

Summary
The connection between integrins and actin is driving the field of cell migration in new directions. Integrins and actin are coupled through a physical linkage, which provides traction for migration. Recent studies show the importance of this linkage in regulating adhesion organization and development. Actin polymerization orchestrates adhesion assembly near the leading edge of a migrating cell, and the dynamic cross-linking of actin filaments promotes adhesion maturation. Breaking the linkage between actin and integrins leads to adhesion disassembly. Recent quantitative studies have revealed points of slippage in the linkage between actin and integrins, showing that it is not always efficient. Regulation of the assembly and organization of adhesions and their linkage to actin relies on signaling pathways that converge on components that control actin polymerization and organization.
Keywords: Actin, Integrin, Migration, Adhesion
Introduction
Cell migration is a highly complex and regulated process, in which intracellular and extracellular signals conjoin to produce a coordinated response (Lauffenburger and Horwitz, 1996; Ridley et al., 2003; Vicente-Manzanares et al., 2005). The migratory cycle consists of well-defined yet integrated steps that include: front-to-back polarization in response to extracellular cues, which are often chemotactic; membrane extension by protrusion and adhesion formation and cell-body translocation; adhesion disassembly; and rear retraction. Front-to-back polarization involves the local activation of signaling proteins such as phosphatidylinositol 3-kinase (PI3K) at the leading edge (Van Haastert and Devreotes, 2004), as well as the activation of the Cdc42-Par6-atypical protein kinase C (aPKC) pathway, which polarizes cellular structures such as the microtubule organizing center (MTOC), Golgi complex and nucleus (Etienne-Manneville and Hall, 2001).
Our current understanding of how protrusions and adhesions form derives largely from focused studies that identify the two as disparate processes. Actin, however, is required for both, and the actin cytoskeleton is emerging as a key component that links protrusion at the leading edge to adhesion assembly, maturation and turnover. Actin is the underlying molecular machinery that drives protrusions; it adopts different organizations in lamellipodia and filopodia, the two main types of protrusive structures. In lamellipodia, the small GTPases Cdc42 and Rac activate adaptor and/or signaling molecules of the Wiskott-Aldrich syndrome family of proteins such as WASP (Wiskott-Aldrich syndrome protein) and WAVE (WASP-family verprolin-homologous protein), which in turn activate the Arp2/3 complex. This complex nucleates actin-filament branches, and results in a broad dendritic-like actin network (Jaffe and Hall, 2005; Pollard and Borisy, 2003). In filopodia, Cdc42 promotes linear actin polymerization via formins and vasodilator-stimulated phosphoprotein (VASP), and fascin organizes polymerized actin into elongated filaments (Mattila and Lappalainen, 2008).
Here, we will focus on integrin-dependent fibroblast-like migration, although integrin-independent migration is observed in other cell types, such as neuronal cells in the brain and tumor cells, under some conditions (Fackler and Grosse, 2008; Hatten, 2002; Lammermann et al., 2008; Maness and Schachner, 2007; Wolf et al., 2007). Fibroblasts migrate on substrates that are coated with matrix molecules (e.g. fibronectin) via integrins, a family of heterodimeric transmembrane receptors that comprise an α- and a β-subunit (Hynes, 2002).
Integrins bind to extracellular ligands via their outer domains, whereas their internal, cytoplasmic domains are linked to a multitude of structural and signaling molecules as well as the actin cytoskeleton (Humphries and Newham, 1998; Hynes, 1992; Zaidel-Bar et al., 2007a). A unique property of integrins is that their affinity is `tunable', that is, they can switch from low to high affinity by activation of adaptor and signaling molecules that induce changes in the conformation of the integrin (Carman and Springer, 2003; Hynes, 2002). In protrusions, integrins can reside in a high-affinity state (Laudanna et al., 2002; Schwartz and Ginsberg, 2002).
Integrins nucleate the formation of supramolecular assemblies of structural and signaling proteins. These integrin-mediated adhesions serve as traction points for contractile or tensional forces through their interaction with actin (Balaban et al., 2001; Beningo et al., 2001). Adhesions are also signaling centers – they regulate the activation of Rho GTPases in protusions (Kraynov et al., 2000; Nalbant et al., 2004), which in turn control the polymerization and organization of actin and actomyosin filaments, as well as adhesion formation and maturation (Jaffe and Hall, 2005). The coordination between adhesion and the organization of the actin cytoskeleton that is required for cell migration is highlighted by the biphasic migratory response to substrate concentration (DiMilla et al., 1993; Huttenlocher et al., 1996; Palecek et al., 1997). Adjustment of the activation of signaling endpoints such as myosin II, which regulates adhesion and actin organization, can compensate for suboptimal adhesive conditions (Gupton and Waterman-Storer, 2006).
Although the existence of the integrin-actin linkage has been known for many years, it is becoming increasingly clear that its role is more subtle and pervasive than originally anticipated. Near the leading edge, the actin cytoskeleton dictates where and how adhesions assemble, mature and disassemble (turnover) by serving as a dynamic scaffolding network. In return, migration-related signaling pathways converge on integrin-mediated adhesions and regulate actin polymerization and organization.
In this Commentary, we will discuss the relationship between the actin cytoskeleton, adhesion formation and composition, and the strength and regulation of the integrin-actin linkage in the context of cell migration. In doing so, we will capture the importance of actin in nearly all facets of cell migration. We will reveal a shift in the current view of the connection between actin and the process of cell adhesion, and suggest that the actin cytoskeleton directs adhesion assembly, disassembly and organization. We will also show that the composition and strength of the linkage between actin and integrins determine the morphology, growth and stability of the adhesions.
The actin-integrin linkage mediates adhesion stability
The linkage between integrin receptors and the actin cytoskeleton appears to be crucial for the integrity and stability of adhesions. The existence of this linkage and the molecules of which it is comprised (e.g. talin, vinculin and α-actinin) have been known for decades (Fig. 1). Despite this, a plethora of biochemical interactions among these molecules has made it difficult to ascertain the precise physical connection between integrins and actin. For example, both talin and α-actinin have been reported to bind to integrin cytoplasmic domains and actin (Hemmings et al., 1996; Horwitz et al., 1986; Otey et al., 1990). Vinculin, another actin-binding protein, also binds to talin and α-actinin (Burridge and Mangeat, 1984; Gilmore and Burridge, 1996; Johnson and Craig, 1995). Thus, it is likely that multiple routes connect integrins and actin (Volberg et al., 1995), and the decision about which connecting pathways to use might vary among cell types and even individual adhesions.
Fig. 1.
