

Abstract
In the most extensive analysis of body size in marine invertebrates to date, we show that the size–frequency distributions of northeastern Pacific bivalves at the provincial level are surprisingly invariant in modal and median size as well as size range, despite a 4-fold change in species richness from the tropics to the Arctic. The modal sizes and shapes of these size–frequency distributions are consistent with the predictions of an energetic model previously applied to terrestrial mammals and birds. However, analyses of the Miocene–Recent history of body sizes within 82 molluscan genera show little support for the expectation that the modal size is an evolutionary attractor over geological time.
Body size influences almost every aspect of the biology of a species, from physiology to life history (1–4), and plays an important role in the organization of ecological communities (5–8). Size–frequency distributions (SFDs) of species within clades and regional biotas represent a macroecological and macroevolutionary expression of the forces operating on body sizes over large temporal and spatial scales, and several models have attempted to explain the shapes of these distributions (9–13). However, little is known about how SFDs of marine invertebrates vary along major environmental gradients such as latitude, and contradictory predictions exist. For example, some authors have argued that size should increase with latitude within and among species even for ectotherms (14–16), whereas species–energy theory predicts decreasing size with latitude (16, 17), and clade-specific or region-specific effects might overwhelm any general trends (18). In the most extensive biogeographic analysis of body size in marine invertebrates to date, we compare the SFDs of northeastern Pacific bivalve faunas among four biogeographic provinces, spanning 75o of latitude. We then compare the shapes of these SFDs to the predictions of a theoretical model of body size based on energetics (10) and test the evolutionary predictions of this and other optimization models.
Latitudinal Trends in Body Size
Methods.
The latitudinal ranges and body sizes of 915 of the ≈950 species of marine bivalves recorded from the tropics to the Arctic along the northeastern Pacific continental shelf (depth < 200 m) were compiled through an extensive search of the primary literature and from major museum collections (19–21). All bivalve trophic groups are represented, including deposit-feeding protobranchs, epifaunal pterioid and infaunal veneroid suspension-feeders, chemosymbiotic lucinoids, and carnivorous septibranchs. As a measure of body size for each species, we used the geometric mean of length and height of the largest known specimen. This standard metric for living and fossil mollusks (22–24) correlates closely with body mass [for the limited mass data available on the bivalves used in this study, log2[(length)1/2 × height] = 5.507 + 0.316 × log2(mass), r2 = 0.81, P < 0.0001; highly significant relationships between linear shell measurements and body mass are also shown in refs. 25 and 26]. We used log2 intervals to divide the size data into convenient categories; this widely used transformation renders analyses less sensitive to sampling and intraspecific variation (7, 27).
Four biotic provinces were defined based on clusters of species' geographic-range endpoints, which coincide with contacts between contrasting water masses or water types (ref. 19, modifying ref. 28). These natural, quantitatively defined biogeographic units, superimposed on a strong latitudinal diversity gradient, are robust to sampling and provide the spatial template for the analyses of SFDs. At this provincial scale, species richness decreases by a factor of 4, from 624 species in the tropical Panamic Province to 169 species in the Arctic Province (Fig. 1).
Figure 1.

SFDs of marine bivalves in each of the four major northeastern Pacific biogeographic provinces. The SFDs do not differ significantly in mode, median, or range (see Table 1 and Fig. 2).
Results.
