

Abstract
Invasive procedures for prenatal diagnosis are associated with increased risk of abortion; thus, development of noninvasive procedures would be beneficial. Based on the observation that embryonic nucleated red blood cell (NRBC) crosses the placenta and enters the circulation of pregnant women, the ability to identify such cell would allow development of such procedures. Identification of NRBCs in blood samples would be possible provided that specific antibodies are available. Here we have isolated recombinant antibodies using phage display. From the panel of antibody fragments specifically recognising ε-Hb, one was chosen for further characterization, DAb1. DAb1 binds to ε-Hb both in Western blots and immunocytochemistry. Several ε-Hb positive cells were detected in a blood sample taken as postchorionic villus sampling (CVS). To evaluate the sensitivity of the method, K562 cells (which express ε-Hb) were spiked in a blood sample followed by staining in solution and FACS analysis.
1. Introduction
Today, prenatal diagnosis of chromosomal anomalies and genetic disorders is routinely performed by invasive procedures (e.g., chorionic villus sampling (CVS) and amniocentesis). These techniques are highly accurate, but sustain risk of miscarriage, ∼0,5% [1]. Isolation and analysis of intact foetal nucleated cells in maternal blood during pregnancies has been suggested as a method of noninvasive prenatal diagnosis. In 1969, foetal cells were first detected circulating the maternal bloodstream [2]. Embryonic erythroblasts (immature red blood cells), embryonic lymphocytes, and trophoblast cells have all been pursued as sources of foetal genomic material. Embryonic erythroblasts have since drawn most attention [3] due to a short and definite life span (15 minutes) resulting in a reduced risk of misdiagnosis from previous pregnancies [4].
The Hb expression profile of embryonic NRBCs differs from that of adult blood cells. The expression of Hb in the embryo and foetus can be divided in three overlapping periods, corresponding to the major haematopoietic organ: mesoblastic (yolk sac), hepatic (liver and spleen), and myeloid (bone marrow) [5, 6]. Human embryonic ζ- and ε-Hb are synthesised in yolk sac-derived primitive erythroid cells [7] and are followed by the hepatic period extending from the 10th to the 24th gestational week. In this period foetal Hbs are expressed and are initiated with expression of β-Hb then γ-Hb and later the adult Hb (α2β2) [8]. Three genes encode the α-like globins (5′-ζ-α2-α1-3′) and five genes the β-like globins (5′-ε-Gγ-Aγ-δ-β-3′), which are expressed in a developmental sequence that parallels their structural arrangement [8–10], giving rise to 141 and 146 amino acid residues, respectively.
The most specific marker for foetal erythroblasts is the ε-Hb chains as expression of the other foetal Hbs has been demonstrated in adult tissue [11–15]. A monoclonal antibody has previously been described, mouse antihaemoglobin (ε-Chain). Applied Imaging, however, a panel of antibodies recognising the same antigen, is advantageous in order to increase specificity and decrease the risk of identification of false positive. Recombinant antibodies offer additional advantages over traditional monoclonal antibodies with respect to generating vast quantities and genetically linking of active moieties.
Since the initial description of systems allowing display of fusion proteins on the surface of filamentous bacteriophage by Smith in 1985 [16], generation of scFv antibodies from phage antibody libraries have proven to be an efficient way to produce specific antibodies. The major advantage of phage display is the linkage between phenotype and genotype [16]. This enables selection of phage displaying antibodies against any given antigen and at the same time isolation of the corresponding gene encoding the antibody.
Here we have used phage display as a tool to select antibodies against the ε-Hb chain, for the purpose of applying these in noninvasive prenatal diagnosis.
2. Materials and Methods
2.1. Production of Recombinant Epsilon Haemoglobin
HBE1 (Haemoglobin, Epsilon 1, RZPD clone ID: IRAKp961B1722Q2) was obtained from RZPD (Deutsches Ressourcenzentrum für Genomforschung GmbH, Germany).
The coding sequence of the HBE1 was amplified by PCR with primers introducing NdeI and XhoI cloning sites and cloned into the expression vector pET-21c. pET-21c-HBE1 was transfected into BL21(DE3)pLysS by electroporation and used for expression. Bacterial expression cultures were grown with 10 μM haemin (Hemin Bovine, Sigma-Aldrich Denmark A/S, Denmark) and induced with 1 mM IPTG (isopropyl β-D-thio-galactopyranoside). Purification was performed according to the manufactures instructions (Batch purification of 6xHis-tagged proteins from E. coli under denaturing conditions; Qiagen, The QIAexpressionist, protocol 17). The amount and purity of purified ε-Hb were checked by Bradford analysis and SDS-PAGE.
2.2. Cell Culture
The human erythroleukemia cell line K562 was grown as described elsewhere [17]. Haemin (Hemin Bovine, Sigma-Aldrich Denmark A/S, Denmark) was supplemented to culture medium to induce Hb expression [18].
2.3. Blood Samples
Foetal and maternal blood samples were obtained from pregnant women after informed consent. The samples from pregnant women were all taken in the first trimester. Embryonic nucleated red blood cells were enriched by MACS (magnetic activated cell sorting) with CD71-Dynabeads (Dynabeads M-450 CD71, Dynal ASA, Oslo, Norway).