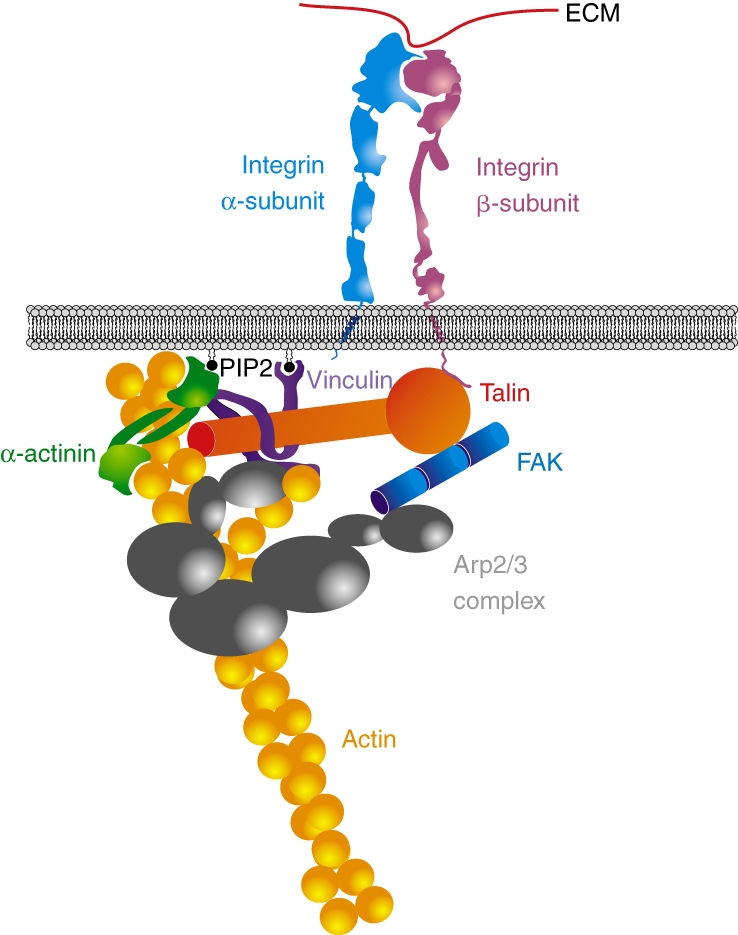
The actin-integrin linkage. The linkage between the extracellular matrix (ECM, red strand on top) and the actin cytoskeleton (represented by yellow beaded coils) is depicted. Integrins (represented by the α- and β-transmembrane subunits in light blue and pink) can bind directly to the talin head domain (red sphere). Through its tail domain (red rod), talin can bind directly to actin as well as to other components of the linkage, such as vinculin (shown in purple). Vinculin can also bind to actin directly, as well as to the actin cross-linker α-actinin (shown as a dimer, in green). Both vinculin and α-actinin are anchored to the membrane, and their activity is modulated by interactions with phosphatidylinositol (4,5)-bisphosphate (PIP2). Finally, vinculin and FAK (shown in blue) can bind to the actin nucleator Arp2/3 (shown as a heptamer in grey).
Disruption of any of the components of the linkage between the substratum and the actin compromises the integrity of adhesions and can lead to their disassembly. Nascent adhesions disassemble as the back edge of the lamellipodium, which is a region of active actin depolymerization and reorganization, passes by them (Choi et al., 2008). Talin, which activates integrins and links them to actin, is required for adhesion stability (Zhang et al., 2008). Calpain, a calcium-dependent protease, mediates adhesion disassembly, at least in part, by its action on talin. When talin is mutated in its calpain-cleavage site and introduced into cells, the adhesions become stable and show impaired disassembly (Franco et al., 2004). Talin-knockout cells are also unable to generate force at stimulated putative adhesion sites that are created by ligand-coated beads in an optical trap (Jiang et al., 2003); they also show unstable adhesions in spreading assays (Zhang et al., 2008).
Disruption of other components of the linkage affects adhesion dynamics in different ways. Vinculin seems to be dispensable for the formation of mature and stable adhesions (Volberg et al., 1995), but its genetic deletion increases cell migration (Xu et al., 1998). In addition, cells in which α-actinin has been knocked down form stable adhesions, but they are small and reside as puncta along actin filaments rather than only at their ends (Choi et al., 2008). Finally, integrin affinity, which is regulated by numerous signaling pathways that lead to integrin activation, also contributes to stability. Activated integrins, which show high affinity for their ligand, appear to reside in adhesions near the leading edge (Kiosses et al., 2001); in lymphocytes, the integrin αLβ2 is in an intermediate-affinity conformation and is also found near the leading edge (Stanley et al., 2008). However, cells in which integrins cannot be activated do not form stable adhesions (Zhang et al., 2008).
Thus, the integrity of the connection between the substratum (via integrins) and the actin cytoskeleton is essential to ensure the proper assembly and disassembly of adhesions, which is required for cell migration. This connection includes the capability of the integrin to be activated, normal expression or function of the scaffold molecules that connect the integrin to the actin, and polymerization and organization of the actin.
Actin organization determines adhesion morphology
Emerging evidence suggests that actin organizes the adhesion rather than vice versa. Cells that do not display highly bundled actin structures, such as leukocytes, tend to have small, less prominent adhesions (Smith et al., 2005). In migrating fibroblasts, the actin is dendritic rather than highly bundled in the lamellipodium, which is located near the leading edge of protrusions (Svitkina and Borisy, 1999). This region is populated by small, punctate, nascent adhesions that remain stationary (Fig. 2). Similar small structures near the lamellipodium, termed `focal complexes', have been observed by others (Nobes and Hall, 1995; Rottner et al., 1999; Zaidel-Bar et al., 2003). They were defined as structures that form in cells that express constitutively active Rac and depend on myosin II activity (Nobes and Hall, 1995; Rottner et al., 1999). Although nascent adhesions require Rac activation (Nayal et al., 2006), they are smaller and reside exclusively inside the lamellipodium.
Fig. 2.

The adhesion lifetime. Adhesions first form in the lamellipodium, where their rate of assembly correlates with the rate of protrusion. As actin disassembles at the rear of the lamellipodium, adhesions turn over. Some adhesions elongate in the region of convergence between the lamellipodium and the lamellum, and mature centripetally along thin actin filaments that are decorated with α-actinin, an actin cross-linker (depicted in red). As the bundles become thicker and more stable owing to enhanced cross-linking, myosin II (depicted in green) enters and the adhesions become larger.
By contrast, adhesions mature by elongation on filamentous actin when the protrusion pauses (Giannone et al., 2007). The organization of the actin appears to be a crucial factor in mediating adhesion elongation. Centripetal maturation occurs when the actin organization changes from dendritic to linear and filamentous in the vicinity of the adhesion (Fig. 2). It occurs along a physical template, which comprises actin filaments and α-actinin, an actin cross-linker. α-actinin is crucial for organizing these templates. In its absence, actin filaments are short, not highly bundled, and randomly oriented rather than centripetal. Corresponding to this change in actin organization the adhesions are no longer elongated; instead, they are a plethora of punctate dots that decorate the actin filaments in their entirety (Choi et al., 2008). Large, organized actin filaments and adhesions are not found in either the α-actinin knockdowns or cells that express dominant interfering mutants (Triplett and Pavalko, 2006). This suggests that actin stability by cross-linking is a defining factor in organizing actin and, in turn, organizing the adhesion.
The tension and bundling that are produced by myosin II activity also contribute to actin organization and adhesion morphology. Large, contractile actomyosin filament bundles terminate at large and stable adhesions and correlate with a proportional increase in adhesion size (Chrzanowska-Wodnicka and Burridge, 1996; Hotulainen and Lappalainen, 2006). Also, cells that display an extended tail contain thick actin filaments that end in large, elongated adhesions; they are generated by myosin II activation at the rear (Vicente-Manzanares et al., 2008). An interesting feature of myosin II is that it does not reside in adhesions; instead, it localizes along actin filaments that interact with the adhesions. Thus, myosin II appears to `act at a distance', perhaps by moving actin filaments together and thereby bundling them. This suggests a prominent role for the cross-linking properties of myosin II in organizing the actin bundles that are associated with maturing adhesions. When the contractility of myosin II is inhibited, actin bundles are no longer prominent and the adhesions are not elongated. However, a non-contractile mutant of myosin II mutants mediates the formation of actin bundles in protrusions and elongation of maturing adhesions even when all contractility is inhibited, again reinforcing the notion that actin cross-linking triggers this process (Choi et al., 2008). Although a role for myosin II in actin bundling appears to be evident, contractility is also important, particularly at later stages (Chrzanowska-Wodnicka and Burridge, 1996; Giannone et al., 2007).
