reuveni et al., 1999a). Muscles were digested with collagenase for 2.5–3 hours and released myofibers were cultured in Vitrogen-coated culture dishes. Typically, a preparation of one mouse resulted in five to six 35-mm tissue culture dishes, with 20–30 myofibers per dish. Myofiber cultures were maintained in basal medium with or without FGF2; medium was replaced every 24 hours to ensure ample supply of FGF2.
Primary myogenic cultures
Cells were dissociated from the fast-twitch tibialis anterior and gastrocnemius muscles using Pronase, following our procedure for adult rodents (Kastner et al., 2000; Yablonka-Reuveni, 2004). Cells were cultured in 24-well dishes (Matrigel pre-coated, as in EDL myofiber cultures) at a density of 5×103 cells per well. Typically, a preparation from a single mouse was cultured into 12 wells per dish (5–6 dishes, one dish per time point), which permitted multiple parallel cultures per experiment. This protocol yields cultures containing 85–90% myogenic cells, based on the expression of early myogenic markers, such as Pax7 or MyoD, on culture day 4 (Yablonka-Reuveni, 2004; present study).
Immunofluorescent analysis
Freshly isolated and cultured EDL and soleus myofibers, and primary myogenic cultures were fixed with 4% paraformaldehyde followed by immunolabeling and counterlabeling of nuceli with DAPI as previously described (Shefer et al., 2004; Shefer and Yablonka-Reuveni, 2005; Yablonka-Reuveni, 2004). Some cultures were analyzed by single- and double-immunofluorescent staining using mouse, rat and rabbit primary antibodies followed by species-specific secondary antibodies. Other cultures were double-labeled with isotype-specific mouse primary antibodies and reactivity was monitored with isotoype-specific secondary antibodies (Shefer and Yablonka-Reuveni, 2005; Yablonka-Reuveni, 2004). In addition, primary cultures were triple-labeled with two isotype-specific mouse antibodies and a rabbit antibody. For double-immunolabeling, secondary antibodies were AlexaFluor 488 and 568. For triple-immunolabeling, secondary antibodies were AlexaFluor 488, 568 and 647. In order to avoid possible false positive signals due to some overlap in emission spectra of the 568 and 647 chromophores, the latter two were applied together only when demarcating antigens localized to different cellular compartments (i.e., nuclear and cytoplasmic). All secondary antibodies (Molecular Probes) were produced in goat and were used at a 1:1000 dilution. Controls consisted of cultures reacted with one primary antibody followed by 2 or 3 secondary antibodies and of cultures reacted with just secondary antibodies.
FDB myofibers were processed as above except that they were fixed with methanol (Shefer and Yablonka-Reuveni, 2005; Yablonka-Reuveni and Rivera, 1994; Yablonka-Reuveni et al., 1999a). Methanol, but not paraformaldehyde, was used in this set of studies since it maintains good adherence between the myofiber and the Vitrogen matrix throughout the immunostaining procedure.
Primary antibodies
Previously characterized primary antibodies were either monoclonal antibodies developed in mouse and rat, or polyclonal antibodies developed in rabbit. The following primary antibodies were used: anti-Pax7 (mouse IgG1, ascites fluid, Developmental Studies Hybridoma Bank [DSHB], 1:2000 dilution); anti-MyoD (mouse IgG1, clone 5.8A, BD Biosciences, 1:800; and rabbit, M-318, Santa Cruz Biotechnology, 1:400; double immunostaining with both anti-MyoD antibodies showed co-staining of the same cells); anti-myogenin (mouse IgG1, clone F5D, hybridoma supernatant, DSHB, 1:2); anti-β-galactosidase (mouse IgG2a, clone JIE7, supernatant, DSHB, 1:16); anti-sarcomeric myosin (mouse IgG2b, MF20, ascites fluid, DSHB, 1:6000); anti-CD34 (rat, BD Biosciences, 1:100); anti-PCNA (mouse IgG, clone 19F4, Boehringer Mannheim, 1:100); and anti-MAPK (ERK1/ERK2), rabbit, 1:1000; (Yablonka-Reuveni et al., 1999a; Yablonka-Reuveni et al., 1999b).
Cell Quantification
Satellite cells were detected in freshly isolated EDL and soleus myofibers by immunolabeling for Pax7. Cultured FDB myofibers were monitored for the number of myofiber-associated proliferating (PCNA+) and differentiating (myogenin+) satellite cells at different time points. Positive cells in freshly isolated and in cultured myofibers were identified and quantified by multi-focal level inspection of each myofiber using a 40x objective. In some cases, myofibers were double-labeled with the anti-MAPK antibody to confirm that satellite cells, and not the myofiber nuclei were positive for the nuclear antigens Pax7, PCNA or myogenin (Yablonka-Reuveni et al., 1999a; Yablonka-Reuveni et al., 1999b).
To quantify cells emanating from myofibers, 1- and 2-week old myofiber cultures were double-immunolabeled with mouse anti-Pax7 and rabbit anti-MyoD. Ten arbitrary microscopic-fields were analyzed with a 20x objective. Four images were acquired per field, using phase and fluorescent channels (red [Pax7], green [MyoD], blue [DAPI]). Since arbitrary fields in some of the cultures prepared from old and senile mice did not contain cells, additional images were acquired to accomplish a total of 10 cell-containing fields.
Primary myogenic cultures were quantified in a similar manner to cultures emanating from myofibers with the exception that in addition to double-labeling for Pax7 and MyoD, parallel cultures were triple-labeled with mouse anti-Pax7 (or anti-myogenin) in combination with rabbit anti-MyoD and mouse anti-sarcomeric myosin (red, green and far-red channels, respectively). During early time points, more then 10 arbitrary fields were analyzed to ensure a minimum number of 500 cells per quantification, and results were averaged per 10 fields.
Microscope and imaging system
Observations were made with an inverted fluorescent microscope (Nikon eclipse, TE2000-S). Images of live clones were acquired with Nikon Coolpix 4500 camera. Images of primary myogenic cultures were acquired with a CoolSNAPES monochrome CCD camera, which is sensitive to the far-red emitted spectra. All other images were acquired with a Qimaging Retiga 1300i Fast 1394 monochrome CCD camera. The CCD camera drive and color acquisition were controlled by MetaView Imaging System (Universal Imaging Corporation). Composites of digitized images were assembled using Adobe Photoshop software.
Statistics
All statistics were performed using Statistica 6.1. Comparisons were carried out with parametric or non-parametric testing depending on the data type. Variables were tested using the parametric or non-parametric MANOVA (multiple analysis of variance; Friedman test was used when data was significantly different from normal distribution). When significant differences were found they were followed by Tukey HSD test for non-equal sample size for comparisons. When percentages were compared, before data were analyzed by ANOVA, they were subjected to an arcsin of square root-transformation for percentages and ratios to meet the criteria of the ANOVA method. For comparisons of the distribution of proportional data, chi-square tests were performed. For all tests, P values less than 0.05 were considered significant.
Results
The number of satellite cells in freshly isolated myofibers declines with age
Satellite cells in individual myofibers of EDL and soleus muscles were quantified based on Pax7 expression (Figs. 1 and 2, Table 1). Pax7 was previously shown to be expressed in cells positioned underneath the basal membrane (i.e., satellite cells) (Halevy et al., 2004; Seale et al., 2000) and Pax7 expression has been used to trace satellite cells in isolated myofibers from young mice (Collins et al., 2005; Shefer et al., 2004). However, it has not been previously determined that Pax7 expression identifies all satellite cells in their in situ position on the myofiber. To investigate this aspect, individual myofibers from mice of different ages were double-labeled with the Pax7 antibody in combination with antibodies against MAPK (Fig. 1A–E) or CD34 (data not shown); the latter two antigens demarcate satellite cells in myofibers (Beauchamp et al., 2000; Yablonka-Reuveni et al., 1999a; Yablonka-Reuveni et al., 1999b). In the present study, over 98% of the Pax7+ cells were co-labeled for MAPK or CD34; only rarely were cells positive for Pax7 but not the other marker and vice versa. Additionally, Pax7 immunostaining was examined in myofibers isolated from Myf5nlacZ/+ mice in which nuclear lacZ expression is driven by the Myf5 promoter and resident satellite cells can be traced by beta-galactosidase staining (Beauchamp et al., 2000; Zammit et al., 2002). When cultures were double-immunostained for Pax7 and β-galactosidase (Fig. 1F–J), Pax7 highlighted all nuclei positive for lacZ (satellite cells). In adult myofibers an additional 6% of the Pax7+ nuclei were negative for lacZ (but positive for MAPK based on parallel double-labeling for Pax7 and MAPK). However, in myofibers from old mice, there were far more Pax7+ cells then lacZ+ cells. Analysis of Myf5lacZ satellite cells was based on 100 myofibers from 5 mice.
Fig. 1.
