

Abstract
Human acyl-CoA oxidase 1 (ACOX1) is a rate-limiting enzyme in peroxisomal fatty acids β-oxidation and its deficiency is associated with a lethal, autosomal ressive disease, called pseudoneonatal-adrenoleukodystrophy. Two mRNA variants, transcribed from a single gene encode ACOX1a or ACOX1b isoforms respectively. Recently, a mutation in a splice site has been reported [1], which results in the defective peroxisomal fatty acids beta-oxidation. Here we show that these mRNA splice variants are expressed differentially in human liver. We investigated the biochemical role of the two human ACOX1 isoforms by heterologous expression of the catalytically active ACOX1a and ACOX1b enzymes in E. coli. ACOX1a seems to be more labile and exhibits only 50% specific activity toward palmitoyl-CoA as compared to ACOX1b.
Keywords: acyl-CoA oxidase, isoform, fatty acids, beta-oxidation, peroxisomes
Introduction
The absence of acyl-CoA oxidase, the rate-limiting enzyme in peroxisomal β-oxidation, is associated with the human disease, called pseudoneonatal-adrenoleukodystrophy, one of the nine identified types of peroxisomal single-enzyme deficiencies [1]. Several patients with ACOX1 deficiency have been reported. Although only few data are available in the literature on the molecular basis of acyl-CoA oxidase deficiency, one patient had a large deletion in the ACOX1 gene, whereas another patient had a single point mutation, resulting in both cases in enzymatically inactive acyl-CoA oxidases with high plasma levels of VLCFA [2]. Recently, a new patient was identified with a mutation in a splice site resulting in skipping of exon 4 in the mature mRNA and revealing a peroxisomal beta-oxidation deficiency in VLCFA [3]. Furthermore, the homozygous ACOX1 null mice have been generated. They are viable, but growth-retarded, infertile and developed a severe hepatic steatosis [4]. The human acyl CoA oxidase gene is located on chromosome 17 and consists of 15 exons and 14 introns, with two exons 3 named 3a and 3b [5]. An alternative splicing of the pre-mRNA leads to the synthesis of two mature mRNAs which encode two ACOX1 isoforms: ACOX1a or ACOX1b [6]. Each mRNA encodes for a polypeptide of 72 kDa (660 aminoacids), which contains a C-terminal peroxisomal targeting signal SKL [7]. It is noteworthy that the occurrence of two isoforms for ACOX1 is conserved through evolution including zebrafish [8], rodents [9] and human [5], and that alternative exons 3a and 3b each code for a 59 aminoacids region located near to the N-terminus [5, 9].
The active enzyme is a dimer of 140 kDa [10], which contains one molecule of a non-covalently bound Flavin Adenine Dinucleotide (FAD) per subunit as prosthetic group. Like mitochondrial acyl-CoA dehydrogenases, ACOX1 catalyzes theα, β-dehydrogenation of acyl-CoA esters, producing the corresponding trans-2-enoyl-CoA esters and FADH2 in the reductive half-reaction. Re-oxidation of the reduced FAD requires molecular oxygen and produces hydrogen peroxide as product. While the mitochondrial acyl-CoA dehydrogenases donate their electrons directly to the mitochondrial respiratory chain, the peroxisomal ACOX activity is considered to participate in cellular respiration and the production of heat [11].
Thus, the occurrence of two isoforms of ACOX1 could be physiologically important for the substrate specificity and selectivity handling for the peroxisomal β-oxidation activities. To investigate the biochemical role of each human ACOX1 isoform, we constructed the full-length human liver cDNAs encoding ACOX1a or ACOX1b respectively and expressed them as catalytically active Histidine-tagged recombinant proteins in E. coli. Subsequently we characterized the recombinant isoforms and determined optimal activities in terms of pH, temperature and substrate specificity.