Pairwise comparisons among the four provincial faunas show no significant differences in the shapes of the SFDs or in their median sizes, modal size class, and range of sizes (Table 1; Figs. 1 and 2). This decoupling of body-size patterns at the provincial scale from the strong latitudinal trend in species richness is surprising for several reasons: (i) extrinsic factors such as mean annual temperature, oxygen availability, seasonality, and productivity, each of which has been argued to affect body size (refs. 14–17 and 30, but see refs. 31 and 32), vary strongly with latitude along the northeastern Pacific margin (33); (ii) the style and intensity of predation evidently change significantly with latitude (34–36), and size can be an important refuge from predation in marine benthos in general and mollusks in particular (34, 35, 37–41); (iii) although adjacent provinces share species, almost complete taxonomic turnover occurs between the Arctic and Panamic Provinces with only four species ranging throughout; (iv) the family-level composition of the modal size class varies considerably with latitude (Fig. 3), indicating further that these patterns also are not shaped by higher-level phylogenetic effects such as the presence of a few species-rich, cosmopolitan families; and (v) at least some provincial boundaries are size-selective such that SFDs of the species that cross a given boundary differ significantly from those that stop at that boundary. For example, at Point Conception, the boundary between the Californian and Oregonian provinces, the median size of bivalve species that cross the boundary is 30 mm but 17 mm for species that stop at the boundary, and the SFDs of these two sets of species differ significantly (P = 0.0019, Mann–Whitney U test). Together with points (iii) and (iv), this further indicates that the similarities of SFDs among provinces do not simply represent the latitudinal attenuation of a single species-pool.
Table 1.
P values for individual pairwise comparisons of SFDs for the northeastern Pacific bivalves
| Panamic | Californian | Oregonian | Arctic | |
|---|---|---|---|---|
| Kolmogorov–Smirnov test | ||||
| Panamic | — | 0.17 | 0.07 | 0.24 |
| Californian | — | — | 0.77 | 0.4 |
| Oregonian | — | — | — | >.999 |
| Mann–Whitney U test | ||||
| Panamic | — | 0.56 | 0.38 | 0.47 |
| Californian | — | — | 0.28 | 0.32 |
| Oregonian | — | — | — | 0.97 |
A sequential Bonferroni test (29) shows that even the smallest observed P value (0.07) is not significant at a 0.10 tablewide level.
Figure 2.

Median body sizes of northeastern Pacific marine bivalves, grouped at the level of individual provinces (Fig. 1), show no significant differences. The 95% confidence intervals were calculated by randomly resampling the original size data (1,000 iterations each).
Figure 3.

Familial composition of the modal and adjacent size classes (defined as log2 units 4–6) as a function of latitude. The contribution of individual families changes with latitude; e.g., Veneridae and Arcidae are important in the lower latitudes whereas other families, such as Astartidae and Sareptidae, are more important at high latitudes. Thus, the similarities of the provincial SFDs are not the result of phylogenetic effects, such as the presence of a few species-rich and latitudinally widespread families.
Shapes of SFDs
The SFDs of northeastern Pacific bivalves for all four provinces are left-skewed (the g1 statistic defined as the third central moment divided by the cube of the SD of the distributions, from south to north, are −0.51, −0.05, −0.26, −0.31), although only the Panamic province is significantly different from lognormal (P < 0.001). This contrast with the right-skewed SFDs documented for birds and mammals (8, 10, 42) is unlikely to derive from sampling biases. Although new species will continue to be found, particularly in the small size classes, the northeastern Pacific shelf is the best-studied latitudinal transect of molluscan faunas in the world, and remaining differences in sampling intensity among provinces argue against strong biases. The Californian province is the most intensively collected of all, including a rich complement of very small species (21), but its mode and median are indistinguishable from those for provinces to the north and south.
Energetics and Modal Size
Methods.
The latitudinal invariance of the SFDs raises intriguing questions about the factors shaping body-size distributions, given that we can rule out, at least for our molluscan data, the many physical and biotic factors known or hypothesized to vary with latitude. Several kinds of models, ranging from passive diffusion to differential diversification to energetics, have been proposed to account for the shapes of SFDs. Among the available energetic models for body-size evolution (10–12) is one developed by Brown et al. (10) and commonly known as the BMT model, which uses measures of resource acquisition and their allocation to maintenance and reproduction to predict an optimal shape and mode for multispecies SFDs in major taxonomic groups. According to this model, large species are more effective than small ones at acquiring resources but the small species are more efficient in converting those resources into offspring (7, 10). Reproductive power, as the tradeoff between acquisition of resources and their conversion into reproduction, therefore is maximized at an intermediate body size that is predicted to coincide with the modal class in any large assemblage of related taxa. Although some aspects of the BMT model have been controversial (11, 43–46), it provides a theoretical framework for understanding body-size patterns at the level of clades or regional faunas and has successfully predicted modal sizes in birds and mammals (refs. 7, 8, 10, and 42, although see ref. 47 for a failure of the model to predict SFDs of African dung beetles, perhaps owing in part to the focus on a single subclade).
