

Abstract
Background
It has been hypothesized that reduced axonal transport contributes to the degeneration of neuronal processes in Parkinson's disease (PD). Mitochondria supply the adenosine triphosphate (ATP) needed to support axonal transport and contribute to many other cellular functions essential for the survival of neuronal cells. Furthermore, mitochondria in PD tissues are metabolically and functionally compromised. To address this hypothesis, we measured the velocity of mitochondrial movement in human transmitochondrial cybrid "cytoplasmic hybrid" neuronal cells bearing mitochondrial DNA from patients with sporadic PD and disease-free age-matched volunteer controls (CNT). The absorption of low level, near-infrared laser light by components of the mitochondrial electron transport chain (mtETC) enhances mitochondrial metabolism, stimulates oxidative phosphorylation and improves redox capacity. PD and CNT cybrid neuronal cells were exposed to near-infrared laser light to determine if the velocity of mitochondrial movement can be restored by low level light therapy (LLLT). Axonal transport of labeled mitochondria was documented by time lapse microscopy in dopaminergic PD and CNT cybrid neuronal cells before and after illumination with an 810 nm diode laser (50 mW/cm2) for 40 seconds. Oxygen utilization and assembly of mtETC complexes were also determined.
Results
The velocity of mitochondrial movement in PD cybrid neuronal cells (0.175 +/- 0.005 SEM) was significantly reduced (p < 0.02) compared to mitochondrial movement in disease free CNT cybrid neuronal cells (0.232 +/- 0.017 SEM). For two hours after LLLT, the average velocity of mitochondrial movement in PD cybrid neurites was significantly (p < 0.003) increased (to 0.224 +/- 0.02 SEM) and restored to levels comparable to CNT. Mitochondrial movement in CNT cybrid neurites was unaltered by LLLT (0.232 +/- 0.017 SEM). Assembly of complexes in the mtETC was reduced and oxygen utilization was altered in PD cybrid neuronal cells. PD cybrid neuronal cell lines with the most dysfunctional mtETC assembly and oxygen utilization profiles were least responsive to LLLT.
Conclusion
The results from this study support our proposal that axonal transport is reduced in sporadic PD and that a single, brief treatment with near-infrared light can restore axonal transport to control levels. These results are the first demonstration that LLLT can increase axonal transport in model human dopaminergic neuronal cells and they suggest that LLLT could be developed as a novel treatment to improve neuronal function in patients with PD.
Background
Sporadic Parkinson's disease (PD) is a disabling, progressive neurodegenerative disease. The neuropathological characterization of PD includes not only the loss of dopaminergic neurons in the substantia nigra and other brainstem nuclei but also the presence of cytoplasmic inclusions that form Lewy bodies and Lewy neurites in surviving neurons. [1,2] Degeneration of nigral neurons begins at synaptic terminals and progresses retrograde to cell bodies before the onset of symptoms. [3] At symptom onset, 60–80% of the striatal dopaminergic terminals have already been lost. Dopaminergic nerve terminals continue to be lost at a rate of 10–12% a year. [4]
Neurons prone to degeneration in PD have axons that are long and poorly myelinated and provide a massive innervation to the striatum (150,000 presynaptic terminals per neuron). [5] Neurons have a high metabolic rate and require a large, uninterrupted supply of ATP. [6] Microtubule-based anterograde axonal transport is ATP-driven and responsible for the delivery of mitochondria and other cellular constituents to distal sites in neurons. Compromised retrograde axonal transport deprives the cell body of vital trophic factors and deprives axons and dendrites of synaptic vesicles, lysosomes and mitochondria. [5,7]
Reduced or compromised axonal transport could underlie the progressive, relentless loss of dopaminergic nerve terminals in sporadic PD. Axonal transport is one aspect of mitochondrial dynamics that is critical for the distribution of functional mitochondria to distal synaptic terminals. Movement, fission, and fusion of mitochondria are functions that are also essential for neuronal survival. [8] Studies at the ultrastructural level in human PD caudate and striatum detected a disruption in the distribution of both microtubules and organelles such as mitochondria. [9] Neurotoxins that are used to model PD pathogenesis (1-methyl-4-phenylpyridinium (MPP+) and rotenone) compromise mitochondrial function by inhibiting complex I of the mtETC. These toxins also have microtubule depolymerizing activities that alter axonal transport. [2,10-14] Complex I of the mtETC is structurally and functionally defective in brain, platelets and other tissues in PD patients. [15,16] Since mitochondrial DNA (mtDNA) encodes components of the mtETC, it is not surprising that mtDNA from PD patients contains increased levels of mutations and deletions. [15,17-19]
It is now known that genes linked to familial PD (α-synuclein, parkin and PINK1) play important roles in mitochondrial transport dynamics and function. [8,20] Pathogenic mutation, hyper-phosphorylation or over-expression of α-synuclein is associated with reduced axonal transport. [21] Liang et al [13] and Feng [22] suggested that the consequences of both genetic and environmental risk factors for familial and sporadic PD disrupt microtubules and axonal transport, leading to the death of dopaminergic neurons.
In an effort to explore the contribution of mitochondria to features of PD pathogenesis such as reduced axonal transport, we have focused our studies on "cytoplasmic hybrid" cybrid cell lines. Cybrid cell lines were created by the fusion of donated platelets from PD or CNT volunteers with human mtDNA-free (Rho0) SH-SY5Y human neuroblastoma or NT2 teratocarcinoma cells. [23,24] Cybrid cell lines have been used to explore the role of mitochondrial dysfunction in PD. [24-28] Using this model of PD pathogenesis, the contribution of mitochondria can be selectively studied and the variability due to nuclear genes and environmental factors can be controlled. [29]
Changes in PD cybrid cell lines correlate with changes seen in PD brain. [29] PD cybrid lines exhibit- A) decreased complex I activity,[26,28] B) increased production of reactive oxygen species (ROS),[26,28,30] C) increased levels of antioxidant enzymes,[26,31] D) increased numbers of morphologically abnormal mitochondria,[32] E) mtDNA with deletions (personal communication, Dr. Bradley Miller), and F) fibrillar and vesicular inclusions called cybrid Lewy bodies that replicate the essential antigenic and structural features of Lewy bodies in PD brain. [27]
Non-dividing, process-bearing, dopaminergic neuronal cells were generated from undifferentiated CNT and PD cybrids using a low concentration of staurosporine. [2,33] These PD and CNT cybrid neuronal cells contain neuronal cytoskeletal proteins (neuron-specific beta tubulin, tau, and microtubule associated protein 2) and dopaminergic markers (tyrosine hydroxylase, dopamine transporter, the dopamine D2 receptor and the vesicular monoamine transporter). [2] Our results show that the velocity of mitochondrial movement is significantly reduced in the processes of PD cybrid neuronal cells compared to CNT. Reduced movement of mitochondria in human PD cybrid neuronal cells supports the proposition that reduced axonal transport plays a role in PD progression.