In summary, the morphology of adhesions depends on the structure of the actin in which they reside. Unbundled actin supports small adhesions, whereas bundled actin, produced either by direct interaction with actin cross-linkers or by distal, myosin-II-generated tension, supports larger, elongated adhesions, which assemble along the actin bundles themselves.
Actin polymerization and adhesion formation
Recent data suggest that actin polymerization drives adhesion assembly. Comet-like actin tails associate with integrins near the leading edge and induce β1-integrin clustering at the tip of newly formed actin ripples or filopodia; cytochalasin D inhibits this movement (Galbraith et al., 2007). Actin treadmilling (i.e. the anterior movement of disassembling actin monomers towards the assembling barbed ends) in these ripples seems to drive integrin clustering and lateral movement. This may constitute an exploratory mechanism for haptotaxis, or substrate-driven migration (Galbraith et al., 2007). The tight correlation between the rate of nascent adhesion assembly and protrusion also points to a role for actin polymerization in adhesion assembly. As the lamellipodium advances via actin polymerization, nascent adhesions assemble at a rate that correlates linearly with the protrusion rate, and cytochalasin D, which inhibits actin polymerization, prevents the formation of new nascent adhesions (Choi et al., 2008).
The coupling between adhesion assembly and actin polymerization has important implications. It is possible that the binding of adhesion precursors to polymerizing actin nucleates adhesions. This points to an alternative model for adhesion assembly. The traditional view of adhesion formation envisions integrin ligation mediating its activation and/or the clustering of adhesion components to initiate a nascent adhesion (Calderwood et al., 2000; Carman and Springer, 2003). However, the observation that adhesion assembly requires actin polymerization suggests that the interaction of a subset of adhesion components with actin nucleates the nascent adhesion, which is then stabilized by its association with integrins. In this context, the direct interaction of focal adhesion kinase (FAK) and vinculin with the Arp2/3 complex (DeMali et al., 2002; Serrels et al., 2007), the main nucleator of actin branching and polymerization in lamellipodia, constitutes a possible mechanism for targeting vinculin and FAK to future adhesion sites. The presence of activated integrins in regions of protrusion outside adhesions suggests that they enter the forming adhesion in an activated state (Galbraith et al., 2007; Kiosses et al., 2001).
The other implication is that adhesions might nucleate actin polymerization. This would provide a mechanism for the formation of actin filaments on which adhesions elongate; these appear to elongate from nascent adhesions at the lamellipodium-lamellum interface. This possibility is supported by the observation that purified integrin-adhesion complexes have actin-polymerization activity (Butler et al., 2006). Although the neutralization of Arp2/3 in β3-integrin-containing adhesion complexes did not impair actin polymerization, targeting of the formin mDia did (Butler et al., 2006).
Thus, actin polymerization and adhesion formation are linked. Actin polymerization determines the rate of adhesion assembly and potentially nucleates adhesions that contain activated integrins; conversely, adhesions provide nucleation points that may support actin polymerization. Adhesions and actin are also physically linked and this linkage coordinates adhesion assembly and disassembly and the processes they regulate.
The integrin-actin linkage functions as a molecular clutch
The linkage between the substratum and actin serves as a molecular clutch that transmits actomyosin-generated force to the substratum. The clutch hypothesis stemmed from the biochemical demonstration of an integrin-actin linkage (Horwitz et al., 1986), and it was originally proposed to explain the forward extension of growth cones (Jay, 2000; Mitchison and Kirschner, 1988). Although the concept is old, the connection between the dynamics of actin filaments with that of adhesions is recent. The implications of this linkage are highly significant for cell migration – it can regulate migration through its effect on protrusion rates, as well as adhesion-dependent mechanotransduction (i.e. signaling that is induced by mechanical forces).
The clutch hypothesis integrates adhesion, retrograde flow and actin polymerization. The retrograde flow of actin, generated by either myosin II contraction or membrane resistance to actin polymerization at the leading edge, counteracts the protrusive effect of polarized actin polymerization in protrusions. Thus, the net extension rate of a protrusion is the difference between the actin polymerization and the retrograde flow rates. Adhesions can modulate the protrusion mechanism by linking actin filaments to a fixed substratum, thereby creating traction points that inhibit actin retrograde flow (Alexandrova et al., 2008). This linkage would, therefore, increase the protrusion rate by shunting the forces that drive retrograde flow to the substratum and thereby inhibiting the retrograde flow (Fig. 3; supplementary material Movies 1 and 2). The weakening of such a substratum-integrin-actin linkage would increase the retrograde flow rate and increase the protrusion rate, whereas decreasing the retrograde flow, by inhibiting myosin II, would increase the overall protrusion rate.
Fig. 3.

The integrin-actin linkage acts as a molecular clutch. (A,B) The linkage is shown in two positions: integrins not linked to actin (disengaged) (A), and integrins linked to actin (engaged) (B). In A, the actin is not anchored to the substratum, and thus the force produced by actin polymerization (P) is counterbalanced by retrograde flow (R) which is caused by myosin contraction and tension on the membrane in the lamellipodium. In the example, they balance and there is no protrusion. In B, actin is coupled to the substratum by the interaction of actin-binding proteins with integrins. Under these conditions, the force generated by the retrograde flow is partially or fully shunted to the substratum (oblique black arrows). The force produced by actin polymerization then exceeds the force that produces retrograde flow, resulting in a higher protrusion rate. New nascent adhesions assemble as the lamellipodium extends. As the protrusion advances, the boundary between the lamellipodium and the lamellum moves forward. In the lamellum, myosin II activity generates a contractile force that drives retrograde flow. Slippage points that result in differential coupling of adhesion proteins to the actin occur at an as-yet-undetermined level between the α-actinin (in green) and the other components of the linkage, and/or at the level of interaction of the integrin with the substrate.
Many observations are in accordance with the clutch notion. Tension is observed in protruding regions in both growth cones and the leading edge of migrating fibroblasts (Beningo et al., 2001). In addition, myosin II inhibition, by many different methods, increases the protrusion rate (Cai et al., 2006; Even-Ram et al., 2007). Finally, inactivating talin, a putative linkage molecule, by CALI (chromophore-assisted laser inactivation) inhibits the protrusion rate in growth cones, pointing to the role of this molecule as a physical component of the linkage (Sydor et al., 1996).