2.4. Single Chain Antibody Selection
An immunotube (Nunc) was coated overnight at 4°C with recombinant ε-Hb in coating buffer (50 mM NaHCO3, pH 9.6). Due to precipitation of the ε-Hb, the proteins were coated by diluting the urea denatured protein 1 : 7 in coating buffer, thus keeping the Urea concentration 1 M. Residual binding was blocked by incubating with 2% MPBS (2% weight/volume low fat milk powder in Phosphate Buffered Saline (PBS)), before 2.5 × 1012 phage particles representing the Griffin 1 scFv library [19] in 2% MPBS were added and incubated for 2 hours. Nonbound phage antibodies were eliminated by washing 10 times with PBS/0,05% tween 20 and 10 times with PBS. Bound phage antibodies were eluted in 0.5 mL trypsin (1 mg/mL) in 50 mM Tris/HCl pH 7, 4, 1 mM CaCl2 and used to infect E. coli, TG-1. Single phage antibody clones were picked and rescued for ELISA using the KM13 helper phage [20]; remaining colonies were scraped of the plate and propagated for phage production. The resulting phage antibodies were applied in a second round of selection. Single phage colonies were picked and grown in 96 well plates for stock and phage production.
2.5. ELISA
Single phage antibody clones were screened for binding to antigen in ELISA as previously described [21], except that phages were rescued with KM13 helper phage [22]. ELISA was carried out by coating extracts of K562 cell, recombinant ε-Hb, BL21 cell extract, or adult blood on ELISA plates (MAXI-sorp, NUNC, Denmark), followed by blocking with 2% MPBS. Fifty μL of precleared culture supernatant from phage antibody clones grown and rescued monoclonally were added together with 50 μL of 4% MPBS and incubated for 2 hours. The ELISA plates were washed and phages were detected with a 1 : 5000 dilution of antiM13-HRP conjugated antibody (Pharmacia). The ELISA was developed with OPD (o-phenylenediamine) tablets (DAKO, Denmark) according to the manufacturer's instructions. In the case of ELISA using pKBJ derivatives of scFv antibodies these were detected with the anti-Myc tag antibody (9E10) and an HRP-conjugated rabbit antimouse antibody (DAKO, Denmark).
2.6. Western- and Dot Blots
Following SDS-PAGE and transfer of proteins to nitrocellulose membranes, membranes were incubated with phage antibodies or purifed scFv derivatives and bound phage or scFv derivatives were detected by incubation with antiM13-HRP conjugated or 9E10 followed by HRP-conjugated rabbit antimouse antibody, respectively. Blots were subsequently developed with ECL plus Western Blotting Detection System according to manufacturers instructions (Amersham Biosciences, UK).
2.7. Expression of Phage Antibodies in pKBJ3 Vector
The antibody genes of the selected clones were recloned into the pKBJ3 vector, as described [23], using the cloning sites Nco1 and Not1. The antibodies were purified by immobilised metal affinity chromatography, Nickel beads (Ni-NTA). The protein concentration was determined with Bradford reagent and the purity by SDS-PAGE. Subsequently, antibodies were further purified on a Q-sepharose column before biotinylation (ECL Protein Biotinylation Module, Amersham Bioscience) or conjugated directly with FITC (Fluorotag FITC Conjugation Kit, Sigma).
2.8. Immunocytochemistry
Either blood or K562 cells were dispersed on to the slides in a thin and uniform layer followed by drying. Cells with or without fixation with 4% paraformaldehyde were permeabilised by submerging into cold (−20°C) methanol overnight at −20°C. Slides were then washed and blocked with 2% MPBS. Subsequently, biotinylated anti-ε-Hb antibody was added in 2% MPBS/0,05% Tween 20 with gentle shaking. Bound antibody was detected with Streptavidin-FITC. Coverglasses were mountered with antifade (DAKO Fluorescent Mounting Medium, DAKO, Denmark) or antifade containing DAPI to counterstain the nuclei, after which the slides were examined in the microscope.
2.9. Fluorescence In Situ Hybridization
X and Y chromosomal analysis was performed to distinguish male embryonic cells from maternal cells. This was done using CEP X (α satellite) SpectrumAqua hybridising centromere of human chromosome X (bands Xp 11.1-q11.1, locus DXZ1; Vysis; Downers Grove, Ill, USA) and CEP Y (α satellite) SpectrumOrange hybridising centromere of human chromosome Y (bands Yq 12, locus DYZ1; Vysis; Downers Grove, Ill, USA). Hybridisations were performed as described elsewhere [24]. Briefly, slides were equilibrated in PBS after immunocytochemistry, refixed in 2% paraformaldehyde for 2 minutes, dehydrated in ethanol, and air-dried. The chromosomespecific repeat probes were used according to the instructions given by the supplier.
2.10. Automated Scanning
Cells stained with biotin-conjugated DAb1 complexed to streptavidin-FITC and DAPI stained nuclei were located by automatic scanning (Applied Imaging) using RCDetect (rare cell detection) scanner developed by MetaSystems (Altlußheim, Germany). The algoritm was modified for embryonic cells in maternal blood. Scanning of slides with blood samples from pre-CVS and post-CVS was performed as described elsewhere [24].