In an effort to normalize this reduction in mitochondrial movement, we exposed PD and CNT cybrid neuronal cells to a low level near-infrared light therapy. In animal studies, low level near-infrared light therapy (LLLT) improved the outcome in retinal damage due to methanol or rotenone toxicity, [33-35] traumatic brain injury,[36] or spinal cord injury,[37] and induced analgesia in dorsal root ganglion neurons. [38] LLLT has improved the survival and function of rat neuronal cells exposed to MPP+ [39] or rotenone. [35] In the human brain, transcranial laser therapy (delivery of near-infrared laser light through the scalp and skull) has been used to successfully treat complex neurological conditions such as ischemic stroke. [40] Research supports the idea that near-infrared laser (750–1400 nm) light is absorbed and stimulates components in the mtETC, increases redox capacity, activates cytochrome c oxidase (complex IV), increases ATP production in dysfunctional cells and initiates long-term changes in neuronal function in human and animal studies. [37,40-43] Our results show that a single treatment with 810 nm laser light was sufficient to restore the velocity of mitochondrial movement to control levels in PD cybrid neuronal cells.
Methods
Patient population
The demographics for PD and CNT volunteers were described previously. [27] The Hoehn and Yahr[44] score for all PD volunteers was stage 2. The mean age for the PD group was 72.14 +/- 4.18SD and 64.4 +/- 5.68SD for the CNT group.
Cybrid growth and differentiation
PD and CNT cybrid cell lines were created by fusing platelets from sporadic PD or disease-free CNT individuals with mtDNA-free Rho0 cells created in SH-SY5Y neuroblastoma cells. [45] Blood samples were collected under an IRB-approved protocol, and details of cybrid creation were described previously. [27] PD and CNT cybrid, SH-SY5Y and Rho0 cell lines were grown in T75 tissue culture flasks (Greiner, PGC Scientific, Gaithersburg, MD) in a culture medium (CM) consisting of Dulbecco's modified Eagle's medium with high glucose (DMEM), 10% characterized fetal bovine serum (FBS, Hyclone, Logan, Utah), 100 μg/ml pyruvate, 50 μg/ml uridine, antibiotic-antimycotic, 100 Units/ml penicillin G, 100 μg/ml streptomycin, 0.25 μg/ml and amphotericin β (Invitrogen, Carlsbad, California, USA). [27]
Cybrid neuron differentiation
Glass bottomed 35 mm dishes (MatTek Corp., Ashland, MA) were washed with sterile H2O then treated with sterile 1 N HCl for 30 minutes, washed three times with sterile H2O and stored at room temperature (RT). Coverslip dishes were coated for 40 min.-overnight with 1.5 mg/ml poly-l-lysine (PL, Sigma-Aldrich, St. Louis, Missouri, USA) dissolved in sterile RT H2O. Dishes were rinsed twice with sterile RT H2O before adding the cell suspension.
Proliferating cybrid cells were harvested from flasks with 0.05% trypsin (Invitrogen, Carlsbad, CA) in phosphate buffered saline (PBS, Sigma-Aldrich, St. Louis, Missouri, USA) for 5 minutes at 37°C. Trypsin activity was stopped by an equal volume of CM. Cells were then centrifuged at 200 g for 5 minutes and re-suspended in CM. 40,000 cells in 2 ml CM were added to each 35 mM dish. After 18–24 hours, CM was removed and the differentiation media (DM, 500 ml of Neurobasal with 10 ml B27 supplements [Invitrogen, Carlsbad, CA] and 0.5 mM glutamine, pyruvate, uridine and antibiotic-antimycotic as described above). A 25 μM stock solution of staurosporine (ST, Sigma-Aldrich Corp, St Louis, Missouri, USA) was made in dimethyl sulfoxide, frozen at -80°C. Unused aliquots were discarded after 60 days. ST (4 nM-8 nM) dilutions were made fresh in DM and replaced every 2–3 days. Differentiation was completed on day 12.
Immunocytochemistry
The MitoProfile® OXPHOS/PDH immunocytochemistry kit (MitoSciences, Eugene, Oregon, USA) was used to detect defective assembly of ETC complexes I-V. The manufacturer's suggested protocol was modified for coverslip dishes. Vectashield mounting medium (Vector Labs, Burlingame, California, USA) was added to coverslip dish after staining. Images of stained cybrid neuronal cells were created using an Olympus Fluoview laser scanning confocal microscope system (Center Valley, Pennsylvania, USA). SYTO 61 (Invitrogen) at a concentration of 5 mM (30 min) was used to stain nucleic acids in the cell nucleus and cytoplasm.
High Resolution Oxygen Respirometry
A two chamber oxygraph O2k (Oroboros, Innsbruck, Austria) was used to measure respiration rates of dissociated CNT, PD and laser-treated CNT and PD cybrid neuronal cells according to manufacturers directions. After 12 days in DM, PD and CNT cybrid neuronal cells were harvested by rinsing with PBS. The cells (4.5 × 106 cells/ml resuspended in phenol-free DMEM and separated into two sterile 35 mm culture dishes) were kept on ice. In this blinded study, one dish received LLLT (see below) and the other dish remained untreated for each cybrid line studied. After the oxygraph was calibrated according to manufacturer's instructions, 2 ml of LLLT or untreated cybrid neuronal cells were added to each chamber. The chambers were closed and cells were equilibrated (37°C). After the basal rate of respiration was established, complex V respiration was inhibited by oligomycin (4 ug/ml final concentration). Respiration was uncoupled by carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP at a final concentration of ~0.325 uM, Sigma St Louis, MO). Complex 1 was inhibited by adding rotenone at a final concentration of 1.0 μM.
Axonal transport
To measure mitochondrial movement, cybrid neuronal cells were incubated with 50 nM MitoTracker CMXRos (MTRed; Invitrogen, Carlsbad, California, USA) for 10 min at 37°C. [46] Time-lapse recordings were made using an Olympus IX70 microscope equipped with epifluorescence and Nomarski optics, a Lambda 10-2 filter wheel, a Photometrics CoolSnap HQ progressive scan CCD camera and a heater/controller to maintain cybrid cells at 37°C during image collection (World Precision Instruments, Inc, Sarasota, Florida, USA). Collection of image stacks, and velocity measurements were made using MetaMorph Imaging System (Molecular Devices, Downingtown, Pennsylvania, USA). For standard recordings, images were collected every 3 seconds for 2 min. Unlike other studies, the velocity of all mitochondria in each cell process was tracked by hand whether they moved or not. The average velocity for individual mitochondria was calculated using intervals where movement occurred. [46] Neutral density filters are used to reduce illumination from the mercury lamp and minimize phototoxicity. [46]
LLLT with near-infrared 810 nm laser
Differentiated neuronal cells from cultures of six PD and five CNT cybrid lines as well as SH-SY5Y were labeled with MtRed in phenol-free DM and 10–15 movies (1–3 neurites per movie, 8–50 mitochondria per process) were collected per dish. A second dish from each line was switched to phenol-free DM and uniformly illuminated at 50 mW/cm2 (measured at the dish surface) with a continuous wave 810nm diode laser for 40 seconds (Acculaser, PhotoThera, Inc, Carlsbad, California, USA). The power output of the laser was measured before each LLLT experiment with an Ohir Optronics power meter (Jerusalem, Israel) to ensure consistent exposure. After LLLT, neuronal cells from six PD, five CNT and SH-SY5Y lines were stained with MTRed as described above. Movies (n = 10–15 movies per experimental condition, 2–3 processes/movie, 8–50 mitochondria per process) were collected at 5 to 10 minute intervals for at least 2 hours for each dish (n = 2–3 experiments +/- LLLT per cybrid line). T-tests assuming equal or unequal variance were used as needed to analyze the data.
