

Abstract
Documenting the development of the functional anatomy underlying error processing is critically important for understanding age-related improvements in cognitive performance. Here we used functional magnetic resonance imaging to examine time courses of brain activity in 77 individuals aged 8–27 years during correct and incorrect performance of an oculomotor task requiring inhibitory control. Canonical eye-movement regions showed increased activity for correct versus error trials but no differences between children, adolescents and young adults, suggesting that core task processes are in place early in development. Anterior cingulate cortex (ACC) was a central focus. In rostral ACC all age groups showed significant deactivation during correct but not error trials, consistent with the proposal that such deactivation reflects suspension of a “default mode” necessary for effective controlled performance. In contrast, dorsal ACC showed increased and extended modulation for error versus correct trials in adults, which, in children and adolescents, was significantly attenuated. Further, younger age groups showed reduced activity in posterior attentional regions, relying instead on increased recruitment of regions within prefrontal cortex. This work suggests that functional changes in dorsal ACC associated with error regulation and error-feedback utilization, coupled with changes in the recruitment of “long-range” attentional networks, underlie age-related improvements in performance.
Keywords: antisaccade, child development, cognitive control, fMRI, prefrontal cortex
Introduction
Elementary cognitive functions are in place in childhood (Diamond and Goldman-Rakic 1989). However, improvements in cognitive control extend into adolescence and young adulthood (Levin et al. 1991). Although much work has been done to identify brain changes underlying improved task performance as a whole (e.g., Fischer et al. 1997; Klein and Foerster 2001; Luna et al. 2004), little attention has been given to understanding developmental changes in the processing of errors. Nevertheless, the emergence of adult-level cognition is known to rely on the development of error-regulatory functions, particularly in the context of being able to voluntarily suppress responses to task-irrelevant stimuli (Williams et al. 1999; Luna et al. 2004). The antisaccade (AS) task (Hallett 1978) is an oculomotor test of such inhibitory control and serves as the task-context in which error processing is examined in the present study.
Correct AS performance requires inhibition of a prepotent saccade toward a briefly presented peripheral stimulus and a voluntary saccade toward the empty mirror location. Behaviorally, AS performance has been shown to improve from childhood, through adolescence, and into young adulthood (Fischer et al. 1997; Munoz et al. 1998; Klein and Foerster 2001; Kramer et al. 2005; Asato et al. 2006). Independent of age, errors typically take the form of a rapid saccade toward the peripheral stimulus, followed by a corrective saccade toward the intended target location, indicating that the task instruction was understood, but that insufficient voluntary control was exerted to suppress the initial automatic saccade. Thus, a considerable advantage of the AS task is that it permits accurate identification of error trials attributable to inhibitory failures rather than to any variety of distraction or inattention.
Prior imaging work in adult humans, consistent with extensive neurophysiological work in monkeys (e.g., Robinson and Goldberg 1978; Bruce and Goldberg 1985), has demonstrated that AS task performance produces robust activation in a network of regions including medial frontal cortex, dorsolateral prefrontal cortex (dlPFC), supplementary motor area (SMA), frontal eye fields (FEF), (lateral) posterior parietal cortex (PPC), striatum, and cerebellum (Luna and Sweeney 1999; Matsuda et al. 2004; Pierrot-Deseilligny et al. 2004; Brown et al. 2006). Error trials have also been shown to generate robust performance-related indices including error negativities (Nieuwenhuis et al. 2001) and increased blood oxygen level–dependent (BOLD) activity in regions of anterior cingulate cortex (ACC; Ford et al. 2005; Polli et al. 2005).
To date, developmental functional magnetic resonance imaging (fMRI) work using the AS task and others taxing inhibitory control has largely focused on blocked performance in which correct and error trials both contribute to signal change estimates (e.g., Luna et al. 2001; Tamm et al. 2002; Marsh et al. 2006), and/or has selectively examined BOLD signal changes associated with correctly performed trials compared with a related “task” baseline thought to control for lower level task components and assumed to be constant across development (e.g., Bunge et al. 2002; Durston et al. 2002). Consequently, extant findings related to the development of inhibitory control are variable and highly task dependent, and the development of error-related processing remains largely unexplored (but see Rubia et al. 2007).
