

Abstract
Background
The var2csa gene encodes a Plasmodium falciparum adhesion receptor which binds chondroitin sulfate A (CSA). This var gene is more conserved than other PfEMP1/var genes and is found in all P. falciparum isolates. In isolates 3D7, FCR3/It4 and HB3, var2csa is transcribed from a sub-telomeric position on the left arm of chromosome 12, but it is not known if this location is conserved in all parasites. Genome sequencing indicates that the var2csa gene is duplicated in HB3, but whether this is true in natural populations is uncertain.
Methodology/Principal Findings
To assess global variation in the VAR2CSA protein, sequence variation in the DBL2X region of var2csa genes in 54 P.falciparum samples was analyzed. Chromosome mapping of var2csa loci was carried out and a quantitative PCR assay was developed to estimate the number of var2csa genes in P.falciparum isolates from the placenta of pregnant women and from the peripheral circulation of other malaria patients. Sequence analysis, gene mapping and copy number quantitation in P.falciparum isolates indicate that there are at least two loci and that both var2csa-like genes can be transcribed. All VAR2CSA DBL2X domains fall into one of two distinct phylogenetic groups possessing one or the other variant of a large (∼26 amino acid) dimorphic motif, but whether either motif variant is linked to a specific locus is not known.
Conclusions/Significance
Two or more related but distinct var2csa-type PfEMP1/var genes exist in many P. falciparum isolates. One gene is on chromosome 12 but additional var2csa-type genes are on different chromosomes in different isolates. Multiplicity of var2csa genes appears more common in infected placentae than in samples from non-pregnant donors indicating a possible advantage of this genotype in pregnancy associated malaria.
Introduction
The var2csa gene is the best characterized of the PfEMP1/var genes and the protein it encodes is a parasite receptor for binding to human placental CSA and thought to play an important role in the pathogenesis of pregnancy associated malaria (PAM) [1]–[3]. VAR2CSA proteins are large (∼350 kDa), antigens, exposed to host antibodies on the erythrocyte surface membrane [4], [5]. They are more variable than most viral antigens known to bind endothelial receptors, but unusually conserved compared to other members of the extremely diverse PfEMP1 antigen family. The var2csa genes are relatively distantly related to other var genes and constitute a distinct group whose recombination with other var genes is suppressed [6], [7]. The var2csa gene is also one of only two var genes (the other being var1csa) which are found in all three of the original P.falciparum reference genome sequences [7]. The var2csa gene is the only PfEMP1/var gene sufficiently conserved to have a recognizable homologue in P.reichenowi, the chimpanzee Plasmodium species that is P. falciparum's closest evolutionary relative [8].
Because it is essential to adhesion of malaria parasites to the placenta [9], VAR2CSA has been proposed as the antigen in an adhesion-blocking vaccine to protect women against malaria during pregnancy [10]–[12]. There is therefore interest in defining the domains involved in adhesion to CSA, such as the DBL2X region [13]. To define variability in the VAR2CSA DBL2X domains we analyzed these sequences in var2csa genes in P.falciparum samples taken from placentae at delivery and from the peripheral circulation of malaria patients, combining these with sequences from database sources.
DBL2X polymorphism consists of defined blocks of variability, divided by regions of sequence conservation, a pattern of variation generally observed in PfEMP1 protein domains [14]–[16]. However the sequence of one substantial region of the VAR2CSA DBL2X domain exists in two dimorphic types. One explanation for this striking dimorphic sequence motif (DSM) might be that there is more than one var2csa-type gene per haploid genome, a tentative conclusion also reached based on the var gene sequence assembly from the HB3 isolate P.falciparum genome sequencing project [7]. This ‘multiple var2csa genes’ hypothesis was therefore tested by mapping the position of var2csa-type genes in pulsed field gel chromosome separations of different parasite isolates. Following confirmation of the existence of two separate loci, a quantitative real-time PCR assay, based on the 2−ΔΔ Ct method, was developed to count the numbers of var2csa-type genes in P.falciparum clinical isolates.
Materials and Methods
Origin and maintenance of P.falciparum isolates
Six placental field-isolates (Tz745, Tz748, Tz752, Tz755, Tz788 and Tz796) and five long-term cultured clones (HB3, FCR3/It4, DD2, 7G8 and 3D7) were grown. Modified Trager-Jensen medium consisting of blood group O+ red blood cells (5% haematocrit), supplemented with 25 mM sodium bicarbonate, 0.125 g L−1 gentamycin and 0.125 g L−1 Albumax II was used. Flasks were kept at 37°C and gassed with 2% oxygen, 5% carbon dioxide in nitrogen. Parasites were harvested at around 5% parasitaemia. Cultures were tested for isolate integrity using nested GLURP and MSP-2–specific primers to measure clone multiplicity by PCR. DBL2X sequences were from placental blood samples collected at delivery in Guediawaye maternity ward, Senegal [17] and stored on filter paper, or extracted from clinical samples collected in Daraweesh, Sudan [18] or Korogwe, Tanzania [19]. Other sequences are from PlasmoDB.
PCR amplification and sequencing of P.falciparum genomic DNA
Genomic DNA was extracted from placental blood samples on filter paper using the Chelex method [20] or extracted directly from patient blood. The var2csa gene was amplified using various combinations of domain specific primers (Table 1). Multi-sequence alignments were made using MAFFT software (http://align.bmr.kyushu-u.ac.jp/mafft/software/) with the “G-INS-I” setting for global alignments, corrected manually.
Table 1. PCR amplification primers.
| Gene: | Primer ID/application: | Primer sequence: | Tm: |
| var2csa- | 557 (Sequencing) | 5′ CGGAATTCAAATGCGACAAATGTAAAT | 54 |
| (DBL2X) | 558 | 3′ ATTTGCGGCCGCCTGATTGTACACATTTATT | 60 |
| 569 | 5′ CGGAATTCGGATCTAGTTCTAATGGTAGTTGT | 61 | |
| 570 | 3′ ATTTGCGGTCGCAATGTTTGAAAAACGAATC | 59 | |
| 653 | 5′ CGGAATTCCTTCAAGAAAATTGTAGTG | 55 | |
| 654 | 3′ ATTTGCGGCCGCGTGTGGTCAATCCCTATAT | 64 | |
| var2csa- | A763 (Sequencing) | 5′ GAAATTGACAATGCAATAA | 56 |
| (DBL5e) | A764 | 3′ CTTCAAGTTCAGCTGGAAT | 54 |
| Tz748-var2csa- | 748A (Transcript quant. assay) | 5′ ATATTAAAAGATGTAAAGGAACCG | 49 |
| (DBL5e) | 748A | 3′ TTTCTTTTTCGTTGTCTTCATTG | 48 |
| 748B | 5′ AATGTATTTAACAATGCAAATGA | 47 | |
| 748B | 3′ CTTCATTTCCGATGTTTGTATAT | 48 | |
| Tz745-var2csa- | 745A (Transcript quant. assay) | 5′ AACAAAACTTGGAGGCAAATG | 49 |
| (DBL5ε) | 745A | 3′ GAGATCCAGCAGTACCAC | 50 |
| 745B | 5′ AGG AAT GTG GAA ACA AAT GTA | 47 | |
| 745B | 3′ GGATCCGGCAGTACCAGT | 53 | |
| seryl-tRNA synthetase | P90 (Transcript quant. Assay) | 5′ AAGTAGCAGGTCATCGTGGTT | 52 |
| (PF07_0073) | P90 | 3′ TTCGGCACATTCTTCCATAA | 48 |
| var2csa- | T12 (PCR copy number assay) | 5′ AATGGGACAAACAAAAAACAAAATAT | 51 |
| (DBL3x) | T13 | 3′ GCTGATATACATTCAGGATAATTTTC | 52 |
| aldolase | P61 (PCR copy number assay) | 5′ TGT ACC ACC AGC CTT ACC AG | 54 |
| (PF14_0425) | P61 | 3′ TTCCTTGCCATGTGTTCAAT | 48 |
Phylogenetic analysis of the domain variants
Phylogenetic analyses were conducted using MEGA version 4.0.2. Input data were sequence alignments of VAR2CSA DBL2X and DBL5ε amino acid sequences. Phylogenetic trees were constructed both as unrooted neighbor joining (NJ) and maximum parsimony trees using a Dayhoff matrix substitution model. Inferred phylogeny was validated by bootstrapping (1,000 replications).