Materiel and methods
mRNA expression of the two human ACOX1 splice variants by RT-PCR
Reverse transcriptase reaction was carried-out using 2 μg of total human liver RNA and the superscript III first strand synthesis kit as recommended by the supplier at 55°C for 50 min in 20 μl reaction volume containing 50μM oligo dT, 10 mM dNTP, 0.1M DTT, 25mM MgCl2 40u/μl RNase-out and 200U/μl of the superscript III enzyme. 1:15 dilution of the RT reaction was used for quantitative real-time in 20 μl reaction volume containing 10 μl of 2X SYBR Green Supermix (Applied Biosystems), 1μl of forward specific ACOX1a or ACOX1b primer and 1μl of common reverse ACOX1 primer. Primers used were for qACOX1a forward 5′-TGCTCAGAAAGAGAAATGGC-3′, qACOX1b forward 5′-CCTCTGGATCTTCACTTGG-3′ and q-ACOX1ab Reverse 5′-TGGGTTTCAGGGTCATACG-3′ all primers were used at 10 μM. PCR Cycling was 95°C for 15 seconds, annealing at 55°C for 30 s and extension at 72°C for 30 seconds followed by dissociation step in ABI 7300 (Applied Biosystems). Two real-time PCR runs were performed in triplicate from 3 pooled human liver samples. Expected length sizes are 132 and 188 bp for the ACOX1a and ACOX1b respectively. The generation of specific PCR products was confirmed by dissociation analysis and relative gene expressions were normalized with 18S ribosomal RNA and measured using the comparative CT method.
Construction of recombinant ACOX plasmids
ACOX cDNAs were synthesized using total RNA isolated from HepG2 cells or from human liver (Ambion), oligo (dT) primers, MMLV reverse transcriptase, forward ACOXKpnI primer 5′-GTCGACGGTACCAACCCGGACCTGCGC-3′ and reverse ACOXHindIII primer 5′-GCCGCAAGCTTATCAGAGCTTGGACTGCAGTG-3′ according to the protocol of suppliers. PCR cycling was denaturation at 95°C for 60 s, annealing at 55°C for 30 s and extension at 72°C for 2.5 minutes for 35 cycles followed by 10 minutes at 72°C. PCR ACOX1 fragment of 2004 bp, was first cloned by T/A cloning into pGEMT easy vector and ApaLI digestion, which cut ACOX1b cDNA but not ACOX1a one, was used as a screening condition. After sequencing, each of ACOX cDNA was subcloned into KpnI-HindIII digested pTriEx-4 (Novagen). Positive clones of pTriEx-4-ACOX1a or pTriEx-4-ACOX1b were used to transform E. coli BL21(DE3) Rosetta (Novagen) or E. coli C41 competent cells respectively.
Expression and purification of his-tagged ACOX proteins
Recombinant proteins were induced by adding of 1mM IPTG to E. coli Luria Bertani broth cultures and further incubation at 23°C for BL21 strain harboring pTriEx-4-ACOX1a or 37°C for C41 strain harboring pTriEx-4-ACOX1b. The bacteria pellet were lyzed by addition of 12.5 ml of buffer A (50mM NaH2PO4, pH 8.0, 300mM NaCl, and 0.05% Tween 20) supplemented with 10mM imidazole, 25μM of FAD and 0.5mM PMSF. After three cycles of freezing thawing in liquid nitrogen and sonication 6×10s with 10 s pauses at 50 W. The pellet was harvested by centrifugation at 31,000 g for 30 minutes at 4°C. The filtred supernatant (0.45μ) was loaded on 1ml Ni-NTA column equilibrated with 1ml of buffer A. The column was washed three times with 1ml of buffer A containing 20mM imidazole. Proteins were eluted with 1ml of elution buffer A containing 250mM imidazole. Fractions were analyzed on SDS-PAGE and stored at −80 °C. All purification buffers were supplemented with 25 μM FAD to protect the enzyme of a loss of activity.
Enzymatic assay
For the pH and temperature optimum and for the temperature denaturation experiments, ACOX1 activity was assayed using UV-spectrophotometric based method [12]. The reaction mixture (500 μl) contained 50 mM of potassium phosphate buffer pH 7.5 (or 50 mM citrate, glycine-glycine or borate buffer for the determination of the pH optimum) and 25 μM FAD.