In the present study we examine the development of inhibitory control and error processing in a large sample (n = 77) of 8–27 year olds using analytic techniques that permit description of the temporal course of activity change without making assumptions about the timing or shape of the hemodynamic response. Time courses of brain activity associated with correctly performed and incorrectly performed, but corrected, AS trials (i.e., error trials) were estimated relative to a “nontask” (constant term) baseline allowing visualization of developmental changes in comparison task responses. Rather than controlling for age-related differences in task difficulty and performance by using individual performance titration (necessitating use of different task parameters across age groups), or by comparing high performing younger participants with low performing older participants, we asked instead, what changes in brain activity associate with the emergence of adult-level performance on a uniform task?
Our approach is largely hypothesis driven. In addition to examining activity changes across the whole brain in an exploratory fashion, we interrogate functionally defined regions known to be implicated in oculomotor control, task-general control and error processing. Of the latter we give considerable emphasis to ACC and proximal regions thought to play a role in the monitoring and regulation of goal directed behavior (for reviews see Carter et al. 1999; Bush et al. 2000; Holroyd and Coles 2002; Botvinick et al. 2004).
Two broad hypotheses have dominated thought about ACC function. The first proposes that ACC detects errors as discrepancies between actual and intended or desired events (Shima and Tanji 1998; Gehring and Willoughby 2002; Holroyd and Coles 2002). The second suggests that dorsal ACC (dACC), in particular, detects and monitors conflict between mutually incompatible response processes such as between incorrect and correct responses (Carter et al. 1998; Botvinick et al. 2001). Both conceptualizations regard ACC function as permitting subsequent adaptive adjustment and, hence, performance optimization, often putatively mediated through dlPFC (MacDonald et al. 2000; Gehring and Fencsik 2001; Kerns et al. 2004). Indeed, the magnitude of error-related activity in ACC has been shown to predict both changes in performance (measured by response times) and the magnitude of activity in dlPFC on trials that immediately follow error commission (Kerns et al. 2004; Kerns 2006). However, descriptions of such associations have, to date, been largely limited to adults.
Evidence from work with humans and animals supports anatomical and physiological maturation of the ACC (Cunningham et al. 2002), possible increased activation of ACC (Adleman et al. 2002; Ladouceur et al. 2004), and increased dopaminergic connections and metabolism in ACC into early adulthood (Benes et al. 1996; Lambe et al. 2000). A primary goal of the present study was to delineate developmental changes in ACC activity associated with performance accuracy. Based on behavioral and electrophysiological findings we hypothesized that children and adolescents would show less differentiation between correct and error trials in ACC, particularly in dACC, versus adults. We expected that these differences would be observed against a backdrop of relatively age-invariant patterns of activity in canonical eye-movement regions thought to be fundamental to task performance, but that dlPFC would show age-related effects related to its close interconnection with dACC (see Taylor et al. 2007 for a review), its role in effortful controlled processing (MacDonald et al. 2000; Badre and Wagner 2004; Cole and Schneider 2007), and based on past developmental studies of cognitive control (e.g., Rubia et al. 2000; Luna et al. 2001; Bunge et al. 2002).
Specifically, we made 2 predictions about activity in dlPFC. First, we expected that activity in dlPFC would be increased for correct versus error trials based on prior results suggesting that neural computations in dlPFC contribute to the biasing or selection of task-appropriate response pathways, particularly when competing response pathways are more automatically available (Milham et al. 2003; Badre and Wagner 2004). Here, our assumption was that errors in part reflect failure to exert sufficient top-down control of this kind. Second, we hypothesized that activity in dlPFC would decrease with age, reflecting age-related improvements in the efficiency of instantiating appropriate response parameters which, we will suggest, may be supported by effective error-related feedback signaling originating in dACC.