Pulsed field gel electrophoresis and Southern blotting
P.falciparum chromosomal DNA blocks were prepared as described [21]. Pulsed field gel electrophoresis was carried out using a Bio-Rad™ CHEF DRII apparatus (0.5X TBE buffer, circulated at 14°C). Chromosomes were separated on 0.7% agarose gels, initially ramping the pulse interval from 90–300s for 24 hours at 4.2 V/cm, followed by 300–720 s for 43 hours at 3.0 V/cm. For Southern blotting, chromosomes were hydrolyzed in the gel (0.25N HCl for 30 mins), prior to denaturation, renaturation and 10X SSC transfer onto Hybond N+ nylon membranes (Amersham GE Healthcare). The DBL3X probe was labeled with dioxygenin-dUTP by PCR using primers forward (pos.4498–4525) and reverse (pos. 4613–4642), based on the 3D7 var2csa sequence [22]. Membranes were pre-hybridized overnight at 42°C in 5X SSC, 5X Denhardt's, 0.5% SDS, 20 mM Tris-HCl pH 7.5, 10% Dextran Sulphate, 35% formamide. Hybridization was carried out in the same buffer with dioxygenin (DIG) (Roche) labeled probe, at 42°C overnight. The blot was washed in SSC at 60°C, with increasing stringency washes, ending with 0.1 X SSC, 0.1% SDS, at room temperature.
Quantitative Real-Time PCR (Q-RT PCR)
Q-RT-PCR was performed using the Rotorgene 6000, version 1.7 system (Corbett Research). Reactions were prepared in 20 µL using Quantitec SYBER Green PCR master mix (Qiagen) and primer concentrations of 1 µM. PCR cycling was 95°C for 15 mins, followed by 40 cycles of 95°C for 30 s, 54°C for 20 s and 65°C for 40 s, with final extension at 68°C for 40 s. The cycle threshold, the PCR cycle at which the product-derived fluorescence intensity crosses a threshold value, was set at 0.025, which reflected the optimal PCR parameters across all runs.
Selection and validation of primers for 2−ΔΔ Ct analysis
The var2csa and aldolase primers (see Table 1) were selected to target known conserved sequences [23]. Primers were optimised to avoid self annealing and hairpin loops formation. Oligonucleotide lengths and melting temperatures were equalized as much as possible and specificity of amplification measured by melting-curve analysis. Primers were BLAST searched against P.falciparum and human databases to ensure specificity. All PCR reactions were checked for the amplification of a single band of the expected size.
The 2−ΔΔ Ct method of relative gene copy number estimation
The 2−ΔΔ Ct method of relative quantitation [24] was adapted to estimate copy number of the var2csa gene. For this calculation to be valid, the amplification efficiency (E) must be close to 100% and E values for target and reference genes (the relative efficiency) must be approximately equal. The measurement of copy number of a target gene in an unknown sample requires a ‘calibrator’ genome with known copy number of both target and reference genes. FCR3/It4 was used as the calibrating parasite genome and the var2csa copy number was determined relative to FCR3/It4, using equations (1) 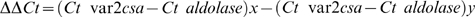 , where x = unknown sample and y = P.falciparum FCR3/It4, and (2) in which the copy number of the unknown target gene is expressed as 2−ΔΔ Ct.
, where x = unknown sample and y = P.falciparum FCR3/It4, and (2) in which the copy number of the unknown target gene is expressed as 2−ΔΔ Ct.
The assay is particularly sensitive to the quality of template DNA preparations. The intrinsic variation between different DNA samples was assessed by making ten separate extractions of DNA, both from filter paper and infected blood, of the FCR3/It4 calibrator, and measuring their Ct values, in triplicate, in three separate runs. The total error, including variation between triplicates, DNA preparations and runs is then included in subsequent calculations of target gene copy numbers, by computing the upper and lower limits of the 95% confidence intervals of both the unknown sample and calibrator. Upper and lower 95% limits of the ΔΔ Ct value were then used to give a more accurate 95% confidence limit on the copy number estimate, given as [2∧−ΔΔCt High− 2∧−ΔΔCt Low]. Three conservative sample inclusion criteria were used. Therefore samples were excluded if the Ct value was >26, if triplicates had a CV >1% and if samples had a 95% confidence copy number interval between integer values 1 and 2.
Transcript quantitation assay
To test whether either or both of the var2csa-type genes were expressed as mRNA in two locus isolates, variant specific primer to amplify reverse transcribed cDNA were designed (Table 1). To compare the transcription of each gene, the amplification efficiency of the two primer pairs were first determined by real-time measurements of serial 10 fold dilutions of plasmid DNA carrying inserts of the two var2csa-type genes (Tz745 and Tz748). Quantitative measurements were then made by comparing the Ct values of the two var2csa-type genes in each isolate with that of the endogenous housekeeping gene (seryl-tRNA synthetase, PF07_0073).
Results
Variation and conservation in var2csa-type genes and proteins
Figure 1 shows a diagram of the VAR2CSA gene and protein. Salanti et al [25] noted that this gene differs from the other var genes in that some of its domains do not conform to the sequence motif spacing-based classification schemes for CIDR and DBL domains used in other PfEMP1 proteins [26]. An estimate of VAR2CSA domain conservation between parasite isolates, calculated on the basis of amino acid sequences, is given by the percentage sequence identity figure shown below each domain. These similarity estimates are based on nine complete var2csa genes assembled in the PlasmoDB database [27] and the P.falciparum genome sequencing projects at the Sanger and Broad Institutes. Most PfEMP1/var genes have <50% amino acid sequence identity between individual DBL domains [7], but significantly higher levels of conservation between isolates are seen in all VAR2CSA domains. The most conserved regions are the central DBL3X and DBL4ε domains. The N and C-terminal regions are somewhat more variable, the membrane-proximal DBL6ε being the most polymorphic domain. Domain functions remain unclear although DBL5ε binds IgM [28]–[30] but whether binding to CSA is mediated by a single or multiple domains is controversial [31]–[36]. Host antibodies bind antigenic structures throughout the molecule [37], [38] and antibodies specific for DBL4ε have been shown to inhibit ligand binding [39].