Results
Mitochondrial movement in differentiated PD and CNT cybrid neurites
The processes selected for study in all conditions were comparable in length and diameter (Figure 1). The range of mitochondrial lengths in PD, CNT and LLLT-treated cybrids was not qualitatively different (Figure 1). Mitochondrial movement in PD, CNT and LLLT and untreated cybrid neurites was bidirectional but primarily anterograde and saltatory (see Additional file 1, Additional file 2, Additional file 3, Additional file 4). The number of mitochondria moving retrograde in any process was minimal. Consequently, velocity was measured irrespective of direction. Average velocities for individual mitochondria less than or equal to 0.075 μm/second were categorized as stationary. In CNT cybrids, 14% of the mitochondria were stationary during the recording period. The number of stationary mitochondria in PD cybrid neurites was elevated compared to CNT cybrid neurites (29%, see Table 1). As shown in Table 1 and Figure 2, the average velocity of mitochondrial movement in PD cybrid neurites (0.175 +/- 0.005SEM) was significantly less than the velocity in CNT cybrid neurites (0.232 +/- 0.017SEM). The total amount of time spent moving and the average distance traveled by mitochondria in PD cybrid neurites was reduced but not significantly different from CNT (Table 1, Figure 2) however, the total distance traveled by mitochondria in PD cybrid neurites was significantly reduced compared to CNT cybrid mitochondria (Figure 2).
Figure 1.
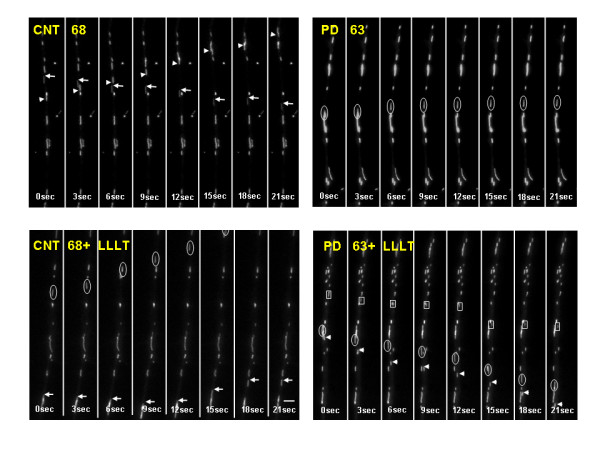
These montages illustrate the movements of MTRed-labeled mitochondria in the processes of CNT68, CNT68-90 minutes after LLLT, PD63 and PD63-85 minutes after LLLT. Moving mitochondria in each montage are indicated by arrowheads, arrows, circles and boxes. The images were taken every 3 seconds. Original movies are shown as additional files. Bar = 1 μm.
Table 1.
Data from studies of mitochondrial movement in LLLT and untreated PD and CNT cybrid neurons
|
PD (n = 6) |
SEM |
PD vs PD+LLLT p value |
CNT (n = 5) |
SEM |
PD vs CNT p value |
|
| Velocity (μm/sec) | 0.175 | 0.005 | 0.001 | 0.232 | 0.017 | 0.02 |
| Percent not moving (v<0.075) | 29 | 3.68 | 0.01 | 14 | 2.42 | 0.01 |
| Average distance traveled | 0.54 | 0.016 | 0.006 | 0.757 | 0.048 | NS |
| Total distance traveled | 9.92 | 0.261 | 0.004 | 17.09 | 1.18 | 0.004 |
| Time spent moving (out of 120 sec) | 45 | 0.358 | NS | 61 | 6.89 | NS |
| Number of neurites/cybrid | 24–53 | 26–44 | ||||
|
PD + LLLT (n = 6) |
SEM |
CNT + LLLT (n = 5) |
SEM |
CNT vs CNT+LLLT p value |
||
| LLLT-50 mW/cm2 for 40 sec (2J/cm2) | ||||||
| Velocity (μm/sec) | 0.224 | 0.02 | 0.226 | 0.017 | NS | |
| Percent not moving (v<0.075) | 12 | 2.15 | 16 | 2.15 | NS | |
| Average distance traveled | 0.682 | 0.033 | 0.782 | 0.065 | NS | |
| Total distance traveled | 12.78 | 0.616 | 16.34 | 1.27 | NS | |
| Time spent moving (out of 120 sec) | 48 | 3.94 | 63 | 6.92 | NS | |
| Number of neurites/cybrid | 25–51 | 28–58 | ||||
Significance was determined using a two-tailed t test assuming unequal variance.
Figure 2.
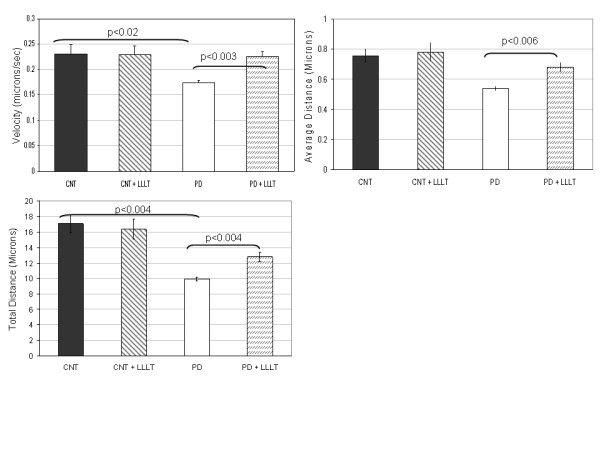
This figure contains three graphs. Graph 2A depicts the average velocity of mitochondrial movement in the processes of CNT (n = 5), CNT+LLLT (n = 5), PD (n = 6) and PD+LLLT (n = 6) cybrid neuronal cell lines. Using a two-tailed t-test assuming unequal variance, mitochondrial movement in PD cybrid neurites was significantly reduced compared to CNT. Mitochondrial movement in PD+LLLT cybrid neurites was significantly increased compared to PD. Mitochondrial movment in CNT cybrid neurites was not significantly different from CNT+LLLT. Differences in culture conditions, cell type, mitochondrial labeling method and method of velocity calculation make it difficult to compare mitochondrial velocities among published papers. Graph 2B depicts the average distance traveled by mitochondria during the recording period. The average distance traveled by mitochondria (calculated by averaging the distance traveled for each three second interval irrespective of direction of travel) in PD+LLLT was significantly increased above PD. Graph 2C depicts the total distance traveled (calculated by summing the distance traveled in each three second interval irrespective of direction of travel) during the recording period by mitochondria in each category. The total distance traveled by PD mitochondria was significantly reduced compared to CNT. The total distance traveled by PD+LLLT mitochondria were significantly increased compared to PD.