Experimental Procedures
Participants
Ninety-eight individuals ranging in age from 8 to 27 years participated in accordance with University of Pittsburgh Institutional Review Board guidelines (28 adults aged 18–27 years; 35 adolescents aged 13–17 years; 35 children aged 8–12 years). No standards exist for defining child and adolescent age ranges, thus these groups were defined based on our past behavioral studies indicating differential cognitive performance (Luna et al. 2004). Participants were native English speakers reporting no history of neurological problems. Three were left-handed. Vision was normal or corrected to normal using magnet compatible glasses or contact lenses. Sixteen participants either failed to complete the study or produced data with sufficient artifacts to preclude further analysis. Four participants were excluded due to equipment-related failure. One adult participant performed at ceiling and was excluded. Thus, we report data from 77 participants (26 adults, 25 adolescents, 26 children). Participants were trained on the AS and prosaccade (PS) tasks in the behavioral laboratory within 3 months of scanning, at which time they also completed assessments of handedness (Annett 1967; Oldfield 1971) and IQ (Wechsler Abbreviated Scale of Intelligence [WASI]; The Psychological Corporation 1999). No significant differences in full-scale IQ were observed across age groups (see Table 1). Immediately prior to scanning, naïve participants spent approximately 15 min in a mock scanner to acclimate them to the MR environment (Rosenberg et al. 1997).
Table 1.
Participant characteristics
| Adults |
Adolescents |
Children |
||||
| Mean | SD | Mean | SD | Mean | SD | |
| Age | 20.81 | 2.79 | 15.32 | 1.63 | 10.50 | 1.39 |
| Handedness: | ||||||
| Annett Handedness Index | 8.64a | 6.48 | 9.83b | 1.93 | 9.11 | 4.79 |
| Edinburgh Handedness Survey | 17.14a | 12.06 | 17.67b | 4.38 | 17.08 | 8.30 |
| General intelligence: | ||||||
| WASI FSIQ | 112.77c | 9.19 | 108.29b | 7.89 | 107.46 | 11.12 |
Note: FSIQ; full-scale IQ.
Data from 4 participants were unavailable.
Data from 1 participant were unavailable.
Data from 5 participants were unavailable.
fMRI Data Acquisition
Data were acquired using a Siemens 3-Tesla MAGNETOM Allegra (Erlangen, Germany) system with a standard circularity-polarized head coil. Pillows and tape minimized head movement. Earplugs dampened scanner noise. The experimenter communicated with participants via a built-in intercom system. A PC (Dell Dimension 8200, Pentium 4, 2 GHz, Windows XP) running E-Prime software (Psychology Software Tools, Pittsburgh, PA) controlled stimulus display. An LCD projector projected stimuli onto a screen at the head of the scanner bore, viewable via a mirror attached to the head coil.
Structural images were acquired first using a sagittal magnetization-prepared rapid gradient-echo (MP-RAGE) T1-weighted sequence (repetition time [TR] = 1570 ms, echo time [TE] = 3.04 ms, flip angle a = 8°, inversion time = 800 ms, voxel size = 0.78125 × 0.78125 × 1 mm). Functional images were then acquired using an echo-planar sequence sensitive to BOLD contrast (T2*) (TR = 1.5 s, TE = 25 ms, flip angle = 70°, voxel size = 3.125 × 3.125 mm in-plane resolution; Kwong et al. 1992; Ogawa et al. 1992). Participants performed 4 functional runs (each, 6 min 15 s in duration), followed by up to 3 runs of an unrelated experiment. During each run, 29 contiguous 4-mm-thick axial images were acquired parallel to the anterior commisure–posterior commisure plane. The first 6 images in each run were discarded to allow stabilization of longitudinal magnetization.
Behavioral Paradigm
During each run, participants alternated between blocked periods of oculomotor task performance and blocked periods of fixation (Chawla et al. 1999; Donaldson et al. 2001; Velanova et al. 2003), performing the PS task during 1 task block, and the AS task during a second (see Fig. 1A). The order of task blocks was counterbalanced across runs (within participant) and across participants. Participants were explicitly told the order of tasks before each run commenced.
Figure 1.

Schematic depictions of run (A) and task trial (B) structures.
Each task block lasted 118.5 s and was preceded by a 3 s cue informing participants about the nature of the upcoming trials (either “Start LOOK-AWAY game” for AS blocks, or “Start LOOK-TOWARD game” for PS blocks). Twelve AS trials or twelve PS trials were presented in each task block, such that, across the 4 runs comprising the experiment, participants performed 48 AS and 48 PS trials. Intervals between trials were planned so that the time between trials (during which a white fixation cross-hair was presented) varied from 3 to 9 s (2–6 frames) and was more often shorter than longer (Dale and Buckner 1997; Dale 1999). This temporal jitter allowed separation of overlapping trial-related signal components and differed from trial to trial for each participant. Trial presentation was time-locked to the onset of successive whole-brain image acquisitions. Each task block ended with a 3 s “task end” cue, alerting participants that a long period of fixation would follow.