Figure 1. Structure of the var2csa gene and protein.

The gene and its exon structures are shown above a protein diagram which has been subdivided into predicted domains [71]. The approximate amount of variation (expressed as percentage amino acid identity) in these domains is shown below each domain block. The sequence similarities were calculated based on the genome sequencing data available (from 3D7, FCR3/It4, HB3, Dd2, PfClin, and P.reichenowi). The trans-membrane region and conserved (>95%) intracellular acidic terminal sequence (ATS) are not included in the analysis.
Variation in the DBL2X domain of VAR2CSA
As part of a study of variation in VAR2CSA, polymorphism in the VAR2CSA DBL2X domain was analyzed. The alignment of the DBL2X domain shows the pattern of interspersed conserved and variable sequences previously seen in the VAR2CSA DBL3X domain [40]. The full alignment of 55 different ∼340 amino acid DBL2X sequences is shown in Figure S1. However it is striking that a relatively large region of the DBL2X domain has one or the other of two variants of a conserved sequence. This clearly differentiates these sequences into two distinct groups. Figure 2 details a ∼62 a.a comparison between 55 VAR2CSA DBL2X domain sequences highlights this dimorphic sequence motif (DSM). Color blocks show regions of >55% sequence identity. Interestingly, the P.reichenowi VAR2CSA DBL2X has conserved this motif and has the FCR3/It4-type variant of the dimorphism. The two closely related sequences derived from the HB3 genome sequencing project are labeled HB3.A and HB3.B. Both are of the FCR3/It4 DSM sub-type.
Figure 2. Dimorphic sequence motif in the DBL 2X domain of VAR2CSA.

The multiple sequence alignment covers a region of 60–62 amino acids and includes 37 VAR2CSA DBL2X sequences from Senegalese (SG) placental isolates (accession numbers GQ358101 – GQ358135) and 18 database-derived sequences, including P.reichenowi (Reich). This region includes amino acids 580–641 of the VAR2CSA sequence encoded by the 3D7 PFL0030c gene. Color blocks mark positions of the alignment with >55% amino acid sequence identity. The dimorphic sequence motif (DSM) is shown between positions 50–78. All sequences contain one or the other DSM variant. A solid line separates FCR3/It4-type sequences from 3D7 type sequences.
Structural models of the VAR2CSA DBL 2X domain
Structural models have been made of the DBL2X and DBL3X domains [41], [42], based on similarity to the DBL domain of the P.falciparum EBA 175 antigen [43]. Recent crystal structures of VAR2CSA DBL3X [44], [45] indicate that the lower-resolution, homology-based model nonetheless agrees well with the experimentally determined structure. Figure 3 illustrates a 3-D model of the DBL2X domain (which has not yet been analyzed by crystallography), similar to that recently published by Bockhorst et al [46], highlighting the position of the DSM which covers an exposed loop and part of an α-helical region of this domain.
Figure 3. Three-dimensional model of the VAR2CSA DBL2X domain.

The three dimensional model of the DBL2X domain displays the position of the dimorphic sequence motif (DSM). The model was constructed using the F1 domain of the P.falciparum Erythrocyte Binding Antigen (EBA 175, Protein data bank code 1ZRO) as a structure template. The VAR2CSA amino acid sequence used is the 3D7 genome sequence (PFL0030c amino acids 535–878).
Phylogenetic relationships between VAR2CSA DBL2X domains
The dimorphism of DBL2X domains was investigated by a standard type of phylogenetic analysis based on the assumption that sequences are evolving largely by mutation. This has been cogently criticized as underestimating the role of very high levels of recombination in generating hyper-variable sequence blocks in P.falciparum PfEMP1/var genes [47]. However, by the standards of PfEMP1/var genes, the VAR2CSA DBL2X domain is not a hypervariable region and in fact shows some highly conserved features, as discussed below. Figure 4 shows the constructed neighbor-joining trees, based on 54 P.falciparum and one P.reichenowi var2csa DBL2X sequences. Figure 4A confirms that these DBL2X sequences will cluster into two distinct phylogenetic subgroups (bootstrap value 97). Sequences fall into one of two reference groups, the 3D7 DSM type, shown in green, or the FCR3/It4 DSM type, shown in red. To test whether this is due to the DSM, or other less obvious variation, the sequences were re-tested, after excising the DSM sequence from the alignments. The neighbor joining tree shows that the division of the sequences into two groups disappears and the branching of the tree is similar to the relationship observed between variants of other DBL domains [48]. Unsurprisingly, the branch lengths support a more distant relationship between the P.reichenowi sequence and all P.falciparum sequences but the conservation of the DSM still results in this particular P.reichenowi sequence clustering with FCR3/It4 sub-type sequences.
Figure 4. Phylogenetic relationships between 44 VAR2CSA DBL2X sequences.

A. Neighbor joining tree illustrating the phylogenetic relationship between the sequences, with representatives of each of the two DSM variants marked in separate colors. 3D7-type sequences are shown in red and FCR3/It4-type in green. The bootstrap value is shown at the main bifurcation. B. Neighbor joining tree based on the same sequences, after excision of the DSM region. The 3D7/red, FCR3/It4/green variant-type coding is maintained.
The majority of the sequences in this analysis are from Dakar, Senegal. However, Figure 4 shows that the branching pattern also accommodates and separates the other African isolates used (e.g. Ghana A and B and SD2O2 and SD124.8 from Sudan), the database sequences (DD2, MC, FCR3/It4, HB3 and 3D7) and the P. reichenowi isolate. These isolates originate from other areas of Africa, Central America and S.E. Asia and indicate that a dimorphic sequence influenced phylogeny is a general feature of the DBL2X domain of var2csa-type genes.
Mapping the var2csa gene onto pulsed field gel separated chromosomes
Alternative hypotheses could explain the existence of the two VAR2CSA DBL2X domain types. Either a single var2csa gene locus encodes a dimorphic series of allelic variants, or multiple loci encode related var2csa-type genes, each having a particular variant of the DSM. The var2csa gene has been mapped to the sub-telomeric region of the left arm of chromosome 12 in isolates FCR3/It4, 3D7 and HB3 but is not known if this location is universal. Genome sequencing projects have detected, but not mapped, two var2csa-type genes in the HB3 isolate [7]. To resolve these uncertainties, a conserved portion of the var2csa gene was used to map the location of the var2csa locus in Southern blots of pulsed field gel (PFG) separated P.falciparum chromosomes.
