

Summary
A key initial event in hair follicle morphogenesis is the localised thickening of the skin epithelium to form a placode, partitioning future hair follicle epithelium from interfollicular epidermis. Although many developmental signalling pathways are implicated in follicle morphogenesis, the role of epidermal growth factor (EGF) and keratinocyte growth factor (KGF, also known as FGF7) receptors are not defined. EGF receptor (EGFR) ligands have previously been shown to inhibit developing hair follicles; however, the underlying mechanisms have not been characterised. Here we show that receptors for EGF and KGF undergo marked downregulation in hair follicle placodes from multiple body sites, whereas the expression of endogenous ligands persist throughout hair follicle initiation. Using embryonic skin organ culture, we show that when skin from the sites of primary pelage and whisker follicle development is exposed to increased levels of two ectopic EGFR ligands (HBEGF and amphiregulin) and the FGFR2(IIIb) receptor ligand KGF, follicle formation is inhibited in a time- and dose-dependent manner. We then used downstream molecular markers and microarray profiling to provide evidence that, in response to KGF and EGF signalling, epidermal differentiation is promoted at the expense of hair follicle fate. We propose that hair follicle initiation in placodes requires downregulation of the two pathways in question, both of which are crucial for the ongoing development of the interfollicular epidermis. We have also uncovered a previously unrecognised role for KGF signalling in the formation of hair follicles in the mouse.
Keywords: Hair follicle, Skin organ culture, EGF, KGF, Mouse
Introduction
Hair follicle (HF) formation begins with local thickening of the epithelium to form a placode, and an associated condensation of the underlying mesenchymal cells termed the dermal condensation (DC); both structures are common to other ectodermal appendages, including feathers, teeth and mammary glands. Individual HF morphogenesis is a tightly regulated process, relying on many highly conserved signalling pathways, including Delta/Notch, Wnt/Frizzled, Hedgehog/Patched, TGFβ/BMP and FGF signalling (Millar, 2002; Schmidt-Ullrich and Paus, 2005), providing a balance of stimulatory and inhibitory influences. Knockout and transgenic animal models have revealed that active Wnt signalling is crucial for the initiation of follicular morphogenesis (Andl et al., 2002; Huelsken et al., 2001; van Genderen et al., 1994; Zhang et al., 2008), whereas studies in mice lacking sonic hedgehog (SHH) indicate that SHH is required after HF initiation, downgrowth of the follicle epithelium and dermal papilla (DP) formation (Chiang et al., 1999; Karlsson et al., 1999; St-Jacques et al., 1998). In addition to individual follicle morphogenesis, the processes that determine whether surface epithelial cells become interfollicular skin epidermis or HFs also establish the spatial distribution of these appendages. In this context, ectodysplasin A (EDA) and its receptor EDAR appear to play a crucial role (Headon et al., 2001; Headon and Overbeek, 1999; Laurikkala et al., 2002). EDA and EDAR interact with members of the bone morphogenetic protein (BMP) family, some of which are inhibitory to follicle development, to establish follicle patterning (Mou et al., 2006; Pummila et al., 2007). In the earliest stages of follicle initiation, there has been an emphasis on determining the molecular factors that distinguish the placode versus the interfollicular epidermis (Nowak et al., 2008; Rhee et al., 2006). Two pathways that have an essential role in these cell fate decisions include EGF and KGF (FGF7 - Mouse Genome Informatics) (Beer et al., 2000; du Cros, 1993; Peus and Pittelkow, 1996; Schneider et al., 2008, Guo et al., 1996). Disruption or blocking of the EGF receptor (EGFR) in several mouse models results in abnormalities occurring late on in follicle development and during the adult hair follicle cycle, leading to the hypothesis that EGF signalling has an important positive influence on follicle development and growth (Miettinen et al., 1995; Murillas et al., 1995). EGFR also has several other activating ligands, including transforming growth factor-α (TGFα), heparin-binding EGF-like growth factor (HBEGF), amphiregulin (AR, AREG - Mouse Genome Informatics), betacellulin (BTC), epiregulin (EPR, EREG - Mouse Genome Informatics) and epigen (EPGN). In mice that lack specific ligands, such as TGFα, the absence of an HF phenotype has been interpreted as functional redundancy among ligands (Mann et al., 1993). By contrast, other work suggests that active EGF signalling can inhibit HF development (Cohen and Elliott, 1963). In skin organ culture, administration of EGF ligands (EGF and TGFα) dramatically inhibits hair morphogenesis in E13.5 mouse skin (Kashiwagi et al., 1997). However, although these two ligands are potent inhibitors of HF morphogenesis, they are not endogenously expressed in the developing skin during follicular morphogenesis. Our previous microarray analysis (Bazzi et al., 2007a) indicated that amphiregulin is present specifically in the epidermis, and its level of expression increases around the onset of follicle initiation. This supports earlier reports of alternate EGFR ligands, including HBEGF and amphiregulin, in skin at this time (Kashiwagi et al., 1997), and suggests that EGFR ligands are expressed dynamically in this developmental window.
Evidence that KGF signalling is functionally important for HF development has come largely from mouse models, in which knockout of the FGFR2(IIIb) receptor results in reduced HF density and retarded follicle development (Petiot et al., 2003). Additionally, overexpression of dominant negative FGFR2 also retards follicle development (Werner et al., 1994), and removal of KGF ligand has a modest effect on hair fibre texture (Guo et al., 1996). As with EGF, reports on the effects of exposure of developing follicles to excess ligand are stage-dependent. When recombinant KGF is injected subcutaneously into adult athymic nude mice over 17 to 18 days, it induces dose-dependent hair growth (Danilenko et al., 1995). However, overexpression of KGF in the basal epidermal and outer root sheath (ORS) keratinocytes via the K14 promoter blocks follicle development (Guo et al., 1993). One interpretation is that the effects of KGF on HF morphogenesis are likely to be dependent on the dose, time and site of KGF production in skin (Botchkareva et al., 1999). A similar hypothesis has been suggested for EGF signalling, whereby the fate of interfollicular versus follicular epithelium at the onset of follicle morphogenesis is determined partly by the levels of exogenous EGFR ligands (Kashiwagi et al., 1997).
In this study, we revisited the role of EGFR and FGFR signalling during normal HF morphogenesis. Initially, we defined the kinetics of expression of the two receptors, both of which were found to be universally downregulated in placodes of all follicle types examined. We identified endogenous ligands associated with EGF and FGF signalling in dermis and epidermis at the start of HF morphogenesis. We found that constitutive activation of both EGF and KGF signalling in the epidermis led to follicle inhibition in a process that was dose dependent but that did not alter the patterning of residual follicles. To probe the molecular events involved in this inhibition, separated epidermis and dermis from treated organ-cultured skins were analysed by whole genome microarray. The resulting data, supported by in situ hybridisation and immunohistochemistry, produced a molecular profile that confirmed the promotion of interfollicular fate at the expense of HF development. Pharmacological inhibition of EGF signalling alone had no influence on normal skin development, highlighting the importance of KGF signalling in these events. We propose that both EGF and KGF receptors are downregulated as part of a mechanism that requires abrogation of signalling via these pathways for placode formation to occur.
MATERIALS AND METHODS
Skin
To investigate in vivo skin, mystacial pad skin aged between E12.5 and E17.5, dorsolateral skin between E13.5 and E17.5, and tail skin at E18.5 were microdissected from C57/Bl6J mice, embedded in TissueTek OCT Compound (Agar Aids), snap frozen and stored at -80°C.
Organ culture
Organ culture of back skin was performed using a procedure modified from that described previously (Kashiwagi et al., 1997). To standardise experiments, discrete and consistently sized pieces of dorsolateral skin (∼1 × 2 mm2) were microdissected from the same region of C57/Bl6J mouse embryos at E13.5 (n=198). Skin was then placed epidermal side up onto rat tail collagen type 1 (Sigma) coated Nucleopore filters (pore size 8 μm; Whatman) floating on 2 ml of DMEM containing 1% Pen-Strep, 1% Fungizone and varying concentrations of EGF (Sigma), HBEGF (R&D Systems), amphiregulin (R&D Systems), KGF/FGF7 (R&D Systems), the EGF receptor inhibitor AG1478 (Calbiochem) or BSA control (Sigma) in a 35 mm dish. Skin samples were moistened with small amounts of media and incubated in a humidified atmosphere containing 5% CO2 for 6, 24 or 72 hours.
Individual skin samples were imaged using a KY-F1030 digital camera (JVC) fitted to a Stemi SVII dissecting microscope (Carl Zeiss) using both bright- and dark-field settings to visualise external follicle structures. When required, intact skin specimens were split with trypsin/pancreatin to facilitate quantitation (below) of developing follicles.
Cultured specimens were embedded in TissueTek OCT Compound, snap frozen and stored at -80°C or fixed in 4% paraformaldehyde in PBS overnight at 4°C for in situ hybridisation or paraffin wax embedding. For each experimental culture condition, at least six replicates were performed, involving a total of 258 back skin samples and 54 mystacial pad specimens.
Follicle analysis
Images of specimens were taken as above and analysed using UTHSCSA ImageTool software (http://ddsdx.uthscsa.edu/dig/itdesc.html); the area was calculated and total follicle numbers were counted. Individual follicles within regions of highest visible follicle density were chosen, and linear distances from the centre of the follicle to the centre of its nearest neighbours were measured. Means were calculated from the results of multiple (>3) measurements.