Three imaging techniques have provided new insights into the nature of the integrin-actin linkage by measuring the velocities of adhesion-associated molecules: one follows the movement of fluorescent clusters, or speckles; another uses correlation methods; and the third observes the directed movement of a small photobleached region. By measuring the dynamic coupling (correlation) between the velocity of several adhesion proteins to that of the actin filaments, the efficiency of the linkage has been revealed, and points of slippage identified (Brown et al., 2006; Hu et al., 2007). Integrins are largely static with respect to the substratum and do not move with actin; by contrast, the movement of α-actinin correlates well with that of actin, implying that they are tightly bound. The movement of vinculin and talin also correlates with that of actin, although not as well as α-actinin. However, other signaling components that are associated with adhesions but interact directly with actin, such as paxillin and FAK, show movement that correlates weakly with that of actin. These observations reveal a slippage in the linkage that probably occurs between α-actinin and another component of the linkage.
When adhesions retract, linkage molecules such as vinculin exhibit a robust coupling with actin, suggesting that the retraction or contractile forces are transmitted to the substratum level by an engaged clutch (Hu et al., 2007). The efficiency of this process is also affected by the adhesiveness of the cell. When more integrins are engaged, by increasing the ligand density on the substratum, the coupling becomes more efficient (Brown et al., 2006). Finally, the coupling changes during protrusion. Zyxin, an α-actinin binding partner, does not flux with actin during protrusion; however, when protrusion comes to a halt, zyxin undergoes retrograde flow together with the actin (Guo and Wang, 2007).
Although all of these observations point to a regulation of the actin-integrin linkage, the mechanism of regulation is presently unknown. This regulation could occur at many levels, including phosphorylation of linkage components such as α-actinin (Izaguirre et al., 2001) or interaction with phosphatidylinositol (4,5)-bisphosphate (PIP2), which regulates the affinity of α-actinin and vinculin for actin and talin, respectively (Fraley et al., 2003; Gilmore and Burridge, 1996). Another point of regulation is the modulation of integrin affinity (Kim et al., 2003), which is activated by talin (Tadokoro et al., 2003), probably through Rap1-RIAM (Rap1 interacting adaptor molecule) or kindlin-2 signaling pathways (Han et al., 2006; Montanez et al., 2008). Tension applied to the linkage could directly regulate its engagement by altering the conformation of adhesion components and thereby change their affinity (Sawada et al., 2006; Yoshigi et al., 2005).
Most studies to date have focused on the cytoskeletal connections, but the interaction between integrins and their ligands is also an important part of the linkage. A relatively weak interaction between integrins and their substrate ligands would result in an inefficient coupling (Jurado et al., 2005), as would the interaction with ligand on a pliable substrate. In addition, the effective efficiency of the linkage depends on the forces that impinge on it. This is determined by the regional organization of actin, the magnitude of the force and the nature of the adhesion itself. For example, small, punctate adhesions at the front of the cell are subjected to different forces than those that reside in other cellular locations associated with thick bundles of actin (Beningo et al., 2001).
The ability of adhesion components to correlate their motion to that of actin provides a potential mechanism for adhesion translocation, disassembly and the movement of material away from the protrusion (Guo and Wang, 2007; Wang, 2007). Other studies also suggest that adhesions can translocate by the treadmilling of components rather than a physical sliding (Ballestrem et al., 2001; Digman et al., 2008).
In summary, the clutch hypothesis proposes a hierarchical linkage of integrins to actin, explains the different movement of adhesion proteins with respect to actin by postulating the existence of slippage points, and suggests that signaling regulates the efficiency of the linkage.
Integrin signaling regulates adhesion and protrusion
Integrin signaling regulates protrusion and adhesion in migrating cells. These signals are generated and localized by adhesions that reside in protruding regions. Although a large signaling network converges on adhesion and protrusion, the signaling generally occurs via Rho GTPases, such as Rac, Rho and Cdc42. The migration-related targets of these Rho GTPases are limited largely to regulators of actin polymerization (the Arp2/3 complex and formins) (Pollard, 2007), actin disassembly (LIM-kinase–cofilin) (Wang et al., 2007), myosin II activation or inactivation [myosin light chain (MLC) kinase (MLCK), MLC phosphatase and MLC] (Clark et al., 2007), and microtubule dynamics (Daub et al., 2001) (Fig. 4). Microtubules mediate adhesion disassembly and polarity (Etienne-Manneville and Hall, 2001; Kaverina et al., 1998).
Fig. 4.

Outline of adhesive signaling in migration. Integrin ligation induces the nucleation of different signaling elements. The major categories (kinases, non-catalytic adaptor proteins and actin-binding proteins) are shown. These categories can influence the recruitment and/or activation of other components of adhesions (represented by red arrows). Most migratory signaling converges on the Rho GTPases, which regulate actin polymerization and stability (via nucleators such as the Arp2/3 complex and formins, or actin-filament-severing proteins such as cofilin), actomyosin contractility (via MLC phosphorylation), and microtubules (not shown).
The adhesions in protrusions contain many phosphorylated proteins that mediate Rho-GTPase-enabled signaling. The phosphorylation of these proteins is lower in mature adhesions outside protrusions, suggesting a spatial specificity of the signals that emanate from them (Ballestrem et al., 2006; Zaidel-Bar et al., 2007b). Paxillin has emerged as a switch for the regulation of adhesion turnover and a major hub for Rac and Rho signaling, because it has several different domains that regulate the activity of these Rho GTPases. For example, phosphorylation on Tyr31 and Tyr118 by FAK or Src recruits the adaptor proteins Crk-II and p130CAS (Schaller and Parsons, 1995), which form a complex that activates Rac via the Rac GTP-exchange factor DOCK180 (Reddien and Horvitz, 2000). Cells that express phosphomimetic mutations in these sites become highly protrusive, with many small adhesions that turn over rapidly near the leading edge. A phosphomimetic mutation on Ser273 produces a similar phenotype. Phosphorylation on Ser273 recruits the G-protein adaptor proteins GIT1-GIT2 and the Rac GTP-exchange factor βPIX, which form another complex that also activates Rac and positions active p21-activated kinase-1 (PAK1) in protrusions (Manabe et al., 2002; Manser et al., 1998; Nayal et al., 2006). PAK1 is an effector of Rac and inactivates myosin II by phosphorylating MLCK (Sanders et al., 1999). GIT1 and GIT2 also have an Arf-GAP domain that is implicated in Rac activation through another small GTPase, Arf6 (Nishiya et al., 2005). Paxillin, therefore, functions to assemble Rac regulator-effector complexes in adhesions.
Integrin-initiated signaling can also activate the small GTPase RhoA (Cox et al., 2001; Ren et al., 1999), which in turn activates ROCK (Rho-associated coiled-coil-containing protein kinase) (Ishizaki et al., 1996; Leung et al., 1996; Matsui et al., 1996). ROCK activates myosin II directly, by phosphorylating the myosin II regulatory light chain (R/MLC) (Amano et al., 1996) and via inhibition of the myosin light chain phosphatase (Kimura et al., 1996). The activation of myosin II leads to the formation of large, stable adhesions and highly bundled actin filaments (Chrzanowska-Wodnicka and Burridge, 1996), which inhibit protrusion.
Finally, the antagonistic function of Rac and RhoA in the coordination of protrusion and the assembly of adhesions illustrates the need for a tight spatiotemporal regulation of the activation-inactivation cycles of these molecules to enable cell migration. Biosensors that localize the activation of Rho-family GTPases have revealed such a tight spatiotemporal regulation (Kraynov et al., 2000; Nalbant et al., 2004; Sabouri-Ghomi et al., 2008).