Figure 5 illustrates a separation of the 14 P.falciparum chromosomes and the corresponding Southern blot, probed with a DBL3X-derived var2csa-specific gene probe. To compensate for gel distortions each sample was run in duplicate. The PFG illustrates the chromosome length polymorphisms that are a feature of P.falciparum [49]. Confirming earlier experiments, chromosome 12 contains a var2csa gene locus, hybridized by the var2csa gene probe in all 10 isolates shown in Figure 5. Two of the isolates, HB3 and Tz748, have one additional locus hybridizing the var2csa probe, on chromosome 1 in HB3 and on what is probably chromosome 8 in Tz748 (hybridizing to the upper region of the poorly resolved chromosomes 5–9, Fig. 5A). The Tz745 has a distinct chromosome 12 var2csa locus and at least one and quite possibly two additional hybridizing loci in the poorly resolved chromosome 5–9 region (Fig. 5B).
Figure 5. Chromosomal location of var2csa-type loci in P.falciparum isolates.

Conditions for the separation of the chromosomes and DIG labeling and detection of the var2csa DBL3X domain-derived probe sequence are given in the Materials and Methods. Gel measuring rulers and superimposed alignment lines are shown. Marker chromosomes are from Hansenula wingei. Note that the two uppermost faint bands are not chromosomes but a PFG artifact. HB3, 3D7, FCR3/It4 and 7G8 are long term laboratory isolates whilst Tz745, Tz748, Tz752, Tz755, Tz788 and Tz796 are recent field isolates from Korogwe, Tanzania.
Gene mapping thus shows that both long-term laboratory isolate HB3 and recent field isolates such as Tz745 and Tz748 have additional loci containing a second var2csa-type gene. We think it likely that isolates containing three var2csa-type genes also exist (Fig. 5B, see Tz745). Since the presence of more than one clone in the culture can be easily detected by PFG analysis, these results also indicate that detection of two var2csa-type genes in these isolates is not caused by the presence of multiple clones in the original isolate.
Genome sequences exist for four established laboratory clones of P.falciparum (3D7, FCR37It4, HB3 and Dd2), for one clinical isolate of Ghanaian origin, and for P.reichenowi [50]–[52]. In HB3, two complete var2csa genes have been assembled (http://www.broad.mit.edu). This agrees with our gene mapping data for HB3 although at this stage of annotation, the two var2csa loci have been placed in head to head orientation on chromosome 12. Our data indicate that the second HB3 var2csa gene is in fact on chromosome 1. Hybridization signals should be proportional to target copy number and the relative intensities observed do not indicate the presence of multiple var2csa-type genes on chromosome 12 in HB3.
PCR assay estimating var2csa gene multiplicity in P.falciparum isolates
PFG-based gene mapping is an unambiguous but unwieldy tool for estimating the number of var2csa-type genes in large numbers of clinical isolates because these are difficult to establish and maintain in culture. It is therefore desirable to have an assay that can be used to measure gene copy number in DNA isolated from preserved blood. The 2−ΔΔ Ct assay does this by normalizing the amount of a variable-copy target gene, against an endogenous single copy gene. After calibration, target gene number in samples can be expressed as n-fold changes in target gene copies, relative to the copy number of the target gene in an endogenous ‘spike’ (known to be 1). Based on our gene mapping and the genome project analysis, P.falciparum FCR3/It4 has the requisite single copy var2csa gene.
Figure 6 shows a 2−ΔΔ Ct assay graph of DNA quantity dependent fluorescence (Rn) versus PCR cycle number. Amplification of the var2csa gene crosses the pre-determined cycle threshold at Ct = 24.0 for P.falciparum HB3. This occurs at Ct = 24.7 for P.falciparum FCR3/It4. The corresponding values for the aldolase gene are Ct 24.3 and 24.0 respectively. Using the ΔΔ Ct equations;
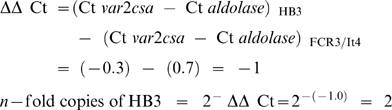 |
Figure 6. 2−ΔΔ Ct assay for counting var2csa-type gene copy number.

Cycling and assay conditions are detailed in the Materials and Methods section. The cycle thresholds (Ct) for detection of var2csa-type target genes and aldolase reference genes are shown as the points where the signal crosses an experimentally predetermined threshold value, in a graph of Rn (normalized reporter by fluorescence) against the number of PCR cycles.
For clone HB3, the 2−ΔΔ Ct assay estimated var2csa copy number agrees with the gene mapping and the genome assembly. With other isolates known to have two loci (Tz748 and Tz745), the assay agrees with the PFG gene mapping. Calibrated with a reference genome ‘spike’, and including primers for a single copy housekeeping gene for normalization, the 2− ΔΔ Ct assay thus appears valid as a gene counting method for var2csa genes.
The polyclonal nature of many P.falciparum infections [53] complicates gene counting and makes sequencing an unreliable method for estimating gene copy numbers. The PCR assay however can not over-estimate the number of isolates containing clones with multiple var2csa-type genes, since a mixture of single and multi-copy isolates can only bring the copy number estimate closer to one. Nevertheless, polyclonality precludes the calculation of actual target gene numbers in mixed genotype infections. Instead, the 2-ΔΔ Ct allows estimation of the proportion of isolates containing parasites which have multiple var2csa-type genes/haploid genome.
We have analyzed a sample set consisting of 43 blood samples from children in Korogwe, Tanzania, 36 placental blood samples from Korogwe and 32 peripheral blood samples from non-pregnant malaria cases in Daraweesh, Sudan. The 2−ΔΔ Ct assay results, showing the proportion of samples containing parasites with multiple var2csa-type genes are presented in Table 2. Parasites derived from the placentae were more likely to contain clones which have multiple var2csa-type genes, although this inequality was not statistically significant. The existence of an advantage of the var2csa multi-locus genotype in the malaria in pregnancy syndrome is therefore neither established nor excluded.
Table 2. var2csa gene multiplicity in samples from placentae at delivery and from non-pregnant P.falciparum donor samples.
| Sample origin | Number of samples | Placental/Non-pregnant samples | Proportion of samples with multiple var2csa-type gene per genome |
| Korogwe, Tanzania | 36 | Placental | 31%, (11/36) |
| Korogwe, Tanzania | 43 | Non-pregnant | 14%, (6/43) |
| Daraweesh, Sudan | 32 | Non-pregnant | 15%, (5/32) |
| Total | 111 | all | 20%, (22/111) |
The relationship between var2csa-type genes located in the same genome
To discover if the multiple var2csa-type genes found in some P.falciparum genomes represent recently duplicated paralogs, we sought to analyze whether genes present in the same genome are more closely related to each other than they are to other var2csa-type genes. Such an analysis requires pre-identification of monoclonal isolates with multiple var2csa loci. Assessing the degree of relationship between var2csa-type genes also requires analyzing domain of the protein that show no evidence of dimorphism, since isolates with two loci, each with the same dimorphic region (e.g. HB3) are less easily recognized as distinct loci. DBL5ε domain sequences were therefore used for this analysis of the relationship between var2csa-type genes in the same genome. After identifying eight monoclonal samples with multiple var2csa-type genes, their DBL5ε coding regions were amplified, sub-cloned and sequenced. These were compared with 25 var2csa DBL5ε-type sequences to estimate the relatedness of var2csa-type genes known to exist within the same genome, as compared to other var2csa-type genes. None of the var2csa-type sequences from multi-copy genomes had any unique polymorphisms absent from other DBL5ε sequences (data not shown).