Immunofluorescence and immunohistochemistry
Cryosections (7 μm) were thaw-mounted on poly-L-lysine coated slides and methanol acetone-fixed. The following antibodies were used for specific labelling: anti-P-cadherin, 1:50 (clone P-CAD, ZYMED); anti-CD44-RPE (clone KM201, Serotec); polyclonal anti-BEK (FGFR2; C17) and polyclonal anti-EGFR (100S, SantaCruz, Autogen Bioclear UK, Wiltshire, UK), polyclonal anti-CD138 (syndecan 1; 281-2, BD Pharmingen, Oxford, UK), polyclonal EDAR (AF745, R&D Systems, Minneapolis, MN, USA), polyclonal anti-LEF1 (CL2A5, Cell Signaling Technologies, Danvers, USA).
Whole mount in situ hybridisation
The β-catenin probe was a kind gift from Dr Sarah Millar (University of Pennsylvania, Philadelphia, Pennsylvania, USA). The Gli1 probe was a kind gift from Dr Alexandra L. Joyner (Memorial Sloan-Kettering Cancer Center, New York, NY, USA). Whole mount in situ hybridisation was performed on cultured embryonic skin as per published protocols (Wilkinson, 1998).
Semi-quantitative RT-PCR
Total RNA was treated with DNaseI (Invitrogen). First strand cDNA was prepared from 25 ng total RNA using 0.5 μg of Oligo(dT) (Invitrogen) and 200 U of SuperScript II RT (Invitrogen). Polymerase chain reactions (PCRs) were performed as per manufacturer's instructions in a Peltier Thermal Cycler (MJ Research, Waltham, MA, USA). The different cDNAs from each of the timepoints were equalised to β-actin. Aliquots were removed from the total reaction after 25, 30 and 35 PCR cycles and electrophoresed on 1.0% agarose/TBE gels containing 0.5 mg/ml of ethidium bromide, and imaged using the Kodak Digital Science Electrophoresis and Documentation System 120.
Primers used for the analysis of endogenous expression of components of the EGF and KGF signalling pathways in the dermis and epidermis of dorsolateral skin (F, forward; R, reverse; 5′-3′): Areg-F, CATCATCCTCGCAGCTATTG; Areg-R, TTGTCCTCAGCTAGGCAATG; β-actin-F, CCTGTATGCCTCTGGTCGTA; β-actin-R, AAGGGTGTAAAACGCAGCTC; Egf-F, GATCCTATCACTGCACATGC; Egf-R, CAGTGCAAGTCTTCCCATCT; Egfr-F, GAGAGTG - ACTGTCTGGTCTGC; Egfr-R, GATGGGGTTGTTGCTGAATC; Fgfr2(IIIb)-F, GATGACCTTCAAGGACTTGG; Fgfr2(IIIb)-R, TTGTT - GATATCCCTGGCCAG; Hbegf-F, GGAAAGGGGTTAGGGAAGAA; Hbegf-R, TCCTCTCCTGTGGTACCTAAACA; Kgf-F, AGGGTGAGA - AGACTGTTCTG; Kgf-R, CTTTCCACCCCTTTGATTGC.
Primers used for the analysis of the expression of components of the EGF and KGF signalling pathways in ligand-treated back skin: β-actin-F, CCTGTATGCCTCTGGTCGTA; β-actin-R, AAGGGTGTAAAACGCAGCTC; Egfr-F, GAGAACCTGCAGATCATCAG; Egfr-R, ACCATGTTGCTTTGTTCTGC; Fgfr2(IIIb)-F, TCCTGGATCAGTGAG - AATGTGGAG; Fgfr2(IIIb)-R, GCTTGGGGGCCCGTGAACACGC; Hbegf-F, CCTTTTCAAAGTTGCTTTCTCC; Hbegf-R, TCCTCTCCTGTGGTACCTAAACA; Kgf-F, CAAACGGCTACGAGTGTGAA; Kgf-R, TCCGCTGTGTGTCCATTTAG.
Microarray analysis of KGF and HBEGF-treated embryonic skin
Triplicate culture experiments for each recombinant protein treatment were performed as described above. Each experimental reiteration contained six individual pieces of embryonic skin taken from littermates. This was incubated as described above for 24 hours in HBEGF and KGF, washed in Earle's media and incubated in a mixture of 0.75% trypsin (without EDTA) (Invitrogen) and filtered 2% pancreatin (Sigma-Aldrich, St Louis, MO, USA) in Earle's media for 20 minutes at 4°C. The epidermis was cleanly separated from the dermis, and each tissue was dissociated in RLT buffer (Qiagen, Valencia, CA, USA). Total RNA was isolated using the RNeasy Mini Kit according to the manufacturer's instructions (Qiagen). cDNA was synthesised and labelled RNA samples were transcribed for hybridisation on microarray chips (MOE430A) using Affymetrix reagents and protocols (Affymetrix, Santa Clara, CA, USA). The data output was normalised and analysed using GeneTraffic software (Iobion Informatics, La Jolla, CA, USA). The BSA-treated control was set as a reference for comparison purposes. The P-value cutoff was set to 0.05 and the significant fold difference was considered twofold higher or lower than baseline.
RESULTS
Localisation of sites of EGFR and FGFR signalling during hair follicle morphogenesis
Our previous study of epidermal morphogenesis by global transcriptional profiling (Bazzi et al., 2007a), revealed changes in the expression of several genes that suggested that the EGF and KGF pathways were active around the period of early follicle morphogenesis. To determine the localisation of the EGF and KGF receptor proteins, immunofluorescence was performed on embryonic skin from different body sites. E13.5 back skin epidermis consisted of a single layer of epithelial cells, which were all expressing EGFR and FGFR2(IIIb) protein (Fig. 1A,G). At E14.5, the epidermis was multiple layered and placodes had been initiated. Intriguingly, EGFR and FGFR2(IIIb) expression was downregulated to the extent that labelling was largely missing in placodes, compared with the interfollicular epidermis (Fig. 1B,H). This receptor downregulation occurs at a timepoint that corresponds with activated Wnt signalling in the placode and DC, indicated by an increased nuclear Lef1 expression in these early HF structures (Fig. 1H, insert). To ascertain whether loss of EGFR and FGFR2(IIIb) was a common feature of all primary back skin follicles, or just a subset, E15.0 skin was colabelled with the placodal marker P-cadherin (cadherin 3 - Mouse Genome Informatics) and either EGFR and FGFR2(IIIb). In all placodes, identified by P-cadherin expression, both receptors were downregulated (n=593 from 3 samples; Fig. 1C,I). In E12.5 mystacial pad skin, the same absence of EGFR and FGFR2(IIIb) protein was observed in the placodes (Fig. 1D,J). This consistent pattern of diminished EGFR and FGFR2(IIIb) expression was observed in the placodes of all follicle types examined, including that of tail skin (Fig. 1E,K) and secondary pelage follicles (Fig. 1F,M). The relative absence of both receptors persisted in the epithelium during the hair germ and peg stages of follicle development (see Fig. S1 in the supplementary material). Splitting and recombining skin is one method of delaying and synchronising follicle morphogenesis (Chuong et al., 1996). When this was used to disrupt and delay follicle morphogenesis (see Fig. S2 in the supplementary material) it did not ultimately affect FGFR2(IIIb) receptor downregulation, highlighting that receptor loss is linked specifically to placode formation, rather than a chronological timepoint.
Fig. 1.

EGFR and FGFR2(IIIb) expression is downregulated in hair follicle placodes. (A-M) Mouse skin sections from back, whisker pad and tail regions labelled with antibodies specific to EGFR (A-F), or FGFR2(IIIb) (G-M) (green). EGFR and FGFR2(IIIb) expression is uniform in back skin epidermis at E13.5 (A,G) and reduced in the placodes (arrows) of back skin follicles by E14.5 (B,H) and in all other developing follicles (D-F,J-M). At E14.5 LEF1 is upregulated in the nucleus of cells in the placode and early DC (H insert), costaining with placodal marker P-cadherin (red) and EGFR (green) (C) or P-cadherin (red) and FGFR2(IIIb) (green) (I). Insert shows receptor staining only. All images show nuclear DAPI (blue). Dotted lines mark the boundaries between epidermis and dermis. Scale bars: 60 μm. ep, epidermal placode.
Analysis of endogenous expression of other components of the EGF and KGF signalling pathways in the dermis and epidermis of dorsolateral skin between E12.5 and E15.5 is shown in Fig. S3 in the supplementary material. HBEGF and amphiregulin were omnipresent in the epidermis. EGFR, the receptor for these ligands, was identified at the transcript level in the developing epidermis at all stages, with a sharp drop in expression at E14.5, coinciding with HF initiation. Transcripts for KGF were only detected in the dermis. KGF receptor (FGFR2) transcripts were present in both the epidermis and dermis at all timepoints.