For the ACOX1’s isoforms acyl-CoA substrate specificity, the ACOX1 activity was assayed by the fluorimetric based assay [13, 14]. The reaction mixture (200 μl) contained 50 mM Tris buffer at pH 8.3, 0.75 mM homovanillic acid, 20 μg.ml-1 horseradish peroxidase, 0.025% Triton X-100, 20 μM delipidated BSA and acyl-CoA substrate as indicated. All reactions were started by the addition of 20 μl of a 0.625 μM enzyme solution.
Expression of his-tagged ACOX1 enzymes in COS-7 cells
COS-7 cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum. Cells on six-well plates were transiently co-transfected as indicated with pTriexACOX1a and/or pTriexACOX1b, and pCMV-bgalactosidase plasmid DNA using Exgen.
Results and Discussion
Differential mRNA splice variants expression of ACOX1 gene in human liver
In order to quantify the level of the ACOX1 gene splice forms, ACOX1a and ACOX1b, in hepatic cells, real time PCR was carried out with two primer pairs specific for each variant. ACOX1a splice mRNA is expressed 1.5 time more than ACOX1b mRNA (figure 1A). For the first time we show that both exon 3 variants are transcribed differentially in liver and that the pre-mRNA is alternatively spliced and expressed permitting the hepatic translation of both ACOX1 isoforms 1a and 1b. The significance of such differential expression is not elucidated yet.
Figure 1.
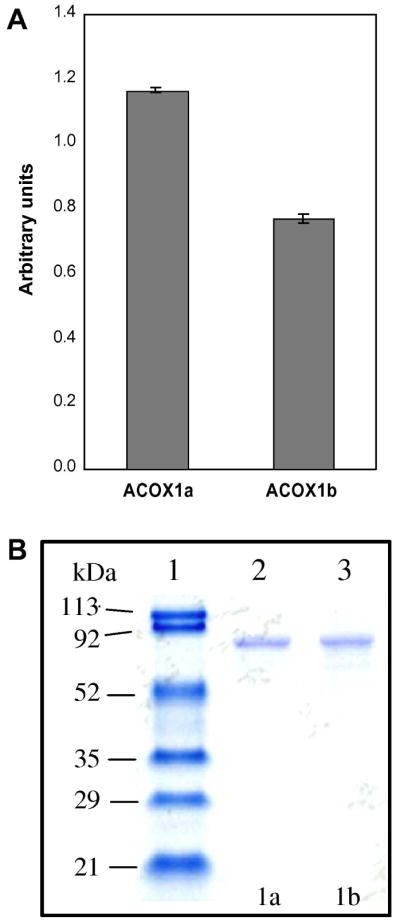
A, Human ACOX1a or ACOX1b mRNAs expression analysis by real time PCR. Messenger RNAs isolated from human liver were reverse transcribed and subjected to quantitative real time PCR using specific primers. Bars represent the relative level of ACOX1a and ACOX1b mRNA using the housekeeping 18S mRNA amplification as a reference gene. The fold change in gene expression in liver was calculated and presented as the mean ± S.D. of two real-time PCR runs performed in triplicate from 3 pooled total RNA samples of human liver (Ambion). B, SDS-PAGE analysis of purified recombinant human peroxisomal ACOX1a and ACOX1b proteins. Lane 1, molecular weight marker (MM) in kDa; Lane 2, ACOX1a (5μg); Lane 3, ACOX1b (5μg).
