

Abstract
Iron is a critical micronutrient, and iron derived from heme contributes a large proportion of the total iron absorbed in a typical Western diet. Heme iron is absorbed by different mechanisms than non-heme iron, but despite considerable study over many years these mechanisms remain poorly understood. This review provides an overview of the importance of heme iron in the diet and discusses the two prevailing hypotheses of heme absorption; namely receptor mediated endocytosis of heme, and direct transport into the intestinal enterocyte by recently discovered heme transporters. A specific emphasis is placed on the questions surrounding the site of heme catabolism and the identity of the enzyme that performs this task. Additionally, we present the hypothesis that a non-heme iron transport protein may be required for heme iron absorption and discuss the experiences of our laboratory in examining this hypothesis.
Keywords: Iron, Heme absorption, Receptor mediated endocytosis, Heme transporter, Heme oxygenase
THE IMPORTANCE OF DIETARY IRON AND HEME
Iron is a vitally important element in biological terms (for review see[1]). Iron is a transition metal with the ability to readily accept and donate electrons, allowing it to function as an oxidant or reductant in a large number of biochemical reactions. In mammals, iron is notably required for oxygen transport as a component of hemoglobin, DNA synthesis as a component of ribonucleotide reductase, and as an electron acceptor/donor in the cytochromes that are essential for energy transduction. Currently, iron deficiency is the most common diet related health problem in the world[2], and the effects on human health are wide ranging. Iron deficiency manifests as anaemia in up to 2 billion people, impairs physical and mental development in children, and can exacerbate many other health problems.
Heme is a biologically important iron containing compound and a key source of dietary iron. Historically, it was doubted that heme iron could be absorbed by the enterocyte and it was not until 1955 that the absorption of heme-derived iron was demonstrated for the first time[3]. Currently, the importance of heme iron in the diet cannot be underestimated. Studies estimate that in Western societies, iron derived from heme sources such as myoglobin and hemoglobin make up two-thirds of the average person’s total iron stores despite only constituting one-third of the iron that is actually ingested[4–6]. This likely explains why vegetarians are more prone to iron deficiency than those who regularly consume red meat[7].
The relative importance of dietary heme is attributable to its high bioavailability compared with non-heme iron in the predominantly alkaline conditions found in the lumen of the small intestine. In aqueous solutions at or above pH 7.0, non-heme iron is present as Fe(II) and Fe(III). Fe(II) readily oxidizes to Fe(III) which precipitates from solution as ferric hydroxide or forms soluble hydroxyl-iron dimers which are not directly available for absorption[8]. Further, many dietary components (particularly humic substances such as tannins and phytate) can chelate iron making it non-bioavailable[9–11], while only select reductants in the diet (such as ascorbate) can act as solubilizing agents[12].
In contrast polymerization of heme, which reduces its absorption, is minimised in alkaline conditions[13] while humic substances[14,15] and chelators such as desferrioxamine[16,17] do not reduce heme bioavailability. Heme solubility is also increased significantly by the presence of protein[13,18–20] which is important con-sidering heme-rich foods typically contain high quantities of protein. Paradoxically, the absorption of heme iron cannot upregulate to the same extent as non-heme iron during iron deficiency[14,15,21–25]. This is possibly due to rate limitations at the step of heme catabolism (see ‘Heme Catabolism’ below) although the extent to which enterocyte adaptation (particularly transfer to the circulation by ferroportin[26–29] and humoral regulation by hepcidin[30–33]) affects heme iron absorption is very poorly characterised compared to non-heme iron.
Despite the clear importance of dietary heme as a source of body iron, the mechanism by which the enterocyte takes up heme and catabolizes it to utilise the iron is still poorly understood. Heme is taken up as an intact metalloporphyrin[34,35] and comparative studies show that a molecule with a similar size, structure, and ionic charge to heme (namely Vitamin B12) was not absorbed by enterocytes because of the absence of specific carriers on the apical membrane, whereas in identical conditions up to 19% of an equivalent dose of heme was absorbed[36]. This strongly suggests that heme uptake is a facilitated process, as opposed to simple diffusion.
Currently, there are two prevailing hypotheses explaining the mechanisms of this process; firstly, a long-standing hypothesis that heme is taken up by receptor mediated endocytosis; secondly, the recent discovery of a heme transporter that may have the capability of transferring heme from the small intestinal lumen directly into the cytoplasm. These pathways are summarised in Figure 1 and discussed in detail below.
Figure 1.
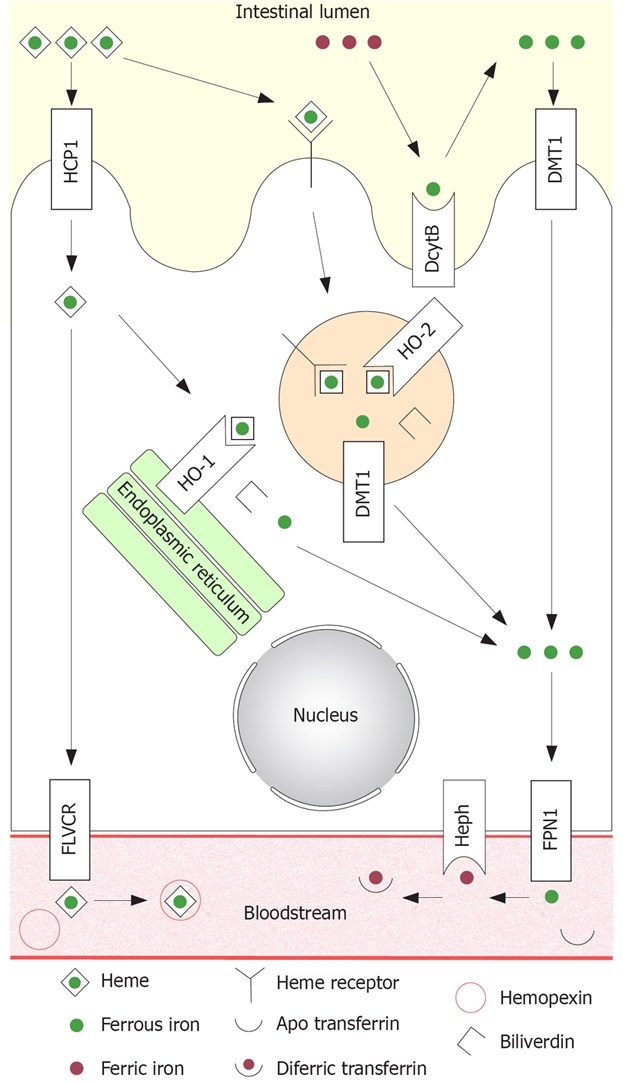
Summary diagram of established and putative iron absorption pathways in the intestinal enterocyte. Non-heme iron: All non-heme iron is ultimately taken up from the lumen by DMT1 situated on the microvillus membrane, before joining the labile iron pool in the cytoplasm. Ferric iron must first be reduced to the ferrous form by DcytB before uptake. Ferrous iron in the labile iron pool is then transferred to the circulation by FPN1, which requires hephaestin for oxidation to the ferric form in order to bind to circulating apotransferrin. Heme iron: Heme iron is hypothesized to be taken up by receptor mediated endocytosis. Internalised heme is degraded by HO-2 inside the vesicles, releasing non-heme iron and generating biliverdin. The non-heme iron is then transported to the cytoplasm by DMT1. Heme iron may also be taken up by PCFT/HCP1 directly into the cytoplasm. Intact heme may be transported across the basolateral membrane by FLVCR where it binds circulating hemopexin. Alternatively, heme may be catabolized to non-heme iron and biliverdin by HO-1 located on the endoplasmic reticulum. Any iron released from heme inside the enterocyte, regardless of the mode of uptake, ultimately joins the labile iron pool and is transferred to the bloodstream by FPN1 in the same fashion as non-heme iron.
HEME RECEPTOR MEDIATED ENDOCYTOSIS
The hypothesis of heme uptake by receptor mediated endocytosis originated in 1979 from the discovery of a heme binding protein on the microvillus membrane of the upper small intestine of both pigs and humans[37]. Characterisation of this binding protein demonstrated that it was not albumin, hemopexin, glutathione-S-transferase, or aggregated heme[38]. The dissociation constant for the binder/heme complex was found to be 10-6 to 10-7 mol/L using radioactive ligands, and 10-9 mol/L in spectral studies[39]. This high affinity binding, along with loss of binding capacity with trypsin digestion, indicated the presence of a protein receptor for heme on the microvillus membrane. Heme uptake by rat duodenal enterocytes increases with iron deficiency and correlates with a modest but statistically significant increase in heme binding capacity[36], suggesting that the quantity of receptor present on the microvillus membrane contributes to regulation of heme iron absorption.