Activation of EGF and FGF signalling by endogenous ligands specifically blocks hair follicle initiation
The focal loss of expression of EGFR and FGFR2(IIIb) within placodes led us to investigate the effects of constitutively activated EGF and KGF signalling on HF morphogenesis. After 72 hours, developing follicles were visible in control skin cultures as rounded external structures distributed across the whole skin surface, including the edges (Fig. 2A). Haematoxylin and Eosin (H&E) staining revealed well-developed hair germs and a multi-layered epidermis starting to cornify (Fig. 2B). EGF, HBEGF and amphiregulin were selected to test the effect of EGFR ligands on the formation of HFs, and each was found to inhibit follicle formation, although at different potencies: 50 ng/ml of recombinant EGF was capable of completely blocking the presence of any discernable follicular structures on the skin surface (Fig. 2C); and 250 ng/ml of HBEGF was required to achieve the same effect (Fig. 2E). Amphiregulin was the least potent, and at concentrations up to 500 ng/ml a small number of follicles were observed externally (Fig. 2G). In addition to follicle inhibition, all of the EGFR ligands elicited numerous folds/wrinkles in the treated skins (Fig. 2C,E,G). When examined histologically, skin cultured with either EGF (50 ng/ml) or HBEGF (250 ng/ml) had no HF structures in 39 out of 43 cultures (91%; Fig. 2D,F) and thicker, well-cornified epidermis. The wrinkles/folds observed on the surface of the skin were clearly visible as deep downward epidermal projections into the dermis (Fig. 2D,F,H).
Fig. 2.

Constitutive EGF and FGF signalling can inhibit hair follicle morphogenesis. (A-L) E13.5 back skin organ-cultured with BSA (A,B) or a recombinant growth factor (C-L) for 72 hours (n=42 for each treatment). (A,C,E,G,I,K) Surface of the skin following culture. For skins treated for 72 hours with BSA (250 ng/ml, A) HFs are visible on the surface. Follicles could not be identified on EGF-(50 ng/ml, C), HBEGF- (250 ng/ml, E), or (I) KGF-(250 ng/ml) treated skins. (G) Amphiregulin (AR) was less potent, and blocked the majority of (but not all) follicles at 500 ng/ml. (K) FGF10 (250 ng/ml) had no visible effect on HF development. (B,D,F,H,J,L) H&E staining of skin sections. Arrows indicate: hair germs (yellow arrows); DCs (black arrows); epidermal folding (blue arrows); epidermal ridges (white arrows). Follicle counts were verified by enzymatically splitting cultured skin and confirming normal follicle numbers in control epidermis (M) and follicle inhibition in treated epidermis (N). Scale bars: 0.5 mm in A,C,E,G,I,K,M,N; 60 μm in B,D,F,H,J,L.
KGF (250 ng/ml) also completely inhibited follicle morphogenesis, as seen externally and histologically. These specimens had cornified epidermis that was uniformly thicker than that of the controls but no folding or wrinkling (Fig. 2I,J). However, another FGFR2(IIIb) ligand, FGF10, was unable to inhibit follicle morphogenesis (Fig. 2K,L) at the same concentration (250 ng/ml). To verify that structures visible externally were developing HFs, cultured skin was enzymatically separated into epidermis and dermis. In all cases, the number of surface bumps equalled the number of obvious developing follicles in the epidermis after separation (Fig. 2M). No follicles were observed in separated skin that had been cultured with 250 ng/ml HBEGF (Fig. 2N) or KGF (data not shown). EGF, HBEGF and KGF administered to E14.5 dorsolateral embryonic skin, in which EGFR- and FGFR2(IIIb)-negative placodes had already formed, elicited no inhibitory effect (see Fig. S4 in the supplementary material).
Inhibition of follicle morphogenesis by EGFR and FGFR2(IIIb) activation is dose dependent, morphology and patterning of residual hair follicles is normal
To investigate the dose-dependent inhibition of HF formation, skin cultures were established with increasing concentrations of each recombinant ligand, and follicle density was calculated. Results for HBEGF and FGF10 are shown in Fig. 3A. Control skin cultured for 72 hours in the absence of ligand developed 38±4.8 follicles per millimetre squared; this density was uniformly distributed across the skin surface. Upon treatment with increasing concentrations of HBEGF, HF numbers were correspondingly reduced. With 50 ng/ml they were halved to 19.5±7.9 (P<0.0005), they dropped to 7.5±5.4 (P<0.000013) with a dose of 125 ng/ml, and no follicles were visible at the highest dose of 250 ng/ml (Fig. 3A). Having observed no inhibitory effects using FGF10 at a concentration at which KGF was inhibitory (250 ng/ml; Fig. 2I), we showed that specimens treated with a range of concentrations of FGF10 up to 500 ng/ml were indistinguishable from controls, and showed no significant difference in follicle density (Fig. 3A).
Fig. 3.

HBEGF, but not FGF10, inhibits follicle morphogenesis in a dose-dependent manner but morphology and patterning of residual hair follicles are normal. (A) The number of follicles against treatment with increasing concentrations of HBEGF or FGF10 after 72 hours in culture; only HBEGF treatment elicits a dose-dependent reduction in follicle numbers. (B) Skin treated with 125 ng/ml of HBEGF stained with H&E (left) or labelled with anti-EDAR antibody (green), nuclei stained with DAPI (blue) (right). Residual follicles that escaped inhibition appeared morphologically indistinguishable from those in control cultured skin. (C) Following treatment of skin with up to 125 ng/ml of HBEGF, residual follicles were present in islands or groups. (D) Graphs represent the distance between at least four follicles from each skin sample (n=6) for each treatment concentration (of either HBEGF or FGF10) and their seven nearest neighbours within these groups (illustrated in C). Scale bars: 60 μm in B; 0.5 mm in C.
Follicles that developed in the presence of lower dosages of HBEGF, AR and KGF were morphologically indistinguishable from those in the control skins and expressed normal placodal markers, including EDAR (Fig. 3B). Intriguingly, on HBEGF-, AR- and KGF-treated skin samples in which follicles had developed, the follicles were not evenly distributed, but instead were localised in clusters. For HBEGF, the distance between the central follicle in each follicular cluster and its seven closest neighbours was quantified for each of the specimens (n=18) at 0 ng/ml, 50 ng/ml (P=0.0006) and 125 ng/ml (P=0.02) (Fig. 3C). No significant increases in interfollicular spacing were found between control and treated samples (Fig. 3D). The same analysis of FGF10-treated samples revealed no increase in follicle spacing between control and treated samples (Fig. 3D).
Absence of hair follicle placodes and dermal condensates in EGF- and FGF-treated skin
To investigate at the molecular level the apparent absence of follicles and whether epidermal folding might represent vestigial follicle structures, we studied the expression of three follicle markers: P-cadherin, which identifies placodes (Jamora et al., 2003); and CD44 and syndecan 1, which both identify dermal condensates (Hayashi et al., 2002; Underhill, 1993). The three proteins were highly expressed in their appropriate HF structures relative to interfollicular dermis and epidermis, both in control cultured skin and equivalently staged in vivo skin (Fig. 4A-D,K-P). Skin cultured with EGF, HBEGF or KGF for 24 or 72 hours revealed an absence of strong differential HF-associated expression of P-cadherin (Fig. 4E-J), CD44 (Fig. 4Q,T,W) and syndecan 1 (Fig. 4R,S,U,V,X,Y). However, KGF treatment resulted in a uniform layer of strong ectopic syndecan 1 expression beneath the basement membrane (Fig. 4V,Y). This is possibly due to an FGF-inducible response element on the syndecan 1 gene (Jaakkola et al., 1997).
Fig. 4.

P-cadherin, CD44 and syndecan 1 confirm the absence of placodes and dermal condensations in ligand-treated skin. (A-Y) P-cadherin, CD44 and syndecan 1 expression (red) in ligand-treated mouse back skin. P-cadherin is upregulated at sites of placode formation during HF morphogenesis (A,B). CD44 and syndecan 1 are expressed only in the DC within the dermal compartment of embryonic skin during HF morphogenesis (K-M). Controls show a distribution of P-cadherin, CD44 and syndecan 1 comparable with in vivo embryonic skin (C,D,N-P). Skin cultured with EGFR ligands EGF (50 ng/ml) or HBEGF (250 ng/ml) displayed no localised upregulated P-cadherin expression (E-H) and no differential dermal expression of CD44 or syndecan 1 (Q-V). Skin cultured with KGF (250 ng/ml) displayed no localised upregulated P-cadherin expression (I,J) or CD44 (W). KGF treatment resulted in a continuous layer of syndecan 1 expression in the dermal cells subjacent to the epidermis (X,Y). P-cadherin, CD44 and syndecan 1 (red), laminin (green), nuclear DAPI (blue). Scale bars: 60 μm.
Inhibition of vibrissa follicle development by activated EGF and FGF signalling occurs in a defined spatiotemporal order
Vibrissa follicles develop at around E12.5 in mice (Ibrahim and Wright, 1975) in a precise pattern of defined rows and columns. The latter are delineated by letter, starting with a (containing the largest and most posterior follicles), and rows are delineated by number, starting with 1 (the most dorsal row) (Oliver, 1966). During development, follicles develop spatiotemporally in a posterior-to-anterior direction, therefore column a and b follicles are the first to appear.
We investigated the effects of HBEGF and KGF ligand treatment on whisker pads from faces of E13.0 embryonic mice (Fig. 5A-C), at which point follicles in columns a, b, and some in column c, had begun to develop. The majority of follicles in column c and all follicles that would make up columns d through to f had yet to form, and therefore these regions were still expressing both EGFR and FGFR2(IIIb). Following 72 hours of culture, the control BSA-treated (250 ng/ml) skin developed in a manner equivalent to the in vivo process, displaying all rows a through f, as well as smaller follicles that appear at the nasal (anterior) end of the face (Fig. 5D,D′). By contrast, skin samples treated with HBEGF or KGF showed arrested follicle patterning. No additional follicles were observed on the skin surface at the anterior end of the whisker pad (Fig. 5E-F′), as confirmed by immunofluorescence analysis (Fig. 5G-I). Those posterior follicles that were present at the start of culture, however, continued to develop normally (Fig. 5E,F,K,L,M).