2.11. Fluorescence-Activated Cell Sorting
Cell staining for FACS was performed with FITC conjugated DAb1. FACS analyses were performed using a Becton Dickenson FACS Aria. FITC and propidium iodide were exited using a 488 nm Laser, FITC was collected through a 530/20 nm band pass filter, and propidium iodide was collected through a 595 nm long pass filter. Data is shown without compensation.
K562 cells were collected and washed with 1xPBS, resuspended in ice-cold methanol and incubated for 10 minutes at −20°C. Subsequently cells were centrifuged and washed with 1xPBS before immunostaining with DAb1 (20 μg/mL). Cells were washed 3 times with PBS and mixed with fixed nonstained K562. Immunostaining of K562 spiked into blood was performed according to same protocol.
3. Results
Recombinant ε-Hb was expressed in BL21(DE3)pLysS giving high protein yields (approximately 10 mg ε-Hb/litre culture). Under various culture conditions inclusion bodies were formed, and therefore purifications were performed under denaturing conditions (8 M urea). Subsequent removal of urea led to aggregation, and selections were carried out on denatured ε-Hb. Selections were carried out with Griffin VHVL library [19], yielding approximately 3000 selected clones, of which 384 were tested monoclonally in ELISA against recombinant ε-Hb. This led to the identification of 26 binders (6.7%), 1 strong, and 25 weak binders. A second round of selection was performed to increase the number of positive clones. From the second round 384 clones were tested monoclonal in ELISA, and 46% of these recognised recombinant ε-Hb. Twelve clones specifically recognised both denatured and native ε-Hb and showed no cross-reactivity to paternal Hbs (α- and β-Hb) (Table 1).
Table 1.
Specificity of the 384 selected antibody clones obtainted from the second round of selection against ε-Hb, tested in ELISA towards different antigens. E: recombinant denatured ε-Hb. RBC: parental red blood cell extract (containing α- and β-Hb, but no ε-Hb), BL21: bacterial control cell extract, and K562: K562 cell extract (containing α-, γ-, δ-, ε-, and ζ-Hb). Neg.: amount of clones not recognising any of the tested antigens. E+RBC, E+BL21, E+K562, and E+K562−BL21−RBC, amount of clones recognising ε-Hb (E) together with other antigens. A total of 178 (46%) of the tested clones (384) gave positive ELISA signal toward ε-Hb, while 57 (15%) recognised K562 cell extract. Only two clones recognised K562 cell extract without recognising ε-Hb. Twelve of the selected antibodies showed specificity of the ε-Hb, since they recognised the denatured recombinant ε-Hb, the K562 extract, containing native ε-Hb and not the parental RBC, nor the BL21 extract.
| E | RBC | BL21 | K562 | E+RBC | E+BL21 | E+K562 | E+K562−BL21−RBC | Neg. |
|---|---|---|---|---|---|---|---|---|
| 178 | 10 | 30 | 57 | 10 | 19 | 55 | 12 | 206 |
Sequencing of these 12 potentially specific antibodies revealed four different clones; one was represented 8 times (DAb1), one was represented twice (1.9A), and two were represented once (3.9G and 4.8E). The repeated representation of clones is normal, when performing more than one round of selection. Clone DAb1, carried an Amber stop codon within the reading frame. This was restored using site directed mutagenesis. The four different phage antibodies, were tested in Western blot against a control protein carrying a His-tag, extract from RBC, K562 cell extract, and the recombinant ε-Hb (Figure 1). The four antibodies all recognised purified ε-Hb together with one or more proteins in the K562 extract. Several bands were present in the lane containing the recombinant ε-Hb, representing monomers, dimmers, and higher complexes of the ε-Hb chain. None of the antibodies recognised the His-tag protein; however, clone 3.9G showed a very weak recognition of one protein in the RBC extract.
Figure 1.

Western blots performed with antibodies, DAb1(a), 1.9A(b), 4.8E(c), and 3.9G(d). The four different clones each recognises the recombinant ε-Hb, giving a correct band of 17 kDa (ε-Hb protein fused with His-Tag; adding 8 amino acids). Furthermore they all recognise a slightly smaller band of 146 amino acid residues in the K562 extract, containing the ε-Hb. A difference is observed for the recognised band in the lane containing the recombinant ε-Hb and the ε-Hb in the K562 extract. Lane H: a control his-tagged protein, R: adult red blood cell extract, K: K562 cell extract, and E: recombinant ε-Hb chain (2,5 μg). (e) Representative ponceau staining of the blotted proteins. M: the marker.
All of the four antibody sequences were subcloned to the pKBJ3 vector [23] and two clones of each antibody were tested for expression. Three were chosen for further studies due to their levels of expression in test experiments: 1.9A.1, 3.9G.2, and DAb1. The purified antibodies were biotinylated and their activity was tested in ELISA, all confirming the activity toward the recombinant ε-Hb (data not shown).