Effects of LLLT on mitochondrial movement in differentiated PD and CNT cybrid neurites
For at least 2 hours after exposure to LLLT (Table 1 and Figure 2), the velocity of mitochondrial movement in PD cybrid neurites was significantly increased and restored to CNT levels. This change in velocity was achieved by reducing the number of stationary mitochondria and increasing the average and total distance traveled (Table 1 and Figure 2). Further studies are needed to determine if more individual stationary mitochondria initiate movement after LLLT or if mitochondria already in motion exhibit increased velocity. The total time PD mitochondria spent moving after LLLT was not significantly different from CNT (Table 1). Of the six PD cybrid lines studied, the response of two lines (PD60, PD61) to LLLT was consistently increased but did not achieve significance in three separate studies. Mitochondrial movement in the five CNT cybrid lines also did not change significantly in response to LLLT (Table 1 and Figure 2).
mtETC assembly in PD, CNT and LLLT-treated cybrid neuronal cells
To better understand the role that mitochondrial dysfunction plays in mitochondrial movement, we double stained differentiated PD and CNT cybrid neuronal cells with antibodies to essential components of mtETC complexes I-V. The presence of staining with monoclonal antibodies directed against labile subunits of each complex (CI-NDUFB4, CII-30 kDa, CIII-Core 2 protein, CIV-subunit I, CV-OSCP) indicates that the complex is correctly assembled. [47] By double staining with antibodies to a control subunit (CV α), it was possible to calculate the percentage of cybrid neuronal cells that had intact assembly of complexes I-V (Figure 3). All five complexes were correctly assembled in CNT cybrid neuronal cells (data not shown). PD cybrid neuron lines (PD60, PD61) exhibited an increase mitochondrial movement after LLLT that did not achieve significance. The same lines also exhibited low levels of complexes I and IV assembly as well as reduced assembly in two other complexes (Figure 3). In contrast, the four PD cybrids (PD63, PD65, PD66, PD67) that exhibited a significant response to LLLT contained more cybrid neuronal cells with correctly assembled complexes I and IV (50–100%). In these four PD lines, at least three complexes were correctly assembled which suggests that the mtETC was more functional (Figure 3).
Figure 3.
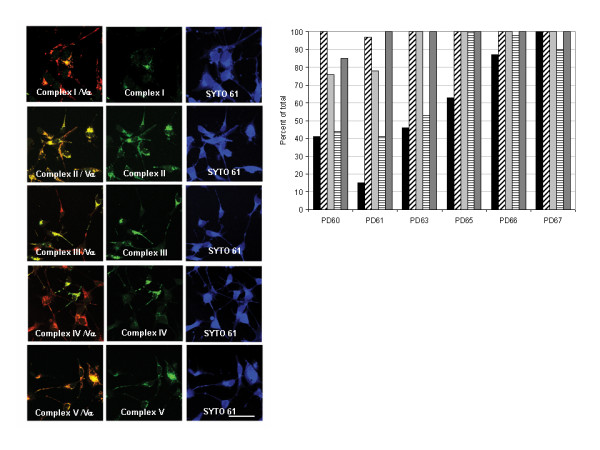
The montage in 3A shows PD61 cybrid neuronal cells stained with the MitoProfile® OXPHOS/PDH immunocytochemistry kit (MitoSciences). Specific antibodies (FITC secondary antibody, green) to Complex I-NDUFB4 (A-C), Complex II-30 kDa (D-F), Complex III-Core 2 protein (G-I), Complex IV-subunit I (J-L), and Complex V-OSCP (M-O) were combined with antibodies to a control subunit (CVα, Texas red secondary, red) to double stain mitochondria in cybrid neuronal cells (see methods). The cell bodies were stained with SYTO 61 (blue). Cybrid neuronal cells in the first column that exhibit double staining are orange or yellow and have intact mETC assembly while cybrid neuronal cells that lack intact assembly of complexs in the mETC are red. Bar = 20 μm. The graph in 3B depicts counts of PD cybrid neuronal cells that contained fully assembled mtETC complexes. In this graph Complex I is black, Complex II has diagonal stripes, Complex III is light gray, Complex IV has horizontal stripes, and Complex V is dark gray. The CNT cybrid neuronal cells exhibited complete assembly of all five complexes (data not shown).
Oxygen utilization studies
An Oxygraph 2K (Oroboros) [48] was used to measure changes in oxygen utilization by Rho0 cells that lack a functional mtETC, two lines of PD cybrid neuronal cells (PD60 and PD61) that did not respond significantly to LLLT, two lines of PD cybrid neuronal cells (PD65 and PD66) that did respond significantly to LLLT and a CNT line (CNT91) of cybrid neuronal cells. These data are shown in Figure 4. Rho0 cells, PD60 and PD61 had reduced routine aerobic respiration (Figure 4A), capacity of the mtETC (Figure 4C) and complex I mediated respiration (Figure 4E) compared to CNT91. [see [48] for details] Non-phosphorylating (oligomycin-inhibited) respiration (LEAK, Figure 4B) did not vary greatly among any of the cell lines tested; however the residual oxygen consumption (ROX, Figure 4D) was higher in Rho0, PD60 and PD61 lines compared to PD65, PD66 or CNT91. In contrast CNT91, PD65 and PD66 had higher routine aerobic respiration, mtETC capacity, and complex I mediated respiration (Figures 4A, 4C and 4E) as well as a reduced level of ROX (Figure 4D). The respiratory control ratio (RCR) which reveals the relative efficiency of ETC coupling (Figure 4F) was diminished in Rho0, PD60 and PD61 compared to cybrid lines that responded significantly to LLLT (PD65, PD66 and CNT91). During the two hour period of measurement after LLLT there was no statistically significant change in respiratory chain activity in any of the cybrid lines tested using the Oxygraph-2k (data not shown).
Figure 4.
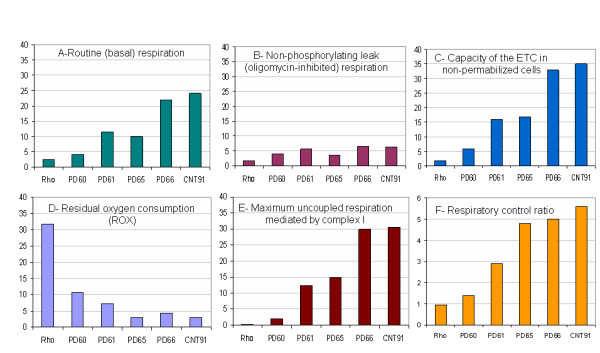
The graphs in this figure illustrate the levels of routine (basal) respiration (A), nonphosphorylating leak (oligomycin inhibited) respiration (B), capacity of the electron transport chain in nonpermeablized cells (C), residual oxygen consumption or ROX (D), maximum uncoupled respiration mediated by complex I (E) and the respiratory control ratio (F) in Rho0 cells, and selected PD and CNT cybrid neuron lines. The Y axis in Figures 4A-E is Respiration [pmol O2/second/10-6 cells]. These data were calculated based on formulas described by Gnaiger. [48]
Discussion
In this study we showed that the velocity of mitochondrial movement by axonal transport was significantly reduced in a human neuronal model of sporadic PD. Other studies have also suggested a role for reduced axonal transport in the pathogenesis of PD. Our data lends additional support for this proposition and for the role that dysfunctional mitochondria play in early PD pathogenesis. We also showed that a single treatment with near-infrared laser light can restore the velocity of mitochondrial movement in PD cybrid neuronal cells to CNT levels. LLLT with near-infrared laser light has been proven safe and effective for the treatment of complex neurological conditions like stroke in humans and rodents. [40,49] Our results provide essential data to support the exploration of LLLT with near-infrared laser light for treatment of neurological conditions like PD.