Fig. 5.

Treatment of organ-cultured E13.0 whisker pads with HBEGF or KGF inhibits new vibrissa follicle initiation but does not influence the development of existing follicles. (A-F′) Whisker pads at E13.0 in which the two most posterior columns of follicles had already begun to develop were cultured for 48 hours with BSA (250 ng/ml; control) (A,D,D′) or either HBEGF (250 ng/ml) (B,E,E′) or KGF (250 ng/ml) (C,F,F′). The control BSA-treated whisker pads (A,D,D′) continued to develop vibrissa follicles in the anterior direction; (D′) yellow dots used to highlight the presence of follicles. Whisker pads treated with HBEGF (B,E,E′) or KGF (C,F,F′) failed to develop any `new' follicles during the 48 hours of culture. (G-L) Whisker pads were sectioned and labelled with antibodies specific to syndecan 1 (red) and laminin (green). Analysis of the follicles in the posterior region showed that HBEGF (K) and KGF (L) had no effect on the development of existing HFs or the formation of the DP. Analysis of HBEGF-(H) or KGF-(I) treated skin in the anterior region of the whisker pads revealed no epidermal HF structures or DCs. Scale bars: 60 μm. dc, dermal condensation; dp, dermal papilla; hf, hair follicle.
Downregulation of Wnt and SHH signalling in EGF- and FGF-treated skin
To interrogate the status of key signalling pathways active during early pelage HF morphogenesis, we combined organ culture, microdissection and microarray analysis. Skin taken from treated and control skin cultures was enzymatically split into dermis and epidermis and transcriptional profiling was performed as we described previously (Bazzi et al., 2007a). After 24 hours of treatment with either KGF or HBEGF, we noted a marked reduction in many of the recognised molecular signatures of HF morphogenesis. These included key effectors in the Wnt, (LEF1, DKK4), sonic hedgehog (SHH, GLI1 and patched homolog 2) and BMP (BMP2) signalling pathways (Table 1). In mice, Wnt signalling is required for the early events of follicle initiation, whereas the SHH pathway is associated with later stages of follicular morphogenesis. To verify the downregulation of the Wnt and SHH pathways whole mount in situ hybridisation was performed. In control skin after 24 hours of culture, β-catenin mRNA had accumulated in developing follicles corresponding to cells in which the Wnt signalling pathway is activated (Fig. 6A). Gli1 mRNA transcript expression represents a readout for activation of the SHH signalling pathway, and at 72 hours Gli1 mRNA was concentrated in a ring pattern (Fig. 6A). Sectioning of skin at both timepoints confirmed focal expression of Gli1 in the dermis of the developing follicles (Fig. 6B). In EGF-, HBEGF- and KGF-treated embryonic skin, no follicular structures and no discrete accumulation of β-catenin beyond background levels was evident (Fig. 6A,B). To investigate the early effects of EGFR or FGFR2(IIIb) activation on Wnt signalling, we cultured day E13.5 skin with control BSA, HBEGF or KGF for 6-hour and 24-hour timepoints and evaluated the nuclear expression of LEF1, an established marker of activated Wnt signalling. At 6 hours, control skin expressed nuclear LEF1 in the epidermis and, at a low level, in some dermal cells. Following 24 hours, cells in placodes and DCs were highly stained by the LEF1 antibody, in a manner comparable to that observed in vivo (Fig. 1H). Treatment with HBEGF or KGF resulted in an absence of discrete LEF1 signalling in both compartments at either the 6-hour or 24-hour timepoints. Interestingly, treated skins continued to express nuclear LEF-1 at all culture timepoints investigated but without a focal distribution (Fig. 6C).
Table 1.
Genes implicated in epidermal differentiation that are downregulated in the epidermis following 24-hour culture with EGF or KGF ligand
| Gene | Control | KGF | HBEGF |
|---|---|---|---|
| sonic hedgehog | 0 | −9 | −9 |
| dickkopf homolog 4 (Xenopus laevis) | 0 | −9 | −13 |
| GLI-Kruppel family member GLI1 | 0 | −8 | −10 |
| periostin, osteoblast specific factor | 0 | −5 | 0 |
| patched homolog 2 | 0 | −5 | −12 |
| bone morphogenetic protein 2 | 0 | −3 | −12 |
| cut-like 1 | 0 | −3 | −3 |
| lymphoid enhancer binding factor 1 | 0 | −2 | −2 |
Embryonic skin was cultured with BSA (control) (250 ng/ml), HBEGF (250 ng/ml) or KGF (250 ng/ml) for 24 hours and genomic transcript expression was profiled by microarray. A number of genes linked to major pathways (Wnt, sonic hedgehog, BMP) involved in HF morphogenesis are downregulated following HBEGF or KGF treatment. 0, baseline levels or no change; numbers, fold of baseline.
Fig. 6.

Wnt and SHH pathways fail to activate in HBEGF- and KGF-treated skin. (A) Whole mounts of cultured embryonic skin. β-catenin and Gli1 transcripts are expressed focally in the control cultures at 24 and 72 hours, respectively. HBEGF-, EGF- or KGF-treated skin samples, which have no external follicle structures, fail to show localised expression of β-catenin and Gli1 transcripts. (B) Sectioned whole-mount skin at 24 hours and 72 hours showing upregulated Gli1 expression in the dermal condensation of BSA-treated controls, but not in HBEGF- or KGF-treated samples. (C) E13.5 back skin was cultured with either BSA or a recombinant growth factor for 6 (top) or 24 (bottom) hours (n=4 for each treatment) and labelled with antibody specific to LEF1 (green). At 6 hours, there is no foci of LEF1 expression in either the epidermis or dermis of all samples. At 24 hours, LEF1 expression was unregulated in a localised manner in the epidermal placode and dermal condensation of BSA-treated controls, but not in the HBEGF- or KGF-treated samples. Scale bars: 60 μm.
HBEGF and KGF treatment promote an interfollicular epidermal phenotype
KGF and HBEGF treatment resulted in the rapid induction of genes involved in epidermal terminal differentiation. These genes included S100a18 (Hrnr - Mouse Genome Informatics), S100a6, loricrin, keratin 6A, Tgm1 K (Tgm1), Lgals3, Lgals7 and filaggrin. Moreover, S100a18 and keratin 6A, which are normally absent until E15.5 in vivo (Bazzi et al., 2007a), were expressed in cultured skin equivalent to E14.5 and therefore appeared prematurely (Table 2). To validate the microarray analysis, immunofluorescence for keratin 6A, filaggrin and loricrin was performed on treated cultures. The expression of each was increased in the epidermis from treated skin compared with the control (see Fig. S5A-I in the supplementary material).
Table 2.
Genes implicated in epidermal differentiation that are upregulated in the epidermis following 24-hour culture with EGF or KGF ligand
| Gene | Control | KGF | HBEGF |
|---|---|---|---|
| filaggrin | 0 | 98 | 29 |
| involucrin | 0 | 38 | 24 |
| loricrin | 0 | 2 | 2 |
| keratin complex 2, basic, gene 6a | 0 | 3 | 5 |
| keratin complex 2, basic, gene 6b | 0 | 20 | 85 |
| keratin complex, acidic, gene 16 | 0 | 64 | 70 |
| S100a18 | 0 | 6 | 5 |
| S100a6 | 0 | 9 | 6 |
| transglutaminase 1, K polypeptide | 0 | 2 | 2 |
| lectin, galactose binding, soluble 7 | 0 | 3 | 2 |
| small proline-rich protein 1A | 0 | 10 | 6 |
| small proline-rich protein 1B | 0 | 5 | 5 |
| small proline-rich protein 2A | 0 | 12 | 18 |
| small proline-rich protein 3 | 0 | 144 | 25 |
Embryonic skin was cultured with BSA (control) (250 ng/ml), HBEGF (250 ng/ml) or KGF (250 ng/ml) for 24 hours and genomic transcript expression was profiled by microarray. Multiple genes involved in normal interfollicular keratinocyte differentiation and barrier formation are upregulated following HBEGF or KGF treatment. 0, baseline levels or no change; numbers, fold of baseline.
Blocking of EGFR using AG1478 has no effect on hair follicle morphogenesis in organ-cultured skin
To investigate the effects of the inhibition of EGFR signalling on HF morphogenesis, E13.5 skin was organ cultured with AG1478, an inhibitor of EGF receptor kinase autophosphorylation. AG1478 is specific for EGFR at concentrations up to 100 μM (Gazit et al., 1991), and was previously shown to cause fusion of interbud rows at a concentration of 50 μM (Atit et al., 2003). Following 72 hours of culture with AG1478 at a concentration of 100 μM, treated skin specimens (n=12) appeared indistinguishable from controls. Skin was split (n=3) to reveal individual hair pegs with no visible fusion (data not shown). Histology and immunofluorescence staining with anti-syndecan showed normal follicles comparable with controls (Fig. 7A,B). Finally, measurement showed that there was no significant difference in follicle number or interfollicular spacing between treated and control skin specimens (Fig. 7D,E).
Fig. 7.