A heme binding protein has also been characterised on the membrane of erythroleukemia cells with very similar properties to that observed in the duodenum[40]. These cell types are capable of internalising heme intact, as evidenced by the binding and uptake of heme-embedded latex beads[41,42]. Heme uptake is also temperature[43] and ATP[44] dependent, and these combined data provide strong evidence for the ability of cells to actively take up heme by endocytosis.
Morphological evidence corroborates this assess-ment. In two similar studies, heme or hemoglobin were administered into closed duodenal loops of rats or dogs, and duodenal tissue samples collected over a time period of up to 3 h thereafter[45,46]. Heme in the duodenal mucosa was reacted with DAB to produce an electron dense precipitate that was then observed by electron microscopy. Both studies reported the appearance of heme initially on the microvillus membrane, then within tubulovesicular structures in the apical cytoplasm, before collecting in vesicles identified as secondary lysosomes[46]. Heme disappeared from within these vesicles approximately 2-3 h after the initial dose. There was no heme observed in the basal cytoplasm or the extracellular space, consistent with heme uptake at the microvillus membrane by an endocytotic pathway and its catabolism within the apical cytoplasm of the cell.
One important criticism of the receptor mediated endocytosis hypothesis is that it assumes iron released from heme is transported out of the internalised vesicles in order to join the labile iron pool. Currently, no such transport process has been identified. However, it is possible that divalent metal transporter 1 (DMT1) may fulfil this role in a manner analogous to its role in the transferrin receptor cycle[47–50] (see ‘A Possible Role for DMT1?’ below).
HEME TRANSPORTERS
In recent years, two mammalian heme transporters have been discovered, namely PCFT/HCP1[51,52] and FLVCR[53]. These appear to function independently of the putative heme receptor and receptor mediated endocytosis in that they act as a direct transfer process across plasma membranes. At this early stage the physiological relevance of these transporters to intestinal heme iron absorption is unclear, but the information that is available will be considered below.
The PCFT/HCP1 cDNA was initially isolated by the subtractive suppressive hybridisation of ileal cDNA from duodenal cDNA in hypotransferrinaemic mice[51]. Heme transport capability by PCFT/HCP1 has been demonstrated in vitro, with expression in Xenopus oocytes and the HeLa cell line resulting in a 2-fold to 3-fold increase in heme uptake. Heme uptake by HeLa cells expressing PCFT/HCP1 was greatly reduced at 4°C compared to 37°C which was interpreted as energy dependence, although without the specific use of metabolic inhibitors to distinguish between temperature dependent kinetic properties of the transporter and a requirement for metabolic fuels, this interpretation is ambiguous.
Uptake of radio-labelled heme by transfected CHO cells was competitive with the uptake of unlabelled heme, zinc protoporphyrin, and protoporphyrin suggesting that transport is selective for the porphyrin ring, although no non-porphyrin competitors appear to have been assessed. However, the addition of PCFT/HCP1 siRNA to CHO cells did not reduce the substantial basal heme uptake of control cells[51]. Clearly, CHO cells have significant pre-existing heme acquisition pathways that are not related to PCFT/HCP1. In vivo, heme uptake from closed duodenal loops in normal and hypoxic mice was modestly reduced (30%-40%) by the addition of a PCFT/HCP1 antibody but no decrease was seen with pre-immune serum. PCFT/HCP1 gene expression was significantly increased by hypoxia but was not significantly altered by iron deficiency which appears contrary to the upregulation of heme binding and uptake shown in other studies[36], although regulation of heme transport may instead be regulated by the sub-cellular location of PCFT/HCP1.
PCFT/HCP1 has been independently characterised as a folate/proton symporter and appears to play a key role in intestinal folate absorption[52,54]. This is evidenced by a 55%-80% reduction in pH dependent folate uptake in the enterocyte-like CaCo-2 cell line following RNA interference for PCFT/HCP1. Additionally, human patients diagnosed with hereditary folate malabsorption carry a point mutation to PCFT/HCP1 that results in the formation of a non-functional splice variant. Interestingly, the folate transport capabilities of PCFT/HCP1 are at least an order of magnitude higher than that observed for heme, suggesting that folate may be the more physiologically relevant target of this transport protein. It is clear that the generation of a knockout model for PCFT/HCP1 is required to assess the importance of this heme/folate transporter in vivo.
FLVCR was initially characterised as the cell surface protein receptor for feline leukaemia virus subgroup C which causes severe anaemia in infected cats[55,56]. It has since been demonstrated that loss of FLVCR function in erythropoietic cells is associated with impairment of erythroid maturation and increased apoptosis, and that heme content of erythropoietic primary cultures is dependent upon FLVCR expression[53]. Since cells expressing FLVCR are capable of actively exporting heme, it was concluded that FLVCR acts as an overflow valve for excess manufactured heme that would otherwise result in cellular toxicity by producing oxidative stress before it can bind to globins for hemoglobin production.
With regard to intestinal heme iron absorption, no studies have yet directly examined FLVCR function in vivo, though the polarized intestinal cell line CaCo-2 does express the protein[53]. It has also been demonstrated that CaCo-2 cells are capable of heme transport in both directions, equivalent to both absorption and secretion in vivo, and that the secretory pathway is significantly more active under control conditions[57]. Presumably, FLVCR is acting on the basolateral membrane to regulate heme content when heme synthesis rates are at their peak just prior to differentiation, and the potential for cell damage from oxidative stress is greatest if HO-1 activity is impaired[58–60] (see ‘Heme Oxygenase’ below). In this case it is highly unlikely that FLVCR is involved with heme uptake at the apical membrane. Studies of FLVCR in vivo are required to confirm this assessment.
HEME CATABOLISM IN THE ENTEROCYTE
It was initially hypothesized that following uptake, heme passed directly into the portal circulation where it bound hemopexin and was most likely sequestered by hepatocytes using the hemopexin receptor and degraded, based on early observations in guinea pigs[23]. However, this theory is questionable for other species, with strong evidence that heme is catabolized within the enterocyte in most omnivorous and carnivorous mammals. This is best demonstrated by experiments in which dogs were administered an intragastric dose of radio-labelled hemoglobin, and 90% of the recoverable radioactivity in samples of portal blood over a period of 3 h was present as non-heme iron[35]. Similar observations have been made in human[14,34] and rat[17] experiments.
The presence of a heme splitting substance in the mucosa was first demonstrated in 1968[61]. The high molecular weight of this substance (MW about 64 kDa) and the kinetic properties of the reaction indicated that the heme splitting substance was an enzyme. Initial studies suggested that xanthine oxidase could play a role by generating hydrogen peroxide to chemically degrade heme, resulting in iron release and a non-specific mixture of four bilirubin isomers[62–64]. However, this hypothesis was problematic since in vivo heme degradation typically results in a single dominant isomer, namely bilirubin IX-α[65,66].
Further research generated a strong case that the heme splitting substance in the mucosa was microsomal heme oxygenase[24]. This is based on the fact that heme oxygenase almost exclusively generates the expected bilirubin IX-α isomer and that heme oxygenase activity is highest in the location where heme iron absorption is highest, the duodenum[17,23,37]. In addition, iron deficiency results in an increase in both heme iron absorption and mucosal heme oxygenase activity, whereas xanthine oxidase activity decreases dramatically.
Based on morphological studies, it appears that heme is degraded inside internalised vesicles within 2-3 h of heme uptake by receptor mediated endocytosis[45,46]. Acid ferrocyanide staining, which exclusively detects non-heme iron, indicates that iron is released from heme inside the vesicle, before transport to the labile iron pool by unknown mechanisms (see ‘A Possible Role for DMT1?’ below). The iron is then thought to undergo identical transport through the enterocyte and into the circulation as for internalised non-heme iron.