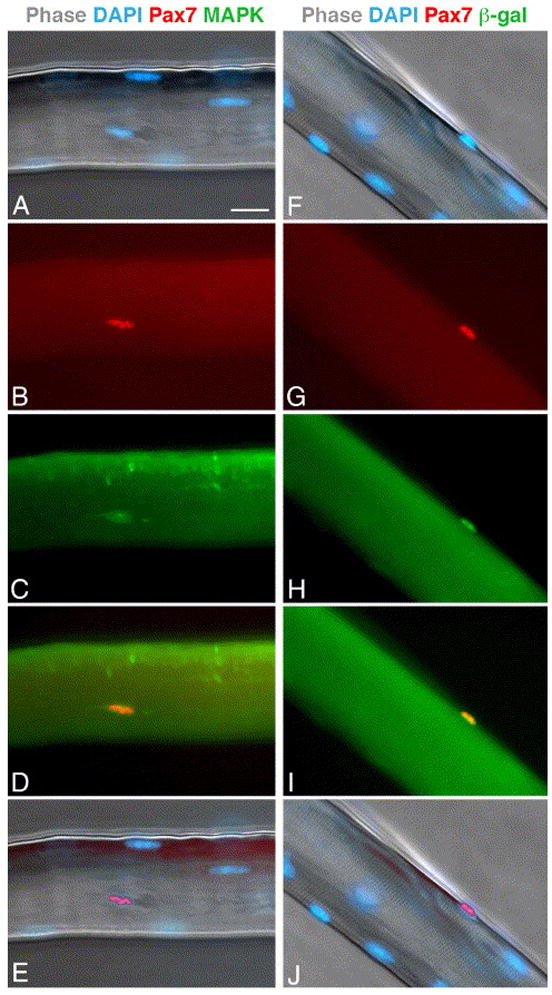
Expression of Pax7 in freshly isolated myofibers. (A–E) Parallel images of an EDL myofiber from an old mouse. The myofiber was stained with DAPI (A) -merged DAPI and phase images), and double-immunolabeled for Pax7 (B) and MAPK (C). Merged images of Pax7 and MAPK labeling (D) and Pax7 and DAPI labeling (E) are also depicted. (F–J) Parallel images of an EDL myofiber from adult Myf5nlacz/+ mouse. The myofiber was stained with DAPI (F, merged with phase image) and double-immunolabeled for Pax7 (G) and β-galactosidase (H); merged images of Pax7 and β-galctosidase (I) and Pax7 and DAPI labeling (J) are also depicted. Bar, 20 μm.
Fig. 2.
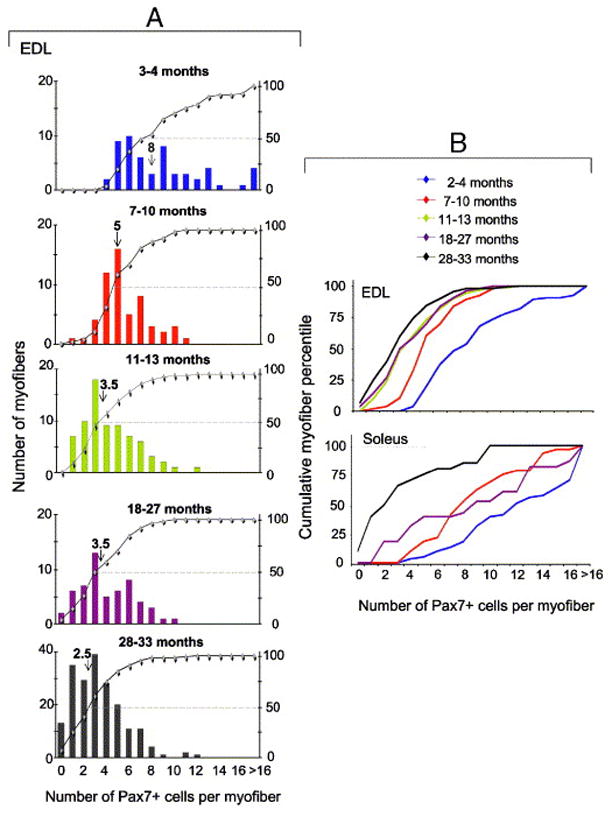
Satellite cell abundance in freshly isolated EDL and soleus myofibers. Satellite cells were enumerated by Pax7 immunolabeling. The number of mice and myofibers used in each analysis along with descriptive statistics are detailed in Table 1. Panel A depicts the distribution of individual EDL myofibers of young, adult (2 age subgroups), old and senile mice. The X-axis shows the number of satellite cells per myofiber. In each inset, myofibers are ranked from left to right according to the number of resident satellite cells they contain. The left Y-axis represents the number of myofibers and each bar shows the number of myofibers with a certain number of satellite cells. The right Y-axis represents the cumulative percentage of individual ranks relative to the total number of myofibers (–◆–); each data point reflects the percent of myofibers with the number of Pax7+ cells corresponding to this data point (as presented in the X-axis) plus all the myofibers with a lower number of satellite cells. For example, in the top panel of Fig. 2A (3–4 month old mice) 25% of the myofibers contain up to 5 Pax7+ cells and 50% of the myofibers contain up to 7 Pax7+ cells. The median number of satellite cells per myofiber is depicted in each inset by an arrow and the median value is depicted above the arrow. Panel B depicts relative cumulative plots of the distribution of EDL and soleus myofibers of all age groups together. EDL data depicted in panel B are based on those shown in each inset of A and are compiled together for comparing the kinetics of satellite cells from EDL and soleus muscle. Soleus data were compiled in the same manner as the EDL data by increments of one satellite cell at a time along the X-axis. The average number of satellite cells per each age group and muscle type and statistical analyses of the differences between the groups are included in Table 1.
Table 1.
Descriptive statistics of the number of Pax7+ cells per myofiber in the different age groups¶
| Age (months) |
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Young | Adult | Old | Senile | ||||||
| (3–6) |
(7–10) |
(11–13) |
(18–27) |
(28–23) |
|||||
| No. of Pax7+ cells§ |
EDL | Sol | EDL | Sol | EDL | EDL | Sol | EDL | Sol |
| Minimum | 4 | 4 | 2 | 4 | 1 | 0 | 2 | 0 | 0 |
| Maximum | 23 | 29 | 11 | 19 | 12 | 10 | 23 | 12 | 10 |
| Median | 8 | 12 | 5 | 8.5 | 3.5 | 3.5 | 9 | 2.5 | 2.5 |
| Average† | 9 b,c,d | 13.7 b,c,d | 5.7 d | 9.2 a,d | 4.2 b,c,d | 4 a | 9.5 a,d | 3 a,b,c | 3.3 a,b,c |
| SD‡ | 4 | 6 | 2 | 3.6 | 2.3 | 2.3 | 5 | 2 | 2 |
| No. of mice | 4 | 6 | 5 | 7 | 6 | 5 | 3 | 17 | 5 |
| No. of myofibers | 56 | 55 | 54 | 30 | 72 | 56 | 23 | 194 | 20 |
Based on data shown in Fig. 2.
A comparison of the number of Pax7+ cells per individual myofiber between the EDL and soleus muscles revealed an overall difference (two-way ANOVA, between group factor; F1,482=84.87, p<0.0001). The effect of age on the number of Pax7+ cells per myofiber in each muscle was also significant (two-way ANOVA, within group factor; F3,482=72.18, p<0.0001). Moreover, a significant interaction (two-way ANOVA, F1,3=9.63, p<0.0001) demonstrates that the decline in satellite cell numbers per myofiber across age in the EDL muscle was steeper from the decline across age in the soleus muscle.
Post-hoc comparisons (Tucky HSD for unequal sample size) within each muscle type are listed below the respective average values; a indicates difference from the young group, b from the adult group, c from the old group and d from the senile group of the same muscle.
Standard deviation of the average values. The large SD values reflect the wide range in the number of satellite cells per myofiber within each animal (as discussed in the Results section).
In view of the above characterizations, Pax7 was used in the present study as a reliable marker of satellite cells. The quantification of satellite cells in different age groups is shown in greater detail for EDL myofibers (Fig. 2A) and the results of EDL and soleus myofibers are then compared and statistically analyzed (Fig. 2B and Table 1). The following main points summarize our findings.
In all age groups, the number of satellite cells varied among individual myofibers within the same muscle. At all ages (except for the senile group), soleus myofibers contained more satellite cells than EDL myofibers. By the senile stage, the range and median values of satellite cells were the same in EDL and soleus myofibers. At all age groups, the myofibers could be broadly divided based on their diameter into large, average and narrow myofibers (with an increase in narrow-diameter myofibers in old and senile muscles); there was no significant correlation between the diameter of myofibers and the number of satellite cells. Regardless of mouse age, the majority of myofibers did not demonstrate centrally localized nuclei or segments of myonuclei chains as typically seen in myofibers from regenerating muscle (Brack et al., 2005; Collins et al., 2005; Yablonka-Reuveni and Anderson, 2006).
Analysis of EDL myofibers revealed a significant age-effect on the number of satellite cells per myofiber with an age-associated shift toward myofibers containing fewer satellite cells (Fig. 2A, B and Table 1). The number of satellite cells per myofiber was significantly higher in the two younger groups (3–4 and 7–10 months), but the median value of satellite cells per myofiber declined nearly to that of senile mice by 1 year of age (i.e, in the 11–13 month group). Moreover, there was an increase in the frequency of myofibers lacking satellite cells in the old and senile groups (chi square12= 311.65; p<0.001).
Analysis of soleus myofibers (Fig. 2B and Table 1) showed that the number of satellite cells in young muscle myofibers was significantly higher than it was in other age groups. Also, only in the senile group was there a drastic shift toward myofibers containing few satellite cells or lacking them completely.
Overall, in EDL myofibers there was a 60% decline in the median value of satellite cells between the young and old groups and a further 17% decline between the old and senile groups. However, in the soleus myofibers there was a 30% decline between the young and old groups and a 72% subsequent decline between the old and senile groups. Collectively, the major decline in the number of satellite cells in EDL myofibers occurred by 1 year of age, while in soleus myofibers it occurred by senile age.