Schematic depictions of task trials (AS and PS) are presented in Figure 1B. The focus of this report, however, is limited to AS trials because PS error rates were so low as to warrant their exclusion from analysis. (Limited behavioral data associated with PS trials are, however, presented for comparison.) Each trial began with a 3-s colored fixation cross-hair (subtending ∼ 0.7° of visual angle) instructing participants to make a PS (green) or an AS (red). This was followed immediately by a 1.5-s peripheral saccade stimulus. No “gap” was interposed between the instruction cue and saccade stimulus to increase the probability of accurate performance in younger participants (Fischer and Weber 1997). Each target stimulus was a yellow spot, subtending ∼0.5°, presented on the horizontal meridian at 1 of 6 eccentricities (at ±3°, 6°, or 9°). Target location order was randomized within each task block. For PS trials, participants' task was to look toward the saccade stimulus. For AS trials, participants were instructed to inhibit saccades toward the saccade stimulus and to look toward the empty location directly opposite.
Eye Tracking
Eye movement measurements were obtained during scanning using a long-range optics eye-tracking system (Model R-LRO6, Applied Science Laboratories, Bedford, MA). Nine-point calibrations were performed at the beginning of the session and between runs as necessary. Eye-movement data were analyzed and scored offline using ILAB (Gitelman 2002) in conjunction with an in-house scoring suite. Eye movements following the presentation of target saccade stimuli were scored for performance accuracy (as correct, incorrect but corrected, incorrect and uncorrected, and scoring omissions for each trial type) and latency.
fMRI Data Preprocessing
Data were preprocessed to remove noise and artifacts. Motion was corrected within and across runs using a rigid-body rotation and translation algorithm (Snyder 1996). Image slices were realigned in time to the midpoint of the first slice using sinc interpolation. Data were normalized to a whole run modal magnitude of 1000 (Ojemann et al. 1997) and were resampled into a standardized atlas space using 2-mm isotropic voxels (see Maccotta et al. 2001, for details of the warping method). Atlas registration involved aligning each subject's T1-weighted image to a custom atlas-transformed target T1-weighted template using a series of affine transforms (Michelon et al. 2003).
Movement Analysis
Measures of head movement were obtained from the output of the rigid-body rotation and translation algorithm. Translations and rotations in the x, y, and z dimensions were averaged across frames and total root mean square (RMS) linear and angular precision measures were calculated for each run. Runs in which a participant's total RMS movement exceeded 1 mm or degree were excluded from further analysis. Four runs of data were included for 25 of the 26 adult participants, 24 of the 25 adolescents, and for 17 of the 26 child participants. The remainder contributed 3 runs. Values for included runs were averaged for each participant, and analysis of variance (ANOVA) were performed testing for differences between age groups.
fMRI Data Analyses
Preprocessed data were analyzed using the general linear model (Friston et al. 1995; Worsley and Friston 1995; Zarahn et al. 1997). Analyses were performed to estimate signal changes associated with correctly performed and incorrectly performed but corrected AS trials using fIDL software (Miezin et al. 2000; Ollinger, Corbetta, et al. 2001; Ollinger, Shulman, et al. 2001). Effects for all analyses are described in terms of percent signal change, defined as signal magnitude divided by the mean of the estimated constant term (1 per run).