The mitochondria in PD cybrids exhibit structural and functional changes that could interfere with normal movement of mitochondria by axonal transport or contribute to the differential response of PD cybrid mitochondria to LLLT. These include-1) reduced activity of complexes I and IV, 2) increased levels of oxidized and nitrated proteins, 3) insufficient ATP, 4) a swollen and abnormal morphology, 5) increased levels of modified α-synuclein, and 6) increased free/polymerized tubulin ratio. [24,28,50,51]
A substantial body of evidence supports the proposal that complex I dysfunction plays a critical role in PD pathogenesis. [52] Analysis of the mtETC revealed reduced assembly of complex I in five out of six PD cybrid neuronal cell lines (Figure 3). Previous studies suggest that mtETC subunit expression was also altered in sporadic PD brain. [30] Complex I subunits were also oxidatively damaged and this damage correlated with decreased function in sporadic PD brain. [30] Findings of reduced capacity of the electron transport chain, routine respiration and complex I mediated oxygen utilization (Figure 4) in PD cybrid neuronal cells, complimented our mtETC assembly data. CNT91 cybrid neuronal cells had oxygen utilization rates comparable to fibroblasts, peritoneal mesothelial cells and hematopoetic cells. [48,53,54]
Complex IV is also misassembled in four out of six PD cybrid lines (Figure 3). Reduced complex IV activity and increased levels of nitrated proteins have been detected in PD tissue and in PD cybrid lines[24,27]. Studies have suggested that nitrative damage plays an important role in PD. [55] Both complexes I and IV are targets of NO. [43] Treatment of neuronal cultures with NO reduces the movement of mitochondria and synaptic vesicles; therefore, it is reasonable to propose that nitrative damage to the mtETC contributes to the loss of mitochondrial movement in PD cybrids. [56-58]
While the degree of mtETC misassembly in each PD cybrid line did not correlate with mitochondrial velocity, mtETC misassembly did relate to the response of PD cybrid neuronal cells to LLLT. All the PD cybrid neuronal cells showed increased mitochondrial velocity after LLLT; however the changes in PD60 and PD61 did not consistently achieve significance (Figure 3). Current understanding suggests that near-infrared light is absorbed by photosensitizers such as the oxidized copper centers in complex IV. This results in the excitation and transfer of electrons to oxygen and the generation of low level changes in ROS expression that activates cellular processes through the release of transcription factors and altered gene expression. [41,59,60] Near-infrared light can also reverse the inhibition of complex IV and cellular respiration caused by NO occupation of the oxygen site. [43] Cybrid neuronal cells PD60 and PD61 had the lowest levels of complex I and IV assembly of all our lines (Figure 3). It's not surprising therefore that the PD cybrid lines with the least intact complex I and IV assembly also have a limited response to near-infrared light. LLLT was able to significantly increase mitochondrial movement in other less crippled PD cybrid neuronal cells (Figure 3). LLLT did not improve oxygen utilization in OxyGraph studies of selected PD or CNT lines of cybrid neurons (data not shown). The two hour period selected for this study may not have been sufficient time for changes in mtETC activity to achieve detectable levels. Further studies will focus on longer post-LLLT intervals.
The velocity of mitochondrial movement in CNT cybrid neuronal cells was not altered by LLLT. LLLT also failed to enhance respiratory function in control animals used for a study of rotenone induced mitochondrial optic neuropathy. [35] Rojas et al[35] suggest that mitochondria that are functioning at a maximal rate are unable to be further stimulated by LLLT. On the other hand, if the mtETC is partially inhibited or dysfunctional, LLLT can increase electron flow by stimulating complex IV. [35] Therefore, we were not surprised when the velocity of mitochondrial movement in CNT cybrid neuronal cells did not respond significantly to LLLT in our study.
Reduced ATP levels stemming from mtETC dysfunction could also limit availability of ATP for use by the motors that drive axonal transport. [61] Esteves et al[24] detected reduced levels of ATP in their undifferentiated NT2 PD cybrids. Recent studies have suggested regional increases in ADP can trigger mitochondrial movement. [62] We used the BioVision Research Products ApoSensor ADP/ATP ratio assay kit (Mountain View, CA) to measure the ADP/ATP ratio but could not detect consistently altered levels of ATP or ADP levels (data not shown) in our PD and CNT cybrid neuronal cells after LLLT. Further studies are needed to verify the role reduced ATP or increased ADP play in the reduced movement of mitochondria in PD cybrid neuronal cells.
The mitochondria in valinomycin-treated cerebellar neuronal cells were swollen and they blocked mitochondrial movement. [63] The mitochondria in PD cybrid neurites were not swollen or rounded (Figure 1) so the reduction in movement can not be attributed to this cause. PD cybrids did contain aggregated, oxidized [27] and phosphorylated α-synuclein (data not shown). In transfected neuronal cells, Saha et al [64] showed that mutated and phosphorylated α-synuclein reduced axonal transport. Modified α-synuclein may contribute to the reduction in mitochondrial movement in PD cybrid neurites but it is unlikely that LLLT could alter α-synuclein aggregation, oxidation or phosphorylation in the period of time studied. Aggregates of α-synuclein can appear in PD cybrid neurites and in rotenone-treated SH-SY5Y neuronal cells, cause localized swelling that resembles a Lewy neurite and disrupt movement of all mitochondria distal to the swelling ([2] and data not shown). Lewy neurite-like swellings were uncommon in PD cybrid neurites and did not account for the reduced movement of mitochondria in PD cybrid neuronal cells.
Conclusion
Speculation that reduced axonal transport plays a role in PD pathogenesis has existed for some time. Our ability to assess the role of axonal transport in PD has improved recently with the development of appropriate models and methods. The outcome of our studies of human cybrid neuronal cells that contain mitochondria derived from CNT and PD volunteers illustrates that axonal transport of mitochondria is significantly reduced in PD cybrid neurites. Reduced axonal transport could underlie the loss of neurites through terminal degeneration and result in the death of dopaminergic neuronal cells in PD. The observation that a single, low level exposure to LLLT can increase the velocity of mitochondrial movement and restore it to CNT levels in PD cybrid neurites is remarkable but consistent with our current understanding of the mechanism of LLLT. In a recent publication, behavioral, metabolic and anatomic measures were improved in a dose-dependent manner in a rat model of rotenone-induced optic neuropathy. [35] LLLT is also currently used to treat a wide range of human conditions that involve injury and inflammation. Recent findings that LLLT can be used to ameliorate the consequences of stroke in phase 2 clinical trials suggest that LLLT with near-infrared light [40] should be explored for the treatment of other neurological diseases such as PD.
Competing interests
JS and LDT are employees and shareholders in PhotoThera, Inc. UO was an employee of PhotoThera, Inc. during 2006–2007. PAT, KMS and MKB have no competing financial interests.
Authors' contributions
PAT designed experiments and conducted the axonal transport studies and wrote the manuscript; KMS prepared differentiated cybrid neuronal cells and conducted the oxygraph studies; MKB produced the cybrid lines and conducted the immunocytochemical analysis of mtETC; LDT and JS contributed to the optical and thermal design (dose selection), and reviewed the manuscript; UO contributed to the optical, thermal and experimental design and reviewed the manuscript.
Supplementary Material
PD63 cybrid neuron process. Time lapse movie of MtRed-labeled mitochondria in a PD63 cybrid neuron process (pass 25) after differentiation with 6 nM staurosporine for 12 days.
PD63 cybrid neuron process 85 minutes after LLLT with 50 mW of 810 nm light for 40 seconds. Time lapse movie of MtRed-labeled mitochondria in a PD63 cybrid neuron process (pass 25) after differentiation with 6 nM staurosporine for 12 days. This movie was recorded 85 minutes after LLLT with 50 mW of 810 nm light for 40 seconds.