The phylogenetic relationship between DBL5ε sequences derived from the same genome and an essentially random sample of other DBL5ε sequences is shown in Figure 7. The tree does not support the hypothesis that multiple var2csa–type genes are the result of recent duplication events in most isolates, although the HB3 and 1627 sequences are quite closely related and could represent relatively more recent duplications. In at least some of the genomes analyzed, the two genes appear to have been evolving independently for sufficient time to have diverged considerably.
Figure 7. Phylogenetic relationships between VAR2CSA DBL5ε sequences.

A neighbor joining tree showing the relationships between 33 different VAR2CSA DBL5ε sequences, including eight sequences derived from samples which are known to have two or more var2csa-type genes/haploid genome. Sequences derived from the same genome are presented in the same color. Bootstrap values for each clade are shown at the nodes of the tree.
Are both var2csa genes capable of being transcribed?
To test whether more than one of the var2csa-type genes are being transcribed into mRNA in multi-loci isolates such as Tz745 and Tz748, var2csa variant-specific primers were designed to amplify cDNA from these cultures. Both parasite isolates are monoclonal, as judged by PCR genotyping and pulsed field gel electrophoresis and both bind CSA on BeWo syncytiotrophoblastic cells. BeWo adhesion-selected Tz745 and Tz748 also show the characteristic parity-dependent serological recognition phenomenon [54]. The amplification efficiency of variant specific primers was determined. cDNA of the var2csa-type genes from both isolates was then amplified using each primer pair, and assayed relative to the amplification of seryl-tRNA synthetase. The results showed that in both adhesion-selected cultures, there is specific transcription from each of two var2csa-type genes (Ct 745.A = 16.1; Ct 745.B = 12.3), (Ct 748.A = 17.1; Ct 748.B = 16.1) and the transcript levels of both genes are increased relative to those measured for the housekeeping gene (Ct 745 = 18.0; Ct 748 = 17.7), after adhesion selection. Each gene thus appears to be transcription-competent and, in these isolates is neither is a pseudo gene.
Discussion
Sequencing projects have been reported for six P.falciparum genomes, each complete and annotated to a different degree. There is generally incomplete coverage of the repeat-rich telomeric sequences (where many var genes lie) and annotation of the multi-gene families such as PfEMP1 and the RIFINS has proven difficult and time-consuming. The first report that the HB3 genome sequencing project detected two var2csa-type genes indicated that these genes are very similar and closely linked on chromosome 12 and thus potentially recent duplications. The biological relevance of such events is uncertain although a recent study using monoclonal antibodies to select VAR2CSA antigen-expressing parasitized erythrocytes concluded that both var2csa-type genes were expressed and up-regulated following this selection [55].
The gene mapping studies shown here prove that two or more related, but not identical var2csa-type genes exist in many P.falciparum isolates. Although there is a conserved locus on chromosome 12, the positions of the other var2csa-type gene loci appear to vary between genomes and are currently mapped to chromosome 1, 8 and the chromosome 5–8 aggregation. In two cultures of monoclonal isolates containing multiple var2csa-type genes (Tz748 and Tz745 respectively), we have shown that both genes are transcribed and neither is an inert pseudo gene. Gene mapping insights into the unexpected complexities of the var2csa-type genes illustrates the need for genetic assays in addition to sequencing when studying gene/phenotype interactions in large multi-gene families. Chromosome mapping has recently revealed another PfEMP1/var gene duplication, in which the Itvar4 gene has been duplicated and transposition via mitotic recombination in culture, from within the chromosome 9–7 aggregation, into a site adjacent to the chromosome 12 var2csa-type gene [56]. Again, the chromosome 12 position of a var2csa-type gene remained conserved in these CS2/EB8 lineage parasites (derived from the FCR3/It4 lineage), where it functions as an expression site for transcription of full-length mRNA from both the Itvar4 and the var2csa-type gene [57].
Our quantitative PCR assays indicate that the expansion of the var2csa sub-family is a common feature of natural populations of P.falciparum isolates. Around twenty percent of the isolates examined and up to a third of Tanzanian placental isolates contain two or more var2csa-type genes/haploid genome. The fact that a higher proportion of placental isolates have multiple var2csa-type genes than occur in samples from non-pregnant individuals may indicate that the multi-locus genotypes have a selective advantage in placental malaria infections. However, larger sample sizes are needed to confirm or reject the hypothesis that multiple var2csa-type genes are a polymorphism which confers greater antigenic variation capacity or a gene dosage and increased adhesion production advantage to the multi-locus genotype parasites in a placental malaria infection, but not in other host infections.
Where two var2csa-type genes exist in one genome, the sequences of each gene do not seem to be more closely related to each other than either is to any other randomly compared var2csa-type sequence in a different genome. In addition, neither of the var2csa-type sequences that have been sequenced in a multi-copy genome displays any unique sequence variation and they have the same polymorphic positions as all other var2csa-type sequences. These results imply that the multiple copies of var2csa-type genes found in some clones are not recently duplicated paralogs experiencing relaxed functional constraints.
Our analysis of the VAR2CSA DBL2X sequences found a 26 amino acid dimorphic sequence, the largest dimorphic region yet identified in a PfEMP1 gene. Structural modeling of one DBL2X variant combined with ligand binding and antibody epitope mapping studies indicate that this dimorphic sequence is surface exposed on the VAR2CSA antigen, binds the glycosaminoglycan ligand CSA and is also a binding site for IgG antibodies from women who had suffered pregnancy associated malaria [58]. This motif also seems to be conserved in the relatively distantly related P.reichenowi var2csa-type gene. Together, these observations argue that this dimorphic sequence and this region of the antigen have some function which has been conserved through a significant period of the evolution of the P.falciparum lineage of malaria parasites. The existence of strong linkage disequilibrium between dimorphic variant types seems unlikely as var2csa-type gene loci occur on different chromosomes and will be randomly re-assorted during the P.falciparum meiosis in the insect vector.
Although it has not previously recognized in PfEMP1 proteins, regions of dimorphic variation have also been reported in several other major P.falciparum antigens, notably the merozoite surface antigens MSP-1 [59], MSP-2 [60], MSP-3 [61], MSP-6 [62] and EBA175 [63]. Various speculative evolutionary explanations for this notable and peculiar type of polymorphism in genes encoding P.falciparum antigens have been proposed. These include adaptation to divergent local host populations, ancient speciation and re-unification, recurrent gene duplication and deletion and population bottlenecks (reviewed in Roy, Ferreira & Hartl [64]).
Our data are noteworthy because the var2csa-type genes are the only PfEMP1/var gene so far shown to have recognized dimorphic sequences. This is possibly related to them also being the most conserved of the var genes. Thus the var2csa-type genes have retained some evidence of dimorphic and, unlike the merozoite antigen genes so far analyzed, are occasionally duplicated. This gives a precedent, and some support for the hypothesis that dimorphism in P.falciparum antigens may be the result of paralogous evolution following gene duplication, followed by later loss of one copy [65].