Cloning and E. coli expression of the human ACOX1a and ACOX1b
The two cDNAs encoding ACOX1a or ACOX1b were amplified by RT-PCR. Full-length amplified cDNAs were cloned in pGEMT plasmid and an ApaLI digestion was used to discriminate between the cDNA clones. The sequencing of the selected clones confirmed the cloning of ACOX1a and ACOX1b cDNAs individually. Each ACOX1 cDNA was further subcloned in the pTriEx-4 expression vector. ACOX1a and ACOX1b recombinant constructs harbor a 5′ histidine tag. Expression assays have shown a clear difference in the behavior of each isoform during the IPTG induced expression, which is dependent on temperature and on induction time of bacterial culture. Soluble and active ACOX1b is produced at 37°C in C41 strain, while ACOX1a is produced as a soluble and active cytosolic protein in BL21 E. coli strain at 23°C. Furthermore, both ACOX1 isoforms were purified by Ni-NTA affinity chromatography, resulting in a homogenous fraction as shown by SDS-PAGE analysis (Figure 1B) with a calculated molecular weight of 81 kDa for both ACOX1 isoforms including the polypeptide tag encoded by the pTriEx plasmid. The presence of FAD in purified ACOX1a and ACOX1b was verified by UV-visible absorption spectrum (data not shown).
Effect of pH on the activities of ACOX1a and ACOX1b
The specific activities of each ACOX1 isoform were determined between pH 6.5 and 10 using palmitoyl-CoA as substrate (Figure 2A). First, the optimum of palmitoyl-CoA oxidase activity for ACOX1a was measured for pH 8.5 while ACOX1b isoform presents an optimum pH ranging from 7.5 to 8.5. This difference in pH behavior could be due to the difference in the amino-acid surrounding of the catalytic glutamate 421 between the two isoforms. Secondly, ACOX1b is at least 2 times more active than ACOX1a. This may be explained by the fact that ACOX1 works in peroxisomes as a dimer [10, 15] and it is not known if the enzyme is present physiologically as two homodimers, one for each isoform, and/or as a heterodimeric form, composed of one chain of ACOX1a and one chain of ACOX1b. So, an ACOX1a homodimer, as produced in this study, is perhaps less active and stable than the heterodimeric counterpart. The existence of heterodimers in the cell is possible since the sequence identity between isoforms is very high (92%) and the presence of the two mRNAs of ACOX1 isoforms in human liver was observed (Figure 1A).
Figure 2.

A, effect of pH on the specific activities of ACOX1a and ACOX1b. Assays were carried out with 5 μg of enzyme purified and palmitoyl-CoA at 50 μM as substrate. Closed circles. B, temperature-dependent activation of ACOX1a and ACOX1b. Assays were carried out with 5 μg of purified enzyme and palmitoyl-CoA at 50 μM as substrate in 50 mM phosphate buffer, pH 7.5 at the indicated temperature. C, denaturation curves of ACOX1a and ACOX1b. Denaturation was started by incubation of purified enzyme in 50 mM phosphate buffer, pH 7.5 for 15 minutes at the indicated temperature followed by an activity assay with palmitoyl-CoA at 50 μM in 50 mM phosphate buffer, pH 7.5 at 30°C using 5 μg of the enzyme. Closed circles, ACOX1a; open circles, ACOX1b.
Effect of temperature on the activities of ACOX1a and ACOX1b
The temperature-dependent activation of acyl-CoA oxidase isoforms was measured at different temperatures in the assay at pH 7.5. The maximum specific activity of 0.1 U.mg−1 for ACOX1a is obtained at 40°C and drops to zero at 50°C, while the ACOX1b isoform has a maximum specific activity of 0.14 U.mg−1 around 37.5°C and retains 57% of its specific activity at 50°C (Figure 2B). Thus ACOX1b seems to be more active at supraphysiological temperature than ACOX1a even if the later shows 70% specific activity regarding ACOX1b at 37.5°C. The denaturation curves for both ACOX isoforms were obtained by incubation of each enzyme at indicated temperature for 15 minutes and then the activity of each sample was measured at 30°C. Figure 2C shows that ACOX1b is more resistant to heat denaturation than ACOX1a and conserves 30% of its specific activity after treatment at 50°C, while incubation of both isoforms between 0°C and 37°C has no effect on their specific activities except that ACOX1a exhibits only 50% specific activity of that of ACOX1b. The weaker stability of ACOX1a, clearly observed in both experiments brings up the question of whether the presence of the homodimer of this isoform is physiologically stable or indicates towards the existence of more stable heterodimers. This hypothesis is supported by purification protocols used for acyl-CoA oxidase from rat liver [10, 15] in which a 55°C heating step is carried out in order to aggregate contaminant proteins but not acyl-CoA oxidase.