A study tracking the absorption of 59Fe-hemoglobin in closed duodenal loops has suggested that heme degradation is the rate limiting step in heme iron absorption, as opposed to hemoglobin degradation, heme uptake or iron transfer to the circulation[67]. This is based on increasing doses of hemoglobin resulting in the accumulation of 59Fe-heme, but not 59Fe, within the mucosa. However, since this study utilized whole-mucosal homogenates to assess relative heme and non-heme iron content there may not be sufficient sensitivity to detect the possible accumulation of non-heme iron inside endocytotic vesicles which would result in decreased heme oxygenase activity by end-product inhibition[61]. Nonetheless, the hypothesis that heme oxygenase is limiting for heme iron absorption is consistent with the decrease in absorption that is observed with inhibitors of heme oxygenase activity[68].
HEME OXYGENASE
Heme oxygenase is a microsomal enzyme (corresponding to the endoplasmic reticulum in vivo) that catalyses the mixed function oxidation of heme using cytochrome P-450, NADPH and molecular oxygen producing CO, iron and biliverdin IX-α which is rapidly reduced to bilirubin IX-α[69–71]. There are two well characterised isoforms of heme oxygenase, referred to as HO-1 and HO-2[72,73], and these isoforms are products of different genes[74,75]. A third isoform has been described as HO-3[76], but this appears to be a brain-specific pseudogene derived from HO-2[77].
HO-1 expression is induced by numerous factors including oxidative stress, inflammation, cytokines, nitric oxide, prostaglandins, an elevated level of substrate, iron deficiency, metals including Cd, Co, Cr, Cu, Fe, Hg, Ni, Pd, Pt, Sn and Zn, hyperoxia, and exposure to UV light (for review see[78]). HO-1 is also induced by hyperthermia, leading to the use of the alternate name heat shock protein 32[79]. Considering these combined factors, induction of HO-1 expression appears to be related to preventing cell damage under many circumstances by reducing levels of the pro-oxidant heme and generating the antioxidant bilirubin[80]. This assessment is confirmed by genetic knockout of HO-1 in mice[81], and humans with impaired HO-1 expression[82–84], which present with reduced defence against external stresses.
In contrast, HO-2 expression is not inducible[79,85]. HO-2 is primarily found in the brain[86] and testis[85] and appears to function as a sensor for O2, CO, and NO[87–90]. In the intestine these functions are relevant in the interstitial cells of Cajal, where HO-2 regulates levels of CO, which in turn affects potassium currents and resting membrane potential of intestinal smooth muscle, and thus intestinal motility (for review see[78]).
In relation to the catabolism of dietary heme, most work regarding heme oxygenase was performed before the different isoforms were known. As such, the specific isoform involved with heme iron absorption has not been established with certainty. However, it has long been assumed that HO-1 is the key player[91] based on corroboration of physiological evidence; specifically, HO-1 and heme iron absorption both upregulate in iron deficiency[92]. Further, HO-1 activity[24] and heme iron absorption[17,23,37] are both highest in the duodenum, which is also the site of highest expression of the putative heme receptor and PCFT/HCP1[37,51,52].
However, HO-1 is generally considered to be a membrane bound protein associated with microsomes[69] with a cytoplasmic catalytic site[93]. Thus under normal circumstances HO-1 would have no topological association with the vesicles that are thought to contain internalised heme, namely endosomes and/or lysosomes. As such, the possibility that HO-2 plays a role in heme iron absorption should be strongly considered, since both HO-1 and HO-2 generate the appropriate isoform of bilirubin[72]. Further, the participation of a non-inducible enzyme could explain why the ability to upregulate heme iron absorption is limited compared to non-heme iron[14,15,21–25], and heme splitting is speculated to be rate limiting[67].
In a recent study our laboratory examined the sub-cellular location of HO-1 and HO-2 in enterocytes in relation to endocytotic markers during the course of heme iron absorption[94]. We observed that HO-1 was distributed evenly throughout the cytoplasm of enterocytes and did not co-localise with endocytotic markers. In contrast, HO-2 presented as a dense band in the apical cytoplasm that co-localised extensively with endosomes. This strongly suggests that HO-2 could be exposed to all heme taken up by the enterocytes, either by way of receptor mediated endocytosis or by transport directly into the cytoplasm. Clearly, the role of HO-2 in heme iron absorption needs to be investigated further using more direct methods, particularly using knockout models for HO-1 and HO-2.
A POSSIBLE ROLE FOR DMT1?
The uptake of non-heme iron by the enterocytes occurs through the function of DMT1[95,96]. DMT1 is also capable of transporting other divalent cations[96–98] and is responsible for cellular iron acquisition during the transferrin receptor cycle[47–50]. DMT1 functions as a Fe(II)/proton symporter[96] and is highly expressed in the duodenum during iron deficiency[99]. This is consistent with the duodenum being the principal site of iron absorption[100] and the least alkaline section of the small intestine due to the close proximity to acidic gastric secretions. Ultimately, Fe(III) is also transported by DMT1 after reduction to Fe(II) by DcytB[101], although the physiological significance of this pathway is the subject of continued debate[102,103].
The physiological relevance of DMT1 in iron metabolism, including iron absorption, is confirmed in the Belgrade (b) rat and mk mouse which both exhibit a microcytic, hypochromic anaemia due to a G185R mutation to DMT1, resulting in a dramatic decrease in DMT1 function[47,104–106]. Considering b/b rats, the primary symptoms are mostly attributable to decreased iron uptake by reticulocytes[107,108] and earlier erythroid precursors[109]. Further research has shown that endosomal iron transport during the transferrin receptor cycle is significantly reduced in b/b rats[108,110–112], and these findings are entirely consistent with the functional role[47,105] and sub-cellular location[48,50] of DMT1 in relation to the transferrin receptor cycle.
In addition to the striking effects on reticulocyte development, b/b rats also exhibit a significant decrease in the quantity of megakaryocytes in their bone marrow[113], and their overall hematological status is similar to that observed in a rare preleukaemic syndrome[114]. The subsequent high clearance rates of prematurely senescent erythrocytes in turn causes splenomegaly. Aside from hematological factors, b/b rats exhibit a universal reduction in iron uptake by body tissues[115], including the brain[116]. The extent to which this affects overall health and development, independent of hematological parameters, is not currently known.
The final important aspect of defective iron metabolism by Belgrade rats is their decreased dietary non-heme iron absorption at the stage of uptake into the enterocytes[115,117]. This is consistent with the location[99] and function[95] of DMT1 on the microvillus membrane of enterocytes. Decreased iron absorption can be viewed as the ‘second hit’ for iron deficiency anaemia in b/b rats. However, the residual activity of mutated DMT1 means that appropriate dietary iron supplementation and iron dextran injections can improve general health and survival rates without returning hematological status to normal[118].
Over the past few years our laboratory has been using the Belgrade rat to examine whether DMT1 is the transporter required to utilise the iron released from heme during receptor mediated endocytosis. This hypothesis suggests that the function of DMT1 is directly comparable to its role in the transferrin receptor cycle[47–50] and that b/b rats would exhibit decreased heme iron absorption compared with +/b and control rats due to a reduced ability to transport iron out of internalised vesicles.
THE BELGRADE RAT AND HEME IRON ABSORPTION
In preliminary experiments, b/b rats exhibited a statistically significant decrease in hemin chloride, hemoglobin, and ferrous iron absorption when compared with +/b and Wistar (Wi) rats, as assessed by whole body retention of an oral gavage of radio-labelled iron. This strongly suggested that there was a requirement for DMT1 in heme iron absorption (see Expt 1 in Figure 2A). However, it was unclear whether this effect was due to a reduction of DMT1 function across the membranes of endocytotic vesicles, or was secondary to pathological effects of the iron deficiency and anaemia experienced by b/b rats. A general defect in the mucosal biology of b/b rats may have consequences for other proteins and biosynthetic processes including reduced expression of the heme receptor and/or heme transporters, reduced capacity to internalise a heme/receptor complex and reduced heme oxygenase activity -all of which would affect heme iron absorption.
Figure 2.