To establish the optimal permeabilisation procedure for cell immunostaining, the antibody fragments were tested in Dot-blots where cells treated with different permabilisation regimes were tested (data not shown). The optimal treatments of the cells were with methanol alone or mixed with acetone (1 : 1 or 1 : 3), followed by storage at −20°C over night. Initially immunocytochemistry with antibody DAb1 performed on methanol fixed and permeabilised K562 culture cells showed intense staining, compared to nonstained cells (data not shown). One of the antibodies gave a significant better staining than the other three antibodies, the DAb1. DAb1 stained approximately 10% of the cells in an embryonic blood sample (first trimester) demonstrating the presence of several ε-Hb containing erythroblasts (data not shown). A blood sample taken post-CVS (week 10 + 3) from a woman carrying a male foetus was enriched for foetal cells by CD-71 directed MACS and subsequently analysed as well. MACS-enriched cells obtained from a 2 mL blood sample were coated on slides. The two slides were permeabilised with methanol, and immunostaining was subsequently performed with DAb1. After confirming that the slides contained stained cells, FISH was performed against the X- and Y-chromosomes. Scanning of the slides revealed 129 cells identified as foetal erythroblasts, since they were positive by both Y-chromosomes directed FISH and immunocytochemistry for ε-Hb (Table 2 and Figure 2). This corresponds to one embryonic NRBC pr 7.8 × 107 maternal blood cells. Several cells were damaged during the FISH procedure because methanol is not strong enough as a fixative for the harsh conditions associated with FISH analysis. Subsequent staining, after paraformaldehyde fixation, revealed as strong or stronger staining of cells (not shown), and makes the cell more resistant towards the FISH procedure, which will lead to a higher FISH efficiency.
Table 2.
Summary of FISH analysis on post-CVS blood. Most identified cells contain an XY signal, while those not containing both a Y and an X-signal were damaged by the FISH procedure. A total of 129 embryonic NRBC were identified from the post-CVS sample. FISH efficiencies were 96% for the Y chromosome and 84% for the X-chromosome.
| FISH-signal | Intact | Damaged | Total |
|---|---|---|---|
| XY | 35 | 73 | 108 |
| XX | 0 | 0 | 0 |
| Y | 0 | 16 | 16 |
| X | 0 | 1 | 1 |
| None | 0 | 4 | 4 |
| Total | 35 | 94 | 129 |
Figure 2.
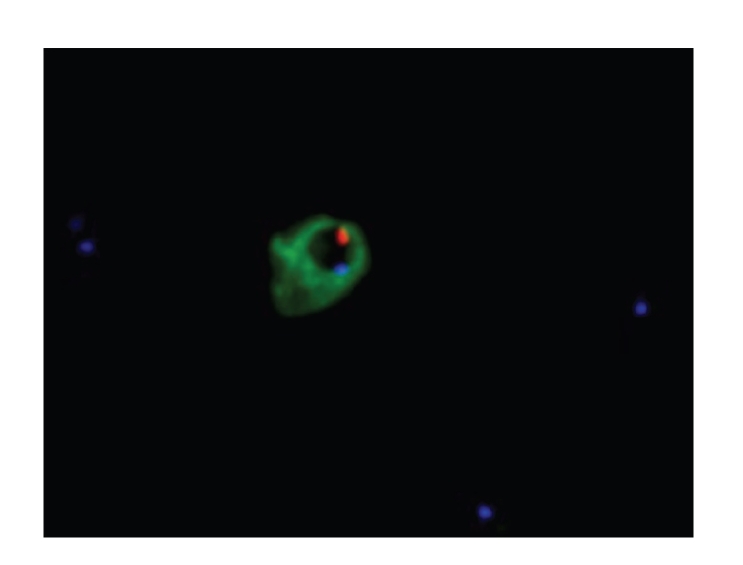
Staining of enriched maternal blood sample taken as post-CVS. First enrichment performed by CD-71 directed MACS. Subsequently cells were mounted on glass slides and permeabilized with methanol followed by immunostaining with FITC conjugated DAb1 (30 μg/mL). Subsequently FISH was performed to karyotype the cells. Embryonic nucleated red blood cells (primitive erythroblasts) identified with DAb1, directly conjugated with FITC (green) is shown. Subsequently, FISH was performed against both the X- (blue) and Y-chromosomes (red), confirming the origin of the immunostained cells. X-chromosomes of nonimmunostained nucleated maternal cells (seen as blue dots), are located around the identified foetal erythroblasts.
Future prenatal diagnosis methods are desired to be applied on maternal blood samples; we therefore obtained blood samples from two pregnant women, pre-CVS. Ten mL of blood, 5 × 1010 blood cells (5 mL each, taken week 8 + 4 and 9 + 0, resp.) was subjected to CD71 directed MACS and stained with DAb1. No ε-Hb positive cells were found when examining ten slides each coated with enriched blood cells from approximately 1 mL blood.