CNT68 cybrid neuron process. Time lapse movie of MtRed-labeled mitochondria in a CNT68 cybrid neuron process (pass 24) after differentiation with 6 nM staurosporine for 12 days.
CNT68 cybrid neuron process 90 minutes after LLLT with 50 mW of 810 nm light for 40 seconds. Time lapse movie of MtRed-labeled mitochondria in a CNT68 cybrid neuron process (pass 24) after differentiation with 6 nM staurosporine for 12 days. This movie was recorded 90 minutes after LLLT with 50 mW of 810 nm light for 40 seconds.
Acknowledgments
Acknowledgements
The authors would like to thank Dr. Nina Solenski, Dr. Bradley Miller and members of the Morris K. Udall Research Center of Excellence at the University of Virginia such as Dr. James Bennett Jr. for insightful comments. Michelle Warthan and Marilena Lekoudis also contributed to this study during their time in the lab.
Funding for this research was provided by PhotoThera, Inc. and a grant from the National Institute of Neurological Disorders and Stroke (NS039788).
Contributor Information
Patricia A Trimmer, Email: pat5q@virginia.edu.
Kathleen M Schwartz, Email: kms7t@virginia.edu.
M Kathleen Borland, Email: mkb2n@virginia.edu.
Luis De Taboada, Email: ldetaboada@photothera.com.
Jackson Streeter, Email: jstreeter@photothera.com.
Uri Oron, Email: UriO@tauex.tau.ac.il.
References
- Foltynie T, Brayne C, Barker RA. The heterogeneity of idiopathic Parkinson's disease. J Neurol. 2002;249:138–145. doi: 10.1007/PL00007856. [DOI] [PubMed] [Google Scholar]
- Borland MK, Trimmer PA, Rubinstein JD, Keeney PM, Mohanakumar K, Liu L, Bennett JP., Jr Chronic, low-dose rotenone reproduces Lewy neurites found in early stages of Parkinson's disease, reduces mitochondrial movement and slowly kills differentiated SH-SY5Y neural cells. Mol Neurodegener. 2008;3:21. doi: 10.1186/1750-1326-3-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrish PK, Sawle GV, Brooks DJ. An [18F]dopa-PET and clinical study of the rate of progression in Parkinson's disease. Brain. 1996;119:585–591. doi: 10.1093/brain/119.2.585. [DOI] [PubMed] [Google Scholar]
- Lang AE, Obeso JA. Challenges in Parkinson's disease: restoration of the nigrostriatal dopamine system is not enough. Lancet Neurol. 2004;3:309–316. doi: 10.1016/S1474-4422(04)00740-9. [DOI] [PubMed] [Google Scholar]
- Sulzer D. Multiple hit hypotheses for dopamine neuron loss in Parkinson's disease. Trends Neurosci. 2007;30:244–250. doi: 10.1016/j.tins.2007.03.009. [DOI] [PubMed] [Google Scholar]
- Mattson MP, Liu D. Mitochondrial potassium channels and uncoupling proteins in synaptic plasticity and neuronal cell death. Biochem Biophys Res Commun . 2003;304:539–549. doi: 10.1016/S0006-291X(03)00627-2. [DOI] [PubMed] [Google Scholar]
- De Vos KJ, Grierson AJ, Ackerley S, Miller CC. Role of axonal transport in neurodegenerative diseases. Annu Rev Neurosci. 2008;31:151–173. doi: 10.1146/annurev.neuro.31.061307.090711. [DOI] [PubMed] [Google Scholar]
- Bueler H. Impaired mitochondrial dynamics and function in the pathogenesis of Parkinson's disease. Exp Neurol. 2009 doi: 10.1016/j.expneurol.2009.03.006. [DOI] [PubMed] [Google Scholar]
- Lach B, Grimes D, Benoit B, Minkiewicz-Janda A. Caudate nucleus pathology in Parkinson's disease: ultrastructural and biochemical findings in biopsy material. Acta Neuropathol(Berl) 1992;83:352–360. doi: 10.1007/BF00713525. [DOI] [PubMed] [Google Scholar]
- Cappelletti G, Surrey T, Maci R. The parkinsonism producing neurotoxin MPP+ affects microtubule dynamics by acting as a destabilising factor. FEBS Lett. 2005;579:4781–4786. doi: 10.1016/j.febslet.2005.07.058. [DOI] [PubMed] [Google Scholar]
- Morfini G, Pigino G, Opalach K, Serulle Y, Moreira JE, Sugimori M, Llinas RR, Brady ST. 1-Methyl-4-phenylpyridinium affects fast axonal transport by activation of caspase and protein kinase C. Proc Natl Acad Sci U S A. 2007;104:2437–2441. doi: 10.1073/pnas.0611231104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang Q, Yan Z, Feng J. Neurotrophic factors stabilize microtubules and protect against rotenone toxicity on dopaminergic neurons. J Biol Chem. 2006;281:29391–29400. doi: 10.1074/jbc.M602740200. [DOI] [PubMed] [Google Scholar]
- Liang Y, Li S, Wen C, Zhang Y, Guo Q, Wang H, Su B. Intrastriatal injection of colchicine induces striatonigral degeneration in mice. J Neurochem. 2008;106:1815–1827. doi: 10.1111/j.1471-4159.2008.05526.x. [DOI] [PubMed] [Google Scholar]
- Schapira AH. Mitochondria in the aetiology and pathogenesis of Parkinson's disease. Lancet Neurol. 2008;7:97–109. doi: 10.1016/S1474-4422(07)70327-7. [DOI] [PubMed] [Google Scholar]
- Thomas B, Beal MF. Parkinson's disease. Hum Mol Genet. 2007;16:R183–194. doi: 10.1093/hmg/ddm159. [DOI] [PubMed] [Google Scholar]
- Vila M, Ramonet D, Perier C. Mitochondrial alterations in Parkinson's disease: new clues. J Neurochem. 2008;107:317–328. doi: 10.1111/j.1471-4159.2008.05604.x. [DOI] [PubMed] [Google Scholar]
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH, Jaros E, Hersheson JS, Betts J, Klopstock T, et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat Genet. 2006;38:515–517. doi: 10.1038/ng1769. [DOI] [PubMed] [Google Scholar]
- Parker WD, Jr, Parks JK. Mitochondrial ND5 mutations in idiopathic Parkinson's disease. Biochem Biophys Res Commun. 2005;326:667–669. doi: 10.1016/j.bbrc.2004.11.093. [DOI] [PubMed] [Google Scholar]
- Kraytsberg Y, Kudryavtseva E, McKee AC, Geula C, Kowall NW, Khrapko K. Mitochondrial DNA deletions are abundant and cause functional impairment in aged human substantia nigra neurons. Nat Genet. 2006;38:518–520. doi: 10.1038/ng1778. [DOI] [PubMed] [Google Scholar]
- Van Laar VS, Berman SB. Mitochondrial dynamics in Parkinson's disease. Exp Neurol. 2009 doi: 10.1016/j.expneurol.2009.03.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung CY, Koprich JB, Siddiqi H, Isacson O. Dynamic changes in presynaptic and axonal transport proteins combined with striatal neuroinflammation precede dopaminergic neuronal loss in a rat model of AAV alpha-synucleinopathy. J Neurosci. 2009;29:3365–3373. doi: 10.1523/JNEUROSCI.5427-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng J. Microtubule: a common target for parkin and Parkinson's disease toxins. Neuroscientist. 2006;12:469–476. doi: 10.1177/1073858406293853. [DOI] [PubMed] [Google Scholar]