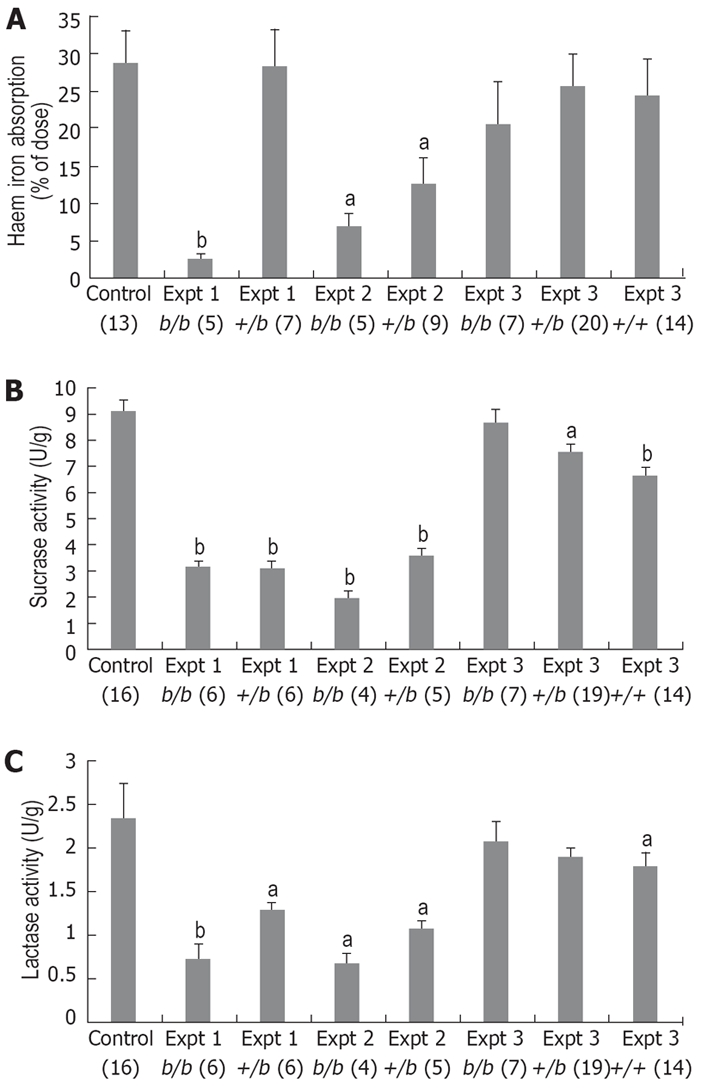
Results from Belgrade rats for heme iron absorption (A), sucrase activity (B) and lactase activity (C) over a series of experiments. Figures in brackets indicate n values, and data is mean ± SE. Groups marked with ‘a’ or ‘b’ are significantly different from the control group (1-way ANOVA aP < 0.05 and bP < 0.005, respectively). In early experiments 1 and 2, b/b and +/b rats had significantly lower heme iron absorption than Wi controls, initially suggesting a possible role for DMT1. However, sucrase and lactase activity was also lower in b/b and +/b rats indicating a more general defect in the mucosa of the Belgrade strain. In experiment 3, +/+ rats were used as an improved control to account for strain differences between Wi and Belgrade strains, but in this (and subsequent) experiments there was an apparent recovery in both heme iron absorption as well as sucrase and lactase activities. We could find no explanation for this dramatic phenotypic change, making it difficult to correlate DMT1 function with heme iron absorption.
It was observed that in addition to reduced heme iron absorption both b/b and +/b rats also exhibited significantly lower activity of sucrase and lactase[119] when compared with Wistar rats (see Expt 1 in Figure 2B, C). Previous studies examining the Belgrade strain have only observed phenotypic differences in b/b rats, whereas +/b rats appear identical to control laboratory rat strains such as Wistar and Sprague-Dawley for all measured parameters[115,117,120,121]. This raised the strong possibility that there are as yet uncharacterized strain differences between Belgrade and other laboratory rat strains that are unrelated to reticulocyte iron uptake and non-heme iron absorption.
Experiments assessing heme iron absorption and the expression of sucrase and lactase were repeated shortly after with similar results; b/b rats again exhibited significantly decreased heme iron absorption compared to Wi, but so did +/b rats contrasting with the previous experiment. Further, both b/b and +/b rats had significantly decreased sucrase and lactase activities compared to Wi controls, making it difficult to correlate DMT1 function with heme iron absorption (see Expt 2 in Figure 2A-C). At this time it was decided to use a more appropriate control than Wi rats in order to account for any strain differences, thus we established a breeding program that would generate b/b, +/b and +/+ Belgrade rats. We concurrently began genotyping all experimental rats by sequencing reverse transcribed DMT1 mRNA; in all cases the measured genotype matched the observed phenotype.
The experiment including +/+ rats yielded unexpected and conflicting results. b/b rats demonstrated a dramatic recovery of both heme iron absorption and sucrase and lactase activity, such that they were directly equivalent to +/b and +/+ rats, and approximately comparable to Wi controls from previous experiments (see Expt 3 in Figure 2A-C). Subsequent experiments were able to replicate this recovery (data not shown). The dramatic phenotypic change in the Belgrade rat strain could not be attributable to any gross developmental characteristics including body mass, spleen mass and mucosa mass, nor any change in the measures of iron deficiency and anaemia. Additionally, we were unable to find evidence for the contraction of a chronic infection, significant changes in other external factors or any variation in our well-controlled measurement of heme iron absorption and enzyme activity.
Thus the colony of Belgrade rats available to us has been unable to provide any conclusive evidence for a role of DMT1 in heme iron absorption. Similarly, we were not able to observe any relocation of DMT1 to endosomes or lysosomes during the course of heme iron absorption using confocal microscopy[94]. However, bearing in mind that there are still large deficiencies in the understanding of heme iron absorption, we feel that a potential role for DMT1 should still be seriously considered in the future. This will require more reliable animal models than the Belgrade rat, possibly including the mk mouse or the Slc11a2int/int mouse which exhibits selective knockout of intestinal DMT1[95]. Further, it would be highly desirable to use more sensitive techniques than confocal microscopy (such as immunoelectron microscopy) to determine whether DMT1 is internalised to endosomes or lysosomes during heme iron absorption.
CONCLUSION
Since the discovery of DMT1 in 1997 iron metabolism has experienced a renaissance, and the subsequent discovery of ferroportin and the humoral regulator hepcidin have provided a very solid and complete theory for the mechanisms of non-heme iron absorption. The absorption of heme iron has yet to undergo a similar revolution, and considerably more work needs to be done in the future for a complete understanding of the absorption of this critical micronutrient. Primarily, the physiological relevance of recently discovered heme transporters will need to be confirmed in vivo, along with continued searching for additional heme transporters and receptors. Concurrent efforts to explicitly identify the enzyme responsible for heme catabolism, as well as the sub-cellular location of this enzyme’s catalytic site, will help ascertain whether heme is taken up by receptor mediated endocytosis or is transported directly into the cytoplasm of enterocytes. Finally, the potential role of iron transporters such as DMT1 in heme iron absorption will need to be investigated further; particularly if receptor mediated endocytosis proves to be an important heme uptake pathway.
Peer reviewers: Trevor Redgrave, Professor, Department of Physiology, School of Biomedical and Chemical Sciences, The University of Western Australia, Perth, WA, Australia; Dr. Nathan Subramaniam, Membrane Transport Laboratory, The Queensland Institute of Medical Research 300 Herston Road, Herston, Brisbane, QLD 4006, Australia; Paul E Sijens, PhD, Associate Professor, Department of Radiology, UMCG, Groningen 9713GZ, Netherlands; Serhan Karvar, MD, Assistant Professor of Medicine, University of Southern California, Keck School of Medicine, Division of Gastrointestinal & Liver Diseases, 2011 Zonal Avenue, HMR 101, Los Angeles, CA 90089, United States
S- Editor Zhong XY L- Editor Mihm S E- Editor Lin YP
References
- 1.Lieu PT, Heiskala M, Peterson PA, Yang Y. The roles of iron in health and disease. Mol Aspects Med. 2001;22:1–87. doi: 10.1016/s0098-2997(00)00006-6. [DOI] [PubMed] [Google Scholar]
- 2.Assessing the iron status of populations. Report of a Joint World Health Organization/Centers for Disease Control and Prevention Technical Consultation on the Assessment of Iron Status at the Population Level [online], 6-8 April 2004 [cited 14 June 2006], Geneva, Switzerland. Available from URL: http://whqlibdoc.who.int/publications/2004/9241593156_eng.pdf.