Determination of the Km and the Vmax of ACOX1a and ACOX1b
Kinetics parameters Km and Vmax were determined for both ACOX1 isoforms using palmitoyl-CoA as substrate. We found a Km of 73 μM and a Vmax of 0.076 U.mg-1 for ACOX1a and a Km of 90 μM and a Vmax of 1.8 U.mg-1 for ACOX1b. Our calculated Km values are closer to that determined by Casteels et al. [16] in human liver homogenate at 62 μM. However, this value has been determined for a mixture of both ACOX1 isoforms present in the homogenate. Human ACOX1 expressed in insect cells showed an apparent Km of 10 μM [5] much lower than our calculated Km for each ACOX1 isoforms. This discrepancy, already noted by Chu et al. [6, 17], is probably due to the absence of BSA in the Chu’s assay conditions. Indeed, van Veldhoven et al. [8] have clearly showed that in vitro ACOX1 activity is BSA dependent.
Substrate specificity of ACOX1a and ACOX1b
Using a fixed concentration of 50 μM saturated fatty acyl-CoA with increasing chain-length from C-4 to C-18, we explored the activities of both ACOX1a and ACOX1b. As shown in Figure 3, both ACOX1a and ACOX1b present a broad range of specificity with maximum of specific activity with lauroyl-CoA (C-12) as substrate. ACOX1b presents a three times higher specific activity for octanoyl-CoA and decanoyl-CoA than ACOX1a. Setoyama et al. [18] have reported that rat ACOX I (counterpart of human ACOX1a) has an optimum of activity toward decanoyl-CoA (C-10) while rat ACOX II (counterpart of human ACOX1b) has an optimum of activity toward myristoyl-CoA (C-14). It is noteworthy that, as for rat ACOX II, a non-negligible activity (0.6 U.mg−1) is observed with octanoyl-CoA for ACOX1b whereas this molecule is thought to be the end product of peroxisomal β-oxidation which is exported from peroxisomes to mitochondria [19]. The slight difference in saturated straight chain fatty acyl-CoA specificity between isoforms of human and rat ACOX1 seems to indicate that both isoforms present a set of common substrates, straight chain fatty acyl-CoA, and that the difference in specificity between isoforms must be searched among fatty acids derivatives. Indeed, previous works related to the enzymatic activity of acyl-CoA oxidase did not look into the existence of the two isoforms and many diverse fatty acid derivatives have been shown as substrate for this peroxisomal pathway [19, 20]. Although mitochondria and peroxisomes possess their own β-oxidation systems, the peroxisomal β-oxidation process has surprisingly large panel of substrates, intervening in the chain shortening of variety of fatty acids derivatives, particularly in the catabolism of VLCFA, eicosanoid derivatives and dicarboxylic acids but also as the obligatory final step in the synthesis of docosahexadecanoic acid [20]. Table 1 shows the specific activities of recombinant human ACOX1a and ACOX1b for various CoA derivatives. The main difference is shown with 16-hydroxy-palmitoyl-CoA, for which ACOX1b specific activity is twice the ACOX1b one. In addition, ACOX1b is 25% more active toward the 1,16-hexadecanodioyl-CoA than ACOX1a, knowing that this substrate has two carboxylic functions activated as co-enzyme A derivative. Each ACOX1 isoforms a or b shows a high specific activity toward the aromatic fatty acid, 6-phenyl-hexoanoyl-CoA (Table1).
Figure 3.

Straight chain acyl-CoA specificity of recombinant human ACOX1a and ACOX1b. Specific activities of ACOX1a (closed circles) and ACOX1b (open circles), measured as described in “Material & Methods”, were plotted against a number of atoms of carbon of acyl-CoA from C4 to C18. Values represent the mean of three assays (± SD). Assay contained acyl-CoA at 50 μM and 1 μg of ACOX1a or ACOX1b.