Myogenesis in cultures emanating from EDL myofibers: number of satellite cell progeny, but not their differentiation potential, is reduced with age
Single myofibers were maintained in culture for 1- or 2-weeks. Cells emanating from individual myofibers (referred to below as “emanating cells”) were quantified, using double immunostaining, for the number of Pax7+/MyoD−, Pax7+/MyoD+, and Pax7−/MyoD+ mononucleated cells, and for the number of nuclei fused into myotubes (Fig. 3A–H and Fig. 4; each bar in Fig. 4 depicts the results of an individual myofiber culture). We analyzed cultures at the 1- and 2-week time points because we wished to determine both amplification and differentiation potential of the emanating cells. Satellite cell proliferation begins within the initial days in culture; further proliferation of progeny along with differentiation onset are detected by the first week in culture as shown in the present study and previous investigations (e.g., Zammit et al., 2002; Shefer et al., 2004). We also aimed at analyzing the progression of satellite cell progeny into the more advance phase of differentiation (i.e., fusion into myotubes) and for that we analyzed 2-week old cultures.
Fig. 3.
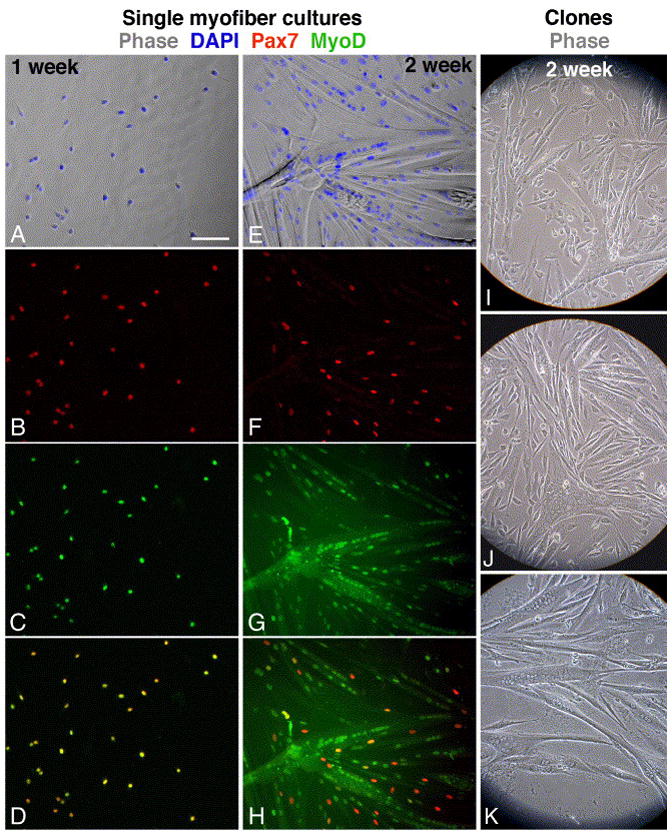
Immunofluorescent and phase images depicting myogenesis in cultures emanating from single EDL myofibers and in live clones of satellite cells that were dissociated from EDL myofibers. Myofibers from a young mouse were cultured for 1- or 2-weeks (A–D and E–H, respectively) and then double-labeled for Pax7 and MyoD. (A, E) Merged phase and DAPI images; (B, F) Pax7; (C, G) MyoD; (D, H) merged Pax7 and MyoD images demonstrating that all cells in the earlier time point co-express Pax7 and MyoD but at the later time point only residual cells are positive for both antigens. Clones were derived from EDL myofibers of young (I), old (J) and senile (K) mice. Bar, 100 μm.
Fig. 4.
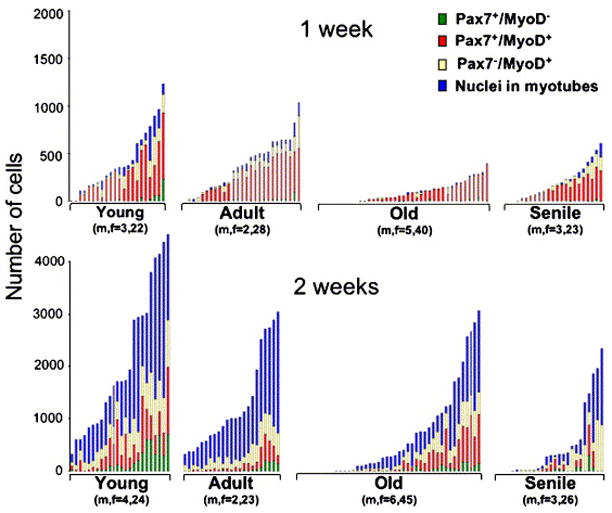
Progression of satellite cell progeny through myogenesis in cultures emanating from individual EDL myofibers. Each stacked bar represents the distribution of cells within a single myofiber culture. The number of mice (n) and the number of myofibers (f) per age group is listed under the respective inset. Data collected by double-immunolabeling and DAPI-counterstaining are shown for 1- and 2-week old cultures (top and bottom panels, respectively). In the inset of each age group, individual myofiber cultures were ranked from left to right according to the total number of myogenic nuclei they encompass. The height of each bar represents the total number of myogenic cells in the corresponding myofiber culture and the different segments in each bar represent the number of cells in each of the four distinct myogenic compartments analyzed. Myofibers that did not give rise to cells are ranked at the left part of each inset. Additional analysis of the data is included in Table 2.
Data show that regardless of age, there was a significant increase in the number of emanating cells between the first and second week in culture (two-way ANOVA, F(1,216)=88.235, p<0.0001) and that emanating cells were able to transit through the myogenic compartments to form myotubes. By the second week in culture there was no significant difference in the proportion of nuclei in each myogenic compartment among all age groups. Table 2 further summarizes the distribution of the myofibers according to the number of emanating cells by week 2 in culture. The data underscore the dramatic difference in the number of cells derived from young myofibers in comparison to the number of cells derived from myofibers of the other age groups. This difference is in agreement with the increased number of satellite cells in the younger age group (Fig. 2 and Table 1).
Table 2.
Distribution of EDL Myofiber-cultures into quartiles based on the number of emanating cells present after 2-weeks in culture¶
| % Myofiber cultures per quartile | Number of emanating cells | |||||
|---|---|---|---|---|---|---|
| Age groups |
1st quartile |
2nd quartile |
3rd quartile |
4th quartile |
Range per quartile |
Overall range |
| Young | 33 | 29 | 17 | 21 | 1174 | 0–4697 |
| Adult | 39 | 35 | 4 | 22 | 761 | 0–3045 |
| Old | 71 | 13 | 7 | 9 | 773 | 0–3092 |
| Senile | 77 | 4 | 8 | 11 | 587 | 0–2340 |
Based on data shown in Fig. 4. The range of number of cells emanating from myofibers within each age group was divided into quartiles and the incidence of myofibers that gave rise to progeny number within the range of each quartile was determined. The first quartile includes cultures with the lowest number of progeny (0–25% of the overall cell range) and the fourth quartile includes cultures with the highest number of progeny (76–100% of the overall cell range).
Notably, in both 1- and 2-week old cultures, the frequency of myofiber cultures containing few or no cells was higher in the old and senile groups than it was in the young and adult groups (Chi square9=76.588, p<0.001; Fig. 2 and Table 2). This finding is in agreement with the increased number of EDL myofibers lacking satellite cells as shown in Fig. 2. Likewise, there was an age-dependent decline in the number of myofibers giving rise to myogenic clones (data not shown). However, there was no significant difference in clone size among the different age groups (data not shown) and in the degree of morphological differentiation (estimated by the extent of myotubes formed, Fig. 3I–K). In sum, the study of cells emanating from single myofibers shows that the pool size of satellite cell progeny, and not the differentiation capacity of the cells, declined with age.
Myogenesis in primary cultures: initial proliferative phase is retarded in progeny of satellite cells from senile muscle but myogenic differentiation is not impaired
The study of cells emanating from single myofibers indicated an enhanced growth potential in myofiber cultures from the younger age group. This enhanced growth could be due to the larger initial number of satellite cells, to the potential presence of better growth signals from the parent myofibers, and/or to inherent differences in the proliferative capacity of the satellite cells founding the myofiber cultures. To exclusively compare the proliferative capacity, myogenic progenitors were isolated from young and senile muscles and cultured at the same initial density in rich growth medium. We studied young and senile age groups since they represent the best phase of muscle maintenance and the most extreme phase of muscle deterioration, respectively.
Primary cultures were analyzed for growth (total cell numbers), proliferation (number of Pax7+/MyoD+ cells), differentiation (number of Pax7−/MyoD+ cells, myogenin+ cells, and nuclei in myotubes) and accumulation of reserve cells (Pax7+/MyoD−). Representative immunostaining images are depicted in Fig. 5 and the data are summarized in Fig. 6. Values in Fig. 6B were calculated based on the total number of DAPI-stained nuclei. The total cell number (Fig 6A) includes remaining cells that did not label with either of the antibodies against the myogenic markers used in this study. These remaining cells have been considered non-myogenic and constitute the extra cells that are not categorized in Fig 6B.
Fig. 5.