Integrin signaling through adhesions is part of a feedback loop. Myosin II activation increases contraction and actin bundling and also increases the signals that mediate myosin II activation. For example, external strain inhibits Rac activation and promotes adhesion growth (Galbraith et al., 2002; Katsumi et al., 2002; Riveline et al., 2001), and induces the formation of large actin bundles and adhesions that inhibit protrusion. The deletion of myosin II in migrating cells decreases actin bundling and adhesion maturation, leaving a rim of small adhesions at the edge of protrusions that are phosphorylated on paxillin and FAK (Vicente-Manzanares et al., 2007).
In summary, integrin-mediated signaling controls the production and organization of actin filaments mainly via the activation and inactivation of Rho GTPases and their downstream effectors. This vast signaling network also includes feedback loops that potentially regulate integrin aggregation and activation, and adhesion assembly and disassembly.
Concluding remarks and future perspectives
The connection between integrins and actin is complex and multifaceted. This perspective has pointed to several new themes in the interplay between the substratum and actin. First, the physical linkage of actin to integrins dictates the stability of adhesions. Second, the local organization of actin dictates the organization of the adhesions rather than vice versa. Third, adhesions mature by elongating along actin filaments. Fourth, the formation of adhesions is coupled to the polymerization of the actin. Fifth, the linkage between the integrin and actin is regulated. The efficiency and regulation of this physical linkage provides a checkpoint that controls protrusion, cell migration, adhesion dynamics and mechanotransduction. Sixth, integrin signaling regulates protrusion and adhesion formation through a small set of targets that regulate actin polymerization-depolymerization and myosin II activity.
These new themes are probably only the tip of the iceberg. The technology is now available to elucidate the interplay between the structural and regulatory relationships between integrins and actin in detail. Answers to the following questions about migrating cells – Why do some cells migrate faster or in a more directed manner than others? What is the nature of the organization of adhesions and actin in cells growing in three dimensions or in vivo? Do changes in the integrin-actin linkage regulate mechanotransduction? – are likely to lie in understanding the integrin-actin linkage, and now seem approachable.
Supplementary Material
Supplementary material available online at http://jcs.biologists.org/cgi/content/full/122/2/199/DC1
The authors acknowledge the work from many colleagues who were not cited because of space constraints. This work was supported by NIH grants GM23244 and the Cell Migration Consortium (U54 GM064346). Deposited in PMC for release after 12 months.
References
- Alexandrova, A. Y., Arnold, K., Schaub, S. B., Vasiliev, J. M., Meister, J.-J., Bershadsky, A. D. and Verkhovsky, A. B. (2008). Comparative dynamics of retrograde actin flow and focal adhesions: formation of nascent adhesions triggers transition from fast to slow flow. PLoS ONE 3, e3234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amano, M., Ito, M., Kimura, K., Fukata, Y., Chihara, K., Nakano, T., Matsuura, Y. and Kaibuchi, K. (1996). Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase). J. Biol. Chem. 271, 20246-20249. [DOI] [PubMed] [Google Scholar]
- Balaban, N. Q., Schwarz, U. S., Riveline, D., Goichberg, P., Tzur, G., Sabanay, I., Mahalu, D., Safran, S., Bershadsky, A., Addadi, L. et al. (2001). Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 3, 466-472. [DOI] [PubMed] [Google Scholar]
- Ballestrem, C., Hinz, B., Imhof, B. A. and Wehrle-Haller, B. (2001). Marching at the front and dragging behind: differential alphaVbeta3-integrin turnover regulates focal adhesion behavior. J. Cell Biol. 155, 1319-1332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballestrem, C., Erez, N., Kirchner, J., Kam, Z., Bershadsky, A. and Geiger, B. (2006). Molecular mapping of tyrosine-phosphorylated proteins in focal adhesions using fluorescence resonance energy transfer. J. Cell Sci. 119, 866-875. [DOI] [PubMed] [Google Scholar]
- Beningo, K. A., Dembo, M., Kaverina, I., Small, J. V. and Wang, Y. L. (2001). Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J. Cell Biol. 153, 881-888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, C. M., Hebert, B., Kolin, D. L., Zareno, J., Whitmore, L., Horwitz, A. R. and Wiseman, P. W. (2006). Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214. [DOI] [PubMed] [Google Scholar]
- Burridge, K. and Mangeat, P. (1984). An interaction between vinculin and talin. Nature 308, 744-746. [DOI] [PubMed] [Google Scholar]
- Butler, B., Gao, C., Mersich, A. T. and Blystone, S. D. (2006). Purified integrin adhesion complexes exhibit actin-polymerization activity. Curr. Biol. 16, 242-251. [DOI] [PubMed] [Google Scholar]
- Cai, Y., Biais, N., Giannone, G., Tanase, M., Jiang, G., Hofman, J. M., Wiggins, C. H., Silberzan, P., Buguin, A., Ladoux, B. et al. (2006). Nonmuscle myosin IIA-dependent force inhibits cell spreading and drives F-actin flow. Biophys. J. 91, 3907-3920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calderwood, D. A., Shattil, S. J. and Ginsberg, M. H. (2000). Integrins and actin filaments: reciprocal regulation of cell adhesion and signaling. J. Biol. Chem. 275, 22607-22610. [DOI] [PubMed] [Google Scholar]
- Carman, C. V. and Springer, T. A. (2003). Integrin avidity regulation: are changes in affinity and conformation underemphasized? Curr. Opin. Cell Biol. 15, 547-556. [DOI] [PubMed] [Google Scholar]
- Choi, C. K., Vicente-Manzanares, M., Zareno, J., Whitmore, L. A., Mogilner, A. and Horwitz, A. R. (2008). Actin and alpha-actinin orchestrate the assembly and maturation of nascent adhesions in a myosin II motor-independent manner. Nat. Cell Biol. 10, 1039-1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrzanowska-Wodnicka, M. and Burridge, K. (1996). Rho-stimulated contractility drives the formation of stress fibers and focal adhesions. J. Cell Biol. 133, 1403-1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark, K., Langeslag, M., Figdor, C. G. and van Leeuwen, F. N. (2007). Myosin II and mechanotransduction: a balancing act. Trends Cell Biol. 17, 178-186. [DOI] [PubMed] [Google Scholar]