Table 1.
Specific activities of recombinant human ACOX1a and ACOX1b for various CoA derivatives. Assay contained acyl-CoA at 50 μM and 1 μg of ACOX1a or ACOX1b. Values represent the mean of three assays (± SD)
| Substrate | Specific activity (U.mg−1) ± SD | ||
|---|---|---|---|
| ACOX1a | ACOX1b | Percent ACOX1b/ACOX1a | |
| C20:5-CoA | 0.186 ± 0.011 | 0.226 ± 0.009 | 121,5 % |
| 4-Me-C9-CoA | 0.237 ± 0.004 | 0.278 ± 0.035 | 117,3 % |
| 16-OH-C16-CoA | 0.150 ± 0.005 | 0.298 ± 0.023 | 198,7 % |
| 4,8,12-trimethyl-C13-CoA | 0.336 ± 0.040 | 0.423 ± 0.090 | 125,9% |
| diC16-CoA | 0.372 ± 0.012 | 0.504 ± 0.030 | 135,5 % |
| 6-phenyl-C6-CoA | 0.699 ± 0.074 | 0.573 ± 0.027 | 81,9 % |
| C16:0-CoA | 0.076 ± 0.005 | 0.166 ± 0.075 | 218.4 % |
C20:5-CoA, eicosapentaenoyl-CoA; 4-Me-C9-CoA, 4-methyl-nonanoyl-CoA; 16-OH-C16-CoA, 16-hydroxy-palmitoyl-CoA; 4,8,12-trimethyl-C13-CoA, 4,8,12-trimethyl-tridecanoyl-CoA; diC16-CoA, 1,16-hexadecadioyl-CoA; 6-phenyl-C6-CoA, 6-phenyl-hexanoyl-CoA; C16:0-CoA, palmitoyl-CoA.
Expression of ACOX1a and ACOX1b in COS-7 cells
Figure 4 shows the overexpression of each isoform or the two isoforms together in COS-7 cells, using the same pTriEx-4 constructs. ACOX1 isoforms were successfully expressed in COS-7 cells (lanes 2 and 3, Figure 4) as revealed by immunoblotting with anti-ACOX1 antibody showing a band of 80 kDa. The latter corresponds to the predicted recombinant ACOX1 size. The corrected specific activity, with palmitoyl-CoA as substrate, was calculated by standardization relative to beta-galactosidase activity and to the amount of ACOX1 from transfected COS cells homogenate. Results show that corrected specific activities for ACOX1a and ACOX1b are similar (ratio ACOX1b/ACOX1a of 1.16) (lanes 2 and 3, Figure 4). By contrast, the co-expression of both isoforms in COS cells results in 50% less corrected specific activity compared to ACOX1b (ratio ACOX1b/ACOX1[a+b] of 2.02) (lanes 3 and 4, Figure 4). Thus, as for the above measured activities with recombinant ACOX1 isoforms, the cell-co-expressed ACOX1a and ACOX1b exhibit 50% corrected specific activity toward palmitoyl-CoA compared to the separated expression of each ACOX1 isoform. In human liver, the higher ACOX1a mRNA level compared to ACOX1b (figure 1A) would result in the production of ACOX1a homodimer and of ACOX1a/ACOX1b heterodimers. The expression in COS cells reveals that the homodimer of each ACOX1 isoform present a similar activity. But the heterodimer of ACOX1a/ACOX1b has 50% less activity that homodimers. Thus in human liver, the regulation of the ratio between ACOX1a-homodimer and ACOX1a/ACOX1b-heterodimer might determine the level of peroxisomal fatty acids beta-oxidation. Thus the production of functional and catalytically active human ACOX1 isoforms is a key step in exploring the substrate specificity and selectivity of each isoform vis-à-vis of a large panel of fatty acids derivatives.
Figure 4.