- Miller SW, Trimmer PA, Parker WD, Jr, Davis RE. Creation and characterization of mitochondrial DNA-depleted cell lines with "neuronal-like" properties. J Neurochem . 1996;67:1897–1907. doi: 10.1046/j.1471-4159.1996.67051897.x. [DOI] [PubMed] [Google Scholar]
- Esteves AR, Domingues AF, Ferreira IL, Januario C, Swerdlow RH, Oliveira CR, Cardoso SM. Mitochondrial function in Parkinson's disease cybrids containing an nt2 neuron-like nuclear background. Mitochondrion. 2008;8:219–228. doi: 10.1016/j.mito.2008.03.004. [DOI] [PubMed] [Google Scholar]
- Barrientos A, Kenyon L, Moraes CT. Human xenomitochondrial cybrids. Cellular models of mitochondrial complex I deficiency. J Biol Chem. 1998;273:14210–14217. doi: 10.1074/jbc.273.23.14210. [DOI] [PubMed] [Google Scholar]
- Esteves AR, Arduino DM, Swerdlow RH, Oliveira CR, Cardoso SM. Oxidative Stress involvement in alpha-synuclein oligomerization in Parkinsons disease cybrids. Antioxid Redox Signal. 2008 doi: 10.1089/ars.2008.2247. [DOI] [PubMed] [Google Scholar]
- Trimmer PA, Borland MK, Keeney PM, Bennett JP, Jr, Parker WD., Jr Parkinson's disease transgenic mitochondrial cybrids generate Lewy inclusion bodies. J Neurochem . 2004;88:800–812. doi: 10.1046/j.1471-4159.2003.02168.x. [DOI] [PubMed] [Google Scholar]
- Swerdlow RH, Parks JK, Miller SW, Tuttle JB, Trimmer PA, Sheehan JP, Bennett JP, Jr, Davis RE, Parker WD., Jr Origin and functional consequences of the complex I defect in Parkinson's disease. Ann Neurol . 1996;40:663–671. doi: 10.1002/ana.410400417. [DOI] [PubMed] [Google Scholar]
- Trimmer PA, Bennett JP., Jr The cybrid model of sporadic Parkinson's disease. Exp Neurol. 2009 doi: 10.1016/j.expneurol.2009.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keeney PM, Xie J, Capaldi RA, Bennett JP., Jr Parkinson's disease brain mitochondrial complex I has oxidatively damaged subunits and is functionally impaired and misassembled. J Neurosci. 2006;26:5256–5264. doi: 10.1523/JNEUROSCI.0984-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cassarino DS, Bennett JP., Jr An evaluation of the role of mitochondria in neurodegenerative diseases: mitochondrial mutations and oxidative pathology, protective nuclear responses, and cell death in neurodegeneration. Brain ResBrain ResRev. 1999;29:1–25. doi: 10.1016/S0165-0173(98)00046-0. [DOI] [PubMed] [Google Scholar]
- Trimmer PA, Swerdlow RH, Parks JK, Keeney P, Bennett JP, Jr, Miller SW, Davis RE, Parker WD., Jr Abnormal mitochondrial morphology in sporadic Parkinson's and Alzheimer's disease cybrid cell lines. Exp Neurol. 2000;162:37–50. doi: 10.1006/exnr.2000.7333. [DOI] [PubMed] [Google Scholar]
- Borland MK, Mohanakumar KP, Rubinstein JD, Keeney PM, Xie J, Capaldi R, Dunham LD, Trimmer PA, Bennett JP., Jr Relationships among molecular genetic and respiratory properties of Parkinson's disease cybrid cells show similarities to Parkinson's brain tissues. Biochim Biophys Acta. 2008;1792:68–74. doi: 10.1016/j.bbadis.2008.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eells JT, Wong-Riley MT, VerHoeve J, Henry M, Buchman EV, Kane MP, Gould LJ, Das R, Jett M, Hodgson BD, et al. Mitochondrial signal transduction in accelerated wound and retinal healing by near-infrared light therapy. Mitochondrion. 2004;4:559–567. doi: 10.1016/j.mito.2004.07.033. [DOI] [PubMed] [Google Scholar]
- Rojas JC, Lee J, John JM, Gonzalez-Lima F. Neuroprotective Effects of Near-Infrared Light in an In Vivo Model of Mitochondrial Optic Neuropathy. J Neurosci. 2008;28:13511–13521. doi: 10.1523/JNEUROSCI.3457-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oron A, Oron U, Streeter J, De TL, Alexandrovich A, Trembovler V, Shohami E. low-level laser therapy applied transcranially to mice following traumatic brain injury significantly reduces long-term neurological deficits. JNeurotrauma. 2007;24:651–656. doi: 10.1089/neu.2006.0198. [DOI] [PubMed] [Google Scholar]
- Byrnes KR, Waynant RW, Ilev IK, Wu X, Barna L, Smith K, Heckert R, Gerst H, Anders JJ. Light promotes regeneration and functional recovery and alters the immune response after spinal cord injury. Lasers SurgMed. 2005;36:171–185. doi: 10.1002/lsm.20143. [DOI] [PubMed] [Google Scholar]
- Chow RT, David MA, Armati PJ. 830 nm laser irradiation induces varicosity formation, reduces mitochondrial membrane potential and blocks fast axonal flow in small and medium diameter rat dorsal root ganglion neurons: implications for the analgesic effects of 830 nm laser. J Peripher Nerv Syst. 2007;12:28–39. doi: 10.1111/j.1529-8027.2007.00114.x. [DOI] [PubMed] [Google Scholar]
- Liang HL, Whelan HT, Eells JT, Wong-Riley MT. Near-infrared light via light-emitting diode treatment is therapeutic against rotenone- and 1-methyl-4-phenylpyridinium ion-induced neurotoxicity. Neuroscience. 2008;153:963–974. doi: 10.1016/j.neuroscience.2008.03.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lampl Y, Zivin JA, Fisher M, Lew R, Welin L, Dahlof B, Borenstein P, Andersson B, Perez J, Caparo C, et al. Infrared laser therapy for ischemic stroke: a new treatment strategy: results of the NeuroThera Effectiveness and Safety Trial-1 (NEST-1) Stroke. 2007;38:1843–1849. doi: 10.1161/STROKEAHA.106.478230. [DOI] [PubMed] [Google Scholar]
- Lane N. Cell biology: power games. Nature. 2006;443:901–903. doi: 10.1038/443901a. [DOI] [PubMed] [Google Scholar]
- Karu TI, Pyatibrat LV, Afanasyeva NI. A novel mitochondrial signaling pathway activated by visible-to-near infrared radiation. PhotochemPhotobiol. 2004;80:366–372. doi: 10.1562/2004-03-25-RA-123.1. [DOI] [PubMed] [Google Scholar]
- Borutaite V, Budriunaite A, Brown GC. Reversal of nitric oxide-, peroxynitrite- and S-nitrosothiol-induced inhibition of mitochondrial respiration or complex I activity by light and thiols. Biochim Biophys Acta. 2000;1459:405–412. doi: 10.1016/S0005-2728(00)00178-X. [DOI] [PubMed] [Google Scholar]