- 3.Walsh RJ, Kaldor I, Brading I, George EP. The availability of iron in meat: some experiments with radioactive iron. Australas Ann Med. 1955;4:272–276. doi: 10.1111/imj.1955.4.4.272. [DOI] [PubMed] [Google Scholar]
- 4.Narasinga BS. Physiology of iron absorption and supplementation. Br Med Bull. 1981;37:25–30. doi: 10.1093/oxfordjournals.bmb.a071671. [DOI] [PubMed] [Google Scholar]
- 5.Bezwoda WR, Bothwell TH, Charlton RW, Torrance JD, MacPhail AP, Derman DP, Mayet F. The relative dietary importance of haem and non-haem iron. S Afr Med J. 1983;64:552–556. [PubMed] [Google Scholar]
- 6.Carpenter CE, Mahoney AW. Contributions of heme and nonheme iron to human nutrition. Crit Rev Food Sci Nutr. 1992;31:333–367. doi: 10.1080/10408399209527576. [DOI] [PubMed] [Google Scholar]
- 7.Gibson S, Ashwell M. The association between red and processed meat consumption and iron intakes and status among British adults. Public Health Nutr. 2003;6:341–350. doi: 10.1079/PHN2002442. [DOI] [PubMed] [Google Scholar]
- 8.Spiro GT, Page L, Saltman P. The hydrolytic polymerisation of ferric citrate. I. The chemistry of the polymer. J Am Chem Soc. 1967;89:5555–5559. [Google Scholar]
- 9.Hallberg L, Solvell L. Absorption of a single dose of iron in man. Acta Med Scand Suppl. 1960;358:19–42. doi: 10.1111/j.0954-6820.1960.tb15571.x. [DOI] [PubMed] [Google Scholar]
- 10.Manis JG, Schachter D. Active transport of iron by intestine: effects of oral iron and pregnancy. Am J Physiol. 1962;203:81–86. doi: 10.1152/ajplegacy.1962.203.1.81. [DOI] [PubMed] [Google Scholar]
- 11.de Alarcon PA, Donovan ME, Forbes GB, Landaw SA, Stockman JA 3rd. Iron absorption in the thalassemia syndromes and its inhibition by tea. N Engl J Med. 1979;300:5–8. doi: 10.1056/NEJM197901043000102. [DOI] [PubMed] [Google Scholar]
- 12.Moore CV, Arrowsmith WR, Welch J, Minnich V. Studies In Iron Transportation And Metabolism. Iv. Observations On The Absorption Of Iron From The Gastro-Intestinal Tract. J Clin Invest. 1939;18:553–580. doi: 10.1172/JCI101069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Conrad ME, Cortell S, Williams HL, Foy AL. Polymerization and intraluminal factors in the absorption of hemoglobin-iron. J Lab Clin Med. 1966;68:659–668. [PubMed] [Google Scholar]
- 14.Turnbull A, Cleton F, Finch CA. Iron absorption. IV. The absorption of hemoglobin iron. J Clin Invest. 1962;41:1897–1907. doi: 10.1172/JCI104646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hallberg L, Solvell L. Absorption of hemoglobin iron in man. Acta Med Scand. 1967;181:335–354. doi: 10.1111/j.0954-6820.1967.tb15161.x. [DOI] [PubMed] [Google Scholar]
- 16.Hwang YF, Brown EB. Effect of desferrioxamine on iron absorption. Lancet. 1965;1:135–137. doi: 10.1016/s0140-6736(65)91093-7. [DOI] [PubMed] [Google Scholar]
- 17.Wheby MS, Suttle GE, Ford KT 3rd. Intestinal absorption of hemoglobin iron. Gastroenterology. 1970;58:647–654. [PubMed] [Google Scholar]
- 18.Hallberg L, Bjorn-Rasmussen E, Howard L, Rossander L. Dietary heme iron absorption. A discussion of possible mechanisms for the absorption-promoting effect of meat and for the regulation of iron absorption. Scand J Gastroenterol. 1979;14:769–779. doi: 10.3109/00365527909181403. [DOI] [PubMed] [Google Scholar]
- 19.Bjorn-Rasmussen E, Hallberg L, Isaksson B, Arvidsson B. Food iron absorption in man. Applications of the two-pool extrinsic tag method to measure heme and nonheme iron absorption from the whole diet. J Clin Invest. 1974;53:247–255. doi: 10.1172/JCI107545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vaghefi N, Nedjaoum F, Guillochon D, Bureau F, Arhan P, Bougle D. Influence of the extent of hemoglobin hydrolysis on the digestive absorption of heme iron. An in vitro study. J Agric Food Chem. 2002;50:4969–4973. doi: 10.1021/jf0109165. [DOI] [PubMed] [Google Scholar]
- 21.Callender ST, Mallett BJ, Smith MD. Absorption of haemoglobin iron. Br J Haematol. 1957;3:186–192. doi: 10.1111/j.1365-2141.1957.tb05785.x. [DOI] [PubMed] [Google Scholar]
- 22.Bannerman RM. Quantitative Aspects Of Hemoglobin-Iron Absorption. J Lab Clin Med. 1965;65:944–950. [PubMed] [Google Scholar]
- 23.Conrad ME, Weintraub LR, Sears DA, Crosby WH. Absorption of hemoglobin iron. Am J Physiol. 1966;211:1123–1130. doi: 10.1152/ajplegacy.1966.211.5.1123. [DOI] [PubMed] [Google Scholar]
- 24.Raffin SB, Woo CH, Roost KT, Price DC, Schmid R. Intestinal absorption of hemoglobin iron-heme cleavage by mucosal heme oxygenase. J Clin Invest. 1974;54:1344–1352. doi: 10.1172/JCI107881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bjorn-Rasmussen E. Iron absorption: present knowledge and controversies. Lancet. 1983;1:914–916. doi: 10.1016/s0140-6736(83)91337-5. [DOI] [PubMed] [Google Scholar]
- 26.Abboud S, Haile DJ. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J Biol Chem. 2000;275:19906–19912. doi: 10.1074/jbc.M000713200. [DOI] [PubMed] [Google Scholar]
- 27.Donovan A, Brownlie A, Zhou Y, Shepard J, Pratt SJ, Moynihan J, Paw BH, Drejer A, Barut B, Zapata A, et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature. 2000;403:776–781. doi: 10.1038/35001596. [DOI] [PubMed] [Google Scholar]
- 28.McKie AT, Marciani P, Rolfs A, Brennan K, Wehr K, Barrow D, Miret S, Bomford A, Peters TJ, Farzaneh F, et al. A novel duodenal iron-regulated transporter, IREG1, implicated in the basolateral transfer of iron to the circulation. Mol Cell. 2000;5:299–309. doi: 10.1016/s1097-2765(00)80425-6. [DOI] [PubMed] [Google Scholar]
- 29.Donovan A, Lima CA, Pinkus JL, Pinkus GS, Zon LI, Robine S, Andrews NC. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005;1:191–200. doi: 10.1016/j.cmet.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 30.Pigeon C, Ilyin G, Courselaud B, Leroyer P, Turlin B, Brissot P, Loreal O. A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload. J Biol Chem. 2001;276:7811–7819. doi: 10.1074/jbc.M008923200. [DOI] [PubMed] [Google Scholar]
- 31.Frazer DM, Wilkins SJ, Becker EM, Vulpe CD, McKie AT, Trinder D, Anderson GJ. Hepcidin expression inversely correlates with the expression of duodenal iron transporters and iron absorption in rats. Gastroenterology. 2002;123:835–844. doi: 10.1053/gast.2002.35353. [DOI] [PubMed] [Google Scholar]
- 32.Ludwiczek S, Theurl I, Artner-Dworzak E, Chorney M, Weiss G. Duodenal HFE expression and hepcidin levels determine body iron homeostasis: modulation by genetic diversity and dietary iron availability. J Mol Med. 2004;82:373–382. doi: 10.1007/s00109-004-0542-3. [DOI] [PubMed] [Google Scholar]
- 33.Nemeth E, Tuttle MS, Powelson J, Vaughn MB, Donovan A, Ward DM, Ganz T, Kaplan J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science. 2004;306:2090–2093. doi: 10.1126/science.1104742. [DOI] [PubMed] [Google Scholar]
- 34.Conrad ME, Benjamin BI, Williams HL, Foy AL. Human absorption of hemoglobin-iron. Gastroenterology. 1967;53:5–10. [PubMed] [Google Scholar]