Inhibition of EGFR signalling has no effect on hair follicle development or density. (A-E) Skins from E13.5 mice were cultured with either BSA (250 ng/ml) or the EGFR inhibitor AG1478 (100 μM). AG1478-treated skin surface (A). Antibody labelling for syndecan 1 (red) and laminin (green), with nuclei labelled with DAPI (blue) (B) and H&E (C) revealed that follicles were indistinguishable from those of the control (see Figs 2, 4). Follicular density and the average distance between follicles and their nearest neighbours were quantified (D,E). No significant difference between treated and control specimens was observed for either parameter. Scale bars: 60 μm.
Interplay between EGF and KGF signalling in ligand-treated organ-cultured skin
To investigate how short-term treatment with HBEGF or KGF influenced the expression of each other, and their own receptors, E13.5 mouse back skin was cultured in the presence of BSA (250 ng/ml), HBEGF (250 ng/ml) or KGF (250 ng/ml) for 6 hours. Dermis and epidermis were then enzymatically separated and the expression of Hbegf, Egfr, Kgf and Fgfr2(IIIb) mRNA transcripts was investigated using semi-quantitative RT-PCR. HBEGF treatment appeared to have no effect on the expression of either the ligands (Hbegf and Kgf) or the receptors [Egfr and Fgfr2(IIIb)] of either pathway when compared with controls (Fig. 8). However, KGF treatment upregulated Kgf expression in the dermis, and in the epidermis it produced increased Hbegf and decreased Fgfr2(IIIb) expression, but no change in Egfr expression.
Fig. 8.
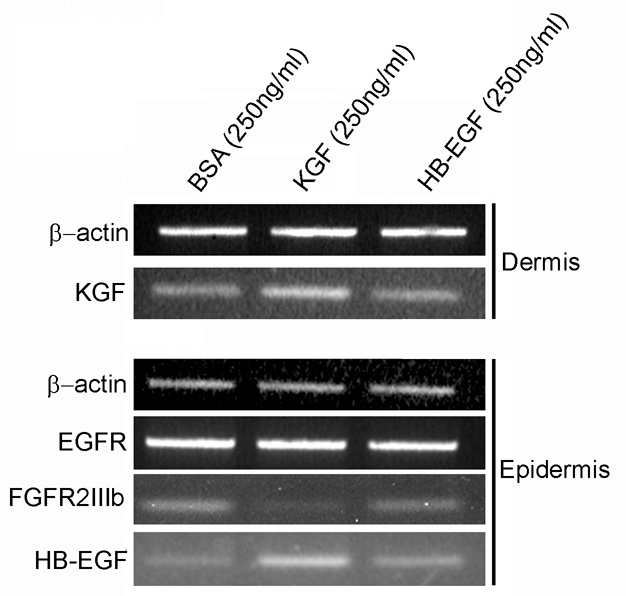
The effects of HBEGF and KGF treatment on mRNA transcript expression of ligands and receptors of EGF and KGF pathways in organ-cultured E13.5 mouse back skin. Semi-quantitative RT-PCR showing Hbegf, Egfr, Kgf and Fgfr2(IIIb) mRNA transcript expression in the appropriate epidermal or dermal compartment of E13.5 mouse back skin following 6 hours culture with BSA (250 ng/ml), KGF (250 ng/ml) or HBEGF (250 ng/ml). Compared with control specimens, skin cultured with HBEGF showed no difference in transcript levels of epidermal expression of Hbegf, the dermal expression of Kgf, the epidermal expression of Egfr or the epidermal expression of Fgfr2(IIIb). Skin cultured with KGF for 6 hours showed an increased epidermal expression of Hbegf, an increase in Kgf dermal expression, and a decrease in epidermal Fgfr2(IIIb) transcript expression. KGF treatment does not alter epidermal expression of Egfr.
DISCUSSION
EGF and KGF signalling pathways are conventionally seen as promoting cell growth and proliferation in the interfollicular epidermis (Beer et al., 2000; du Cros, 1993; Peus and Pittelkow, 1996; Schneider et al., 2008). However, mouse models with elements of these pathways disrupted in skin have indicated that activated EGF and KGF signalling are both crucial for HF morphogenesis and development (Danilenko et al., 1995; Guo et al., 1996; Miettinen et al., 1995; Murillas et al., 1995; Petiot et al., 2003; Werner et al., 1994). Here we demonstrate for the first time that FGFR2(IIIb), as well as EGFR, are coordinately downregulated in placodes of all mouse follicles. In a series of functional organ culture experiments, we show that when skin from the sites of primary pelage and whisker follicle development is exposed to increased levels of two endogenous EGFR ligands (HBEGF and amphiregulin) and the FGFR2(IIIb) receptor ligand KGF, follicle formation is inhibited in a dose- and time-dependent manner. These data demonstrate that active EGF and KGF signalling in the placodes is prohibitive of HF formation. Since alterations in the dermal condensate could alter placode formation, the question arises as to whether the effects of the EGF and KGF ligands are due to activation of signalling in the epidermis or the dermis? Our analysis of placode and dermal condensate markers in treated skin at short timepoints have revealed that condensates can briefly persist in the dermis, but placodes are absent from the epidermis (see Fig. S6 in the supplementary material). This, together with the exclusive epidermal expression of the receptors, leads us to suggest that the inhibitory effects of EGF and KGF signalling act initially on the epidermis. Microarray profiling, and analysis of Wnt and SHH pathway expression in treated skin, supports a model in which failure to downregulate EGFR and FGFR2(IIIb) in HF placodes leads to adoption of an interfollicular epidermal fate.
EGFR and FGFR2(IIIb) are coordinately downregulated during placode initiation
Previous reports have shown that the epidermal growth factor receptor is downregulated in the basal cells of primary human follicle placodes (Nanney et al., 1990). However, the concomitant downregulation of FGFR2(IIIb) receptor specifically within the epithelial placodes of developing primary follicles is a novel observation that differs from previous reports (Danilenko et al., 1995). We showed the same kinetics of expression in secondary follicles, whisker follicles and tail skin follicles. Our results showed that synchronised downregulation of EGFR and FGFR2(IIIb) appear to be a universal process in follicle development, unlike for example the tabby/downless/crinkled (EDA/EDAR/EDARADD) pathway, which is associated with the development of a specific subset of follicles (Mou et al., 2006). In order for one or both pathways to be active, a receptor/ligand pair must be present in the tissue. Our finding that expression of the EGF ligands HBEGF and amphiregulin in the epidermis, and of KGF in the dermis, was coincident with pelage follicle formation, is in general agreement with previous observations on developing mouse (Kashiwagi et al., 1997), rat (Dang et al., 2003) and human (Piepkorn et al., 1995) skin.
Inhibition of placodes by EGFR and FGFR2(IIIb) ligands: effects of exposure to ligands are dose- and time-dependent but do not perturb morphogenesis and pattern formation
Kashiwangi et al. (Kashiwangi et al., 1997) demonstrated that two EGFR ligands, EGF and TGFα, inhibited follicle development in a dose-dependent manner in embryo skin organ culture. These authors postulated that two other ligands, HBEGF and amphiregulin, could be endogenous mediators of HF and epidermal development. Our study showed that treating embryonic mouse skin with recombinant HBEGF, amphiregulin or the FGFR2(IIIb) ligand, KGF, prior to follicle initiation inhibited the development of follicular structures dose dependently. One interpretation of the pronounced epidermal folds observed in the EGF ligand-treated samples is that they were aberrant or vestigial follicle structures. However, the absence of placode (P-cadherin) (Jamora et al., 2003) and condensation (syndecan 1 and CD44) (Trautman et al., 1991) markers showed that this was not the case. Our findings correlate to some degree with mouse models, whereby transgenic mice expressing the human amphiregulin gene displayed a distinctive loss of follicles in the most severely affected regions (Cook et al., 1997). Targeting of KGF overexpression in mouse epidermis also resulted in a marked suppression of HF morphogenesis (Guo et al., 1993). Intriguingly, we showed that treatment with FGF10, an alternate ligand that binds to the FGFR2(IIIb) receptor, produced no observable effect on follicle development. Mice lacking FGF10 show a whisker follicle (Ohuchi et al., 2003) but not a body HF (Suzuki et al., 2000) phenotype.
In agreement with previous studies using EGF (Kashiwagi et al., 1997), we showed that when follicles from HBEGF-, amphiregulin- and KGF-treated skin did escape inhibition (in the presence of lower concentrations of ligand) they developed normally and expressed normal HF-related markers. The fact that these residual follicles were clustered with normal density suggests that there is localised failure of follicular initiation in any given area, rather than a generalised expansion of interfollicular epidermis increasing the distance between follicles. Interestingly, some K14-hKGF (human KGF) transgenic mice showed a mosaic pattern of skin and hair development, with regions containing normal HFs alternating with areas possessing thicker epidermis devoid of HF structures (Guo et al., 1993).
Previous work reported that after the start of follicle initiation, skin is unresponsive to the inhibitory effects of EGF treatment (Kashiwagi et al., 1997). Here, we showed that the same was true of skin treated with the endogenously expressed ligands HBEGF and KGF (see Fig. S4 in the supplementary material), emphasising that follicle initiation is crucial in relation to EGF and KGF pathway signalling and follicle morphogenesis. With vibrissa follicles on whisker pads we were able to simultaneously show three different results (loss of receptors in placodes, inhibition of HF induction with excess ligand and continuation of normal development and patterning in committed follicles), reinforcing both the findings with primary pelage follicles and the universal nature of these signalling events.