- Cox, E. A., Sastry, S. K. and Huttenlocher, A. (2001). Integrin-mediated adhesion regulates cell polarity and membrane protrusion through the Rho family of GTPases. Mol. Biol. Cell 12, 265-277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daub, H., Gevaert, K., Vandekerckhove, J., Sobel, A. and Hall, A. (2001). Rac/Cdc42 and p65PAK regulate the microtubule-destabilizing protein stathmin through phosphorylation at serine 16. J. Biol. Chem. 276, 1677-1680. [DOI] [PubMed] [Google Scholar]
- DeMali, K. A., Barlow, C. A. and Burridge, K. (2002). Recruitment of the Arp2/3 complex to vinculin: coupling membrane protrusion to matrix adhesion. J. Cell Biol. 159, 881-891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Digman, M. A., Brown, C. M., Horwitz, A. R., Mantulin, W. W. and Gratton, E. (2008). Paxillin dynamics measured during adhesion assembly and disassembly by correlation spectroscopy. Biophys. J. 94, 2819-2831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiMilla, P. A., Stone, J. A., Quinn, J. A., Albelda, S. M. and Lauffenburger, D. A. (1993). Maximal migration of human smooth muscle cells on fibronectin and type IV collagen occurs at an intermediate attachment strength. J. Cell Biol. 122, 729-737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etienne-Manneville, S. and Hall, A. (2001). Integrin-mediated activation of Cdc42 controls cell polarity in migrating astrocytes through PKCzeta. Cell 106, 489-498. [DOI] [PubMed] [Google Scholar]
- Even-Ram, S., Doyle, A. D., Conti, M. A., Matsumoto, K., Adelstein, R. S. and Yamada, K. M. (2007). Myosin IIA regulates cell motility and actomyosin-microtubule crosstalk. Nat. Cell Biol. 9, 299-309. [DOI] [PubMed] [Google Scholar]
- Fackler, O. T. and Grosse, R. (2008). Cell motility through plasma membrane blebbing. J. Cell Biol. 181, 879-884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraley, T. S., Tran, T. C., Corgan, A. M., Nash, C. A., Hao, J., Critchley, D. R. and Greenwood, J. A. (2003). Phosphoinositide binding inhibits alpha-actinin bundling activity. J. Biol. Chem. 278, 24039-24045. [DOI] [PubMed] [Google Scholar]
- Franco, S. J., Rodgers, M. A., Perrin, B. J., Han, J., Bennin, D. A., Critchley, D. R. and Huttenlocher, A. (2004). Calpain-mediated proteolysis of talin regulates adhesion dynamics. Nat. Cell Biol. 6, 977-983. [DOI] [PubMed] [Google Scholar]
- Galbraith, C. G., Yamada, K. M. and Sheetz, M. P. (2002). The relationship between force and focal complex development. J. Cell Biol. 159, 695-705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galbraith, C. G., Yamada, K. M. and Galbraith, J. A. (2007). Polymerizing actin fibers position integrins primed to probe for adhesion sites. Science 315, 992-995. [DOI] [PubMed] [Google Scholar]
- Giannone, G., Dubin-Thaler, B. J., Rossier, O., Cai, Y., Chaga, O., Jiang, G., Beaver, W., Dobereiner, H. G., Freund, Y., Borisy, G. et al. (2007). Lamellipodial actin mechanically links myosin activity with adhesion-site formation. Cell 128, 561-575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilmore, A. P. and Burridge, K. (1996). Regulation of vinculin binding to talin and actin by phosphatidyl-inositol-4-5-bisphosphate. Nature 381, 531-535. [DOI] [PubMed] [Google Scholar]
- Guo, W. H. and Wang, Y. L. (2007). Retrograde fluxes of focal adhesion proteins in response to cell migration and mechanical signals. Mol. Biol. Cell 18, 4519-4527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupton, S. L. and Waterman-Storer, C. M. (2006). Spatiotemporal feedback between actomyosin and focal-adhesion systems optimizes rapid cell migration. Cell 125, 1361-1374. [DOI] [PubMed] [Google Scholar]
- Han, J., Lim, C. J., Watanabe, N., Soriani, A., Ratnikov, B., Calderwood, D. A., Puzon-McLaughlin, W., Lafuente, E. M., Boussiotis, V. A., Shattil, S. J. et al. (2006). Reconstructing and deconstructing agonist-induced activation of integrin alphaIIbbeta3. Curr. Biol. 16, 1796-1806. [DOI] [PubMed] [Google Scholar]
- Hatten, M. E. (2002). New directions in neuronal migration. Science 297, 1660-1663. [DOI] [PubMed] [Google Scholar]
- Hemmings, L., Rees, D. J., Ohanian, V., Bolton, S. J., Gilmore, A. P., Patel, B., Priddle, H., Trevithick, J. E., Hynes, R. O. and Critchley, D. R. (1996). Talin contains three actin-binding sites each of which is adjacent to a vinculin-binding site. J. Cell Sci. 109, 2715-2726. [DOI] [PubMed] [Google Scholar]
- Horwitz, A., Duggan, K., Buck, C., Beckerle, M. C. and Burridge, K. (1986). Interaction of plasma membrane fibronectin receptor with talin-a transmembrane linkage. Nature 320, 531-533. [DOI] [PubMed] [Google Scholar]
- Hotulainen, P. and Lappalainen, P. (2006). Stress fibers are generated by two distinct actin assembly mechanisms in motile cells. J. Cell Biol. 173, 383-394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu, K., Ji, L., Applegate, K. T., Danuser, G. and Waterman-Storer, C. M. (2007). Differential transmission of actin motion within focal adhesions. Science 315, 111-115. [DOI] [PubMed] [Google Scholar]
- Humphries, M. J. and Newham, P. (1998). The structure of cell-adhesion molecules. Trends Cell Biol. 8, 78-83. [PubMed] [Google Scholar]
- Huttenlocher, A., Ginsberg, M. H. and Horwitz, A. F. (1996). Modulation of cell migration by integrin-mediated cytoskeletal linkages and ligand-binding affinity. J. Cell Biol. 134, 1551-1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hynes, R. O. (1992). Integrins: versatility, modulation, and signaling in cell adhesion. Cell 69, 11-25. [DOI] [PubMed] [Google Scholar]
- Hynes, R. O. (2002). Integrins: bidirectional, allosteric signaling machines. Cell 110, 673-687. [DOI] [PubMed] [Google Scholar]
- Ishizaki, T., Maekawa, M., Fujisawa, K., Okawa, K., Iwamatsu, A., Fujita, A., Watanabe, N., Saito, Y., Kakizuka, A., Morii, N. et al. (1996). The small GTP-binding protein Rho binds to and activates a 160 kDa Ser/Thr protein kinase homologous to myotonic dystrophy kinase. EMBO J. 15, 1885-1893. [PMC free article] [PubMed] [Google Scholar]
- Izaguirre, G., Aguirre, L., Hu, Y. P., Lee, H. Y., Schlaepfer, D. D., Aneskievich, B. J. and Haimovich, B. (2001). The cytoskeletal/non-muscle isoform of alpha-actinin is phosphorylated on its actin-binding domain by the focal adhesion kinase. J. Biol. Chem. 276, 28676-28685. [DOI] [PubMed] [Google Scholar]
- Jaffe, A. B. and Hall, A. (2005). Rho GTPases: biochemistry and biology. Annu. Rev. Cell Dev. Biol. 21, 247-269. [DOI] [PubMed] [Google Scholar]
- Jay, D. G. (2000). The clutch hypothesis revisited: ascribing the roles of actin-associated proteins in filopodial protrusion in the nerve growth cone. J. Neurobiol. 44, 114-125. [DOI] [PubMed] [Google Scholar]
- Jiang, G., Giannone, G., Critchley, D. R., Fukumoto, E. and Sheetz, M. P. (2003). Two-piconewton slip bond between fibronectin and the cytoskeleton depends on talin. Nature 424, 334-337. [DOI] [PubMed] [Google Scholar]