- 35.Brown EB, Hwang YF, Nicol S, Ternberg J. Absorption of radiation-labeled hemoglobin by dogs. J Lab Clin Med. 1968;72:58–64. [PubMed] [Google Scholar]
- 36.Roberts SK, Henderson RW, Young GP. Modulation of uptake of heme by rat small intestinal mucosa in iron deficiency. Am J Physiol. 1993;265:G712–G718. doi: 10.1152/ajpgi.1993.265.4.G712. [DOI] [PubMed] [Google Scholar]
- 37.Grasbeck R, Kouvonen I, Lundberg M, Tenhunen R. An intestinal receptor for heme. Scand J Haematol. 1979;23:5–9. doi: 10.1111/j.1600-0609.1979.tb02845.x. [DOI] [PubMed] [Google Scholar]
- 38.Tenhunen R, Grasbeck R, Kouvonen I, Lundberg M. An intestinal receptor for heme: its parital characterization. Int J Biochem. 1980;12:713–716. doi: 10.1016/0020-711x(80)90149-4. [DOI] [PubMed] [Google Scholar]
- 39.Grasbeck R, Majuri R, Kouvonen I, Tenhunen R. Spectral and other studies on the intestinal haem receptor of the pig. Biochim Biophys Acta. 1982;700:137–142. doi: 10.1016/0167-4838(82)90089-9. [DOI] [PubMed] [Google Scholar]
- 40.Galbraith RA, Sassa S, Kappas A. Heme binding to murine erythroleukemia cells. Evidence for a heme receptor. J Biol Chem. 1985;260:12198–12202. [PubMed] [Google Scholar]
- 41.Majuri R, Grasbeck R. A rosette receptor assay with haem-microbeads. Demonstration of a haem receptor on K562 cells. Eur J Haematol. 1987;38:21–25. doi: 10.1111/j.1600-0609.1987.tb01418.x. [DOI] [PubMed] [Google Scholar]
- 42.Majuri R. Heme-binding plasma membrane proteins of K562 erythroleukemia cells: adsorption to heme-microbeads, isolation with affinity chromatography. Eur J Haematol. 1989;43:220–225. doi: 10.1111/j.1600-0609.1989.tb00286.x. [DOI] [PubMed] [Google Scholar]
- 43.Worthington MT, Cohn SM, Miller SK, Luo RQ, Berg CL. Characterization of a human plasma membrane heme transporter in intestinal and hepatocyte cell lines. Am J Physiol Gastrointest Liver Physiol. 2001;280:G1172–G1177. doi: 10.1152/ajpgi.2001.280.6.G1172. [DOI] [PubMed] [Google Scholar]
- 44.Vaghefi N, Guillochon D, Bureau F, Neuville D, Lebrun F, Arhan P, Bougle D. The effect of cysteine and 2,4-dinitrophenol on heme and nonheme absorption in a rat intestinal model. J Nutr Biochem. 2000;11:562–567. doi: 10.1016/s0955-2863(00)00120-0. [DOI] [PubMed] [Google Scholar]
- 45.Parmley RT, Barton JC, Conrad ME, Austin RL, Holland RM. Ultrastructural cytochemistry and radioautography of hemoglobin--iron absorption. Exp Mol Pathol. 1981;34:131–144. doi: 10.1016/0014-4800(81)90070-8. [DOI] [PubMed] [Google Scholar]
- 46.Wyllie JC, Kaufman N. An electron microscopic study of heme uptake by rat duodenum. Lab Invest. 1982;47:471–476. [PubMed] [Google Scholar]
- 47.Su MA, Trenor CC, Fleming JC, Fleming MD, Andrews NC. The G185R mutation disrupts function of the iron transporter Nramp2. Blood. 1998;92:2157–2163. [PubMed] [Google Scholar]
- 48.Gruenheid S, Canonne-Hergaux F, Gauthier S, Hackam DJ, Grinstein S, Gros P. The iron transport protein NRAMP2 is an integral membrane glycoprotein that colocalizes with transferrin in recycling endosomes. J Exp Med. 1999;189:831–841. doi: 10.1084/jem.189.5.831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ponka P, Lok CN. The transferrin receptor: role in health and disease. Int J Biochem Cell Biol. 1999;31:1111–1137. doi: 10.1016/s1357-2725(99)00070-9. [DOI] [PubMed] [Google Scholar]
- 50.Tabuchi M, Yoshimori T, Yamaguchi K, Yoshida T, Kishi F. Human NRAMP2/DMT1, which mediates iron transport across endosomal membranes, is localized to late endosomes and lysosomes in HEp-2 cells. J Biol Chem. 2000;275:22220–22228. doi: 10.1074/jbc.M001478200. [DOI] [PubMed] [Google Scholar]
- 51.Shayeghi M, Latunde-Dada GO, Oakhill JS, Laftah AH, Takeuchi K, Halliday N, Khan Y, Warley A, McCann FE, Hider RC, et al. Identification of an intestinal heme transporter. Cell. 2005;122:789–801. doi: 10.1016/j.cell.2005.06.025. [DOI] [PubMed] [Google Scholar]
- 52.Qiu A, Jansen M, Sakaris A, Min SH, Chattopadhyay S, Tsai E, Sandoval C, Zhao R, Akabas MH, Goldman ID. Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption. Cell. 2006;127:917–928. doi: 10.1016/j.cell.2006.09.041. [DOI] [PubMed] [Google Scholar]
- 53.Quigley JG, Yang Z, Worthington MT, Phillips JD, Sabo KM, Sabath DE, Berg CL, Sassa S, Wood BL, Abkowitz JL. Identification of a human heme exporter that is essential for erythropoiesis. Cell. 2004;118:757–766. doi: 10.1016/j.cell.2004.08.014. [DOI] [PubMed] [Google Scholar]
- 54.Inoue K, Nakai Y, Ueda S, Kamigaso S, Ohta KY, Hatakeyama M, Hayashi Y, Otagiri M, Yuasa H. Functional characterization of PCFT/HCP1 as the molecular entity of the carrier-mediated intestinal folate transport system in the rat model. Am J Physiol Gastrointest Liver Physiol. 2008;294:G660–G668. doi: 10.1152/ajpgi.00309.2007. [DOI] [PubMed] [Google Scholar]
- 55.Tailor CS, Willett BJ, Kabat D. A putative cell surface receptor for anemia-inducing feline leukemia virus subgroup C is a member of a transporter superfamily. J Virol. 1999;73:6500–6505. doi: 10.1128/jvi.73.8.6500-6505.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Quigley JG, Burns CC, Anderson MM, Lynch ED, Sabo KM, Overbaugh J, Abkowitz JL. Cloning of the cellular receptor for feline leukemia virus subgroup C (FeLV-C), a retrovirus that induces red cell aplasia. Blood. 2000;95:1093–1099. [PubMed] [Google Scholar]
- 57.Uc A, Stokes JB, Britigan BE. Heme transport exhibits polarity in Caco-2 cells: evidence for an active and membrane protein-mediated process. Am J Physiol Gastrointest Liver Physiol. 2004;287:G1150–G1157. doi: 10.1152/ajpgi.00157.2004. [DOI] [PubMed] [Google Scholar]
- 58.Hartmann F, Owen R, Bissell DM. Characterization of isolated epithelial cells from rat small intestine. Am J Physiol. 1982;242:G147–G155. doi: 10.1152/ajpgi.1982.242.2.G147. [DOI] [PubMed] [Google Scholar]
- 59.Ferris CD, Jaffrey SR, Sawa A, Takahashi M, Brady SD, Barrow RK, Tysoe SA, Wolosker H, Baranano DE, Dore S, et al. Haem oxygenase-1 prevents cell death by regulating cellular iron. Nat Cell Biol. 1999;1:152–157. doi: 10.1038/11072. [DOI] [PubMed] [Google Scholar]
- 60.Uc A, Britigan BE. Does heme oxygenase-1 have a role in Caco-2 cell cycle progression? Exp Biol Med (Maywood) 2003;228:590–595. doi: 10.1177/15353702-0322805-52. [DOI] [PubMed] [Google Scholar]
- 61.Weintraub LR, Weinstein MB, Huser HJ, Rafal S. Absorption of hemoglobin iron: the role of a heme-splitting substance in the intestinal mucosa. J Clin Invest. 1968;47:531–539. doi: 10.1172/JCI105749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mills GC. Hemoglobin catabolism. III. Conversion of hemoglobin to choleglobin by rat liver preparations. J Biochem. 1962;51:41–47. doi: 10.1093/oxfordjournals.jbchem.a127498. [DOI] [PubMed] [Google Scholar]
- 63.Awai M, Brown EB. Examination of the role of xanthine oxidase in iron absorption by the rat. J Lab Clin Med. 1969;73:366–378. [PubMed] [Google Scholar]