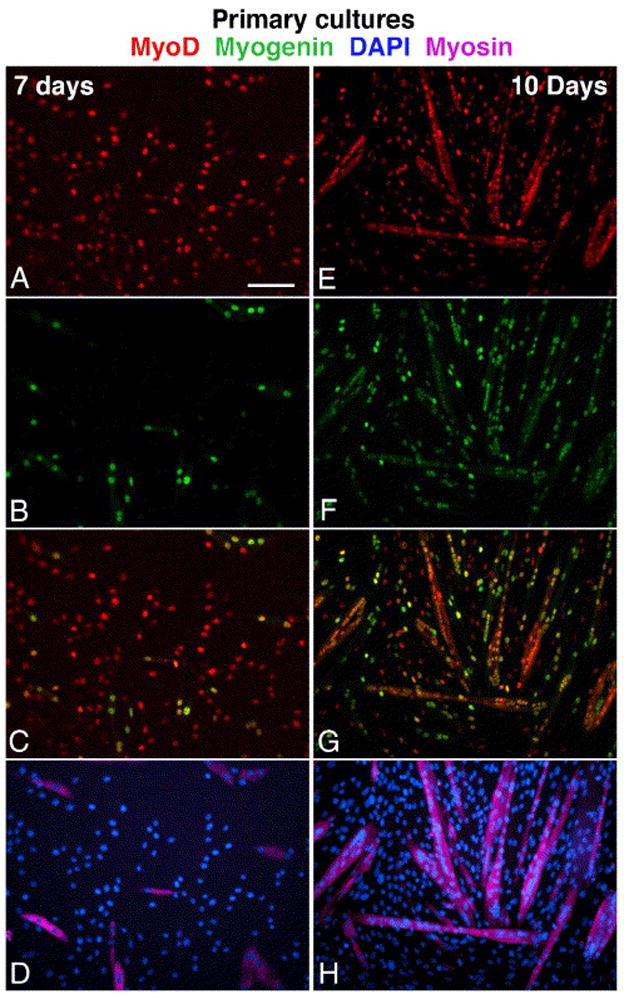
Parallel immunofluorescent images depicting myogenesis in primary cultures prepared from the tibialis anterior and gastrocnemius muscles of senile mouse. Cultures shown were fixed on day 7 (A–D) and day 10 (E–H). Cultures were triple-immunolabled for MyoD (A, E), myogenin (B, F) and sarcomeric myosin (D, H; these images merged with parallel DAPI stain). Merged images of MyoD and myogenin are also shown (C–G). The same immunostaining patterns were observed in cultures from young mice, but with an earlier onset of differentiation. Bar, 100 μm.
Fig. 6.
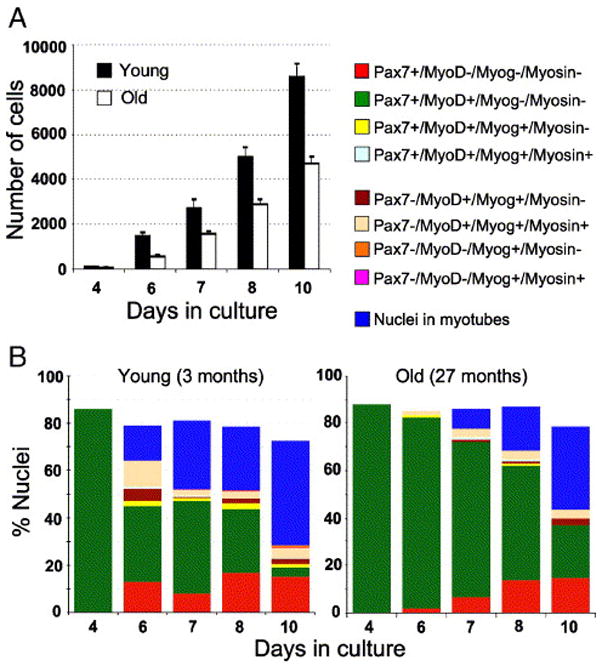
Dynamics of cell growth and myogenesis in primary cultures from young and senile muscle. Cultures were initiated in parallel from one young and one senile mouse. Total cell numbers (Panel A) and myogenic progression (Panel B) were quantified based on DAPI- and immuno-labeling. The total cell number includes remaining cells that did not label with either of the antibodies against the myogenic markers used in this study. These remaining cells were considered non-myogenic. Panel A depicts the mean number (±SEM) of total nuclei based on six parallel cultures per time point. Panel B depicts the kinetics of myogenic progression in the cultures from young (left inset) and senile (right inset) muscles; for each time point, the different segments of the stack bar reflect the percent of nuclei in different myogenic compartments; the residual cells that were not identified by the antibodies are most likely non-myogenic. At each time point, the following antibody combinations were used for double- and triple-immunolabeling of parallel cultures: Pax7/MyoD, Pax7/MyoD/myosin, MyoD/myogenin/myosin (repeating each combination twice). While cells were not co-labeled for Pax7 and myogenin, the combinations of the four markers could be inferred from the direct counts. For example, the proportion of Pax7−/MyoD+/myogenin+/myosin+ cells was calculated based on the set of cultures labeled for MyoD/myogenin/myosin (which demonstrated that all cells positive for both MyoD and myosin were also positive for myogenin) and the set of cultures labeled for Pax7/MyoD/myosin (which identified the percent of cells that were Pax7−/MyoD+/myosin+); the latter cells were thus considered to be Pax7−/MyoD+/myogenin+/myosin+. The proportion of Pax7+/MyoD+/myogenin+/myosin+ cells was deduced in the same manner, based on the number of Pax7+/MyoD+/myosin+ cells. Data shown are representative of three independent experiments.
Between the 6th and 4th day after plating, the number of cells (i.e., total DAPI-stained nuclei) in cultures from young mice increased 15-fold whereas numbers in senile cultures increased only 10-fold (Fig. 6A). These data show a more robust growth rate in the young compared to the senile group during early days in culture. The increase in cell numbers between successive time points eventually slowed-down to a 1.8-fold value in cultures from both age groups (along with differentiation and fusion into myotubes), but the decline was more gradual in the senile group. While the decline to the 1.8-fold value occurred between days 6 to 7 in the young group, it occurred between days 7 to 8 in the senile group. Altogether, there was a persistent 24-hour lag before cultures of senile muscle contained the same cell number as those of young cultures. In accordance with the dynamics of cell accumulation, the onset of differentiation in the senile cultures was delayed by 24 hours in comparison to their young counterparts; however, there was no significant difference in the transition through the myogenic compartments (Fig. 6B). Collectively, the study of primary cultures suggests that the delay in differentiation was a result of a reduced proliferative – but not a reduced differentiative – potential of satellite cell progeny from the senile muscle. The intrinsic ability to transit through the myogenic program and yield myotubes was not impaired with age. However, the kinetics of the increase in total cell number over time in culture pointed to a possible inherent impairment in the initial amplification capacity of satellite cells of senile muscle.
Myogenesis in myofibers from the FDB muscle: satellite cell performance in senile myofibers is rejuvenated when supplemented with FGF2
The FDB myofiber culture system facilitated further insight into the myogenic capacity of satellite cells from senile versus young muscle. As opposed to the EDL myofiber culture system in which satellite cells emigrate from the parent myofiber, these short (fast-twitch) FDB myofibers retain their satellite cells when cultured on Vitrogen and maintained in basal medium. Satellite cell activation, proliferation and differentiation occur within the FDB myofiber unit in a highly synchronized manner (Yablonka-Reuveni and Rivera, 1997b; Yablonka-Reuveni et al., 1999a). This feature has enabled a precise analysis of the schedule of satellite cell proliferation and differentiation within early days in culture. Furthermore, the use of a basal medium has allowed the assessment of growth promoting factors required for satellite cell proliferation (Kastner et al., 2000). Notably, the number of satellite cells associated with the short FDB myofibers is drastically lower than that associated with the longer EDL or soleus myofibers (Shefer and Yablonka-Reuveni, 2005). Regardless of myofiber length and satellite cell numbers, FDB myofibers were shown to manifest all features of longer myofibers, including dystrophy-related damage and repair (Yablonka-Reuveni and Anderson, 2005).
In the present study, proliferation and differentiation of satellite cells from young and senile muscle was quantified by immunolabeling for PCNA and myogenin, respectively (Figs. 7 and 8). Since we were interested in comparing the response of satellite cells to FGF2, myofibers that did not display associated satellite cells (based on MAPK immunostaining) were excluded from the final data; such myofibers were more prevalent in the old age group. A comparison between myofiber cultures prepared from young and senile FDB muscles (Figs. 8A and 8B) revealed an overall difference in the number of PCNA+ and of myogenin+ cells per myofiber (Friedman test, chi square2=743.95, P<0.00001). Specifically, post-hoc comparisons revealed that without FGF supplementation, the number of PCNA+ satellite cells (or subsequently myogenin+ cells) in myofibers from senile mice was significantly lower in comparison to the number in cultures from young mice (p<0.01). However, when supplemented with FGF2 there was no statistical difference between young and senile myofibers in the number of cells and their kinetics of first proliferation (PCNA+ cells) followed by differentiation (myogenin+ cells). These data suggest that the inherent capacity of satellite cells to respond to external cues (at least to FGF2) is not impaired with age. The infrequent myofibers in cultures from senile mice that showed the highest number of satellite cell progeny, even in comparison with myofibers from the young muscle, may represent “newer” myofibers that were formed much later in life due to injury.
Fig. 7.
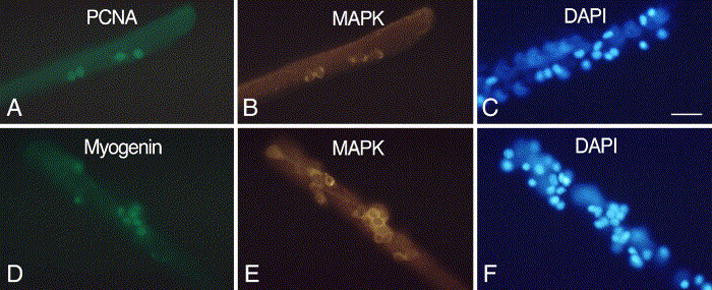
Parallel immunofluorescent and DAPI-stained images of FDB myofibers from senile mice. Myofibers were maintained in basal medium containing 2 ng/ml FGF2. Cultures were immunolabeled for MAPK in combination with PCNA (A–C, 2-day old culture) or myogenin (D–F, 4-day old culture). Not all DAPI-stained nuclei or immuno-positive cells are in the same focal plane when the myofibers are photographed. Bar, 34 μm.