In the first application of the BMT model to marine invertebrates, we
calculated reproductive power for our observed body-size spectrum by
using empirical estimates for the model parameters. According to the
BMT model, reproductive power is given by dW/dt =
(Co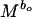 C1
C1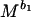 )/(Co
)/(Co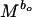 + C1
+ C1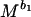 ).
).
bo, which scales the rate of energy acquisition in excess of maintenance needs, can be taken to be 0.75 for all animals, and b1, which scales the rate of transformation of energy to reproductive work, can be taken as −0.25 (10, 42, 43). M is body mass and Co and C1 are taxon-specific constants. We estimated M for individual bivalve species by using the relationship between size and mass given above. There is considerable debate on the estimation of Co and C1 (11, 42–47). We have followed refs. 10 and 42 in calculating these coefficients because the parameters used in those studies are readily available for some species of bivalves. Thus, we assumed that egg production is energetically the most expensive component of the biomass invested in offspring (see ref. 42) for bivalves and used data on mass and gonad production of individual species (e.g., ref. 48) to estimate Co as 0.03 (ref. 10 used milk production in mammals and ref. 42 used egg formation in birds to calculate Co; also see ref. 46). We estimated C1, the conversion component of reproductive power (10, 42), for bivalves as 0.09 from a regression of mass and population production/biomass ratio (e.g., refs. 49 and 50). We also used an alternative estimate of C1 as 0.005 by using a larger sample of “non-insect invertebrates” (appendix VIIIc of ref. 1).
Results.
As shown in Fig. 4, reproductive power is maximized at or near the modal size category for bivalves, for the overall and the provincial SFDs. Further, the shape of the reproductive power curve is consistent with the slightly left-skewed shape of the northeastern Pacific bivalve SFDs. The correspondence in modal size as well as skewness between the size data and the calculated power curve provides impressive corroboration for a model developed for endothermic taxa that have the opposite skew and very different modal values. Coincidence of model output with empirical observations in itself is not strong verification, but the ability of a model to make predictions in new systems can be an important step in the exploration of a model's potential to explain biological phenomena. Thus, although “there is no a priori reason to believe that the most common body size in an animal group is in some sense the best” (46), our analysis empirically supports the BMT model and suggests that energetics play an important role in structuring the spatial distribution of SFDs of northeastern Pacific marine bivalves at the provincial scale. Latitudinally correlated environmental and biotic factors still may be important at smaller spatial scales, of course, such as within-province variations in body size.
Figure 4.

A comparison of the shapes of SFDs predicted by the BMT model (line) and the empirical SFD (histogram) for all of the bivalve species used in this study. The predicted curve in Left is derived by using parameter estimates from Peters (ref. 1; see text) whereas that in Right is based on an alternative estimate of the same parameter (see text). Both predicted curves assume the relationship log2[(length)1/2 × height] = 5.507 + 0.316 × log2(mass). Note that both predicted and empirical distributions are left-skewed and the predicted mode is close to the observed one. Similar results were obtained for individual provinces (not shown).
Evolutionary Trends in Body Size
Methods.