Our data indicate that dimorphic sequences tends to disappear more rapidly from the fastest evolving genes encoding the recombinogenic [66] and hyper-variable [67] PfEMP1 antigens than from the genes encoding the highly polymorphic, but relatively more slowly evolving merozoite surface antigens. The fact that this distinct dimorphic region still distinguishes the DBL2X domain of all known VAR2CSA sequences argues that each dimorphic variant has some essential function. The VAR2CSA DBL2X dimorphic sequence is in fact an immuno-dominant region of that antigen [58]. It is interesting that this experimental observation was predicted by earlier models for the maintenance of such dimorphic antigenic polymorphisms in P.falciparum evolution [68]–[70].
Supporting Information
Sequence alignment of full-length VAR2CSA DBL2X domains. In the P.falciparum 3D7 reference genome this domain includes amino acids 535–878 of the VAR2CSA sequence encoded by the PFL0030c gene. The multiple sequence alignment of these approximately 343 amino acids includes 37 VAR2CSA DBL2X sequences derived from Senegalese placental isolates and 18 database-derived sequences, including P.reichenowi. Color blocks mark alignment positions with >55% amino acid sequence identity. The dimorphic sequence motif (DSM) is shown between positions 50–78.
(10.42 MB TIF)
Acknowledgments
We thank Jonas Fjeldbye and Nahla Chehabi for excellent technical assistance and Stuart McNeill at the University of Copenhagen Biocenter for allowing us to use his apparatus for pulsed field gel electrophoresis.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: The work was funded by a Danish National Research Fundation (http://www.dg.dk/) Niels Bohr Foundation Visiting Professorship (Project: 312000-50-64920) to DEA. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Duffy PE, Fried M. Plasmodium falciparum adhesion in the placenta. Curr Opin Microbiol. 2003;6:371–376. doi: 10.1016/s1369-5274(03)00090-0. [DOI] [PubMed] [Google Scholar]
- 2.Hviid L, Salanti A. VAR2CSA and protective immunity against pregnancy-associated Plasmodium falciparum malaria. Parasitology. 2007;134:1871–1876. doi: 10.1017/S0031182007000121. [DOI] [PubMed] [Google Scholar]
- 3.Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, et al. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
- 4.Bengtsson D, Sowa KM, Salanti A, Jensen AT, Joergensen L, et al. A method for visualizing surface-exposed and internal PfEMP1 adhesion antigens in Plasmodium falciparum infected erythrocytes. Malar J. 2008;7:101. doi: 10.1186/1475-2875-7-101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, et al. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
- 6.Lavstsen T, Salanti A, Jensen AT, Arnot DE, Theander TG. Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J. 2003;2:27. doi: 10.1186/1475-2875-2-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kraemer SM, Kyes SA, Aggarwal G, Springer AL, Nelson SO, et al. Patterns of gene recombination shape var gene repertoires in Plasmodium falciparum: comparisons of geographically diverse isolates. BMC Genomics. 2007;8:45. doi: 10.1186/1471-2164-8-45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rowe JA, Kyes SA. The role of Plasmodium falciparum var genes in malaria in pregnancy. Mol Microbiol. 2004;53:1011–1019. doi: 10.1111/j.1365-2958.2004.04256.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Viebig NK, Levin E, Dechavanne S, Rogerson SJ, Gysin J, et al. Disruption of var2csa gene impairs placental malaria associated adhesion phenotype. PLoS ONE. 2007;2:e910. doi: 10.1371/journal.pone.0000910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Duffy PE, Fried M. Plasmodium falciparum adhesion in the placenta. Curr Opin Microbiol. 2003;6:371–376. doi: 10.1016/s1369-5274(03)00090-0. [DOI] [PubMed] [Google Scholar]
- 11.Rogerson SJ, Mwapasa V, Meshnick SR. Malaria in pregnancy: linking immunity and pathogenesis to prevention. Am J Trop Med Hyg. 2007;77:14–22. [PubMed] [Google Scholar]
- 12.Tuikue NN, Deloron P. Towards a vaccine against pregnancy-associated malaria. Parasite. 2008;15:515–521. doi: 10.1051/parasite/2008153515. [DOI] [PubMed] [Google Scholar]
- 13.Dahlback M, Rask TS, Andersen PH, Nielsen MA, Ndam NT, et al. Epitope mapping and topographic analysis of VAR2CSA DBL3X involved in P. falciparum placental sequestration. PLoS Pathog. 2006;2:e124. doi: 10.1371/journal.ppat.0020124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lavstsen T, Salanti A, Jensen AT, Arnot DE, Theander TG. Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J. 2003;2:27. doi: 10.1186/1475-2875-2-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ward CP, Clottey GT, Dorris M, Ji DD, Arnot DE. Analysis of Plasmodium falciparum PfEMP-1/var genes suggests that recombination rearranges constrained sequences. Mol Biochem Parasitol. 1999;102:167–177. doi: 10.1016/s0166-6851(99)00106-1. [DOI] [PubMed] [Google Scholar]
- 16.Baruch DI, Ma XC, Singh HB, Bi X, Pasloske BL, et al. Identification of a region of PfEMP1 that mediates adherence of Plasmodium falciparum infected erythrocytes to CD36: conserved function with variant sequence. Blood. 1997;90:3766–3775. [PubMed] [Google Scholar]
- 17.Tuikue NN, Bischoff E, Proux C, Lavstsen T, Salanti A, et al. Plasmodium falciparum transcriptome analysis reveals pregnancy malaria associated gene expression. PLoS ONE. 2008;3:e1855. doi: 10.1371/journal.pone.0001855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Roper C, Richardson W, Elhassan IM, Giha H, Hviid L, et al. Seasonal changes in the Plasmodium falciparum population in individuals and their relationship to clinical malaria: a longitudinal study in a Sudanese village. Parasitology. 1998;116 ( Pt 6):501–510. doi: 10.1017/s0031182098002650. [DOI] [PubMed] [Google Scholar]
- 19.Magistrado P, Salanti A, Tuikue Ndam NG, Mwakalinga SB, Resende M, et al. VAR2CSA expression on the surface of placenta-derived Plasmodium falciparum-infected erythrocytes. J Infect Dis. 2008;198:1071–1074. doi: 10.1086/591502. [DOI] [PubMed] [Google Scholar]
- 20.Roper C, Richardson W, Elhassan IM, Giha H, Hviid L, et al. Seasonal changes in the Plasmodium falciparum population in individuals and their relationship to clinical malaria: a longitudinal study in a Sudanese village. Parasitology. 1998;116 ( Pt 6):501–510. doi: 10.1017/s0031182098002650. [DOI] [PubMed] [Google Scholar]
- 21.Corcoran LM, Forsyth KP, Bianco AE, Brown GV, Kemp DJ. Chromosome size polymorphisms in Plasmodium falciparum can involve deletions and are frequent in natural parasite populations. Cell. 1986;44:87–95. doi: 10.1016/0092-8674(86)90487-3. [DOI] [PubMed] [Google Scholar]