- 64.Dawson RB, Rafal S, Weintraub LR. Absorption of hemoglobin iron: the role of xanthine oxidase in the intestinal heme-splitting reaction. Blood. 1970;35:94–103. [PubMed] [Google Scholar]
- 65.Gray CH, Nicholson DC, Tipton G. Degradation of haem compounds to bile pigments. Nat New Biol. 1972;239:5–8. doi: 10.1038/newbio239005a0. [DOI] [PubMed] [Google Scholar]
- 66.Bonnett R, McDonagh AF. The meso-reactivity of porphyrins and related compounds. VI. Oxidative cleavage of the haem system. The four isomeric biliverdins of the IX series. J Chem Soc Perkin 1. 1973;9:881–888. doi: 10.1039/p19730000881. [DOI] [PubMed] [Google Scholar]
- 67.Wheby MS, Spyker DA. Hemoglobin iron absorption kinetics in the iron-deficient dog. Am J Clin Nutr. 1981;34:1686–1693. doi: 10.1093/ajcn/34.9.1686. [DOI] [PubMed] [Google Scholar]
- 68.Boni RE, Huch Boni RA, Galbraith RA, Drummond GS, Kappas A. Tin-mesoporphyrin inhibits heme oxygenase activity and heme-iron absorption in the intestine. Pharmacology. 1993;47:318–329. doi: 10.1159/000139113. [DOI] [PubMed] [Google Scholar]
- 69.Tenhunen R, Marver HS, Schmid R. Microsomal heme oxygenase. Characterization of the enzyme. J Biol Chem. 1969;244:6388–6394. [PubMed] [Google Scholar]
- 70.Tenhunen R, Marver HS, Schmid R. The enzymatic catabolism of hemoglobin: stimulation of microsomal heme oxygenase by hemin. J Lab Clin Med. 1970;75:410–421. [PubMed] [Google Scholar]
- 71.Tenhunen R, Marver H, Pimstone NR, Trager WF, Cooper DY, Schmid R. Enzymatic degradation of heme. Oxygenative cleavage requiring cytochrome P-450. Biochemistry. 1972;11:1716–1720. doi: 10.1021/bi00759a029. [DOI] [PubMed] [Google Scholar]
- 72.Maines MD, Trakshel GM, Kutty RK. Characterization of two constitutive forms of rat liver microsomal heme oxygenase. Only one molecular species of the enzyme is inducible. J Biol Chem. 1986;261:411–419. [PubMed] [Google Scholar]
- 73.Trakshel GM, Maines MD. Multiplicity of heme oxygenase isozymes. HO-1 and HO-2 are different molecular species in rat and rabbit. J Biol Chem. 1989;264:1323–1328. [PubMed] [Google Scholar]
- 74.Cruse I, Maines MD. Evidence suggesting that the two forms of heme oxygenase are products of different genes. J Biol Chem. 1988;263:3348–3353. [PubMed] [Google Scholar]
- 75.Rotenberg MO, Maines MD. Isolation, characterization, and expression in Escherichia coli of a cDNA encoding rat heme oxygenase-2. J Biol Chem. 1990;265:7501–7506. [PubMed] [Google Scholar]
- 76.McCoubrey WK Jr, Huang TJ, Maines MD. Isolation and characterization of a cDNA from the rat brain that encodes hemoprotein heme oxygenase-3. Eur J Biochem. 1997;247:725–732. doi: 10.1111/j.1432-1033.1997.00725.x. [DOI] [PubMed] [Google Scholar]
- 77.Hayashi S, Omata Y, Sakamoto H, Higashimoto Y, Hara T, Sagara Y, Noguchi M. Characterization of rat heme oxygenase-3 gene. Implication of processed pseudogenes derived from heme oxygenase-2 gene. Gene. 2004;336:241–250. doi: 10.1016/j.gene.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 78.Oates PS, West AR. Heme in intestinal epithelial cell turnover, differentiation, detoxification, inflammation, carcinogenesis, absorption and motility. World J Gastroenterol. 2006;12:4281–4295. doi: 10.3748/wjg.v12.i27.4281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ewing JF, Maines MD. Rapid induction of heme oxygenase 1 mRNA and protein by hyperthermia in rat brain: heme oxygenase 2 is not a heat shock protein. Proc Natl Acad Sci USA. 1991;88:5364–5368. doi: 10.1073/pnas.88.12.5364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Stocker R, Yamamoto Y, McDonagh AF, Glazer AN, Ames BN. Bilirubin is an antioxidant of possible physiological importance. Science. 1987;235:1043–1046. doi: 10.1126/science.3029864. [DOI] [PubMed] [Google Scholar]
- 81.Poss KD, Tonegawa S. Heme oxygenase 1 is required for mammalian iron reutilization. Proc Natl Acad Sci USA. 1997;94:10919–10924. doi: 10.1073/pnas.94.20.10919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Yachie A, Niida Y, Wada T, Igarashi N, Kaneda H, Toma T, Ohta K, Kasahara Y, Koizumi S. Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. J Clin Invest. 1999;103:129–135. doi: 10.1172/JCI4165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Yamada N, Yamaya M, Okinaga S, Nakayama K, Sekizawa K, Shibahara S, Sasaki H. Microsatellite polymorphism in the heme oxygenase-1 gene promoter is associated with susceptibility to emphysema. Am J Hum Genet. 2000;66:187–195. doi: 10.1086/302729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Chen YH, Lin SJ, Lin MW, Tsai HL, Kuo SS, Chen JW, Charng MJ, Wu TC, Chen LC, Ding YA, et al. Microsatellite polymorphism in promoter of heme oxygenase-1 gene is associated with susceptibility to coronary artery disease in type 2 diabetic patients. Hum Genet. 2002;111:1–8. doi: 10.1007/s00439-002-0769-4. [DOI] [PubMed] [Google Scholar]
- 85.Trakshel GM, Kutty RK, Maines MD. Purification and characterization of the major constitutive form of testicular heme oxygenase. The noninducible isoform. J Biol Chem. 1986;261:11131–11137. [PubMed] [Google Scholar]
- 86.Sun Y, Rotenberg MO, Maines MD. Developmental expression of heme oxygenase isozymes in rat brain. Two HO-2 mRNAs are detected. J Biol Chem. 1990;265:8212–8217. [PubMed] [Google Scholar]
- 87.Ding Y, McCoubrey WK Jr, Maines MD. Interaction of heme oxygenase-2 with nitric oxide donors. Is the oxygenase an intracellular 'sink' for NO? Eur J Biochem. 1999;264:854–861. doi: 10.1046/j.1432-1327.1999.00677.x. [DOI] [PubMed] [Google Scholar]
- 88.Adachi T, Ishikawa K, Hida W, Matsumoto H, Masuda T, Date F, Ogawa K, Takeda K, Furuyama K, Zhang Y, et al. Hypoxemia and blunted hypoxic ventilatory responses in mice lacking heme oxygenase-2. Biochem Biophys Res Commun. 2004;320:514–522. doi: 10.1016/j.bbrc.2004.05.195. [DOI] [PubMed] [Google Scholar]
- 89.Shrestha Dangol D, Chen HP. Role of hemeoxygenase-2 in pregnancy-induced hypertension. Int J Gynaecol Obstet. 2004;85:44–46. doi: 10.1016/S0020-7292(03)00196-6. [DOI] [PubMed] [Google Scholar]
- 90.Williams SE, Wootton P, Mason HS, Bould J, Iles DE, Riccardi D, Peers C, Kemp PJ. Hemoxygenase-2 is an oxygen sensor for a calcium-sensitive potassium channel. Science. 2004;306:2093–2097. doi: 10.1126/science.1105010. [DOI] [PubMed] [Google Scholar]
- 91.Latunde-Dada GO, Simpson RJ, McKie AT. Recent advances in mammalian haem transport. Trends Biochem Sci. 2006;31:182–188. doi: 10.1016/j.tibs.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 92.Collins JF, Franck CA, Kowdley KV, Ghishan FK. Identification of differentially expressed genes in response to dietary iron deprivation in rat duodenum. Am J Physiol Gastrointest Liver Physiol. 2005;288:G964–G971. doi: 10.1152/ajpgi.00489.2004. [DOI] [PubMed] [Google Scholar]