The rich fossil record of marine bivalves allows a direct test of one evolutionary expectation of the BMT model. The modal size has been interpreted as an optimum or adaptive peak that should act as a taxonwide evolutionary attractor, so that lineages should tend statistically to evolve toward, or remain on, the mode over time, albeit with interference from incumbents and other competitors (13, 51, 52). This expectation of directionality contrasts with a stochastic model in which evolutionary patterns in the range of body sizes occupied by clades arise via diffusion during diversification (53–57). To test whether the modal size represents a groupwide evolutionary attractor for bivalves, we first identified the modal size class for 550 species of Miocene northeastern Pacific bivalves drawn from a similar array of families as the Recent fauna. Size data for the Miocene species were collected through an extensive literature search (see refs. 58–60 and references cited therein). To increase sample size and provide geographic coverage from Alaska to the tropics we treated the Miocene as a single time bin, rather than using finer temporal resolution. For each fossil species we used the largest size reported for the Miocene. We then compared, for each of 82 genera having a good Miocene record, the envelope defined by the largest and smallest species in the Miocene with that of the present day (following refs. 24 and 61). Log2-transformed size data for Miocene species yield a median size of 5.28 units, the modal size category of between 5 and 5.5, and a size range from 0.41 to 7.89. Thus, the modal size class for the Miocene species assemblage was the same as the modern one, and the range of sizes was very similar. Therefore, it is appropriate to test the attractor hypothesis by tracking the behavior of individual genera within this framework.
Results.
We found no significant tendency for the genera to evolve toward the modal size over the past 15 million years (i.e., to fall preferentially into the shaded areas in Fig. 5). Numbers are small, but only 7% (95% binomial confidence range of 0–22%) of the genera that started above the mode showed a directional shift toward it (Fig. 5C). Of the genera starting below the mode, 32% (10–54%) showed a directional shift toward the mode, but this is not significantly different from the 21% of lineages (4–41%) that started below the mode and increased their size range in both directions (Fig. 5B). Of the larger sample of genera whose size ranges spanned the mode in the Miocene, only 17% (7–27%) narrow their size range to a tighter focus on the mode, as would be expected if the mode was a selective target. An equal proportion of genera expand both upper and lower bounds, and almost 50% show a directional shift away from the mode, with most of those actually leaving the modal size class completely (Fig. 5D). Thus, our data show that the Miocene–Recent interval was sufficient to encompass considerable evolutionary change in the size range of our 82 lineages and that the mode and range of the SFD of the bivalve fauna was stable despite the failure of the mode to act as an evolutionary attractor. The alternative that the mode is an attractor where entry is generally blocked by established species (7) is also undermined by the paleontological evidence for the highly dynamic behavior of bivalve lineages relative to the modal size.
Figure 5.

Evolutionary size change in 82 Miocene–Recent genera and subgenera of bivalves, with each quadrant representing a different evolutionary pattern as diagrammed in A; the shaded quadrants represent those that would be most heavily occupied if the modal size was an evolutionary attractor. (A) Graphical representation of four potential patterns of body-size evolution, from an earlier t1 to a later t2. As in Jablonski (24), the vertical axis is the change in the upper bound of the size distribution of species in a lineage, and the horizontal axis is the change in the lower bound of the size distribution, so that each clade is plotted as a point determined by the behavior of its upper and lower bounds. (B) Temporal changes in genera whose Miocene maximum size was smaller than the present-day (and Miocene) modal class; the shaded quadrant represents the directional size increase toward the modal class, the expectation if the mode was an evolutionary attractor. (C) Temporal changes in genera whose Miocene minimum size was greater than the modal size class; the shaded quadrant represents directional size decrease toward the modal class. (D) Temporal changes in genera whose size range spanned the modal size class during the Miocene; the shaded quadrant represents a narrowing of size ranges around the mode.
These results are consistent with those from a well-studied Late Cretaceous provincial fauna, where the modal class differs from the Recent fauna by only half a log2 unit despite 65 million years and a major extinction event separating the two intervals, but, again, does not appear to be an evolutionary attractor (61). When the data of Jablonski (61) are analyzed using the protocol of this paper, the modal size class for the Cretaceous bivalves is between 4.5 and 5 units (as opposed to 5–5.5 units for the living species) and median size is 4.48 units. For the Cretaceous bivalve genera starting above these values, 19 ± 11% evolved away from the mode, 30 ± 14% evolved toward the mode, and 35 ± 15% expanded in both directions. The lack of size selectivity in these faunas at the mass extinction boundary suggests that the mode was not an attractor during the recovery phase either, but this requires more detailed investigation.