Specifically, for each participant, the BOLD response to each trial type (i.e., correct responses, incorrect but corrected responses, and incorrect and uncorrected responses + scoring omissions for both AS and PS trials) was estimated by coding a different regressor (delta function) for each of the 15 time points (i.e., image acquisitions) immediately following each stimulus onset. Regressors were also coded to account for cues at the beginning and end of each task block, the linear trend and runwise constant terms (Donaldson et al. 2001; Visscher et al. 2003; Velanova et al. 2006). This procedure produced 1 time course estimate (over 15 time points) per voxel, per trial condition. No hemodynamic response shape was assumed. Time course values associated with AS trials were then entered into voxel-by-voxel ANOVA based on a mixed-effects model with subjects as a random factor. Significant activation at the voxel level was defined as a significant main effect of time (indicating significant modulation of signal) or a significant interaction between any factor and time (indicating differential modulation of signal across levels of the factor interacting with time; Schlaggar et al. 2002). Levels of time corresponded to frames of the measured time course. Correctly performed and incorrectly performed but corrected AS trials (i.e., correct trials and error trials) corresponded to levels of the response type factor. Age group served as a between subjects factor. z-maps, based on F-statistics, were then visualized. These exploratory statistical images were smoothed with a Gaussian filter of 8-mm full width at half maximum, and corrections for multiple comparisons and sphericity were applied. The criterion for the former was based on a previous simulation study using a Monte Carlo method which stipulated a cutoff of z = 3 at a 110-voxel extent to maintain a significance probability of P < 0.05 (Forman et al. 1995; McAvoy et al. 2001). No voxels in the age group by time interaction map survived corrections required to maintain an acceptable false positive rate. Thus, an additional analysis identifying age-group differences based on endpoints (i.e., voxels showing a significant time by “age-group” effect, where levels of the age-group factor were restricted to adults and children) was conducted (see Fair et al. 2006 for use of a similar approach).
Analyses in Regions of Interest
Functional regions of interest (ROIs) were identified by taking uncorrected (F)z-statistical images for the main effect of time, for the interaction of response type and time, and for the interaction of “age group” and time (based on endpoints), preblurring using a 4-mm hard-sphere kernel, and applying an automated (local extremum) search algorithm that located peaks exceeding a significance threshold of P < 0.0027 (i.e., z = 3). Peaks separated by less than 10 mm were consolidated by averaging coordinates. Regions were defined by including all voxels in the uncorrected images within a 10 mm radius of an identified peak, then excluding voxels that failed to pass multiple comparisons and sphericity corrections (i.e., that did not appear in the corrected exploratory image derived as described above). This process defined ROIs that changed activity based on the effects represented in the base statistical images (Burgund et al. 2006).
Regions associated with oculomotor control (SMA/preSMA, FEF, PPC, and putamen), task-general control (dlPFC), and error processing (medial frontal gyrus/rostral ACC [medFG/rACC] and dACC) were specified a priori as foci of interest (see Table 2). All a priori specified regions were derived from the main effect of time, except regions associated with error processing, which, because they were not represented in the main effect of time image, were derived based on the response type by time interaction. Regions derived from the main effect of time were prioritized because, although indicating modulation of signal, they are unbiased with respect to all effects of interest. A region in primary visual cortex (V1; Brodmann area [BA] 17, centered on 10, −80, 5) served as a control. Regionwise analyses based on mixed-effects models were performed across all voxels within each region to identify region-specific time courses and levels of significance for the effect of time, and for interactions of time with the response type and age-group factors. Sphericity corrected levels of significance are reported.
Table 2.
Regions of interest
| Region | Approximate locationa | Peak coordinates |
Reference papersb | ||
| x | y | z | |||
| SMA/preSMA | Medial frontal gyrus; BA 6/8 | −1 | 2 | 56 | 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 |
| Bilateral FEF | L precentral gyrus; BA 6 | −28 | −9 | 56 | |
| R precentral gyrus; BA 6 | 26 | −7 | 57 | ||
| R PPC | R (lateral) PPC; BA 7/39 | 31 | −52 | 46 | |
| Bilateral striatum | L putamen | −22 | 2 | 7 | |
| R putamen | 21 | 3 | 10 | ||
| medFG/rACC | Cingulate cortex; BA 24/32 | −3 | 40 | 17 | 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 |
| dACC | Cingulate cortex; BA 32 | −2 | 22 | 38 | |
| R dlPFC | R middle and precentral frontal gyrus; BA 9/46/6 | 47 | 1 | 36 | 23, 24, 25, 26, 27, 28, 29, 30, 31, 32 |
Regions are named based on their approximate BA in the Talairach and Tournoux (1988) atlas.