ACOX1 isoforms expression in COS-7 cells. Western blot analysis and corrected specific activity measurements in COS-7 crude extracts (100 μg proteins). ACOX activities were standardized by beta-galactosidase activity and corrected by the amount of ACOX1 expressed in COS-7 cells evaluated by densitometry. The ratio was calculated by dividing the value of corrected specific activity of lane 3 by the value of lane 2 or lane 4. Lane 1, control COS-7 cells transfected with pTriex4 empty vector; lanes 2, 3 and 4 COS-7 cells transfected with pTriEx-4-ACOX1a, pTriEx-4-ACOX1b or both vectors respectively; lane 5, control recombinant ACOX1b produced in bacteria with the pTriEx-4-ACOX1b vector.
Acknowledgments
David Oaxaca-Castillo was supported by a fellowship from CONACyT (Mexico) program. Aurore Vluggens and this project were supported by the European Union project “Peroxisome” LSHG-CT-2004-512018 and in part by the Regional Council of Burgundy and the MESR. We thank Dr. Sailesh Surapureddi (NIH/NIEHS) and Dr Ronald J. Wanders (University of Amsterdam) for the critical reading of the manuscript and giving helpful suggestions.
Abbreviations
- ACOX1
acyl-CoA oxidase 1
- ACOX1a
ACOX1 isoform a
- ACOX1b
ACOX1 isoform b
- VLCFA
very long chain fatty acids
- FAD
flavin adenine dinucleotide
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Wanders RJ. Peroxisomes, lipid metabolism, and peroxisomal disorders. Mol Genet Metab. 2004;83:16–27. doi: 10.1016/j.ymgme.2004.08.016. [DOI] [PubMed] [Google Scholar]
- 2.Funato M, Shimozawa N, Nagase T, Takemoto Y, Suzuki Y, Imamura Y, Matsumoto T, Tsukamoto T, Kojidani T, Osumi T, Fukao T, Kondo N. Aberrant peroxisome morphology in peroxisomal beta-oxidation enzyme deficiencies. Brain Dev. 2006;28:287–292. doi: 10.1016/j.braindev.2005.10.010. [DOI] [PubMed] [Google Scholar]
- 3.Rosewich H, Waterham HR, Wanders RJ, Ferdinandusse S, Henneke M, Hunneman D, Gartner J. Pitfall in metabolic screening in a patient with fatal peroxisomal beta-oxidation defect. Neuropediatrics. 2006;37:95–98. doi: 10.1055/s-2006-923943. [DOI] [PubMed] [Google Scholar]
- 4.Fan CY, Pan J, Chu R, Lee D, Kluckman KD, Usuda N, Singh I, Yeldandi AV, Rao MS, Maeda N, Reddy JK. Hepatocellular and hepatic peroxisomal alterations in mice with a disrupted peroxisomal fatty acyl-coenzyme A oxidase gene. J Biol Chem. 1996;271:24698–24710. doi: 10.1074/jbc.271.40.24698. [DOI] [PubMed] [Google Scholar]
- 5.Varanasi U, Chu R, Chu S, Espinosa R, LeBeau MM, Reddy JK. Isolation of the human peroxisomal acyl-CoA oxidase gene: organization, promoter analysis, and chromosomal localization. Proc Natl Acad Sci U S A. 1994;91:3107–3111. doi: 10.1073/pnas.91.8.3107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chu R, Varanasi U, Chu S, Lin Y, Usuda N, Rao MS, Reddy JK. Overexpression and characterization of the human peroxisomal acyl-CoA oxidase in insect cells. J Biol Chem. 1995;270:4908–4915. doi: 10.1074/jbc.270.9.4908. [DOI] [PubMed] [Google Scholar]
- 7.Subramani S, Koller A, Snyder WB. Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem. 2000;69:399–418. doi: 10.1146/annurev.biochem.69.1.399. [DOI] [PubMed] [Google Scholar]