The lack of identified cells could be a result of the enrichment procedure; however, in order to avoid performing such enrichment, methods with a higher throughput such as FACS have to be applied. To explore the feasibility of using DAb1 to stain cells for flow cytometric analysis we mixed K562 cells stained with DAb1 with nonstained K562 cells, followed by FACS analysis. As can be seen in Figure 3(a) two pools appeared. Furthermore FACS was performed on blood sample spiked with K562 (Figure 3(b)). A population of cells is seen in the figure, consisting of K562 cells, confirmed by staining and morphology by microscopy (data not shown), thus providing proof of concept.
Figure 3.

Flow cytometric analysis of (a) stained and nonstained K562. The lower circle shows the non-DAb1 stained cells while upper circle shows the stained cells (P1). All cells have been fixed, permeabilised, and PI stained before cytometry. (b) K562 cells spiked into paternal blood sample. Figure 3 shows 106 cell events. The P1 shows the population of stained K562 among the blood cells.
4. Discussion
Previous attempts have been performed to select specific antibodies against foetal or embryonic erythroblasts, both by others [25, 26] and ourselves (data not shown). In our case we did not succeed, due to a limited availability of foetal cell. Instead we selected on the only known specific antigen of the embryonic erythroblast; ε-Hb and difficulties has likewise been reported with regard to the antibodies selected by Huie et al. as discussed by Bianchi and Hanson [27].
It has previously been shown that the ε-Hb chain is difficult to purify [28]. We overcame this problem by purifying recombinant ε-Hb chain under denaturing conditions in 8 M urea buffers, leading to a yield, high enough for performing antibody selections by phage display. The selections were performed using urea denatured purified antigen directly coated on a solid surface preventing aggregation. Previously reports have described selections made on denatured antigen, but only as a side effect of purification. In some cases the antigen was purified by SDS-PAGE [29, 30]. In other cases antigens were transferred to nitrocellulose membranes after SDS-PAGE [31, 32]. In all cases, antibodies were able to recognise antigen in Western blot.
We succeeded in selecting a specific human antibody against the ε-Hb chain by the use of phage display. Comparison with previous staining obtained with mouse anti-ε-Hb antibody [24] the selected human antibody fragment, DAb1, gives similar staining of NRBC. Previous attempts resulted in isolation of antibody fragments with cross-reactivity to other Hb-chains [33]. Our selected antibody, DAb1 revealed only one band against the K562 extract, in Western blot (Figure 1). This band corresponds to the higher of the two haemoglobin bands, seen on the ponceau staining. This represents a mixture of the β like globin genes, which contain the ε-Hb, while the lower band represents the α like globin genes.
This study illustrates that “you get what you select for.” Here we have shown that antibodies that recognise ε-Hb chain specifically can be selected, and established that (i) it is feasible to select antibodies against urea denatured antigens and (ii) selections can be directed towards recognition of sequence instead of structure. This new selection method can be very useful in future studies, where it is essential to obtain antibodies that recognise the antigen in Western blots. Also, there are many cases in which it is difficult to obtain pure native folded protein, avoiding aggregation, in such cases this method will allow the production of specific antibody fragments.
The usage of automated scanning offers a high-throughput screening method for the identification of foetal cells in a maternal blood sample, either by scanning for immunostained cells or Y-chromosomes containing cells after FISH [34–36]. However, it is a time-consuming and laborious work to validate the identified cells due to a high percentage of false positives. Enrichment procedures reducing the amount of maternal cells are therefore required for usage of automated scanning in noninvasive prenatal diagnostics.
Immunocytochemistry, combined with automated scanning, will enable us to estimate the number of embryonic NRBCs in blood samples taken from pregnant women post- and pre-CVS. Comparing the numbers of NRBCs in the samples taken as post-CVS corresponds well with those reported by others. In the staining on the pre-CVS blood samples no embryonic NRBCs were detected. To date there is no conclusive evidence on the number of embryonic NRBCs in the maternal bloodstream during pregnancy. The reports are many, and quantitations have been performed in different ways. The first reliable report performed on a pre-CVS sample estimated around 1 cell per mL maternal blood [3]. Later reports estimate between 1–5 cells per mL maternal blood [15, 37, 38], depending of the method of enrichment. Others have however found foetal NRBC at much lower frequencies [39–41].
Traditionally CD71 have been used to enrich the fraction of foetal cells. Results obtained by Choolani et al. [39], have shown that embryonic, like foetal erythroblasts were GPA+, CD47+, CD45−, and CD35−, whereas CD71 expression was almost undetectable on embryonic erythroblast but strongly expressed by foetal erythroblasts [39]. Furthermore, all embryonic erythroblasts expressed ε-Hb, whereas less than 0.06% of the foetal erythroblasts expressed this Hb chain. This raises the question whether the CD71 selection actually is appropriate for selection of ε-Hb positive NRBCs.
By FISH we were able to establish that at least 96.1% of the antibody stained NRBCs were of embryonic origin. This efficiency is comparable with those obtained by others [42]. The FISH procedure partly destroyed several of the cells, because fixation of the cells was only performed with methanol, making the cells fragile to the hard treatment of the FISH procedure. Most important is however the fact that no identified antibody stained cells were detected with two X-chromosomes, confirming that ε-Hb exclusively is found in NRBC of embryonic origin. Alternatively, the combination of markers could potentially be used with expression of ε-Hb for FACS isolation of NRBCs. It is likely that the combination of markers will enable the isolation of an essentially pure population of cells.