Paleontological data, therefore, suggest that although the distribution of molluscan body sizes conforms to a framework evidently set by energetic and/or life-history parameters, among lineages, the dynamics of size evolution are more stochastic in nature. The failure of the lineages to exhibit consistent directional shifts toward the mode suggests that the orderly maintenance of regional SFDs over time may involve species sorting (differential origination and extinction within and among size classes; refs. 13, 53, and 62), although the paleontological data are not yet adequate for rigorous testing of this higher-level dynamic in the northeast Pacific.
Conclusions
Studies of latitudinal patterns of biological diversity tend to concentrate on quantifying species richness rather than patterns of morphological and ecological diversity (63). Our results show that for marine bivalves, latitudinal patterns of species richness are decoupled from patterns of body size, a fundamental aspect of species ecology and life history. Regional SFDs are statistically indistinguishable despite significant changes in species richness and in a wide array of variables held to influence body size, such as temperature, seasonality, and productivity. The compositions of provincial faunas presumably are shaped by migration and net diversification rates (speciation less extinction), and in northeastern Pacific bivalves these processes have given rise to remarkably stable patterns of body size over 5,000 km of continental shelf, from the equator to the polar sea. The BMT model (10) successfully predicts both the mode and the shape of these invariant SFDs, but the simplest evolutionary predictions of this and other optimization models, that the mode and presumed optimum will tend to act as an evolutionary attractor, are not met. The density of occupation of a size range, or any other part of the morphospace, need not reflect a long-term microevolutionary optimum, and our paleontological data suggest that the dynamics of size evolution differ across hierarchical levels. These molluscan lineages appear to behave diffusively relative to the mode, as suggested by earlier macroevolutionary models (53–57), but, in contrast to those models, many lineages show a general net size increase, particularly in the upper size bound—regardless of initial position relative to the mode. More importantly, the sorting of lineages appears to operate according to energetic rules and, thus, is nonrandom, so that the stable SFDs evidently are maintained by origination and extinction dynamics within and among size classes (i.e., taxon sorting) rather than strong, directional evolution of lineages toward, or long-term stasis on, the mode. Future work on body-size evolution might focus on the fuller integration of energetic parameters at the level of individuals and populations with the factors governing origination/extinction rates within and among clades.
Acknowledgments
We thank T. Brey for providing the raw data for the tabulation in ref. 48; J. H. Brown, M. Foote, S. M. Kidwell, M. LaBarbera, J. W. Valentine, and an anonymous reviewer for valuable comments; E. V. Coan and P. V. Scott for early access to their volume on northeast Pacific bivalves; and the National Center for Ecological Analysis and Synthesis working group on body size (F. A. Smith, Principal Investigator) for discussions about size evolution. This work was supported by National Science Foundation grants (to K.R. and D.J.), a National Science Foundation Graduate Fellowship (to K.M.), and a John Simon Guggenheim Memorial Fellowship (to D.J.).