- 22.Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, et al. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
- 23.Dahlback M, Rask TS, Andersen PH, Nielsen MA, Ndam NT, et al. Epitope mapping and topographic analysis of VAR2CSA DBL3X involved in P. falciparum placental sequestration. PLoS Pathog. 2006;2:e124. doi: 10.1371/journal.ppat.0020124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 25.Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, et al. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
- 26.Smith JD, Subramanian G, Gamain B, Baruch DI, Miller LH. Classification of adhesive domains in the Plasmodium falciparum erythrocyte membrane protein 1 family. Mol Biochem Parasitol. 2000;110:293–310. doi: 10.1016/s0166-6851(00)00279-6. [DOI] [PubMed] [Google Scholar]
- 27.Bahl A, Brunk B, Coppel RL, Crabtree J, Diskin SJ, et al. PlasmoDB: the Plasmodium genome resource. An integrated database providing tools for accessing, analyzing and mapping expression and sequence data (both finished and unfinished). Nucleic Acids Res. 2002;30:87–90. doi: 10.1093/nar/30.1.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Creasey AM, Staalsoe T, Raza A, Arnot DE, Rowe JA. Nonspecific immunoglobulin M binding and chondroitin sulfate A binding are linked phenotypes of Plasmodium falciparum isolates implicated in malaria during pregnancy. Infect Immun. 2003;71:4767–4771. doi: 10.1128/IAI.71.8.4767-4771.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Semblat JP, Raza A, Kyes SA, Rowe JA. Identification of Plasmodium falciparum var1CSA and var2CSA domains that bind IgM natural antibodies. Mol Biochem Parasitol. 2006;146:192–197. doi: 10.1016/j.molbiopara.2005.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ghumra A, Shi J, McIntosh RS, Rasmussen IB, Braathen R, et al. Structural requirements for the interaction of human IgM and IgA with the human Fcalpha/mu receptor. Eur J Immunol. 2009;39:1147–1156. doi: 10.1002/eji.200839184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dahlback M, Rask TS, Andersen PH, Nielsen MA, Ndam NT, et al. Epitope mapping and topographic analysis of VAR2CSA DBL3X involved in P. falciparum placental sequestration. PLoS Pathog. 2006;2:e124. doi: 10.1371/journal.ppat.0020124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gowda AS, Madhunapantula SV, Achur RN, Valiyaveettil M, Bhavanandan VP, et al. Structural basis for the adherence of Plasmodium falciparum-infected erythrocytes to chondroitin 4-sulfate and design of novel photoactivable reagents for the identification of parasite adhesive proteins. J Biol Chem. 2007;282:916–928. doi: 10.1074/jbc.M604741200. [DOI] [PubMed] [Google Scholar]
- 33.Higgins MK. The structure of a chondroitin sulfate-binding domain important in placental malaria. J Biol Chem. 2008;283:21842–21846. doi: 10.1074/jbc.C800086200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bentley GA, Gamain B. How does Plasmodium falciparum stick to CSA? Let's see in the crystal. Nat Struct Mol Biol. 2008;15:895–897. doi: 10.1038/nsmb0908-895. [DOI] [PubMed] [Google Scholar]
- 35.Gamain B, Smith JD, Avril M, Baruch DI, Scherf A, et al. Identification of a 67-amino-acid region of the Plasmodium falciparum variant surface antigen that binds chondroitin sulphate A and elicits antibodies reactive with the surface of placental isolates. Mol Microbiol. 2004;53:445–455. doi: 10.1111/j.1365-2958.2004.04145.x. [DOI] [PubMed] [Google Scholar]
- 36.Gamain B, Trimnell AR, Scheidig C, Scherf A, Miller LH, et al. Identification of multiple chondroitin sulfate A (CSA)-binding domains in the var2CSA gene transcribed in CSA-binding parasites. J Infect Dis. 2005;191:1010–1013. doi: 10.1086/428137. [DOI] [PubMed] [Google Scholar]
- 37.Barfod L, Bernasconi NL, Dahlback M, Jarrossay D, Andersen PH, et al. Human pregnancy-associated malaria-specific B cells target polymorphic, conformational epitopes in VAR2CSA. Mol Microbiol. 2007;63:335–347. doi: 10.1111/j.1365-2958.2006.05503.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sharling L, Enevold A, Sowa KM, Staalsoe T, Arnot DE. Antibodies from malaria-exposed pregnant women recognize trypsin resistant epitopes on the surface of Plasmodium falciparum-infected erythrocytes selected for adhesion to chondroitin sulphate A. Malar J. 2004;3:31. doi: 10.1186/1475-2875-3-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nielsen MA, Pinto VV, Resende M, Dahlback M, Ditlev SB, et al. Induction of adhesion-inhibitory antibodies against placental Plasmodium falciparum parasites using single domains of VAR2CSA. Infect Immun. 2009 doi: 10.1128/IAI.00159-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dahlback M, Rask TS, Andersen PH, Nielsen MA, Ndam NT, et al. Epitope mapping and topographic analysis of VAR2CSA DBL3X involved in P. falciparum placental sequestration. PLoS Pathog. 2006;2:e124. doi: 10.1371/journal.ppat.0020124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bockhorst J, Lu F, Janes JH, Keebler J, Gamain B, et al. Structural polymorphism and diversifying selection on the pregnancy malaria vaccine candidate VAR2CSA. Mol Biochem Parasitol. 2007;155:103–112. doi: 10.1016/j.molbiopara.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 42.Dahlback M, Rask TS, Andersen PH, Nielsen MA, Ndam NT, et al. Epitope mapping and topographic analysis of VAR2CSA DBL3X involved in P. falciparum placental sequestration. PLoS Pathog. 2006;2:e124. doi: 10.1371/journal.ppat.0020124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Singh SK, Hora R, Belrhali H, Chitnis CE, Sharma A. Structural basis for Duffy recognition by the malaria parasite Duffy-binding-like domain. Nature. 2006;439:741–744. doi: 10.1038/nature04443. [DOI] [PubMed] [Google Scholar]
- 44.Bentley GA, Gamain B. How does Plasmodium falciparum stick to CSA? Let's see in the crystal. Nat Struct Mol Biol. 2008;15:895–897. doi: 10.1038/nsmb0908-895. [DOI] [PubMed] [Google Scholar]
- 45.Higgins MK. The structure of a chondroitin sulfate-binding domain important in placental malaria. J Biol Chem. 2008;283:21842–21846. doi: 10.1074/jbc.C800086200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bockhorst J, Lu F, Janes JH, Keebler J, Gamain B, et al. Structural polymorphism and diversifying selection on the pregnancy malaria vaccine candidate VAR2CSA. Mol Biochem Parasitol. 2007;155:103–112. doi: 10.1016/j.molbiopara.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 47.Bockhorst J, Lu F, Janes JH, Keebler J, Gamain B, et al. Structural polymorphism and diversifying selection on the pregnancy malaria vaccine candidate VAR2CSA. Mol Biochem Parasitol. 2007;155:103–112. doi: 10.1016/j.molbiopara.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 48.Lavstsen T, Salanti A, Jensen AT, Arnot DE, Theander TG. Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J. 2003;2:27. doi: 10.1186/1475-2875-2-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Corcoran LM, Forsyth KP, Bianco AE, Brown GV, Kemp DJ. Chromosome size polymorphisms in Plasmodium falciparum can involve deletions and are frequent in natural parasite populations. Cell. 1986;44:87–95. doi: 10.1016/0092-8674(86)90487-3. [DOI] [PubMed] [Google Scholar]