The paraformaldehyde fixation of the K562 cells resulted in a background of autofluorescence, as seen on Figure 3. A small background is also observed on a variety of blood cell types. Although the autofluorescence on blood cells has been observed to be almost undetectable on the majority of cell types, it is unlikely that FACS can be used as a method for obtaining pure samples of ε-Hb positive cells, but as an enrichment method. To ensure that no cells are lost before the FACS, it is vital to perform FACS on nonenriched blood samples. However, removal of erythrocytes would make the enrichment procedure much faster and increase the purity. If FACS procedure is to be successfully applied, as a high-throughput enrichment method, it will have to be combined with another enrichment procedure that does not lead to a loss of any embryonic erythroblasts.
We have, by the FACS of the K562 immunostained by DAb1 and the spiked samples, obtained a gate for sorting out ε-Hb antibody stained cells. This gate will make it possible to enrich ε-Hb positive embryonic NRBCs from a maternal blood sample. Following FACS enrichment identification of individual embryonic NRBC can be performed by microscopic analysis.
We have shown that DAb1 recognises ε-Hb chain specifically, confirmed by FISH that ε-Hb positive cells are of foetal origin and validated ε-Hb as a cellular marker for embryonic NRBCs. In order to apply the anti-ε-Hb antibody, DAb1, for prenatal diagnosis high-throughput methods like FACS analysis would have to be introduced due to the low frequency of circulating cells.
5. Conclusion
The inherent problem with any noninvasive prenatal diagnosis procedure is the low abundance of foetal cells in the maternal blood, thus requiring sensitive high-throughput methods. Here we have isolated a recombinant antibody, DAb1, with a potential use in high-throughput noninvasive prenatal diagnosis, when combined with new fast instrumentation for FACS. The recombinant antibody can be produced by bacterial expression, thus allowing cost effective large scale production. By applying the recombinant antibody in prenatal diagnosis, it is possible to remove the increased risk of abortion associated with invasive procedures.
Acknowledgments
The authors would like to acknowledge Dr. Greg Winter, MRC Centre, Cambridge for providing the Griffin phage antibody library. Furthermore, support from the Novo Nordisk Foundation and the Danish Research Council is kindly acknowledged.
Abbreviations
- NRBC:
Nucleated red blood cells
- RBC:
Red blood cells
- Hb:
Haemoglobin
- FACS:
Fluorescence-activated cell sorter
- CVS:
Chorionic villus sampling
- MACS:
Magnetic Activated Cell Sorting
- FISH:
Fluorescence in situ hybridization
- ELISA:
Enzyme-linked immunosorbent assays.
References
- 1.Borgatti M, Bianchi N, Mancini I, Feriotto G, Gambari R. New trends in non-invasive prenatal diagnosis: applications of dielectrophoresis-based Lab-on-a-chip platforms to the identification and manipulation of rare cells (Review) International Journal of Molecular Medicine. 2008;21(1):3–12. [PubMed] [Google Scholar]
- 2.Walknowska J, Conte FA, Grumbach MM. Practical and theoretical implications of fetal-maternal lymphocyte transfer. The Lancet. 1969;1(7606):1119–1122. doi: 10.1016/s0140-6736(69)91642-0. [DOI] [PubMed] [Google Scholar]
- 3.Bianchi DW, Flint AF, Pizzimenti MF, Knoll JHM, Latt SA. Isolation of fetal DNA from nucleated erythrocytes in maternal blood. Proceedings of the National Academy of Sciences of the United States of America. 1990;87(9):3279–3283. doi: 10.1073/pnas.87.9.3279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Slunga-Tallberg A, El-Rifai W, Keinanen M, et al. Maternal origin of nucleated erythrocytes in peripheral venous blood of pregnant women. Human Genetics. 1995;96(1):53–57. doi: 10.1007/BF00214186. [DOI] [PubMed] [Google Scholar]
- 5.Wood WG. Haemoglobin synthesis during human fetal development. British Medical Bulletin. 1976;32(3):282–287. doi: 10.1093/oxfordjournals.bmb.a071376. [DOI] [PubMed] [Google Scholar]
- 6.Zanjani ED, Ascensao JL, Tavassoli M. Liver-derived fetal hematopoietic stem cells selectively and preferentially home to the fetal bone marrow. Blood. 1993;81(2):399–404. [PubMed] [Google Scholar]
- 7.Luo HY, Liang XL, Frye C, et al. Embryonic hemoglobins are expressed in definitive cells. Blood. 1999;94(1):359–361. [PubMed] [Google Scholar]
- 8.Palis J, Segel GB. Developmental biology of erythropoiesis. Blood Reviews. 1998;12(2):106–114. doi: 10.1016/s0268-960x(98)90022-4. [DOI] [PubMed] [Google Scholar]