HBEGF and KGF promote epidermal fate at the expense of HF development
We performed global transcriptional profiling to identify gene pathways that are activated or inhibited in the epidermis after EGFR and FGFR2(IIIb) signalling. Given the well-established role of Wnt/β-catenin signalling for the initiation of HF development (Andl et al., 2002; Zhang et al., 2008) it was noteworthy that Lef1, which is a downstream effector of Wnt signalling, and the Wnt regulator Dkk4 (Bazzi et al., 2007b), were both downregulated. Sonic hedgehog, Gli1 and patched 2, key components of the SHH signalling pathway involved later in follicular development (Chiang et al., 1999; Karlsson et al., 1999; St-Jacques et al., 1998), were also downregulated. These observations were verified with key downstream mediators of the Wnt and SHH pathways, respectively, using whole mount in situ hybridisation and antibody staining. This showed an absence of distinct focal HF-related expression. They demonstrate at a molecular level the complete failure of follicular development following exposure to EGF and KGF signalling at the pre-placode stage. Although there is a failure of placodal Wnt signalling when E13.5 skin is cultured with ligands, we are not suggesting that the activation of either receptor directly inhibits the Wnt signalling pathway. Interestingly, as the epidermis of the treated skins expressed nuclear LEF1 at all culture timepoints investigated, albeit at low levels (Fig. 6C), it is attractive to suggest that EGFR and FGFR2(IIIb) signalling does not inhibit the activation of the Wnt signalling pathway.
Contrasting with this was the strong expression of genes associated with epidermal differentiation, including S100 proteins and small proline-rich proteins. Filaggrin, which is expressed relatively late on in the stratification process (Bickenbach et al., 1995), emerged prematurely in response to stimulation by both ligands. Likewise, keratins 6 and 16, known to be involved in epidermal homeostasis, as well as being expressed during the `activation' of keratinocytes in wound healing, were also upregulated. Taken together, these findings suggest that exposure to EGF ligands and KGF promotes an interfollicular epidermal fate, at the expense of HF morphogenesis (Guo et al., 1993; Schneider et al., 2008).
The FGF pathway acts synergistically on the EGF pathway but not vice versa
Our examination of the short-term influence of HBEGF and KGF ligands on both pathways demonstrated that KGF upregulated its own mRNA levels in E13.5 dermis, and that this was coupled with decreased transcript levels of its receptor in the epidermis. Interestingly, KGF treatment also strongly increased mRNA levels of multiple EGFR ligands, including Hbegf, suggesting that KGF can directly influence the EGF pathway. It has been shown previously that KGF treatment of keratinocytes induces the production of secreted TGFα protein, and that this is linked to the downregulation of the EGF receptor (Dlugosz et al., 1994). The reason we observed no EGF receptor downregulation could be a matter of timing, since the above study was performed over days compared with 6 hours here.
By contrast, HBEGF treatment appeared to have no effect on the expression of either the ligands (HBEGF and KGF) or the receptors [EGFR and FGFR2(IIIb)] of either pathway when compared with control specimens (Fig. 8). However, auto- and cross-induction, and autocrine signalling, occurs extensively among the EGF receptor ligands (Barnard et al., 1994; Piepkorn et al., 1998) with HBEGF in particular having a proposed juxtacrine interaction with its receptor (Harris et al., 2003), which might be pertinent to placode formation and the creation of a follicular/non-follicular cell boundary in adjacent basal epidermal cells. Crucially, some EGF ligands are able to downregulate the EGFR receptor (Singh and Harris, 2005) and, more generally, different ligands are known to alter the magnitude and duration of EGFR signals (Wells, 1999).
Denticles to feathers to hairs: alternating regions of EGFR signalling establish regional specificity throughout evolution
Treatment of embryonic chick skin with exogenous EGF suppresses feather formation (Atit et al., 2003) in a manner similar to that seen here with other EGF ligands and mouse HFs. Chick skin explant cultures treated with AG1478, a recognised inhibitor of EGFR signalling (Zieske et al., 2000) causes fusion of feather buds, implying loss of interfollicular epidermal fate. In our study, treatment of mouse skin with AG1478 at varying concentrations failed to elicit any effect on development and patterning of pelage and vibrissa follicles (data not shown). Similarly, in mice with targeted disruption of EGFR, follicle morphogenesis is not disrupted (Hansen et al., 1997). Thus, although in embryonic chick skin the EGFR pathway might be sufficient to control interfollicular fate, aspects of our work support the model that in mouse it does so in conjunction with KGF signalling.
Why then, given the other well-established pathways that are known to be involved in HF placode formation, do EGF and FGF signalling downregulate locally in the placode, and what interactions occur with other pathways? Concerning the first question, a trivial answer is that it is a neutral response to a switch of fate involving other mechanisms. Indeed, evidence from the literature suggests that EDAR, for example, does not directly interact with EGF during follicle initiation. A recent study showed that although both BMP4 and EGF are able to block the rescue of follicle formation by EDA in Eda-/- mouse skin, BMPs but not EGF are able to repress Edar expression in embryonic mouse skin prior to follicle patterning (Mou et al., 2006). Among TGFβ family members, the TGFβ2 isoform appears to be particularly important for follicle morphogenesis, as Tgfb2 null mice have reduced hair follicle numbers, and adding TGFβ2 to developing skin in organ cultures promotes epidermal hyperplasia and follicle induction (Foitzik et al., 1999). Since all TGFβ isoforms have inhibitory effects on keratinocyte cell proliferation in vitro, Foitzik et al. postulate that the TGFβ2-specific influence on follicle development in vivo may be indirectly influencing other growth factor pathways via different cell types. Certainly, in vitro evidence militates against the idea that TGFβ2 directly stimulates the KGF and EGF pathways; for example, TGFβ2 dose-dependently inhibits corneal epithelial cell proliferation promoted by KGF and EGF (Honma et al., 1997). Likewise, in relation to early follicle development there are no reports of direct interaction between noggin, an important antagonist of BMP activity located in the mesenchyme (Botchkarev et al., 1999) and KGF and EGF signalling. However, it is interesting that the Notch pathway is linked to early follicle development (Favier et al., 2000) and that EGFR has been recently identified as a key negative regulator of notch1 gene expression in primary human keratinocytes (Kolev et al., 2008). It is also noteworthy that AP2α, which is a negative regulator of the EGF receptor (Wang et al., 2006), also becomes strongly expressed in the emerging HF placode (Panteleyev et al., 2003).
Guo et al. (Guo et al., 1993) suggest that `elevated growth response might block the mesenchymal-epithelial signalling necessary for HF morphogenesis'. Certainly, one of the features associated with placode formation is a reduction in epidermal cell division (Mustonen et al., 2004; Wessells and Roessner, 1965).Therefore there may be a requirement to interrupt cell division, to permit the transition from interfollicular to follicular status and to create a new stem cell pool.
The Fuchs group has elegantly shown some of the cellular changes that occur during hair placode initiation (Jamora et al., 2003). These include a switch in expression from E-cadherin (cadherin 1 - Mouse Genome Informatics), an adhesion molecule closely linked with the EGFR pathway (Hazan and Norton, 1998), to P-cadherin, coincident with other cytoskeletal and behavioural changes to the cells. Therefore, the temporary downregulation of EGF and FGF signalling may be integral to global changes to cell behaviour and directional cell movements that occur in the placode.
Alternating regions of heightened and diminished EGFR signalling is an evolutionarily conserved mechanism, for example, it establishes vein patterning in Drosophila morphogenesis (Blair, 2007). Alternating regions of Wnt and EGFR signalling are also hallmarks of patterning the denticles (Payre et al., 1999). These mechanisms are reprised in the chick, where EGF signalling specifies the interbud fate (Atit et al., 2003). That similar mechanisms have been adapted for the formation of mammalian HFs is perhaps not unexpected for EGF signalling. Now we have uncovered a crucial overlapping role for KGF signalling in mouse HF initiation, adding a new layer of complexity to an already exquisitely regulated developmental process.
Supplementary material
Supplementary material for this article is available at http://dev.biologists.org/cgi/content/full/136/13/2153/DC1
Supplementary Material
We thank members of the LSSU at Durham for their assistance. At Columbia University we thank Mr Ming Zhang for excellent technical assistance and Drs Vladan Miljkovic and Yonghui Zhang for expert assistance in microarray analysis. This work was partly supported by a generous grant from the Steven and Michelle Kirsch Foundation (to A.M.C. and C.A.B.J.), by the New York State Foundation for Science Technology and Innovation (NYSTAR, to A.M.C.) and by the BBSRC (to C.A.B.J.). Deposited in PMC for release after 6 months.