Abbreviation
- SFD
size–frequency distribution
References
- 1.Peters R H. The Ecological Implications of Body Size. Cambridge, U.K.: Cambridge Univ. Press; 1983. [Google Scholar]
- 2.Calder W A., III . Size, Function, and Life History. Cambridge, MA: Harvard Univ. Press; 1984. [Google Scholar]
- 3.Schmidt-Nielsen K. Scaling. New York: Cambridge Univ. Press; 1984. [Google Scholar]
- 4.LaBarbera M. In: Patterns and Processes in the History of Life. Raup D M, Jablonski D, editors. Berlin: Springer; 1986. pp. 69–98. [Google Scholar]
- 5.Lawton J H. Philos Trans R Soc London B. 1990;330:283–291. [Google Scholar]
- 6.Blackburn T M, Brown V K, Doube B M, Greenwood J J D, Lawton J H, Stork N E. J Anim Ecol. 1993;62:519–528. [Google Scholar]
- 7.Brown J H. Macroecology. Chicago: Univ. of Chicago Press; 1995. [Google Scholar]
- 8.Maurer B A. Untangling Ecological Complexity. Chicago: Univ. of Chicago Press; 1998. [Google Scholar]
- 9.Hutchinson G E, MacArthur R H. Am Nat. 1959;93:117–125. [Google Scholar]
- 10.Brown J H, Marquet P A, Taper M L. Am Nat. 1993;142:573–584. doi: 10.1086/285558. [DOI] [PubMed] [Google Scholar]
- 11.Kozlowski J. Am Nat. 1996;147:1087–1091. [Google Scholar]
- 12.Kozlowski J, Weiner J. Am Nat. 1997;149:352–380. [Google Scholar]
- 13.Gardezi T, da Silva J. Am Nat. 1999;153:110–123. doi: 10.1086/303150. [DOI] [PubMed] [Google Scholar]
- 14.Atkinson D. Adv Ecol Res. 1994;25:1–58. [Google Scholar]
- 15.Blackburn T M, Gaston K J, Loder N. Diversity Distributions. 1999;5:165–174. [Google Scholar]
- 16.Cushman J H, Lawton J H, Manly B J F. Oecologia. 1993;95:30–37. doi: 10.1007/BF00649503. [DOI] [PubMed] [Google Scholar]
- 17.Turner J R G, Lennon J J. Nature (London) 1989;340:351. [Google Scholar]
- 18.Hawkins B A, Lawton J H. Oecologia. 1995;102:31–36. doi: 10.1007/BF00333307. [DOI] [PubMed] [Google Scholar]
- 19.Roy K, Jablonski D, Valentine J W. Proc Natl Acad Sci USA. 1994;91:8871–8874. doi: 10.1073/pnas.91.19.8871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Roy K, Jablonski D, Valentine J W, Rosenberg G. Proc Natl Acad Sci USA. 1998;95:3699–3702. doi: 10.1073/pnas.95.7.3699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Coan E V, Scott P V, Bernard F R. Santa Barbara Museum of Natural History Monographs, Studies in Biodiversity. 2000;2:1–764. [Google Scholar]
- 22.Stanley S M. Paleobiology. 1986;12:89–110. [Google Scholar]
- 23.Jackson J B C, Jung P, Fortunato H. In: Evolution and Environment in Tropical America. Jackson J B C, Budd A F, Coates A G, editors. Chicago: Univ. of Chicago Press; 1996. pp. 234–270. [Google Scholar]
- 24.Jablonski D. Nature (London) 1997;385:250–252. [Google Scholar]
- 25.Powell E N, Stanton R J., Jr Palaeontology. 1985;28:1–34. [Google Scholar]
- 26.Tokeshi M, Ota N, Kawai T. J Zool London. 2000;251:31–38. [Google Scholar]
- 27.Brown J H, Nicoletto P F. Am Nat. 1991;138:1478–1512. [Google Scholar]
- 28.Valentine J W. Limnol Oceanogr. 1966;11:198–211. [Google Scholar]
- 29.Rice W R. Evolution. 1989;43:223–225. doi: 10.1111/j.1558-5646.1989.tb04220.x. [DOI] [PubMed] [Google Scholar]