- Johnson, R. P. and Craig, S. W. (1995). F-actin binding site masked by the intramolecular association of vinculin head and tail domains. Nature 373, 261-264. [DOI] [PubMed] [Google Scholar]
- Jurado, C., Haserick, J. R. and Lee, J. (2005). Slipping or gripping? Fluorescent speckle microscopy in fish keratocytes reveals two different mechanisms for generating a retrograde flow of actin. Mol. Biol. Cell 16, 507-518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katsumi, A., Milanini, J., Kiosses, W. B., del Pozo, M. A., Kaunas, R., Chien, S., Hahn, K. M. and Schwartz, M. A. (2002). Effects of cell tension on the small GTPase Rac. J. Cell Biol. 158, 153-164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaverina, I., Rottner, K. and Small, J. V. (1998). Targeting, capture, and stabilization of microtubules at early focal adhesions. J. Cell Biol. 142, 181-190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, M., Carman, C. V. and Springer, T. A. (2003). Bidirectional transmembrane signaling by cytoplasmic domain separation in integrins. Science 301, 1720-1725. [DOI] [PubMed] [Google Scholar]
- Kimura, K., Ito, M., Amano, M., Chihara, K., Fukata, Y., Nakafuku, M., Yamamori, B., Feng, J., Nakano, T., Okawa, K. et al. (1996). Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho-kinase). Science 273, 245-248. [DOI] [PubMed] [Google Scholar]
- Kiosses, W. B., Shattil, S. J., Pampori, N. and Schwartz, M. A. (2001). Rac recruits high-affinity integrin alphavbeta3 to lamellipodia in endothelial cell migration. Nat. Cell Biol. 3, 316-320. [DOI] [PubMed] [Google Scholar]
- Kraynov, V. S., Chamberlain, C., Bokoch, G. M., Schwartz, M. A., Slabaugh, S. and Hahn, K. M. (2000). Localized Rac activation dynamics visualized in living cells. Science 290, 333-337. [DOI] [PubMed] [Google Scholar]
- Lammermann, T., Bader, B. L., Monkley, S. J., Worbs, T., Wedlich-Soldner, R., Hirsch, K., Keller, M., Forster, R., Critchley, D. R., Fassler, R. et al. (2008). Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 453, 51-55. [DOI] [PubMed] [Google Scholar]
- Laudanna, C., Kim, J. Y., Constantin, G. and Butcher, E. (2002). Rapid leukocyte integrin activation by chemokines. Immunol. Rev. 186, 37-46. [DOI] [PubMed] [Google Scholar]
- Lauffenburger, D. A. and Horwitz, A. F. (1996). Cell migration: a physically integrated molecular process. Cell 84, 359-369. [DOI] [PubMed] [Google Scholar]
- Leung, T., Chen, X. Q., Manser, E. and Lim, L. (1996). The p160 RhoA-binding kinase ROK alpha is a member of a kinase family and is involved in the reorganization of the cytoskeleton. Mol. Cell. Biol. 16, 5313-5327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manabe, R., Kovalenko, M., Webb, D. J. and Horwitz, A. R. (2002). GIT1 functions in a motile, multi-molecular signaling complex that regulates protrusive activity and cell migration. J. Cell Sci. 115, 1497-1510. [DOI] [PubMed] [Google Scholar]
- Maness, P. F. and Schachner, M. (2007). Neural recognition molecules of the immunoglobulin superfamily: signaling transducers of axon guidance and neuronal migration. Nat. Neurosci. 10, 19-26. [DOI] [PubMed] [Google Scholar]
- Manser, E., Loo, T. H., Koh, C. G., Zhao, Z. S., Chen, X. Q., Tan, L., Tan, I., Leung, T. and Lim, L. (1998). PAK kinases are directly coupled to the PIX family of nucleotide exchange factors. Mol. Cell 1, 183-192. [DOI] [PubMed] [Google Scholar]
- Maruyama, K. and Ebashi, S. (1965). Alpha-actinin, a new structural protein from striated muscle. II. Action on actin. J. Biochem. 58, 13-19. [DOI] [PubMed] [Google Scholar]
- Matsui, T., Amano, M., Yamamoto, T., Chihara, K., Nakafuku, M., Ito, M., Nakano, T., Okawa, K., Iwamatsu, A. and Kaibuchi, K. (1996). Rho-associated kinase, a novel serine/threonine kinase, as a putative target for small GTP binding protein Rho. EMBO J. 15, 2208-2216. [PMC free article] [PubMed] [Google Scholar]
- Mattila, P. K. and Lappalainen, P. (2008). Filopodia: molecular architecture and cellular functions. Nat. Rev. Mol. Cell Biol. 9, 446-454. [DOI] [PubMed] [Google Scholar]
- Mitchison, T. and Kirschner, M. (1988). Cytoskeletal dynamics and nerve growth. Neuron 1, 761-772. [DOI] [PubMed] [Google Scholar]
- Montanez, E., Ussar, S., Schifferer, M., Bosl, M., Zent, R., Moser, M. and Fassler, R. (2008). Kindlin-2 controls bidirectional signaling of integrins. Genes Dev. 22, 1325-1330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nalbant, P., Hodgson, L., Kraynov, V., Toutchkine, A. and Hahn, K. M. (2004). Activation of endogenous Cdc42 visualized in living cells. Science 305, 1615-1619. [DOI] [PubMed] [Google Scholar]
- Nayal, A., Webb, D. J., Brown, C. M., Schaefer, E. M., Vicente-Manzanares, M. and Horwitz, A. R. (2006). Paxillin phosphorylation at Ser273 localizes a GIT1-PIX-PAK complex and regulates adhesion and protrusion dynamics. J. Cell Biol. 173, 587-589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiya, N., Kiosses, W. B., Han, J. and Ginsberg, M. H. (2005). An alpha4 integrin-paxillin-Arf-GAP complex restricts Rac activation to the leading edge of migrating cells. Nat. Cell Biol. 7, 343-352. [DOI] [PubMed] [Google Scholar]
- Nobes, C. D. and Hall, A. (1995). Rho, Rac, and Cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 81, 53-62. [DOI] [PubMed] [Google Scholar]
- Otey, C. A., Pavalko, F. M. and Burridge, K. (1990). An interaction between alpha-actinin and the beta 1 integrin subunit in vitro. J. Cell Biol. 111, 721-729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palecek, S. P., Loftus, J. C., Ginsberg, M. H., Lauffenburger, D. A. and Horwitz, A. F. (1997). Integrin-ligand binding properties govern cell migration speed through cell-substratum adhesiveness. Nature 385, 537-540. [DOI] [PubMed] [Google Scholar]
- Pollard, T. D. (2007). Regulation of actin filament assembly by Arp2/3 complex and formins. Annu. Rev. Biophys. Biomol. Struct. 36, 451-477. [DOI] [PubMed] [Google Scholar]
- Pollard, T. D. and Borisy, G. G. (2003). Cellular motility driven by assembly and disassembly of actin filaments. Cell 112, 453-465. [DOI] [PubMed] [Google Scholar]
- Reddien, P. W. and Horvitz, H. R. (2000). CED-2/CrkII and CED-10/Rac control phagocytosis and cell migration in Caenorhabditis elegans. Nat. Cell Biol. 2, 131-136. [DOI] [PubMed] [Google Scholar]
- Ren, X. D., Kiosses, W. B. and Schwartz, M. A. (1999). Regulation of the small GTP-binding protein Rho by cell adhesion and the cytoskeleton. EMBO J. 18, 578-585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridley, A. J., Schwartz, M. A., Burridge, K., Firtel, R. A., Ginsberg, M. H., Borisy, G., Parsons, J. T. and Horwitz, A. R. (2003). Cell migration: integrating signals from front to back. Science 302, 1704-1709. [DOI] [PubMed] [Google Scholar]