- 8.Morais S, Knoll-Gellida A, Andre M, Barthe C, Babin PJ. Conserved expression of alternative splicing variants of peroxisomal acyl-CoA oxidase 1 in vertebrates and developmental and nutritional regulation in fish. Physiol Genomics. 2007;28:239–252. doi: 10.1152/physiolgenomics.00136.2006. [DOI] [PubMed] [Google Scholar]
- 9.Osumi T, Ishii N, Miyazawa S, Hashimoto T. Isolation and structural characterization of the rat acyl-CoA oxidase gene. J Biol Chem. 1987;262:8138–8143. [PubMed] [Google Scholar]
- 10.Van Veldhoven PP, Vanhove G, Assselberghs S, Eyssen HJ, Mannaerts GP. Substrate specificities of rat liver peroxisomal acyl-CoA oxidases: palmitoyl-CoA oxidase (inducible acyl-CoA oxidase), pristanoyl-CoA oxidase (non-inducible acyl-CoA oxidase), and trihydroxycoprostanoyl-CoA oxidase. J Biol Chem. 1992;267:20065–20074. [PubMed] [Google Scholar]
- 11.Brown LA, Baker A. Peroxisome biogenesis and the role of protein import. J Cell Mol Med. 2003;7:388–400. doi: 10.1111/j.1582-4934.2003.tb00241.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Osumi T, Hashimoto T, Ui N. Purification and properties of acyl-CoA oxidase from rat liver. J Biochem (Tokyo) 1980;87:1735–1746. doi: 10.1093/oxfordjournals.jbchem.a132918. [DOI] [PubMed] [Google Scholar]
- 13.Vamecq J. Fluorometric assay of peroxisomal oxidases. Anal Biochem. 1990;186:340–349. doi: 10.1016/0003-2697(90)90092-n. [DOI] [PubMed] [Google Scholar]
- 14.Brocard C, Es-Souni M, Ramirez LC, Latruffe N, Bournot P. Stimulation of peroxisomal palmitoyl-CoA oxidase activity by ciprofibrate in hepatic cell lines: comparative studies in Fao, MH1C1 and HepG2 cells. Biol Cell. 1993;77:37–41. doi: 10.1016/s0248-4900(05)80172-8. [DOI] [PubMed] [Google Scholar]
- 15.Inestrosa NC, Bronfman M, Leighton F. Purification of the peroxisomal fatty acyl-CoA oxidase from rat liver. Biochem Biophys Res Commun. 1980;95:7–12. doi: 10.1016/0006-291x(80)90696-8. [DOI] [PubMed] [Google Scholar]
- 16.Casteels M, Schepers L, Van Veldhoven PP, Eyssen HJ, Mannaerts GP. Separate peroxisomal oxidases for fatty acyl-CoAs and trihydroxycoprostanoyl-CoA in human liver. J Lipid Res. 1990;31:1865–1872. [PubMed] [Google Scholar]
- 17.Chu R, Usuda N, Reddy MK, Liu C, Hashimoto T, Alvares K, Rao MS, Reddy JK. Functional expression of rat peroxisomal acyl-CoA oxidase in Spodoptera frugiperda cells. Biochem Biophys Res Commun. 1994;200:178–186. doi: 10.1006/bbrc.1994.1431. [DOI] [PubMed] [Google Scholar]
- 18.Setoyama C, Tamaoki H, Nishina Y, Shiga K, Miura R. Functional expression of two forms of rat acyl-CoA oxidase and their substrate specificities. Biochem Biophys Res Commun. 1995;217:482–487. doi: 10.1006/bbrc.1995.2801. [DOI] [PubMed] [Google Scholar]
- 19.Wanders RJ, Waterham HR. Biochemistry of mammalian peroxisomes revisited. Annu Rev Biochem. 2006;75:295–332. doi: 10.1146/annurev.biochem.74.082803.133329. [DOI] [PubMed] [Google Scholar]
- 20.Ferdinandusse S, Denis S, Mooijer PA, Zhang Z, Reddy JK, Spector AA, Wanders RJ. Identification of the peroxisomal beta-oxidation enzymes involved in the biosynthesis of docosahexaenoic acid. J Lipid Res. 2001;42:1987–1995. [PubMed] [Google Scholar]