- 9.He Z, Russell JE. Expression, purification, and characterization of human hemoglobins Gower-1 (ζ2ε2), Gower-2 (α2ε2), and Portland-2 (ζ2β2) assembled in complex transgenic-knockout mice. Blood. 2001;97(4):1099–1105. doi: 10.1182/blood.v97.4.1099. [DOI] [PubMed] [Google Scholar]
- 10.Lorkin PA. Fetal and embryonic haemoglobins. Journal of Medical Genetics. 1973;10(1):50–64. doi: 10.1136/jmg.10.1.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Al-Mufti R, Hambley H, Farzaneh F, Nicolaides KH. Fetal and embryonic hemoglobins in erythroblasts from fetal blood and fetal cells enriched from maternal blood in fetal anemia. Haematologica. 2001;86(12):1270–1276. [PubMed] [Google Scholar]
- 12.Chung S-W, Wong SC, Clarke BJ, Patterson M, Walker WH, Chui DH. Human embryonic ζ-globin chains in adult patients with α-thalassemias. Proceedings of the National Academy of Sciences of the United States of America. 1984;81(19):6188–6191. doi: 10.1073/pnas.81.19.6188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Høgh AM, Hviid TVF, Christensen B, et al. ζ-, ε-, and γ-globin mRNA in blood samples and CD71+ cell fractions from fetuses and from pregnant and nonpregnant women, with special attention to identification of fetal erythroblasts. Clinical Chemistry. 2001;47(4):645–653. [PubMed] [Google Scholar]
- 14.Pembrey ME, Weatherall DJ, Clegg JB. Maternal synthesis of haemoglobin F in pregnancy. The Lancet. 1973;1(7816):1350–1354. doi: 10.1016/s0140-6736(73)91675-9. [DOI] [PubMed] [Google Scholar]
- 15.Mavrou A, Kolialexi A, Antsaklis A, Korantzis A, Metaxotou C. Identification of fetal nucleated red blood cells in the maternal circulation during pregnancy using anti-hemoglobin-ε antibody. Fetal Diagnosis and Therapy. 2003;18(5):309–313. doi: 10.1159/000071971. [DOI] [PubMed] [Google Scholar]
- 16.Smith GP. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science. 1985;228(4705):1315–1317. doi: 10.1126/science.4001944. [DOI] [PubMed] [Google Scholar]
- 17.Smith RD, Malley JD, Schechter AN. Quantitative analysis of globin gene induction in single human erythroleukemic cells. Nucleic Acids Research. 2000;28(24):4998–5004. doi: 10.1093/nar/28.24.4998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fibach E, Kollia P, Schechter AN, Noguchi CT, Rodgers GP. Hemin-induced acceleration of hemoglobin production in immature cultured erythroid cells: preferential enhancement of fetal hemoglobin. Blood. 1995;85(10):2967–2974. [PubMed] [Google Scholar]
- 19.Griffiths AD, Williams SC, Hartley O, et al. Isolation of high affinity human antibodies directly from large synthetic repertoires. The EMBO Journal. 1994;13(14):3245–3260. doi: 10.1002/j.1460-2075.1994.tb06626.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jensen KB, Jensen ON, Ravn P, Clark BF, Kristensen P. Identification of keratinocyte-specific markers using phage display and mass spectrometry. Molecular & Cellular Proteomics. 2003;2(2):61–69. doi: 10.1074/mcp.M200049-MCP200. [DOI] [PubMed] [Google Scholar]
- 21.Marks JD, Hoogenboom HR, Bonnert TP, McCafferty J, Griffiths AD, Winter G. By-passing immunization: human antibodies from V-gene libraries displayed on phage. Journal of Molecular Biology. 1991;222(3):581–597. doi: 10.1016/0022-2836(91)90498-u. [DOI] [PubMed] [Google Scholar]
- 22.Kristensen P, Winter G. Proteolytic selection for protein folding using filamentous bacteriophages. Folding and Design. 1998;3(5):321–328. doi: 10.1016/S1359-0278(98)00044-3. [DOI] [PubMed] [Google Scholar]
- 23.Jensen KB, Larsen M, Pedersen JS, et al. Functional improvement of antibody fragments using a novel phage coat protein III fusion system. Biochemical and Biophysical Research Communications. 2002;298(4):566–573. doi: 10.1016/s0006-291x(02)02484-1. [DOI] [PubMed] [Google Scholar]
- 24.Christensen B, Philip J, Lykke-Hansen L, Kølvraa S. Sensitivity and specificity of the identification of fetal cells in maternal blood by combined staining with antibodies against beta-, gamma-and epsilon-globin chains. Fetal Diagnosis and Therapy. 2003;18(6):479–484. doi: 10.1159/000073147. [DOI] [PubMed] [Google Scholar]
- 25.Huie MA, Cheung M-C, Muench MO, Becerril B, Kan YW, Marks JD. Antibodies to human fetal erythroid cells from a nonimmune phage antibody library. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(5):2682–2687. doi: 10.1073/pnas.051631798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alvarez FV, Olander J, Crimmins D, et al. Development, characterization, and use of monoclonal antibodies made to antigens expressed on the surface of fetal nucleated red blood cells. Clinical Chemistry. 1999;45(9):1614–1620. [PubMed] [Google Scholar]