Selected reference papers motivating interest in the regions: 1. Brown et al. 2006; 2. Bruce and Goldberg 1985; 3. Curtis and D'Esposito 2003; 4. Ettinger et al. 2007; 5. Ford et al. 2005; 6. Luna and Sweeney 2004; 7. Luna et al. 2001; 8. Matsuda et al. 2004; 9. Miller et al. 2005; 10. Muri et al. 1996; 11. O'Driscoll et al. 1995; 12. Pierrot-Deseilligny et al. 2004; 13. Botvinick et al. 2004; 14. Brown and Braver 2005; 15. Carter et al. 1998; 16. Gehring and Fencsik 2001; 17. Holroyd et al. 2004; 18. Ito et al. 2003; 19. Kerns 2006; 20. Pierrot-Deseilligny et al. 2004; 21. Polli et al. 2005; 22. Raichle et al. 2001; 23. Badre and Wagner 2004; 24. Bunge et al. 2002; 25. Cole and Schneider 2007; 26. Durston et al. 2006; 27. Koechlin et al. 2003; 28. MacDonald et al. 2000; 29. Miller and Cohen 2001; 30. Pierrot-Deseilligny et al. 2005; 31. Rubia et al. 2006; 32. Tamm et al. 2002.
Additional analyses were then conducted using methods described above (though nonselectively) to identify regions derived from the main effect of time image and from images corresponding to the interactions of response type and time, and “age-group” (endpoints) and time in which time courses of activity changed with age. Specifically, we identified regions where time courses of activity for correct and error trials showed parallel developmental trajectories (i.e., regions where interactions of age group and time were observed in the absence of any higher order interaction with the response type factor), and regions where time courses of activity for correct and error trials showed differing developmental trajectories (i.e., regions where interactions of age group, time, and response type were observed). These are presented in Table 3.
Table 3.
Regions showing significant age-related effects
| Region source map | Effect | Approximate location | Peak atlas coordinates |
n voxels | z at peak | P(effect) | Directiona | Orderb | ||
| x | y | z | ||||||||
| Time | Age group × time (parallel trajectories) | R Frontal: precentral and middle frontal gyrusc | 47 | 1 | 36 | 469 | 12.6 | 0.03 | + | C > T = Ad |
| R Frontal: precentral gyrus | 55 | −13 | 34 | 340 | 4.9 | 0.004 | + | C > T > A | ||
| R Middle frontal gyrus | 45 | 16 | 28 | 366 | 5.7 | 0.03 | + | C > T = A | ||
| R Superior frontal gyrus | 16 | 23 | 57 | 438 | 11.2 | 0.02 | − | C > T = A | ||
| L Inferior frontal gyrus | −48 | 15 | 22 | 431 | 7.7 | 0.05 | − | T > A = C | ||
| L Parietal: postcentral gyrus | −60 | −15 | 33 | 187 | 4.1 | 0.04 | −/+ | A = T > C | ||
| R Superior temporal gyrus | 40 | 7 | −14 | 219 | 4.8 | 0.01 | − | C > T > A | ||
| R Inferior occipital gyrus | 28 | −97 | −7 | 355 | 17.5 | 0.05 | − | A > T > C | ||
| Age group × time × response type (differential trajectories) | L Frontal: paracentral lobule | −6 | −27 | 57 | 444 | 11.7 | 0.04 | − | AS+e: T = C > A | |
| AS−: A = T > C | ||||||||||
| R Parietal: postcentral gyrus | 41 | −29 | 25 | 505 | 15.1 | 0.04 | − | AS+: T > A = C | ||
| AS−: T > A = C | ||||||||||
| L Parietal: postcentral gyrus | −31 | −29 | 63 | 469 | 12.1 | 0.03 | − | AS+: T = C > A | ||
| AS−: T = C > A | ||||||||||
| L Parietal: postcentral gyrus | −17 | −34 | 65 | 486 | 10.6 | 0.02 | − | AS+: T > A = C | ||
| AS−: T > A = C | ||||||||||
| R Parahippocampal gyrus | 36 | −31 | −10 | 372 | 7.7 | 0.05 | − | AS+: A = T = C | ||