References
- Andl, T., Reddy, S. T., Gaddapara, T. and Millar, S. E. (2002). WNT signals are required for the initiation of hair follicle development. Dev. Cell 2, 643-653. [DOI] [PubMed] [Google Scholar]
- Atit, R., Conlon, R. A. and Niswander, L. (2003). EGF signaling patterns the feather array by promoting the interbud fate. Dev. Cell 4, 231-240. [DOI] [PubMed] [Google Scholar]
- Barnard, J. A., Graves-Deal, R., Pittelkow, M. R., DuBois, R., Cook, P., Ramsey, G. W., Bishop, P. R., Damstrup, L. and Coffey, R. J. (1994). Auto- and cross-induction within the mammalian epidermal growth factor-related peptide family. J. Biol. Chem. 269, 22817-22822. [PubMed] [Google Scholar]
- Bazzi, H., Fantauzzo, K. A., Richardson, G. D., Jahoda, C. A. and Christiano, A. M. (2007a). Transcriptional profiling of developing mouse epidermis reveals novel patterns of coordinated gene expression. Dev. Dyn. 236, 961-970. [DOI] [PubMed] [Google Scholar]
- Bazzi, H., Fantauzzo, K. A., Richardson, G. D., Jahoda, C. A. and Christiano, A. M. (2007b). The Wnt inhibitor, Dickkopf 4, is induced by canonical Wnt signaling during ectodermal appendage morphogenesis. Dev. Biol. 305, 498-507. [DOI] [PubMed] [Google Scholar]
- Beer, H. D., Gassmann, M. G., Munz, B., Steiling, H., Engelhardt, F., Bleuel, K. and Werner, S. (2000). Expression and function of keratinocyte growth factor and activin in skin morphogenesis and cutaneous wound repair. J. Investig. Dermatol. Symp. Proc. 5, 34-39. [DOI] [PubMed] [Google Scholar]
- Bickenbach, J. R., Greer, J. M., Bundman, D. S., Rothnagel, J. A. and Roop, D. R. (1995). Loricrin expression is coordinated with other epidermal proteins and the appearance of lipid lamellar granules in development. J. Invest. Dermatol. 104, 405-410. [DOI] [PubMed] [Google Scholar]
- Blair, S. S. (2007). Wing vein patterning in Drosophila and the analysis of intercellular signaling. Annu. Rev. Cell Dev. Biol. 23, 293-319. [DOI] [PubMed] [Google Scholar]
- Botchkarev, V. A., Botchkareva, N. V., Roth, W., Nakamura, M., Chen, L. H., Herzog, W., Lindner, G., McMahon, J. A., Peters, C., Lauster, R. et al. (1999). Noggin is a mesenchymally derived stimulator of hair-follicle induction. Nat. Cell Biol. 1, 158-164. [DOI] [PubMed] [Google Scholar]
- Botchkareva, N. V., Botchkarev, V. A., Chen, L. H., Lindner, G. and Paus, R. (1999). A role for p75 neurotrophin receptor in the control of hair follicle morphogenesis. Dev. Biol. 216, 135-153. [DOI] [PubMed] [Google Scholar]
- Chiang, C., Swan, R. Z., Grachtchouk, M., Bolinger, M., Litingtung, Y., Robertson, E. K., Cooper, M. K., Gaffield, W., Westphal, H., Beachy, P. A. et al. (1999). Essential role for Sonic hedgehog during hair follicle morphogenesis. Dev. Biol. 205, 1-9. [DOI] [PubMed] [Google Scholar]
- Chuong, C. M., Widelitz, R. B., Ting-Berreth, S. and Jiang, T. X. (1996). Early events during avian skin appendage regeneration: dependence on epithelial-mesenchymal interaction and order of molecular reappearance. J. Invest. Dermatol. 107, 639-646. [DOI] [PubMed] [Google Scholar]
- Cohen, S. and Elliott, G. A. (1963). The stimulation of epidermal keratinization by a protein isolated from the submaxillary gland of the mouse. J. Invest. Dermatol. 40, 1-5. [DOI] [PubMed] [Google Scholar]
- Cook, P. W., Piepkorn, M., Clegg, C. H., Plowman, G. D., DeMay, J. M., Brown, J. R. and Pittelkow, M. R. (1997). Transgenic expression of the human amphiregulin gene induces a psoriasis-like phenotype. J. Clin. Invest. 100, 2286-2294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dang, C. M., Beanes, S. R., Soo, C., Ting, K., Benhaim, P., Hedrick, M. H. and Lorenz, H. P. (2003). Decreased expression of fibroblast and keratinocyte growth factor isoforms and receptors during scarless repair. Plast. Reconstr. Surg. 111, 1969-1979. [DOI] [PubMed] [Google Scholar]
- Danilenko, D. M., Ring, B. D., Yanagihara, D., Benson, W., Wiemann, B., Starnes, C. O. and Pierce, G. F. (1995). Keratinocyte growth factor is an important endogenous mediator of hair follicle growth, development, and differentiation. Normalization of the nu/nu follicular differentiation defect and amelioration of chemotherapy-induced alopecia. Am. J. Pathol. 147, 145-154. [PMC free article] [PubMed] [Google Scholar]
- Dlugosz, A. A., Cheng, C., Denning, M. F., Dempsey, P. J., Coffey, R. J., Jr and Yuspa, S. H. (1994). Keratinocyte growth factor receptor ligands induce transforming growth factor alpha expression and activate the epidermal growth factor receptor signaling pathway in cultured epidermal keratinocytes. Cell Growth Differ. 5, 1283-1292. [PubMed] [Google Scholar]
- du Cros, D. L. (1993). Fibroblast growth factor and epidermal growth factor in hair development. J. Invest. Dermatol. 101, 106S-113S. [DOI] [PubMed] [Google Scholar]
- Favier, B., Fliniaux, I., Thelu, J., Viallet, J. P., Demarchez, M., Jahoda, C. A. and Dhouailly, D. (2000). Localisation of members of the notch system and the differentiation of vibrissa hair follicles: receptors, ligands, and fringe modulators. Dev. Dyn. 218, 426-437. [DOI] [PubMed] [Google Scholar]
- Foitzik, K., Paus, R., Doetschman, T. and Dotto, G. P. (1999). The TGF-beta2 isoform is both a required and sufficient inducer of murine hair follicle morphogenesis. Dev. Biol. 212, 278-289. [DOI] [PubMed] [Google Scholar]
- Gazit, A., Osherov, N., Posner, I., Yaish, P., Poradosu, E., Gilon, C. and Levitzki, A. (1991). Tyrphostins. 2. Heterocyclic and alpha-substituted benzylidenemalononitrile tyrphostins as potent inhibitors of EGF receptor and ErbB2/neu tyrosine kinases. J. Med. Chem. 34, 1896-1907. [DOI] [PubMed] [Google Scholar]
- Guo, L., Yu, Q. C. and Fuchs, E. (1993). Targeting expression of keratinocyte growth factor to keratinocytes elicits striking changes in epithelial differentiation in transgenic mice. EMBO J. 12, 973-986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo, L., Degenstein, L. and Fuchs, E. (1996). Keratinocyte growth factor is required for hair development but not for wound healing. Genes Dev. 10, 165-175. [DOI] [PubMed] [Google Scholar]
- Hansen, L. A., Alexander, N., Hogan, M. E., Sundberg, J. P., Dlugosz, A., Threadgill, D. W., Magnuson, T. and Yuspa, S. H. (1997). Genetically null mice reveal a central role for epidermal growth factor receptor in the differentiation of the hair follicle and normal hair development. Am. J. Pathol. 150, 1959-1975. [PMC free article] [PubMed] [Google Scholar]
- Harris, R. C., Chung, E. and Coffey, R. J. (2003). EGF receptor ligands. Exp. Cell Res. 284, 2-13. [DOI] [PubMed] [Google Scholar]
- Hayashi, K., Mochizuki, M., Nomizu, M., Uchinuma, E., Yamashina, S. and Kadoya, Y. (2002). Inhibition of hair follicle growth by a laminin-1 G-domain peptide, RKRLQVQLSIRT, in an organ culture of isolated vibrissa rudiment. J. Invest. Dermatol. 118, 712-718. [DOI] [PubMed] [Google Scholar]
- Hazan, R. B. and Norton, L. (1998). The epidermal growth factor receptor modulates the interaction of E-cadherin with the actin cytoskeleton. J. Biol. Chem. 273, 9078-9084. [DOI] [PubMed] [Google Scholar]
- Headon, D. J. and Overbeek, P. A. (1999). Involvement of a novel Tnf receptor homologue in hair follicle induction. Nat. Genet. 22, 370-374. [DOI] [PubMed] [Google Scholar]
- Headon, D. J., Emmal, S. A., Ferguson, B. M., Tucker, A. S., Justice, M. J., Sharpe, P. T., Zonana, J. and Overbeek, P. A. (2001). Gene defect in ectodermal dysplasia implicates a death domain adapter in development. Nature 414, 913-916. [DOI] [PubMed] [Google Scholar]
- Honma, Y., Nishida, K., Sotozono, C. and Kinoshita, S. (1997). Effect of transforming growth factor-beta1 and -beta2 on in vitro rabbit corneal epithelial cell proliferation promoted by epidermal growth factor, keratinocyte growth factor, or hepatocyte growth factor. Exp. Eye Res. 65, 391-396. [DOI] [PubMed] [Google Scholar]
- Huelsken, J., Vogel, R., Erdmann, B., Cotsarelis, G. and Birchmeier, W. (2001). beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell 105, 533-545. [DOI] [PubMed] [Google Scholar]
- Ibrahim, L. and Wright, E. A. (1975). The growth of rats and mice vibrissae under normal and some abnormal conditions. J. Embryol. Exp. Morphol. 33, 831-844. [PubMed] [Google Scholar]