Fig. 8.
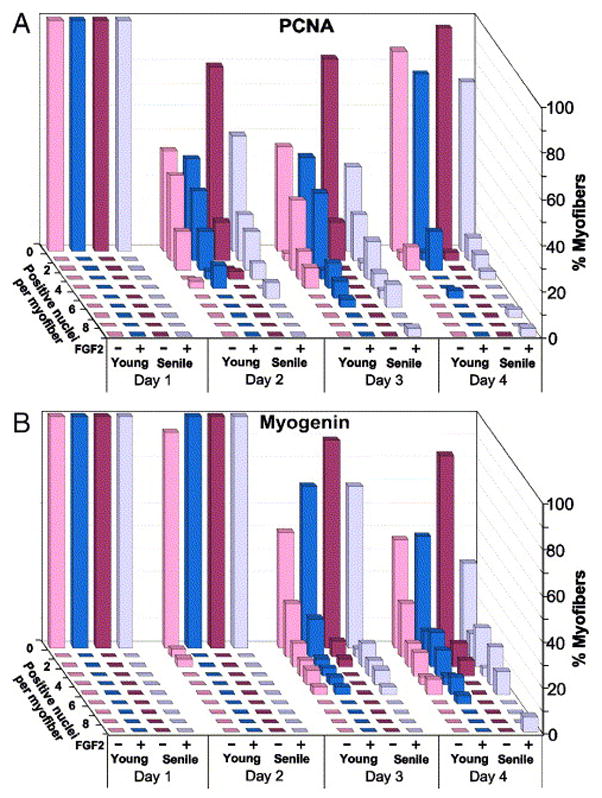
Kinetics of proliferation and differentiation of satellite cells in single FDB myofibers. Myofibers were isolated in parallel from young and senile muscle (two mice each) and cultured in basal medium (± FGF2). Parallel cultures were quantified for the number of PCNA+ cells (panel A) or myogenin+ cells (panel B) to identify proliferating and differentiating satellite cells. In order to specifically investigate the potency of myofibers containing satellite cells, arbitrary myofibers were first inspected for the presence of MAPK+ cells and the distribution data were then compiled (30–40 myofibers per time point). In this 3-dimensional data representation, each row of bars represents the distribution of myofibers with a specific number of satellite cells within a time point. The X-axis identifies the time in culture (days), the age of the donor mice, and whether (+/−) FGF was added. The Y-axis depicts the percent of myofibers
Discussion
This study evaluated age-related changes in satellite cells. Specifically, we investigated the size and performance of the satellite-cell pool by employing model systems that permit satellite cells to display their potential to proliferate and differentiate. The present comparison of satellite cell dynamics in the fast-twitch EDL myofibers versus the slow-twitch soleus muscle permitted insight into the relationships between age-dependent satellite-cell pool size and the extent of myofiber atrophy in different muscle types.
Aging is a process of decreased functioning that develops throughout an individual’s life (Florez-Duquet and McDonald, 1998; Troen, 2003). Accordingly, we analyzed age ranges from maturity to senility, which facilitated comprehensive insight into aging as a dynamic process. Our data show that the satellite cell pool is exhausted with age in both fast and slow muscles, resulting in the accumulation of myofibers that do not contain satellite cells. However, the remaining satellite cells maintain their potential to respond to growth promoting cues, undergo differentiation, fuse into myotubes and give rise to reserve cells throughout life. Moreover, this study establishes that the age-related deterioration in the capacity of the myofiber unit to support satellite cell amplification can be alleviated upon addition of growth-promoting agents such as FGF2. We conclude that although satellite cell performance can be rejuvenated by supplementing their micro-environment with growth-promoting agents, the low number of satellite cells may become the bottleneck of myofiber maintenance during aging.
The size of the satellite-cell pool declines with age
In this study, satellite cells were quantified along the entire length of individual myofibers, based on monitoring Pax7-expressing cells in a large sample size. Most previous reports on age-related changes in the satellite cell pool have relied on electron microscopy to identify satellite cells in muscle sections (Gibson and Schultz, 1983; Kadi et al., 2004; Roth et al., 2000). In these reports, satellite cell quantification has been represented as ratios between the number of satellite cells and a variety of factors such as the number of myonuclei in the same microscopic fields, myofiber diameter, and myofiber unit length. In the absence of a common denominator, discrepancies in the reported density of satellite cells per myofiber often arise even when analyzing the same muscles. In other studies, the number of progeny emanating from myofibers was used as a measure for the initial number of satellite cells; however, this approach has not taken into consideration the possibility that not all satellite cells emigrated from the myofibers and/or synchronously entered proliferation (Bockhold et al., 1998). Here, we determined the absolute number of satellite cells – an approach that permitted accurate fingerprinting of satellite cell profiles within and among muscles of different ages.
We show that satellite cell numbers in myofibers from both EDL and soleus muscle severely decreased and the frequency of myofibers lacking or having just one satellite cell significantly increased towards the end of the mouse lifespan. This “end of life crisis” in satellite-cell pool size may reflect an overall deterioration in body systems. However, before the onset of this “crisis”, there are clear differences in the age-associated pattern of satellite cell loss between EDL and soleus muscles. In EDL muscle, a major decline in the number of satellite cells occurred by 1 year of age. Conversely, in the soleus muscle, a drastic decrease in satellite cell numbers occurs only near 2.5 years of age. Furthermore, while a significant number of myofibers lacking satellite cells was apparent in the EDL muscle of the old-age group; such myofibers were present in the soleus muscle only at the senile stage. Altogether, the dynamics of age-linked satellite cell loss in EDL and soleus muscles are in accordance with established fiber-type dependent atrophy that occurs during muscle aging, reflecting a potential role for satellite cell abundance in the differential atrophy of fast and slow muscles.
Dual-immunolabeling with an antibody against syndecan-4 and MyoD was recently used as a means to quantify satellite cells in TA myofibers (Brack et al., 2005). Images from that study indicated that many additional entities in freshly-isolated TA myofibers, which seemed to be myofiber nuclei, were dually-labeled for syndecan-4 and MyoD. It is thus unclear how the distinction between satellite cells and myofiber nuclei was achieved. Indeed, we noted syndecan-4 expression associated with myonuclei when studying freshly-isolated myofibers. Nevertheless, many of these nuclei were not co-immunolabeled with Pax7 or within MAPK+ cells (G. Shefer and Z. Yablonka-Reuveni, unpublished; antibody was kindly provided by Dr. A. Rapraeger, University of Wisconsin, Madison). Therefore, we did not utilize syndecan-4 expression as a general and unequivocal marker of satellite cells. Also, since only infrequent satellite cells were positive for MyoD in freshly-isolated myofibers, we do not recognize MyoD expression as a general marker of quiescent satellite cells in such freshly-isolated myofibers. In contrast, Pax7-immunolabeling showed an excellent match between our data and those of Collins et al. (2005) where myofibers from the young-age group were studied. Collectively, we conclude that Pax7 immunolabeling provides a highly reliable means for identification of satellite cells in isolated myofibers from limb muscles, regardless the age of the animal being studied.
Satellite cell amplification, but not differentiation potential, declines with age
Based on a comprehensive investigation of satellite cells in several culture systems we conclude that progeny of satellite cells of all ages maintains the potential to proliferate, differentiate and fuse into myotubes, as well as the potential to give rise to reserve (Pax7+/MyoD−) cells.
In the FDB culture system, where myofibers were maintained in basal medium, proliferation of satellite cells of senile origin was greatly reduced unless the cultures were supplemented with FGF2. Hence, the aging myofiber unit does not produce sufficient growth-promoting agents to sustain satellite cell proliferation. The impaired capacity of aging muscle to support satellite cell recruitment/proliferation was suggested to be a consequence of a decline in IGF-I (Barton-Davis et al., 1998; Chakravarthy et al., 2000; Musaro et al., 2001) or a reduction in notch signaling (Conboy et al., 2003; Conboy and Rando, 2002). The present study introduces a novel insight into the impact of FGF depletion in the aging muscle.
Early studies of satellite cells isolated from 1-year old rats by enzymatic digestion, proposed that a decline in available hepatocyte growth factor (HGF) is responsible for the age-linked proliferative-latency of satellite cells and that this latency could be overcome by the addition of HGF but not FGF. These latter studies suggest that HGF promotes upregulation of FGF receptors (Allen et al., 1995; Johnson and Allen, 1995). We have demonstrated a similar promoting effect of FGF and HGF on satellite cell proliferation in myofibers both from young rats (2–3 month old; Kastner et al., 2000) and from adult rats (9–12 month old (Yablonka-Reuveni et al., 1999b). Hence, both FGF and HGF may regulate satellite cell recruitment in the younger and the older rodents. However, the FGF receptor system may be more sensitive to the enzymatic and physical trituration steps involved in satellite cell isolation, resulting in the reported preferential proliferative response to HGF (compared to FGF) in primary cultures of freshly isolated satellite cells. Differently, the gentler procedure of myofiber isolation may preserve the FGF receptors. Collectively, the present study reinforces our earlier conclusion that the FGF system indeed plays a role in satellite cell recruitment and amplification.