- 27.Bianchi D, Hanson J. Sharpening the tools: a summary of a National Institutes of Health workshop on new technologies for detection of fetal cells in maternal blood for early prenatal diagnosis. Journal of Maternal-Fetal and Neonatal Medicine. 2006;19(4):199–207. doi: 10.1080/14767050600676851. [DOI] [PubMed] [Google Scholar]
- 28.Gale RE, Clegg JB, Huehns ER. Human embryonic haemoglobins Gower 1 and Gower 2. Nature. 1979;280(5718):162–164. doi: 10.1038/280162a0. [DOI] [PubMed] [Google Scholar]
- 29.Pini A, Viti F, Santucci A, et al. Design and use of a phage display library: antibodies with subnanomolar affinity against a marker of angiogenesis eluted from a two-dimensional gel. The Journal of Biological Chemistry. 1998;273(34):21769–21776. doi: 10.1074/jbc.273.34.21769. [DOI] [PubMed] [Google Scholar]
- 30.Spillner E, Deckers S, Grunwald T, Bredehorst R. Paratope-based protein identification by antibody and peptide phage display. Analytical Biochemistry. 2003;321(1):96–104. doi: 10.1016/s0003-2697(03)00439-1. [DOI] [PubMed] [Google Scholar]
- 31.Liu B, Marks JD. Applying phage antibodies to proteomics: selecting single chain Fv antibodies to antigens blotted on nitrocellulose. Analytical Biochemistry. 2000;286(1):119–128. doi: 10.1006/abio.2000.4788. [DOI] [PubMed] [Google Scholar]
- 32.Ravn P, Kjær S, Jensen KH, et al. Identification of phage antibodies toward the Werner protein by selection on Western blots. Electrophoresis. 2000;21(3):509–516. doi: 10.1002/(SICI)1522-2683(20000201)21:3<509::AID-ELPS509>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 33.Parsons HL, Earnshaw JC, Wilton J, et al. Directing phage selections towards specific epitopes. Protein Engineering. 1996;9(11):1043–1049. doi: 10.1093/protein/9.11.1043. [DOI] [PubMed] [Google Scholar]
- 34.Seppo A, Frisova V, Ichetovkin I, et al. Detection of circulating fetal cells utilizing automated microscopy: potential for noninvasive prenatal diagnosis of chromosomal aneuploidies. Prenatal Diagnosis. 2008;28(9):815–821. doi: 10.1002/pd.1987. [DOI] [PubMed] [Google Scholar]
- 35.Kilpatrick MW, Tafas T, Evans MI, et al. Automated detection of rare fetal cells in maternal blood: eliminating the false-positive XY signals in XX pregnancies. American Journal of Obstetrics and Gynecology. 2004;190(6):1571–1581. doi: 10.1016/j.ajog.2004.03.055. [DOI] [PubMed] [Google Scholar]
- 36.Johnson KL, Stroh H, Khosrotehrani K, Bianchi DW. Spot counting to locate fetal cells in maternal blood and tissue: a comparison of manual and automated microscopy. Microscopy Research and Technique. 2007;70:585–588. doi: 10.1002/jemt.20439. [DOI] [PubMed] [Google Scholar]
- 37.Collarini EJ, Cain CA, Gammon D, et al. Comparison of methods for erythroblast selection: application to selecting fetal erythroblasts from maternal blood. Cytometry. 2001;45(4):267–276. doi: 10.1002/1097-0320(20011201)45:4<267::aid-cyto10023>3.0.co;2-d. [DOI] [PubMed] [Google Scholar]
- 38.Kitagawa M, Sugiura K, Omi H, et al. New technique using galactose-specific lectin for isolation of fetal cells from maternal blood. Prenatal Diagnosis. 2002;22(1):17–21. doi: 10.1002/pd.222. [DOI] [PubMed] [Google Scholar]
- 39.Choolani M, O'Donoghue K, Talbert D, et al. Characterization of first trimester fetal erythroblasts for non-invasive prenatal diagnosis. Molecular Human Reproduction. 2003;9(4):227–235. doi: 10.1093/molehr/gag027. [DOI] [PubMed] [Google Scholar]
- 40.Christensen B, Kølvraa S, Lykke-Hansen L, et al. Studies on the isolation and identification of fetal nucleated red blood cells in the circulation of pregnant women before and after chorion villus sampling. Fetal Diagnosis and Therapy. 2003;18(5):376–384. doi: 10.1159/000071983. [DOI] [PubMed] [Google Scholar]
- 41.Rodriguez de Alba M, Palomino P, Gonzalez-Gonzalez C, et al. Prenatal diagnosis on fetal cells from maternal blood: practical comparative evaluation of the first and second trimesters. Prenatal Diagnosis. 2001;21(3):165–170. doi: 10.1002/1097-0223(200103)21:3<165::aid-pd29>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 42.Choolani M, O'Donnell H, Campagnoli C, et al. Simultaneous fetal cell identification and diagnosis by epsilon-globin chain immunophenotyping and chromosomal fluorescence in situ hybridization. Blood. 2001;98(3):554–557. doi: 10.1182/blood.v98.3.554. [DOI] [PubMed] [Google Scholar]