- 30.Chapelle G, Peck L S. Nature (London) 1999;399:114–115. [Google Scholar]
- 31.Spicer J I, Gaston K J. Ecol Lett. 1999;2:397–401. [Google Scholar]
- 32.Poulin R. Biol J Linn Soc. 1995;54:231–244. [Google Scholar]
- 33.Robinson A R, Brink K H, editors. The Sea—Vol. 11: The Global Coastal Ocean. New York: Wiley; 1998. [Google Scholar]
- 34.Vermeij G J. Biogeography and Adaptation. Cambridge, MA: Harvard Univ. Press; 1978. [Google Scholar]
- 35.Vermeij G J. Evolution and Escalation. Princeton: Princeton Univ. Press; 1987. [Google Scholar]
- 36.Stachowicz J J, Hay M E. Am Nat. 2000;156:59–71. doi: 10.1086/303366. [DOI] [PubMed] [Google Scholar]
- 37.Paine R T. Ecology. 1976;57:858–873. [Google Scholar]
- 38.Palmer A R. Evolution. 1979;33:697–713. doi: 10.1111/j.1558-5646.1979.tb04722.x. [DOI] [PubMed] [Google Scholar]
- 39.Boulding E G. J Exp Mar Biol Ecol. 1984;76:201–223. [Google Scholar]
- 40.Juanes F. Mar Ecol Prog Ser. 1992;87:239–249. [Google Scholar]
- 41.Arsenault D J, Himmelman J H. Mar Ecol Prog Ser. 1996;140:115–122. [Google Scholar]
- 42.Maurer B A. Evol Ecol. 1998;12:935–944. [Google Scholar]
- 43.Brown J H, Taper M L, Marquet P A. Am Nat. 1996;147:1092–1097. [Google Scholar]
- 44.Jones K E, Purvis A. Funct Ecol. 1997;11:751–756. [Google Scholar]
- 45.Symonds M R E. Funct Ecol. 1999;13:508–513. [Google Scholar]
- 46.Chown S L, Gaston K J. Funct Ecol. 1997;11:365–375. [Google Scholar]
- 47.Chown S L, Steenkamp H E. Afr Entomol. 1996;4:203–212. [Google Scholar]
- 48.Brey T. Mar Ecol Prog Ser. 1995;125:87–93. [Google Scholar]
- 49.Urban H J, Campos B. Mar Ecol Prog Ser. 1994;115:93–102. [Google Scholar]
- 50.Urban H J. J Shellfish Res. 1996;15:719–727. [Google Scholar]
- 51.Kelt D A. Biol J Linn Soc. 1997;62:15–37. [Google Scholar]
- 52.Alroy J. Science. 1998;280:731–734. doi: 10.1126/science.280.5364.731. [DOI] [PubMed] [Google Scholar]
- 53.Maurer B A, Brown J H, Rusler R D. Evolution. 1992;46:939–953. doi: 10.1111/j.1558-5646.1992.tb00611.x. [DOI] [PubMed] [Google Scholar]
- 54.Stanley S M. Evolution. 1973;27:1–26. doi: 10.1111/j.1558-5646.1973.tb05912.x. [DOI] [PubMed] [Google Scholar]
- 55.McKinney M L. In: Evolutionary Trends. McNamara K J, editor. Tucson: Univ. of Arizona Press; 1990. pp. 75–118. [Google Scholar]
- 56.McShea D W. Evolution. 1994;48:1747–1763. doi: 10.1111/j.1558-5646.1994.tb02211.x. [DOI] [PubMed] [Google Scholar]
- 57.Gould S J. J Paleontol. 1988;62:319–329. [Google Scholar]
- 58.Marincovich L. Bull Am Paleontol. 1983;84:59–155. [Google Scholar]
- 59.Moore E J. U S Geol Surv Prof Paper. 1983–1992;1228:A–E. [Google Scholar]
- 60.Woodring W P. U S Geol Surv Prof Paper. 1973–1982;306:E–F. [Google Scholar]
- 61.Jablonski D. In: Evolutionary Paleobiology. Jablonski D, Erwin D H, Lipps J H, editors. Chicago: Univ. of Chicago Press; 1996. pp. 256–289. [Google Scholar]
- 62.Grantham T A. Annu Rev Ecol Syst. 1995;26:301–321. [Google Scholar]
- 63.Roy K, Foote M. Trends Ecol Evol. 1997;12:277–281. doi: 10.1016/s0169-5347(97)81026-9. [DOI] [PubMed] [Google Scholar]