- Riveline, D., Zamir, E., Balaban, N. Q., Schwarz, U. S., Ishizaki, T., Narumiya, S., Kam, Z., Geiger, B. and Bershadsky, A. D. (2001). Focal contacts as mechanosensors: externally applied local mechanical force induces growth of focal contacts by an mDia1-dependent and ROCK-independent mechanism. J. Cell Biol. 153, 1175-1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rottner, K., Hall, A. and Small, J. V. (1999). Interplay between Rac and Rho in the control of substrate contact dynamics. Curr. Biol. 9, 640-649. [DOI] [PubMed] [Google Scholar]
- Sabouri-Ghomi, M., Wu, Y., Hahn, K. and Danuser, G. (2008). Visualizing and quantifying adhesive signals. Curr. Opin. Cell Biol. 20, 541-550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, L. C., Matsumura, F., Bokoch, G. M. and de Lanerolle, P. (1999). Inhibition of myosin light chain kinase by p21-activated kinase. Science 283, 2083-2085. [DOI] [PubMed] [Google Scholar]
- Sawada, Y., Tamada, M., Dubin-Thaler, B. J., Cherniavskaya, O., Sakai, R., Tanaka, S. and Sheetz, M. P. (2006). Force sensing by mechanical extension of the Src family kinase substrate p130Cas. Cell 127, 1015-1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaller, M. and Parsons, J. (1995). pp125FAK-dependent tyrosine phosphorylation of paxillin creates a high-affinity binding site for Crk. Mol. Cell. Biol. 15, 2635-2645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz, M. A. and Ginsberg, M. H. (2002). Networks and crosstalk: integrin signalling spreads. Nat. Cell Biol. 4, E65-E68. [DOI] [PubMed] [Google Scholar]
- Serrels, B., Serrels, A., Brunton, V. G., Holt, M., McLean, G. W., Gray, C. H., Jones, G. E. and Frame, M. C. (2007). Focal adhesion kinase controls actin assembly via a FERM-mediated interaction with the Arp2/3 complex. Nat. Cell Biol. 9, 1046-1056. [DOI] [PubMed] [Google Scholar]
- Smith, A., Carrasco, Y. R., Stanley, P., Kieffer, N., Batista, F. D. and Hogg, N. (2005). A talin-dependent LFA-1 focal zone is formed by rapidly migrating T lymphocytes. J. Cell Biol. 170, 141-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanley, P., Smith, A., McDowall, A., Nicol, A., Zicha, D. and Hogg, N. (2008). Intermediate-affinity LFA-1 binds alpha-actinin-1 to control migration at the leading edge of the T cell. EMBO J. 27, 62-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svitkina, T. M. and Borisy, G. G. (1999). Arp2/3 complex and actin depolymerizing factor/cofilin in dendritic organization and treadmilling of actin filament array in lamellipodia. J. Cell Biol. 145, 1009-1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sydor, A. M., Su, A. L., Wang, F. S., Xu, A. and Jay, D. G. (1996). Talin and vinculin play distinct roles in filopodial motility in the neuronal growth cone. J. Cell Biol. 134, 1197-1207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tadokoro, S., Shattil, S. J., Eto, K., Tai, V., Liddington, R. C., de Pereda, J. M., Ginsberg, M. H. and Calderwood, D. A. (2003). Talin binding to integrin beta tails: a final common step in integrin activation. Science 302, 103-106. [DOI] [PubMed] [Google Scholar]
- Triplett, J. W. and Pavalko, F. M. (2006). Disruption of alpha-actinin-integrin interactions at focal adhesions renders osteoblasts susceptible to apoptosis. Am. J. Physiol. Cell Physiol. 291, C909-C921. [DOI] [PubMed] [Google Scholar]
- Van Haastert, P. J. and Devreotes, P. N. (2004). Chemotaxis: signalling the way forward. Nat. Rev. Mol. Cell Biol. 5, 626-634. [DOI] [PubMed] [Google Scholar]
- Vicente-Manzanares, M., Webb, D. J. and Horwitz, A. R. (2005). Cell migration at a glance. J. Cell Sci. 118, 4917-4919. [DOI] [PubMed] [Google Scholar]
- Vicente-Manzanares, M., Zareno, J., Whitmore, L., Choi, C. K. and Horwitz, A. F. (2007). Regulation of protrusion, adhesion dynamics, and polarity by myosins IIA and IIB in migrating cells. J. Cell Biol. 176, 573-580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vicente-Manzanares, M., Koach, M. A., Whitmore, L., Lamers, M. L. and Horwitz, A. F. (2008). Segregation and activation of myosin IIB creates a rear in migrating cells. J. Cell Biol. 183, 543-554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volberg, T., Geiger, B., Kam, Z., Pankov, R., Simcha, I., Sabanay, H., Coll, J. L., Adamson, E. and Ben-Ze'ev, A. (1995). Focal adhesion formation by F9 embryonal carcinoma cells after vinculin gene disruption. J. Cell Sci. 108, 2253-2260. [DOI] [PubMed] [Google Scholar]
- Wachsstock, D. H., Wilkins, J. A. and Lin, S. (1987). Specific interaction of vinculin with alpha-actinin. Biochem. Biophys. Res. Commun. 146, 554-560. [DOI] [PubMed] [Google Scholar]
- Wang, W., Eddy, R. and Condeelis, J. (2007). The cofilin pathway in breast cancer invasion and metastasis. Nat. Rev. Cancer 7, 429-440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Y. L. (2007). Flux at focal adhesions: slippage clutch, mechanical gauge, or signal depot. Sci STKE 2007, pe10. [DOI] [PubMed] [Google Scholar]
- Wolf, K., Wu, Y. I., Liu, Y., Geiger, J., Tam, E., Overall, C., Stack, M. S. and Friedl, P. (2007). Multi-step pericellular proteolysis controls the transition from individual to collective cancer cell invasion. Nat. Cell Biol. 9, 893-904. [DOI] [PubMed] [Google Scholar]
- Xu, W., Baribault, H. and Adamson, E. D. (1998). Vinculin knockout results in heart and brain defects during embryonic development. Development 125, 327-337. [DOI] [PubMed] [Google Scholar]
- Yoshigi, M., Hoffman, L. M., Jensen, C. C., Yost, H. J. and Beckerle, M. C. (2005). Mechanical force mobilizes zyxin from focal adhesions to actin filaments and regulates cytoskeletal reinforcement. J. Cell Biol. 171, 209-215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaidel-Bar, R., Ballestrem, C., Kam, Z. and Geiger, B. (2003). Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J. Cell Sci. 116, 4605-4613. [DOI] [PubMed] [Google Scholar]
- Zaidel-Bar, R., Itzkovitz, S., Ma'ayan, A., Iyengar, R. and Geiger, B. (2007a). Functional atlas of the integrin adhesome. Nat. Cell Biol. 9, 858-867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaidel-Bar, R., Milo, R., Kam, Z. and Geiger, B. (2007b). A paxillin tyrosine phosphorylation switch regulates the assembly and form of cell-matrix adhesions. J. Cell Sci. 120, 137-148. [DOI] [PubMed] [Google Scholar]
- Zhang, X., Jiang, G., Cai, Y., Monkley, S. J., Critchley, D. R. and Sheetz, M. P. (2008). Talin depletion reveals independence of initial cell spreading from integrin activation and traction. Nat. Cell Biol. 10, 1062-1068. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.