There was a time lag in the accumulation of progeny in primary cultures from senile muscle in comparison to those from young muscle, even though cultures initially had the same cell density and were grown in potent growth medium that contained high levels of growth-promoting agents, including FGF2 (Yablonka-Reuveni, 1995). Conversely, proliferation of satellite cells from the senile muscle occurred in a single bout in the FDB myofiber culture system when supplemented with FGF2. Together, these studies establish that the association with the parent myofiber provides a competent environment for satellite cell activation and cell cycle progression, regardless of age. Such a promoting environment may be a result of paracrine interplay between the parent myofiber and its resident cells. Nevertheless, it is possible that the efficiency of the proliferative machinery is reduced with age, and this decline, combined with the impact of the enzymatic digestion required for satellite cell isolation, may lengthen the recovery time upon cell culturing in the primary culture model. This scenario may account for the reduced synchronization of senile muscle satellite cell proliferation during early days in culture, as observed in this study. Clearly, the present study has not addressed the possibility that the rich growth medium used in the primary culture model may contain a factor that can selectively delay proliferation of satellite cells from senile muscle and this issue is avoided in FDB cultures where a different medium is being used. We do not favor the latter possibility because the proliferative rate is reduced not only when comparing senile to young groups but also when comparing adult to young groups during early days in primary cultures (Allen et al., 1995; our unpublished results); thus, proliferative latency is not unique to the senile age. Notably, myofiber injuries resulting from day-to-day muscle utilization do not involve detachment of satellite cells from their natural niche (as when establishing primary cultures). Therefore, the FDB myofiber system may better represent the proliferative potential of satellite cells in adult and aging muscle.
Present results with the FDB system indicate the proliferation of satellite cells of senile origin can be as efficient as that of satellite cells from young muscle, when proper growth-promoting agents are provided. While the present investigation focused on FGF, other growth factors known to promote satellite cell proliferation may also enhance the mitogenic milieu in aging muscle. A mere analysis of types and levels of growth factor (FGF isoforms included) expressed in old muscle will not necessarily provide a clear insight into the role of growth-promoting factors in aging muscle. For example, while IGF-I overexpression reduces age-linked myofiber atrophy, IGF-I expression level is not reduced in aging muscle; IGF-I signaling can be impaired by cytokines and the latter are known to increase systemically with age (Barton-Davis et al., 1998; Chakravarthy et al., 2000; Edstrom and Ulfhake, 2005; Grounds, 2002; Musaro et al., 2001). Studies with genetic models of inducible growth factor over/under expression are now required to evaluate the effect of growth promoting agents on enhancing satellite cell function during aging.
The present clonal study indicates that clonable satellite cells preserve their amplification and differentiation potential throughout life. Conversely, it has been suggested that the proliferative capacity of satellite cells from old muscle is impaired, based on the smaller number of cells that developed in satellite cell clones from old muscle as compared to clones from young muscle (Schultz and Lipton, 1982). However, the number of cells per clone was studied only during the early days in culture; therefore, a lag in the onset of proliferation, rather than an inherent reduced proliferative capacity, may account for the smaller clones derived from old muscle. Indeed, our analysis of clones at later time points (i.e., 10–14 days in culture) did not reveal apparent differences in clonal growth or in the degree of fusion into myotubes regardless of the age of the donor mouse. Furthermore, RT-PCR analysis of these clones did not reveal any apparent differences in expression levels of muscle-specific transcription factors, structural proteins or various growth factors shown to regulate myogenesis (ongoing studies). In another study reporting on the age-associated decline in clonal size, cultures were expanded long-term, prior to the establishment of clones (Chakravarthy et al., 2000). Therefore, the identity of the subpopulation of cells that founded these clones could not be clearly evaluated since they were not derived directly from satellite cells (Machida et al., 2004). Furthermore, nearly all cell culture studies on the differentiation of satellite cells from aging muscles, have relied on cells that were propagated for long-term (i.e., passaged cells), in order to amplify cell numbers for morphological and biochemical analysis (e.g., Beggs et al., 2004; Chakravarthy et al., 2000; Lees et al., 2005; Taylor-Jones et al., 2002). This propagation process may not only result in reduced myogenicity of satellite-cell progeny (Machida et al., 2004), but also preferentially enhance the growth of non-myogenic cells. The presence of non-myogenic cells can bias conclusions about age-dependent properties of satellite-cell progeny.
Collectively, our results show that, although satellite cell amplification is impaired within the aging muscle, satellite cell functioning can be rejuvenated when the cells receive proper growth-promoting signals.
Satellite cell performance in aging muscle: a proposed mechanism for the decline in satellite-cell pool size
The present study supports the conclusion that both the myofiber’s growth-promoting capacity and satellite cell abundance decline during aging. The deterioration of these two elements may together underlie age-associated impairment of muscle maintenance and repair. Most likely, in the context of sarcopenia every functional satellite cell is critical for the repair of subtle and focal damages that occur in normal day-to-day muscle utilization. However, more studies are required to determine if there is a causal link between the decline in satellite cell numbers with age and sarcopenia. Age-linked muscle atrophy was reduced when the muscle environment was modified by manipulating the level of the growth regulators IGF-I or myostatin (Barton-Davis et al., 1998; Chakravarthy et al., 2000; Musaro et al., 2001; Wagner et al., 2005). Although enhanced satellite cell potency was suggested to underlie the improved muscle quality, these rodent studies did not establish such a direct connection. We aim to investigate age-linked dynamics of satellite cell numbers in models of decreased sarcopenia. This type of research would further define the interplay between muscle mass and satellite cell numbers. Based on the present study, investigations on means to reduce the age-associated decline in the size of the satellite cell pool should focus on the first phase, which occurs in EDL myofibers by 1 year of age, and the second phase of a further increase in myofibers lacking satellite cells, which occurs in EDL myofibers by 1.5–2 year of age. Hence, such studies may not necessarily require the analysis of very old mice.
We demonstrate here that the inherent propensity of satellite cells to undergo proliferation and differentiation is maintained at all ages. Therefore, it is probable that fortifying the aging environment with the right growth-promoting agents would encourage satellite cells to enter myogenesis, resulting in improved muscle quality throughout life. The present study shows that satellite cell progeny emanating from single myofibers or developing in primary cultures are able to progress through the different compartments of myogenesis to eventually yield myotubes. Notably, satellite cells of senile muscle contribute reserve cells (Pax7-expressing cells that are negative for MyoD) similarly to satellite cells of young muscle. Ongoing work in our laboratory (K. Day and Z. Yablonka-Reuveni) has demonstrated that the reserve cells (Pax7+/MyoD−) consist of two populations: i) quiescent cells that may be equivalent to satellite cells, and ii) proliferating cells that may be equivalent to Pax7-expressing cells identified in developing muscle. The latter cells end up in the satellite cell niche (Kassar-Duchossoy et al., 2005; Relaix et al., 2005). Thus, reserve cells observed in our studies seem to represent distinct phenotypic compartments, which are possible intermediates of satellite cells. Present observations that reserve cells are produced in culture regardless of age underscore the importance of these cells with regard to the myogenic program and further point to an inherent capacity of satellite cells to execute the full myogenic program even near the natural end of an organism’s life.
While satellite cells retain their capacity to progress through the myogenic program regardless of age, it remains unclear whether satellite cells can execute such a complete myogenic program during ongoing muscle maintenance. It might be that satellite cell progeny are able to recapitulate the entire myogenic program of proliferation, differentiation and renewal only as a consequence of overt muscle injury. This capacity has been shown recently in adult muscle upon transplantation of isolated myofibers to injured host muscle (Collins et al., 2005). However, it has not been proven that the satellite cell pool can be replenished (by self-renewal or from non-satellite cell sources) during normal muscle utilization throughout life. Asymmetric cell division was proposed as a means to renew satellite cells during the active growth phase of postnatal muscle (Moss and Leblond, 1971). Yet, a follow-up study of growing muscle noted only minimal numbers of satellite cells that underwent more than one round of cell division before fusion with myofibers. Moreover, the number of repetitive proliferation rounds was limited (Schultz, 1996). Hence, it seems that there is minimal evidence for continuous renewal of the satellite cell pool even during the normal growth phase.
We propose that while satellite cells retain myogenic stem cell function, they are not adequately replenished during normal muscle utilization. As a result, the satellite cell pool is exhausted with age. Based on the emergence of the reserve cell phenotype (Pax7+/MyoD−) only after Pax7+/MyoD+ cells are present (seen also in chicken clonal studies, Halevy et al., 2004), we propose the model depicted in Fig. 9, where satellite cell renewal would require proliferative activity of satellite cell progeny. Presently, we are focusing on establishing in vivo model systems that permit satellite cell renewal throughout life or, alternatively, lead to premature elimination of satellite cells. Availability of such models will enable better understanding of the role of satellite cells in myofiber maintenance in adult and aging muscle.
Fig. 9.
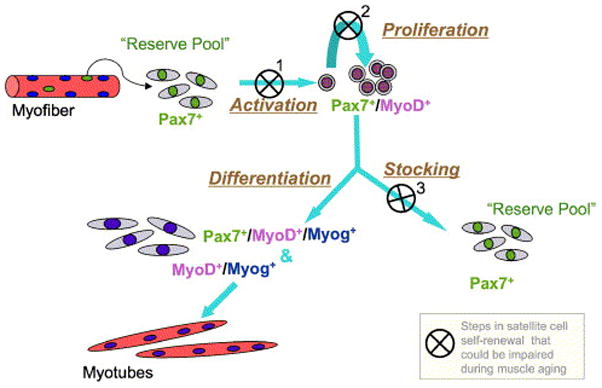
A model depicting possible mechanism(s) for age-linked depletion of satellite cells based on impaired self-renewal. Activation and entry of satellite cells into a proliferative state is essential for renewal of the stem cell pool. This renewal may be impaired in the aging muscle when the original satellite cells fuse with existing myofibers or with each other without producing replacement cells for satellite cell re-stocking. The latter process may occur when: 1. the original satellite cell cannot be activated to express MyoD+ (first step in the model); and/or 2. if satellite cells fail to undergo at least minimal clonal expansion by proliferation (second step in the model), and/or 3. when satellite cells do proliferate but reserve cells are not formed (third step in the model). Impaired efficiency of the three steps identified in the model might be inflicted by the aging environment and not necessarily due to inherent changes in the cells themselves.