- Hoehn MM, Yahr MD. Parkinsonism: onset, progression and mortality. Neurology. 1967;17:427–442. doi: 10.1212/wnl.17.5.427. [DOI] [PubMed] [Google Scholar]
- Swerdlow RH. Mitochondria in cybrids containing mtDNA from persons with mitochondriopathies. J Neurosci Res. 2007;85:3416–3428. doi: 10.1002/jnr.21167. [DOI] [PubMed] [Google Scholar]
- Trimmer PA, Borland MK. Differentiated Alzheimer's disease transmitochondrial cybrid cell lines exhibit reduced organelle movement. Antioxid Redox Signal. 2005;7:1101–1109. doi: 10.1089/ars.2005.7.1101. [DOI] [PubMed] [Google Scholar]
- Capaldi RA, Murray J, Byrne L, Janes MS, Marusich MF. Immunological approaches to the characterization and diagnosis of mitochondrial disease. Mitochondrion. 2004;4:417–426. doi: 10.1016/j.mito.2004.07.006. [DOI] [PubMed] [Google Scholar]
- Gnaiger E. Drug-Induced Mitochondrial Dysfunction. Will JADaY: John Wiley & Sons, Inc; 2008. Polarographic oxygen sensor, the oxygraph, and high-resolution respirometry to assess mitochondrial function; pp. 327–352. [Google Scholar]
- Ilic S, Leichliter S, Streeter J, Oron A, Detaboada L, Oron U. Effects of power densities, continuous and pulse frequencies, and number of sessions of low-level laser therapy on intact rat brain. PhotomedLaser Surg. 2006;24:458–466. doi: 10.1089/pho.2006.24.458. [DOI] [PubMed] [Google Scholar]
- Cassarino DS, Fall CP, Swerdlow RH, Smith TS, Halvorsen EM, Miller SW, Parks JP, Parker WD, Jr, Bennett JP., Jr Elevated reactive oxygen species and antioxidant enzyme activities in animal and cellular models of Parkinson's disease. Biochim Biophys Acta. 1997;1362:77–86. doi: 10.1016/s0925-4439(97)00070-7. [DOI] [PubMed] [Google Scholar]
- Sheehan JP, Swerdlow RH, Parker WD, Miller SW, Davis RE, Tuttle JB. Altered calcium homeostasis in cells transformed by mitochondria from individuals with Parkinson's disease. J Neurochem . 1997;68:1221–1233. doi: 10.1046/j.1471-4159.1997.68031221.x. [DOI] [PubMed] [Google Scholar]
- Pathak RU, Davey GP. Complex I and energy thresholds in the brain. Biochim Biophys Acta. 2008;1777:777–782. doi: 10.1016/j.bbabio.2008.05.443. [DOI] [PubMed] [Google Scholar]
- Hutter E, Renner K, Pfister G, Stockl P, Jansen-Durr P, Gnaiger E. Senescence-associated changes in respiration and oxidative phosphorylation in primary human fibroblasts. Biochem J. 2004;380:919–928. doi: 10.1042/BJ20040095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stadlmann S, Renner K, Pollheimer J, Moser PL, Zeimet AG, Offner FA, Gnaiger E. Preserved coupling of oxidative phosphorylation but decreased mitochondrial respiratory capacity in IL-1beta-treated human peritoneal mesothelial cells. Cell Biochem Biophys. 2006;44:179–186. doi: 10.1385/CBB:44:2:179. [DOI] [PubMed] [Google Scholar]
- Guix FX, Uribesalgo I, Coma M, Muñoz FJ. The physiology and pathophysiology of nitric oxide in the brain. Progress in Neurobiology. 2005;76:126–152. doi: 10.1016/j.pneurobio.2005.06.001. [DOI] [PubMed] [Google Scholar]
- Rintoul GL, Bennett VJ, Papaconstandinou NA, Reynolds IJ. Nitric oxide inhibits mitochondrial movement in forebrain neurons associated with disruption of mitochondrial membrane potential. JNeurochem. 2006;97:800–806. doi: 10.1111/j.1471-4159.2006.03788.x. [DOI] [PubMed] [Google Scholar]
- Zanelli SA, Trimmer PA, Solenski NJ. Nitric oxide impairs mitochondrial movement in cortical neurons during hypoxia. JNeurochem. 2006;97:724–736. doi: 10.1111/j.1471-4159.2006.03767.x. [DOI] [PubMed] [Google Scholar]
- Stagi M, Dittrich PS, Frank N, Iliev AI, Schwille P, Neumann H. Breakdown of axonal synaptic vesicle precursor transport by microglial nitric oxide. J Neurosci. 2005;25:352–362. doi: 10.1523/JNEUROSCI.3887-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lubart R, Breitbart H. Biostimulative effects of low-energy lasers and their implications for medicine. Drug Development Research. 2000;50:471–475. doi: 10.1002/1098-2299(200007/08)50:3/4<471::AID-DDR30>3.0.CO;2-E. [DOI] [Google Scholar]
- Karu TI. Mitochondrial Signaling in Mammalian Cells Activated by Red and Near-IR Radiation. Photochem Photobiol. 2008;84:1091–1099. doi: 10.1111/j.1751-1097.2008.00394.x. [DOI] [PubMed] [Google Scholar]
- Hollenbeck PJ, Saxton WM. The axonal transport of mitochondria. JCell Sci. 2005;118:5411–5419. doi: 10.1242/jcs.02745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mironov SL. ADP regulates movements of mitochondria in neurons. Biophys J. 2007;92:2944–2952. doi: 10.1529/biophysj.106.092981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaasik A, Safiulina D, Choubey V, Kuum M, Zharkovsky A, Veksler V. Mitochondrial swelling impairs the transport of organelles in cerebellar granule neurons. J Biol Chem. 2007;282:32821–32826. doi: 10.1074/jbc.M702295200. [DOI] [PubMed] [Google Scholar]
- Saha AR, Hill J, Utton MA, Asuni AA, Ackerley S, Grierson AJ, Miller CC, Davies AM, Buchman VL, Anderton BH, Hanger DP. Parkinson's disease alpha-synuclein mutations exhibit defective axonal transport in cultured neurons. JCell Sci. 2004;117:1017–1024. doi: 10.1242/jcs.00967. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
PD63 cybrid neuron process. Time lapse movie of MtRed-labeled mitochondria in a PD63 cybrid neuron process (pass 25) after differentiation with 6 nM staurosporine for 12 days.
PD63 cybrid neuron process 85 minutes after LLLT with 50 mW of 810 nm light for 40 seconds. Time lapse movie of MtRed-labeled mitochondria in a PD63 cybrid neuron process (pass 25) after differentiation with 6 nM staurosporine for 12 days. This movie was recorded 85 minutes after LLLT with 50 mW of 810 nm light for 40 seconds.
CNT68 cybrid neuron process. Time lapse movie of MtRed-labeled mitochondria in a CNT68 cybrid neuron process (pass 24) after differentiation with 6 nM staurosporine for 12 days.
CNT68 cybrid neuron process 90 minutes after LLLT with 50 mW of 810 nm light for 40 seconds. Time lapse movie of MtRed-labeled mitochondria in a CNT68 cybrid neuron process (pass 24) after differentiation with 6 nM staurosporine for 12 days. This movie was recorded 90 minutes after LLLT with 50 mW of 810 nm light for 40 seconds.