- 50.Aurrecoechea C, Brestelli J, Brunk BP, Dommer J, Fischer S, et al. PlasmoDB: a functional genomic database for malaria parasites. Nucleic Acids Res. 2009;37:D539–D543. doi: 10.1093/nar/gkn814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bahl A, Brunk B, Coppel RL, Crabtree J, Diskin SJ, et al. PlasmoDB: the Plasmodium genome resource. An integrated database providing tools for accessing, analyzing and mapping expression and sequence data (both finished and unfinished). Nucleic Acids Res. 2002;30:87–90. doi: 10.1093/nar/30.1.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gardner MJ, Hall N, Fung E, White O, Berriman M, et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature. 2002;419:498–511. doi: 10.1038/nature01097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Arnot DE. The influence of the genetic complexity of Plasmodium falciparum infections on the epidemiology of malaria. Trans R Soc Trop Med Hyg. 2002;96(Suppl 1):S131–S136. doi: 10.1016/s0035-9203(02)90064-7. [DOI] [PubMed] [Google Scholar]
- 54.Hviid L. The immuno-epidemiology of pregnancy-associated Plasmodium falciparum malaria: a variant surface antigen-specific perspective. Parasite Immunol. 2004;26:477–486. doi: 10.1111/j.0141-9838.2004.00733.x. [DOI] [PubMed] [Google Scholar]
- 55.Soerli J, Barfod L, Lavstsen T, Bernasconi N, Lanzavecchia A, et al. IgG selection of Plasmodium falciparum for expression of placental malaria-specific variant surface antigens. Parasite Immunol. 2009;31:341–346. doi: 10.1111/j.1365-3024.2009.01097.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Duffy MF, Byrne TJ, Carret C, Ivens A, Brown GV. Ectopic recombination of a malaria var gene during mitosis associated with an altered var switch rate. J Mol Biol. 2009;389:453–469. doi: 10.1016/j.jmb.2009.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Duffy MF, Byrne TJ, Carret C, Ivens A, Brown GV. Ectopic recombination of a malaria var gene during mitosis associated with an altered var switch rate. J Mol Biol. 2009;389:453–469. doi: 10.1016/j.jmb.2009.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Resende M, Nielsen MA, Dahlback M, Ditlev SB, Andersen P, et al. Identification of glycosaminoglycan binding regions in the Plasmodium falciparum encoded placental sequestration ligand, VAR2CSA. Malar J. 2008;7:104. doi: 10.1186/1475-2875-7-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Tanabe K, Mackay M, Goman M, Scaife JG. Allelic dimorphism in a surface antigen gene of the malaria parasite Plasmodium falciparum. J Mol Biol. 1987;195:273–287. doi: 10.1016/0022-2836(87)90649-8. [DOI] [PubMed] [Google Scholar]
- 60.Smythe JA, Coppel RL, Day KP, Martin RK, Oduola AM, et al. Structural diversity in the Plasmodium falciparum merozoite surface antigen 2. Proc Natl Acad Sci U S A. 1991;88:1751–1755. doi: 10.1073/pnas.88.5.1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.McColl DJ, Anders RF. Conservation of structural motifs and antigenic diversity in the Plasmodium falciparum merozoite surface protein-3 (MSP-3). Mol Biochem Parasitol. 1997;90:21–31. doi: 10.1016/s0166-6851(97)00130-8. [DOI] [PubMed] [Google Scholar]
- 62.Pearce JA, Triglia T, Hodder AN, Jackson DC, Cowman AF, et al. Plasmodium falciparum merozoite surface protein 6 is a dimorphic antigen. Infect Immun. 2004;72:2321–2328. doi: 10.1128/IAI.72.4.2321-2328.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ware LA, Kain KC, Lee Sim BK, Haynes JD, Baird JK, et al. Two alleles of the 175-kilodalton Plasmodium falciparum erythrocyte binding antigen. Mol Biochem Parasitol. 1993;60:105–109. doi: 10.1016/0166-6851(93)90033-t. [DOI] [PubMed] [Google Scholar]
- 64.Roy SW, Ferreira MU, Hartl DL. Evolution of allelic dimorphism in malarial surface antigens. Heredity. 2008;100:103–110. doi: 10.1038/sj.hdy.6800887. [DOI] [PubMed] [Google Scholar]
- 65.Roy SW, Ferreira MU, Hartl DL. Evolution of allelic dimorphism in malarial surface antigens. Heredity. 2008;100:103–110. doi: 10.1038/sj.hdy.6800887. [DOI] [PubMed] [Google Scholar]
- 66.Bockhorst J, Lu F, Janes JH, Keebler J, Gamain B, et al. Structural polymorphism and diversifying selection on the pregnancy malaria vaccine candidate VAR2CSA. Mol Biochem Parasitol. 2007;155:103–112. doi: 10.1016/j.molbiopara.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 67.Barry AE, Leliwa-Sytek A, Tavul L, Imrie H, Migot-Nabias F, et al. Population genomics of the immune evasion (var) genes of Plasmodium falciparum. PLoS Pathog. 2007;3:e34. doi: 10.1371/journal.ppat.0030034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.McKenzie FE, Ferreira MU, Baird JK, Snounou G, Bossert WH. Meiotic recombination, cross-reactivity, and persistence in Plasmodium falciparum. Evolution. 2001;55:1299–1307. doi: 10.1111/j.0014-3820.2001.tb00652.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ferreira MU, da Silva NM, Wunderlich G. Antigenic diversity and immune evasion by malaria parasites. Clin Diagn Lab Immunol. 2004;11:987–995. doi: 10.1128/CDLI.11.6.987-995.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Roy SW, Ferreira MU, Hartl DL. Evolution of allelic dimorphism in malarial surface antigens. Heredity. 2008;100:103–110. doi: 10.1038/sj.hdy.6800887. [DOI] [PubMed] [Google Scholar]
- 71.Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, et al. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol. 2003;49:179–191. doi: 10.1046/j.1365-2958.2003.03570.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Sequence alignment of full-length VAR2CSA DBL2X domains. In the P.falciparum 3D7 reference genome this domain includes amino acids 535–878 of the VAR2CSA sequence encoded by the PFL0030c gene. The multiple sequence alignment of these approximately 343 amino acids includes 37 VAR2CSA DBL2X sequences derived from Senegalese placental isolates and 18 database-derived sequences, including P.reichenowi. Color blocks mark alignment positions with >55% amino acid sequence identity. The dimorphic sequence motif (DSM) is shown between positions 50–78.
(10.42 MB TIF)