Acknowledgments
We express gratitude to Drs. Peter Rabinovitch, Norman Wolf and Warren Ladigen, and their team members (University of Washington) for generously providing or sharing mice throughout the study. We are also grateful to Drs. Michael Rudnicki (Ottawa Health Research Institute) and Shahragim Tajbakhsh (Pasteur Institute) for the Myf5nlacZ/+ mouse strain.
This work was supported by grants to Z.Y.-R. from the National Institute on Aging (AG21566 and AG13798) and the USDA Cooperative State Research, Education and Extension Service (NRI, 99-35206-7934).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Agbulut O, Noirez P, Beaumont F, Butler-Browne GS. Myosin heavy chain isoforms in postnatal muscle development of mice. Biol Cell. 2003;95:399–406. doi: 10.1016/s0248-4900(03)00087-x. [DOI] [PubMed] [Google Scholar]
- Allen DL, Harrison BC, Sartorius C, Byrnes WC, Leinwand LA. Mutation of the IIB myosin heavy chain gene results in muscle fiber loss and compensatory hypertrophy. Am J Physiol, Cell Physiol. 2001;280:C637–664. doi: 10.1152/ajpcell.2001.280.3.C637. [DOI] [PubMed] [Google Scholar]
- Allen RE, McAllister PK, Masak KC. Myogenic potential of satellite cells in skeletal muscle of old rats. A brief note Mech Ageing Dev. 1980;13:105–109. doi: 10.1016/0047-6374(80)90053-6. [DOI] [PubMed] [Google Scholar]
- Allen RE, Sheehan SM, Taylor RG, Kendall TL, Rice GM. Hepatocyte growth factor activates quiescent skeletal muscle satellite cells in vitro. J Cell Physiol. 1995;165:307–312. doi: 10.1002/jcp.1041650211. [DOI] [PubMed] [Google Scholar]
- Alnaqeeb MA, Goldspink G. Changes in fibre type, number and diameter in developing and ageing skeletal muscle. J Anat. 1987;153:31–45. [PMC free article] [PubMed] [Google Scholar]
- Barton-Davis ER, Shoturma DI, Musaro A, Rosenthal N, Sweeney HL. Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muscle function. Proc Natl Acad Sci USA. 1998;95:15603–1567. doi: 10.1073/pnas.95.26.15603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beauchamp JR, Heslop L, Yu DS, Tajbakhsh S, Kelly RG, Wernig A, Buckingham ME, Partridge TA, Zammit PS. Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. J Cell Biol. 2000;151:1221–1234. doi: 10.1083/jcb.151.6.1221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beggs ML, Nagarajan R, Taylor-Jones JM, Nolen G, Macnicol M, Peterson CA. Alterations in the TGFbeta signaling pathway in myogenic progenitors with age. Aging Cell. 2004;3:353–361. doi: 10.1111/j.1474-9728.2004.00135.x. [DOI] [PubMed] [Google Scholar]
- Bockhold KJ, Rosenblatt JD, Partridge TA. Aging normal and dystrophic mouse muscle: analysis of myogenicity in cultures of living single fibers. Muscle Nerve. 1998;21:173–183. doi: 10.1002/(sici)1097-4598(199802)21:2<173::aid-mus4>3.0.co;2-8. [DOI] [PubMed] [Google Scholar]
- Brack AS, Bildsoe H, Hughes SM. Evidence that satellite cell decrement contributes to preferential decline in nuclear number from large fibres during murine age-related muscle atrophy. J Cell Sci. 2005;118:4813–4821. doi: 10.1242/jcs.02602. [DOI] [PubMed] [Google Scholar]
- Brooks SV, Faulkner JA. Skeletal muscle weakness in old age: underlying mechanisms. Med Sci Sports Exerc. 1994;26:432–439. [PubMed] [Google Scholar]
- Brooks SV, Faulkner JA. The magnitude of the initial injury induced by stretches of maximally activated muscle fibres of mice and rats increases in old age. J Physiol. 1996;497:573–580. doi: 10.1113/jphysiol.1996.sp021790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlson BM, Dedkov EI, Borisov AB, Faulkner JA. Skeletal muscle regeneration in very old rats. J Gerontol A Biol Sci Med Sci. 2001;56:B224–233. doi: 10.1093/gerona/56.5.b224. [DOI] [PubMed] [Google Scholar]
- Carlson BM, Faulkner JA. Muscle transplantation between young and old rats: age of host determines recovery. Am J Physiol. 1989;256:C1262–C1266. doi: 10.1152/ajpcell.1989.256.6.C1262. [DOI] [PubMed] [Google Scholar]
- Cartee GD. What insights into age-related changes in skeletal muscle are provided by animal models? J Gerontol A Biol Sci Med Sci. 1995;50:137–141. doi: 10.1093/gerona/50a.special_issue.137. [DOI] [PubMed] [Google Scholar]
- Chakravarthy MV, Davis BS, Booth FW. IGF-I restores satellite cell proliferative potential in immobilized old skeletal muscle. J Appl Physiol. 2000;89:1365–1379. doi: 10.1152/jappl.2000.89.4.1365. [DOI] [PubMed] [Google Scholar]
- Collins CA, Olsen I, Zammit PS, Heslop L, Petrie A, Partridge TA, Morgan JE. Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell. 2005;122:1–3. doi: 10.1016/j.cell.2005.05.010. [DOI] [PubMed] [Google Scholar]
- Conboy IM, Conboy MJ, Smythe GM, Rando TA. Notch-mediated restoration of regenerative potential to aged muscle. Science. 2003;302:1575–1577. doi: 10.1126/science.1087573. [DOI] [PubMed] [Google Scholar]
- Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weissman IL, Rando TA. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature. 2005;433:760–764. doi: 10.1038/nature03260. [DOI] [PubMed] [Google Scholar]
- Conboy IM, Rando TA. The regulation of Notch signaling controls satellite cell activation and cell fate determination in postnatal myogenesis. Dev Cell. 2002;3:397–409. doi: 10.1016/s1534-5807(02)00254-x. [DOI] [PubMed] [Google Scholar]
- Dhawan J, Rando TA. Stem cells in postnatal myogenesis: molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends Cell Biol. 2005;15:666–673. doi: 10.1016/j.tcb.2005.10.007. [DOI] [PubMed] [Google Scholar]
- Edstrom E, Ulfhake B. Sarcopenia is not due to lack of regenerative drive in senescent skeletal muscle. Aging Cell. 2005;4:65–77. doi: 10.1111/j.1474-9728.2005.00145.x. [DOI] [PubMed] [Google Scholar]
- Florez-Duquet M, McDonald RB. Cold-Induced Thermoregulation and Biological Aging. Physiol Rev. 1998;78:339–358. doi: 10.1152/physrev.1998.78.2.339. [DOI] [PubMed] [Google Scholar]
- Gallegly JC, Turesky NA, Strotman BA, Gurley CM, Peterson CA, Dupont-Versteegden EE. Satellite cell regulation of muscle mass is altered at old age. J Appl Physiol. 2004;97:1082–1090. doi: 10.1152/japplphysiol.00006.2004. [DOI] [PubMed] [Google Scholar]
- Gibson MC, Schultz E. Age-related differences in absolute numbers of skeletal muscle satellite cells. Muscle Nerve. 1983;6:574–580. doi: 10.1002/mus.880060807. [DOI] [PubMed] [Google Scholar]
- Goodrick CL. Lifespan and the inheritance of longevity of inbred mice. J Gerontol. 1975;30:257–263. doi: 10.1093/geronj/30.3.257. [DOI] [PubMed] [Google Scholar]
- Grounds MD. Age-associated changes in the response of skeletal muscle cells to exercise and regeneration. Ann N Y Acad Sci. 1998;854:78–91. doi: 10.1111/j.1749-6632.1998.tb09894.x. [DOI] [PubMed] [Google Scholar]
- Grounds MD. Reasons for the degeneration of ageing skeletal muscle: a central role for IGF-1 signalling. Biogerontology. 2002;3:19–24. doi: 10.1023/a:1015234709314. [DOI] [PubMed] [Google Scholar]
- Grounds MD, Yablonka-Reuveni Z. Molecular and cell biology of skeletal muscle regeneration. Mol Cell Biol Hum Dis Ser. 1993;3:210–256. doi: 10.1007/978-94-011-1528-5_9. [DOI] [PubMed] [Google Scholar]
- Halevy O, Piestun Y, Allouh MZ, Rosser BW, Rinkevich Y, Reshef R, Rozenboim I, Wleklinski-Lee M, Yablonka-Reuveni Z. Pattern of Pax7 expression during myogenesis in the posthatch chicken establishes a model for satellite cell differentiation and renewal. Dev Dyn. 2004;231:489–502. doi: 10.1002/dvdy.20151. [DOI] [PubMed] [Google Scholar]
- Hawke TJ, Garry DJ. Myogenic satellite cells: physiology to molecular biology. J Appl Physiol. 2001;91:534–551. doi: 10.1152/jappl.2001.91.2.534. [DOI] [PubMed] [Google Scholar]
- Johnson SE, Allen RE. Activation of skeletal muscle satellite cells and the role of fibroblast growth factor receptors. Exp Cell Res. 1995;219:449–453. doi: 10.1006/excr.1995.1251. [DOI] [PubMed] [Google Scholar]
- Kadi F, Charifi N, Denis C, Lexell J. Satellite cells and myonuclei in young and elderly women and men. Muscle Nerve. 2004;29:120–127. doi: 10.1002/mus.10510. [DOI] [PubMed] [Google Scholar]