- Jaakkola, P., Vihinen, T., Maatta, A. and Jalkanen, M. (1997). Activation of an enhancer on the syndecan-1 gene is restricted to fibroblast growth factor family members in mesenchymal cells. Mol. Cell. Biol. 17, 3210-3219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamora, C., DasGupta, R., Kocieniewski, P. and Fuchs, E. (2003). Links between signal transduction, transcription and adhesion in epithelial bud development. Nature 422, 317-322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlsson, L., Bondjers, C. and Betsholtz, C. (1999). Roles for PDGF-A and sonic hedgehog in development of mesenchymal components of the hair follicle. Development 126, 2611-2621. [DOI] [PubMed] [Google Scholar]
- Kashiwagi, M., Kuroki, T. and Huh, N. (1997). Specific inhibition of hair follicle formation by epidermal growth factor in an organ culture of developing mouse skin. Dev. Biol. 189, 22-32. [DOI] [PubMed] [Google Scholar]
- Kolev, V., Mandinova, A., Guinea-Viniegra, J., Hu, B., Lefort, K., Lambertini, C., Neel, V., Dummer, R., Wagner, E. F. and Dotto, G. P. (2008). EGFR signalling as a negative regulator of Notch1 gene transcription and function in proliferating keratinocytes and cancer. Nat. Cell Biol. 10, 902-911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laurikkala, J., Pispa, J., Jung, H. S., Nieminen, P., Mikkola, M., Wang, X., Saarialho-Kere, U., Galceran, J., Grosschedl, R. and Thesleff, I. (2002). Regulation of hair follicle development by the TNF signal ectodysplasin and its receptor Edar. Development 129, 2541-2553. [DOI] [PubMed] [Google Scholar]
- Mann, G. B., Fowler, K. J., Gabriel, A., Nice, E. C., Williams, R. L. and Dunn, A. R. (1993). Mice with a null mutation of the TGF alpha gene have abnormal skin architecture, wavy hair, and curly whiskers and often develop corneal inflammation. Cell 73, 249-261. [DOI] [PubMed] [Google Scholar]
- Miettinen, P. J., Berger, J. E., Meneses, J., Phung, Y., Pedersen, R. A., Werb, Z. and Derynck, R. (1995). Epithelial immaturity and multiorgan failure in mice lacking epidermal growth factor receptor. Nature 376, 337-341. [DOI] [PubMed] [Google Scholar]
- Millar, S. E. (2002). Molecular mechanisms regulating hair follicle development. J. Invest. Dermatol. 118, 216-225. [DOI] [PubMed] [Google Scholar]
- Mou, C., Jackson, B., Schneider, P., Overbeek, P. A. and Headon, D. J. (2006). Generation of the primary hair follicle pattern. Proc. Natl. Acad. Sci. USA 103, 9075-9080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murillas, R., Larcher, F., Conti, C. J., Santos, M., Ullrich, A. and Jorcano, J. L. (1995). Expression of a dominant negative mutant of epidermal growth factor receptor in the epidermis of transgenic mice elicits striking alterations in hair follicle development and skin structure. EMBO J. 14, 5216-5223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustonen, T., Ilmonen, M., Pummila, M., Kangas, A. T., Laurikkala, J., Jaatinen, R., Pispa, J., Gaide, O., Schneider, P., Thesleff, I. et al. (2004). Ectodysplasin A1 promotes placodal cell fate during early morphogenesis of ectodermal appendages. Development 131, 4907-4919. [DOI] [PubMed] [Google Scholar]
- Nanney, L. B., Stoscheck, C. M., King, L. E., Jr, Underwood, R. A. and Holbrook, K. A. (1990). Immunolocalization of epidermal growth factor receptors in normal developing human skin. J. Invest. Dermatol. 94, 742-748. [DOI] [PubMed] [Google Scholar]
- Nowak, J. A., Polak, L., Pasolli, H. A. and Fuchs, E. (2008). Hair follicle stem cells are specified and function in early skin morphogenesis. Cell Stem Cell 3, 33-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohuchi, H., Tao, H., Ohata, K., Itoh, N., Kato, S., Noji, S. and Ono, K. (2003). Fibroblast growth factor 10 is required for proper development of the mouse whiskers. Biochem. Biophys. Res. Commun. 302, 562-567. [DOI] [PubMed] [Google Scholar]
- Panteleyev, A. A., Mitchell, P. J., Paus, R. and Christiano, A. M. (2003). Expression patterns of the transcription factor AP-2alpha during hair follicle morphogenesis and cycling. J. Invest. Dermatol. 121, 13-19. [DOI] [PubMed] [Google Scholar]
- Payre, F., Vincent, A. and Carreno, S. (1999). ovo/svb integrates Wingless and DER pathways to control epidermis differentiation. Nature 400, 271-275. [DOI] [PubMed] [Google Scholar]
- Petiot, A., Conti, F. J., Grose, R., Revest, J. M., Hodivala-Dilke, K. M. and Dickson, C. (2003). A crucial role for Fgfr2-IIIb signalling in epidermal development and hair follicle patterning. Development 130, 5493-5501. [DOI] [PubMed] [Google Scholar]
- Peus, D. and Pittelkow, M. R. (1996). Growth factors in hair organ development and the hair growth cycle. Dermatol. Clin. 14, 559-572. [DOI] [PubMed] [Google Scholar]
- Piepkorn, M., Underwood, R. A., Henneman, C. and Smith, L. T. (1995). Expression of amphiregulin is regulated in cultured human keratinocytes and in developing fetal skin. J. Invest. Dermatol. 105, 802-809. [DOI] [PubMed] [Google Scholar]
- Piepkorn, M., Pittelkow, M. R. and Cook, P. W. (1998). Autocrine regulation of keratinocytes: the emerging role of heparin-binding, epidermal growth factor-related growth factors. J. Invest. Dermatol. 111, 715-721. [DOI] [PubMed] [Google Scholar]
- Pummila, M., Fliniaux, I., Jaatinen, R., James, M. J., Laurikkala, J., Schneider, P., Thesleff, I. and Mikkola, M. L. (2007). Ectodysplasin has a dual role in ectodermal organogenesis: inhibition of Bmp activity and induction of Shh expression. Development 134, 117-125. [DOI] [PubMed] [Google Scholar]
- Rhee, H., Polak, L. and Fuchs, E. (2006). Lhx2 maintains stem cell character in hair follicles. Science 312, 1946-1949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt-Ullrich, R. and Paus, R. (2005). Molecular principles of hair follicle induction and morphogenesis. BioEssays 27, 247-261. [DOI] [PubMed] [Google Scholar]
- Schneider, M. R., Werner, S., Paus, R. and Wolf, E. (2008). Beyond wavy hairs: the epidermal growth factor receptor and its ligands in skin biology and pathology. Am. J. Pathol. 173, 14-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh, A. B. and Harris, R. C. (2005). Autocrine, paracrine and juxtacrine signaling by EGFR ligands. Cell. Signal. 17, 1183-1193. [DOI] [PubMed] [Google Scholar]
- St-Jacques, B., Dassule, H. R., Karavanova, I., Botchkarev, V. A., Li, J., Danielian, P. S., McMahon, J. A., Lewis, P. M., Paus, R. and McMahon, A. P. (1998). Sonic hedgehog signaling is essential for hair development. Curr. Biol. 8, 1058-1068. [DOI] [PubMed] [Google Scholar]
- Suzuki, K., Yamanishi, K., Mori, O., Kamikawa, M., Andersen, B., Kato, S., Toyoda, T. and Yamada, G. (2000). Defective terminal differentiation and hypoplasia of the epidermis in mice lacking the Fgf10 gene. FEBS Lett. 481, 53-56. [DOI] [PubMed] [Google Scholar]
- Trautman, M. S., Kimelman, J. and Bernfield, M. (1991). Developmental expression of syndecan, an integral membrane proteoglycan, correlates with cell differentiation. Development 111, 213-220. [DOI] [PubMed] [Google Scholar]
- Underhill, C. B. (1993). Hyaluronan is inversely correlated with the expression of CD44 in the dermal condensation of the embryonic hair follicle. J. Invest. Dermatol. 101, 820-826. [DOI] [PubMed] [Google Scholar]
- van Genderen, C., Okamura, R. M., Farinas, I., Quo, R. G., Parslow, T. G., Bruhn, L. and Grosschedl, R. (1994). Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev. 8, 2691-2703. [DOI] [PubMed] [Google Scholar]
- Wang, X., Bolotin, D., Chu, D. H., Polak, L., Williams, T. and Fuchs, E. (2006). AP-2alpha: a regulator of EGF receptor signaling and proliferation in skin epidermis. J. Cell Biol. 172, 409-421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wells, A. (1999). EGF receptor. Int. J. Biochem. Cell Biol. 31, 637-643. [DOI] [PubMed] [Google Scholar]
- Werner, S., Smola, H., Liao, X., Longaker, M. T., Krieg, T., Hofschneider, P. H. and Williams, L. T. (1994). The function of KGF in morphogenesis of epithelium and reepithelialization of wounds. Science 266, 819-822. [DOI] [PubMed] [Google Scholar]
- Wessells, N. K. and Roessner, K. D. (1965). Nonproliferation in dermal condensations of mouse vibrissae and pelage hairs. Dev. Biol. 12, 419-433. [DOI] [PubMed] [Google Scholar]
- Wilkinson, D. (1998). In Situ Hybridization: A Practical Approach. Oxford: Oxford University Press.
- Zhang, Y., Andl, T., Yang, S. H., Teta, M., Liu, F., Seykora, J. T., Tobias, J. W., Piccolo, S., Schmidt-Ullrich, R., Nagy, A. et al. (2008). Activation of beta-catenin signaling programs embryonic epidermis to hair follicle fate. Development 135, 2161-2172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zieske, J. D., Takahashi, H., Hutcheon, A. E. and Dalbone, A. C. (2000). Activation of epidermal growth factor receptor during corneal epithelial migration. Invest. Ophthalmol. Vis. Sci. 41, 1346-1355. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}