- 93.Baranano DE, Wolosker H, Bae BI, Barrow RK, Snyder SH, Ferris CD. A mammalian iron ATPase induced by iron. J Biol Chem. 2000;275:15166–15173. doi: 10.1074/jbc.275.20.15166. [DOI] [PubMed] [Google Scholar]
- 94.West AR, Oates PS. Subcellular location of heme oxygenase 1 and 2 and divalent metal transporter 1 in relation to endocytotic markers during heme iron absorption. J Gastroenterol Hepatol. 2008;23:150–158. doi: 10.1111/j.1440-1746.2007.05047.x. [DOI] [PubMed] [Google Scholar]
- 95.Gunshin H, Fujiwara Y, Custodio AO, Direnzo C, Robine S, Andrews NC. Slc11a2 is required for intestinal iron absorption and erythropoiesis but dispensable in placenta and liver. J Clin Invest. 2005;115:1258–1266. doi: 10.1172/JCI24356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Gunshin H, Mackenzie B, Berger UV, Gunshin Y, Romero MF, Boron WF, Nussberger S, Gollan JL, Hediger MA. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature. 1997;388:482–488. doi: 10.1038/41343. [DOI] [PubMed] [Google Scholar]
- 97.Forbes JR, Gros P. Iron, manganese, and cobalt transport by Nramp1 (Slc11a1) and Nramp2 (Slc11a2) expressed at the plasma membrane. Blood. 2003;102:1884–1892. doi: 10.1182/blood-2003-02-0425. [DOI] [PubMed] [Google Scholar]
- 98.Sacher A, Cohen A, Nelson N. Properties of the mammalian and yeast metal-ion transporters DCT1 and Smf1p expressed in Xenopus laevis oocytes. J Exp Biol. 2001;204:1053–1061. doi: 10.1242/jeb.204.6.1053. [DOI] [PubMed] [Google Scholar]
- 99.Trinder D, Oates PS, Thomas C, Sadleir J, Morgan EH. Localisation of divalent metal transporter 1 (DMT1) to the microvillus membrane of rat duodenal enterocytes in iron deficiency, but to hepatocytes in iron overload. Gut. 2000;46:270–276. doi: 10.1136/gut.46.2.270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Brown EB, Jr Justus BW. In vitro absorption of radioiron by everted pouches of rat intestine. Am J Physiol. 1958;194:319–326. doi: 10.1152/ajplegacy.1958.194.2.319. [DOI] [PubMed] [Google Scholar]
- 101.McKie AT, Barrow D, Latunde-Dada GO, Rolfs A, Sager G, Mudaly E, Mudaly M, Richardson C, Barlow D, Bomford A, et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science. 2001;291:1755–1759. doi: 10.1126/science.1057206. [DOI] [PubMed] [Google Scholar]
- 102.Frazer DM, Wilkins SJ, Vulpe CD, Anderson GJ. The role of duodenal cytochrome b in intestinal iron absorption remains unclear. Blood. 2005;106:4413; author reply 4414. doi: 10.1182/blood-2005-07-2923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Gunshin H, Starr CN, Direnzo C, Fleming MD, Jin J, Greer EL, Sellers VM, Galica SM, Andrews NC. Cybrd1 (duodenal cytochrome b) is not necessary for dietary iron absorption in mice. Blood. 2005;106:2879–2883. doi: 10.1182/blood-2005-02-0716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Fleming MD, Trenor CC 3rd, Su MA, Foernzler D, Beier DR, Dietrich WF, Andrews NC. Microcytic anaemia mice have a mutation in Nramp2, a candidate iron transporter gene. Nat Genet. 1997;16:383–386. doi: 10.1038/ng0897-383. [DOI] [PubMed] [Google Scholar]
- 105.Fleming MD, Romano MA, Su MA, Garrick LM, Garrick MD, Andrews NC. Nramp2 is mutated in the anemic Belgrade (b) rat: evidence of a role for Nramp2 in endosomal iron transport. Proc Natl Acad Sci USA. 1998;95:1148–1153. doi: 10.1073/pnas.95.3.1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Touret N, Martin-Orozco N, Paroutis P, Furuya W, Lam-Yuk-Tseung S, Forbes J, Gros P, Grinstein S. Molecular and cellular mechanisms underlying iron transport deficiency in microcytic anemia. Blood. 2004;104:1526–1533. doi: 10.1182/blood-2004-02-0731. [DOI] [PubMed] [Google Scholar]
- 107.Edwards JA, Garrick LM, Hoke JE. Defective iron uptake and globin synthesis by erythroid cells in the anemia of the Belgrade laboratory rat. Blood. 1978;51:347–357. [PubMed] [Google Scholar]
- 108.Edwards JA, Sullivan AL, Hoke JE. Defective delivery of iron to the developing red cell of the Belgrade laboratory rat. Blood. 1980;55:645–648. [PubMed] [Google Scholar]
- 109.Pavlovic-Kentera V, Basara N, Biljanovic-Paunovic L, Vasiljevska M, Rolovic Z. Erythroid progenitors in anemic Belgrade laboratory (b/b) rats. Exp Hematol. 1989;17:812–815. [PubMed] [Google Scholar]
- 110.Garrick MD, Gniecko K, Liu Y, Cohan DS, Garrick LM. Transferrin and the transferrin cycle in Belgrade rat reticulocytes. J Biol Chem. 1993;268:14867–14874. [PubMed] [Google Scholar]
- 111.Bowen BJ, Morgan EH. Anemia of the Belgrade rat: evidence for defective membrane transport of iron. Blood. 1987;70:38–44. [PubMed] [Google Scholar]
- 112.Edwards J, Huebers H, Kunzler C, Finch C. Iron metabolism in the Belgrade rat. Blood. 1986;67:623–628. [PubMed] [Google Scholar]
- 113.Rolovic Z, Jovanovic T, Stankovic Z, Marinkovic N. Abnormal megakaryocytopoiesis in the Belgrade laboratory rat. Blood. 1985;65:60–64. [PubMed] [Google Scholar]
- 114.Tulliez M, Testa U, Rochant H, Henri A, Vainchenker W, Touboul J, Breton-Gorius J, Dreyfus B. Reticulocytosis, hypochromia, and microcytosis: an unusual presentation of the preleukemic syndrome. Blood. 1982;59:293–299. [PubMed] [Google Scholar]
- 115.Farcich EA, Morgan EH. Diminished iron acquisition by cells and tissues of Belgrade laboratory rats. Am J Physiol. 1992;262:R220–R224. doi: 10.1152/ajpregu.1992.262.2.R220. [DOI] [PubMed] [Google Scholar]
- 116.Burdo JR, Martin J, Menzies SL, Dolan KG, Romano MA, Fletcher RJ, Garrick MD, Garrick LM, Connor JR. Cellular distribution of iron in the brain of the Belgrade rat. Neuroscience. 1999;93:1189–1196. doi: 10.1016/s0306-4522(99)00207-9. [DOI] [PubMed] [Google Scholar]
- 117.Oates PS, Morgan EH. Defective iron uptake by the duodenum of Belgrade rats fed diets of different iron contents. Am J Physiol. 1996;270:G826–G832. doi: 10.1152/ajpgi.1996.270.5.G826. [DOI] [PubMed] [Google Scholar]
- 118.Garrick M, Scott D, Walpole S, Finkelstein E, Whitbred J, Chopra S, Trivikram L, Mayes D, Rhodes D, Cabbagestalk K, et al. Iron supplementation moderates but does not cure the Belgrade anemia. Biometals. 1997;10:65–76. doi: 10.1023/a:1018370804882. [DOI] [PubMed] [Google Scholar]
- 119.West AR, Oates PS. Decreased sucrase and lactase activity in iron deficiency is accompanied by reduced gene expression and upregulation of the transcriptional repressor PDX-1. Am J Physiol Gastrointest Liver Physiol. 2005;289:G1108–G1114. doi: 10.1152/ajpgi.00195.2005. [DOI] [PubMed] [Google Scholar]
- 120.Sladic-Simic D, Martinovitch PN, Zivkovic N, Pavic D, Martinovic J, Kahn M, Ranney HM. A thalassemia-like disorder in Belgrade laboratory rats. Ann N Y Acad Sci. 1969;165:93–99. doi: 10.1111/j.1749-6632.1969.tb27779.x. [DOI] [PubMed] [Google Scholar]
- 121.Sladic-Simic D, Zivkovic N, Pavic D, Marinkovic D, Martinovic J, Martinovitch PN. Hereditary hypochromic microcytic anemia in the laboratory rat. Genetics. 1966;53:1079–1089. doi: 10.1093/genetics/53.6.1079. [DOI] [PMC free article] [PubMed] [Google Scholar]