- Karakelides H, Nair KS. Sarcopenia of aging and its metabolic impact. Curr Top Dev Biol. 2005;68:123–148. doi: 10.1016/S0070-2153(05)68005-2. [DOI] [PubMed] [Google Scholar]
- Kassar-Duchossoy L, Giacone E, Gayraud-Morel B, Jory A, Gomes D, Tajbakhsh S. Pax3/Pax7 mark a novel population of primitive myogenic cells during development. Genes Dev. 2005;19:1426–1431. doi: 10.1101/gad.345505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kastner S, Elias MC, Rivera AJ, Yablonka-Reuveni Z. Gene expression patterns of the fibroblast growth factors and their receptors during myogenesis of rat satellite cells. J Histochem Cytochem. 2000;48:1079–1096. doi: 10.1177/002215540004800805. [DOI] [PubMed] [Google Scholar]
- Kunstyr I, Leuenberger HGW. Gerontological data on C57BL/6 mice. I Sex differences in survival curves. J Gerontol. 1975;30:157–162. doi: 10.1093/geronj/30.2.157. [DOI] [PubMed] [Google Scholar]
- Lees SJ, Rathbone CR, Booth FW. Age-Associated Decrease in Muscle Precursor Cell Differentiation. Am J Physiol, Cell Physiol. 2005;290:C609–615. doi: 10.1152/ajpcell.00408.2005. [DOI] [PubMed] [Google Scholar]
- Lexell J. Human aging, muscle mass, and fiber type composition. J Gerontol A Biol Sci Med Sci. 1995;50:11–16. doi: 10.1093/gerona/50a.special_issue.11. [DOI] [PubMed] [Google Scholar]
- Machida S, Spangenburg EE, Booth FW. Primary rat muscle progenitor cells have decreased proliferation and myotube formation during passages. Cell Prolif. 2004;37:267–277. doi: 10.1111/j.1365-2184.2004.00311.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mewissen D. Natural tumor incidence in a population of mice as a reference index. Fed Proc. 1971;30:311. [Google Scholar]
- Moss FP, Leblond CP. Satellite cells as the source of nuclei in muscles of growing rats. Anat Rec. 1971;170:421–435. doi: 10.1002/ar.1091700405. [DOI] [PubMed] [Google Scholar]
- Mouly V, Aamiri A, Bigot A, Cooper RN, Di Donna S, Furling D, Gidaro T, Jacquemin V, Mamchaoui K, Negroni E, Perie S, Renault V, Silva-Barbosa SD, Butler-Browne GS. The mitotic clock in skeletal muscle regeneration, disease and cell mediated gene therapy. Acta Physiol Scand. 2005;184:3–15. doi: 10.1111/j.1365-201X.2005.01417.x. [DOI] [PubMed] [Google Scholar]
- Musaro A, McCullagh K, Paul A, Houghton L, Dobrowolny G, Molinaro M, Barton ER, Sweeney HL, Rosenthal N. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat Genet. 2001;27:195–200. doi: 10.1038/84839. [DOI] [PubMed] [Google Scholar]
- Musaro A, Rosenthal N. Transgenic mouse models of muscle aging. Exp Gerontol. 1999;34:147–156. doi: 10.1016/s0531-5565(98)00079-5. [DOI] [PubMed] [Google Scholar]
- Nikulina EM, Skrinskaya JA, Popova NK. Role of genotype and dopamine receptors in behaviour of inbred mice in a forced swimming test. Psychopharmacology. 1991;105:525–529. doi: 10.1007/BF02244374. [DOI] [PubMed] [Google Scholar]
- Relaix F, Rocancourt D, Mansouri A, Buckingham M. A Pax3/Pax7-dependent population of skeletal muscle progenitor cells. Nature. 2005;435:948–953. doi: 10.1038/nature03594. [DOI] [PubMed] [Google Scholar]
- Renault V, Thornell LE, Eriksson PO, Butler-Brown G, Mouly V. Regenerative potential of human skeletal muscle during aging. Aging Cell. 2002;1:132–139. doi: 10.1046/j.1474-9728.2002.00017.x. [DOI] [PubMed] [Google Scholar]
- Roth SM, Martel GF, Ivey FM, Lemmer JT, Metter EJ, Hurley BF, Rogers MA. Skeletal muscle satellite cell populations in healthy young and older men and women. Anat Rec. 2000;260:351–358. doi: 10.1002/1097-0185(200012)260:4<350::AID-AR30>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- Schultz E. Satellite cell proliferative compartments in growing skeletal muscles. Dev Biol. 1996;175:84–94. doi: 10.1006/dbio.1996.0097. [DOI] [PubMed] [Google Scholar]
- Schultz E, Lipton BH. Skeletal muscle satellite cells: changes in proliferation potential as a function of age. Mech Ageing Dev. 1982;20:377–383. doi: 10.1016/0047-6374(82)90105-1. [DOI] [PubMed] [Google Scholar]
- Scott W, Stevens J, Binder-Macleod SA. Human skeletal muscle fiber type classifications. Phys Ther. 2001;81:1810–1816. [PubMed] [Google Scholar]
- Seale P, Sabourin LA, Girgis-Gabardo A, Mansouri A, Gruss P, Rudnicki MA. Pax7 is required for the specification of myogenic satellite cells. Cell. 2000;102:777–786. doi: 10.1016/s0092-8674(00)00066-0. [DOI] [PubMed] [Google Scholar]
- Shefer G, Wleklinski-Lee M, Yablonka-Reuveni Z. Skeletal muscle satellite cells can spontaneously enter an alternative mesenchymal pathway. J Cell Sci. 2004;117:5393–5404. doi: 10.1242/jcs.01419. [DOI] [PubMed] [Google Scholar]
- Shefer G, Yablonka-Reuveni Z. Isolation and culture of skeletal muscle myofibers as a means to analyze satellite cells. Methods Mol Biol. 2005;290:281–304. doi: 10.1385/1-59259-838-2:281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storer JB. Longevity and gross pathology at death in 22 inbred strains of mice. J Gerontol. 1966;21:404–409. doi: 10.1093/geronj/21.3.404. [DOI] [PubMed] [Google Scholar]
- Taylor-Jones JM, McGehee RE, Rando TA, Lecka-Czernik B, Lipschitz DA, Peterson CA. Activation of an adipogenic program in adult myoblasts with age. Mech Ageing Dev. 2002;123:649–661. doi: 10.1016/s0047-6374(01)00411-0. [DOI] [PubMed] [Google Scholar]
- Troen BR. The biology of aging. Mt Sinai J Med. 2003;70:3–22. [PubMed] [Google Scholar]
- Wagner KR, Liu X, Chang X, Allen RE. Muscle regeneration in the prolonged absence of myostatin. Proc Natl Acad Sci USA. 2005;102:2519–2524. doi: 10.1073/pnas.0408729102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z. Myogenesis in the chicken: the onset of differentiation of adult myoblasts is influenced by tissue factors. Basic Appl Myol (BAM) 1995;5:33–42. [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z. Isolation and characterization of stem cells from adult skeletal muscle. In: Lanza R, Blau H, Melton D, Moore M, Thomas ED, Verfaillie C, Weissman I, West M, editors. Handbook of Stem Cells. Vol. 2. Elsevier-Academic Press; San Diego: 2004. pp. 571–580. [Google Scholar]
- Yablonka-Reuveni Z, Anderson JE. Satellite cells from dystrophic (mdx) mice display accelerated differentiation in primary cultures and in isolated myofibers. Dev Dyn. 2006;235:203–212. doi: 10.1002/dvdy.20602. [DOI] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, Rivera AJ. Temporal expression of regulatory and structural muscle proteins during myogenesis of satellite cells on isolated adult rat fibers. Dev Biol. 1994;164:588–603. doi: 10.1006/dbio.1994.1226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, Rivera AJ. Influence of PDGF-BB on proliferation and transition through the MyoD-myogenin-MEF2A expression program during myogenesis in mouse C2 myoblasts. Growth Factors. 1997a;15:1–27. doi: 10.3109/08977199709002109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, Rivera AJ. Proliferative dynamics and the role of FGF2 during myogenesis of rat satellite cells on isolated fibers. Basic Appl Myol (BAM) 1997b;7:176–189. [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, Rudnicki MA, Rivera AJ, Primig M, Anderson JE, Natanson P. The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Dev Biol. 1999a;210:440–455. doi: 10.1006/dbio.1999.9284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yablonka-Reuveni Z, Seger R, Rivera AJ. Fibroblast growth factor promotes recruitment of skeletal muscle satellite cells in young and old rats. J Histochem Cytochem. 1999b;47:23–42. doi: 10.1177/002215549904700104. [DOI] [PubMed] [Google Scholar]
- Zammit PS, Golding JP, Nagata Y, Hudon V, Partridge TA, Beauchamp JR. Muscle satellite cells adopt divergent fates: a mechanism for self-renewal? J Cell Biol. 2004;166:347–357. doi: 10.1083/jcb.200312007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zammit PS, Heslop L, Hudon V, Rosenblatt JD, Tajbakhsh S, Buckingham ME, Beauchamp JR, Partridge TA. Kinetics of myoblast proliferation show that resident satellite cells are competent to fully regenerate skeletal muscle fibers. Exp Cell Res. 2002;281:39–49. doi: 10.1006/excr.2002.5653. [DOI] [PubMed] [Google Scholar]