| AS−: A > T = C | ||||||||||
| R Cerebellum: posterior lobe | 45 | −77 | −31 | 199 | 3.7 | 0.02 | + | AS+: C = T = A | ||
| AS−: C = T > A | ||||||||||
| Response type × time | Age group × time (parallel trajectories) | L Frontal: precentral gyrus | −57 | −9 | 36 | 212 | 3.8 | 0.002 | + | C > T > A |
| R Parietal: postcentral gyrus | 25 | −49 | 66 | 272 | 6.0 | 0.03 | − | T > A = C | ||
| R Parietal: postcentral gyrus | 36 | −42 | 61 | 300 | 5.4 | 0.004 | + | A > T > C | ||
| R Inferior parietal lobule | 38 | −43 | 46 | 281 | 4.6 | 0.02 | + | A > T > C | ||
| L Parietal: precuneus | −6 | −78 | 45 | 204 | 3.8 | 0.02 | + | A > T > C | ||
| L Occipital: lingual gyrus | −9 | −62 | 0 | 451 | 7 | 0.02 | + | A = T > C | ||
| R Occipital: cuneus | 12 | −62 | 8 | 425 | 5.0 | 0.01 | + | A = T > C | ||
| L Anterior insula | −32 | 15 | −5 | 324 | 4.3 | 0.003 | + | A = T > C | ||
| Age group × time × response type (differential trajectories) | L Cingulate gyrusc | −2 | 22 | 38 | 430 | 6.3 | 0.05 | + | AS+: C > A = T | |
| AS−: C > A > T | ||||||||||
| R Superior frontal gyrus | 3 | 16 | 61 | 368 | 5.2 | 0.04 | + | AS+: A = T = C | ||
| AS−: A > T = C | ||||||||||
| L Posterior insula | −49 | −39 | 20 | 355 | 5.6 | 0.04 | − | AS+: C = T = A | ||
| AS−: C > T = A | ||||||||||
| “Age group” by time (adults vs. children) | Age group × time (parallel trajectories) | L Medial frontal gyrus | −3 | 36 | 40 | 242 | 5.8 | 0.00001 | − | A > T = C |
| L Superior frontal gyrus | −3 | 27 | 54 | 189 | 5.1 | 0.00001 | − | A > T = C | ||
| R Superior frontal gyrus | 23 | 39 | 43 | 118 | 3.9 | 0.02 | − | C > T = A | ||
| L Middle frontal gyrus | −43 | 11 | 46 | 221 | 5.0 | 0.00001 | − | A > T = C | ||
| L Superior temporal gyrus | −44 | −4 | −8 | 213 | 5.0 | 0.00001 | − | C > T > A | ||
| L Superior temporal gyrus | −35 | 5 | −22 | 212 | 4.7 | 0.00001 | − | C > T = A | ||
| R Inferior parietal lobule | 49 | −57 | 45 | 168 | 4.1 | 0.001 | − | A > T > C | ||
| L Inferior parietal lobule | −39 | −68 | 43 | 171 | 5.1 | 0.0001 | − | A > T = C | ||
| L Parietal: precuneus | −5 | −75 | 43 | 119 | 3.9 | 0.03 | + | A > T > C | ||
| R Cerebellum: anterior lobe | 3 | −39 | −24 | 115 | 4.2 | 0.02 | + | C > T = A | ||
| Age group × time × response type (differential trajectories) | R Middle frontal gyrus | 48 | 22 | 33 | 115 | 4.2 | 0.03 | + | AS+: C > T > A | |
| AS−: C > T = A | ||||||||||
Refers to the direction of effects, that is, whether activity is primarily positive (+) or negative (−) at the maximal peak. −/+ Indicates time courses that show a negative peak and then a positive peak (that differ by >0.03% signal change).
Refers to the order of absolute peak levels of signal change, and not to statistical differences. Peaks are considered equal (=) when they differ by <0.03% signal change.
Region included in ROI analyses.
C; children, T; adolescents, A; adults.
AS+; correctly performed AS trials, AS−: AS error trials.
Exploratory Analyses over Time
In order to visualize the evolution of activity over time in each age group, voxelwise t-tests were conducted assessing the extent to which activity at time points 6, 7, and 8 differed from the baseline constant term for correctly performed AS trials, incorrectly performed but corrected AS trials, and for their difference. The resulting (t)z-statistical images were smoothed as above and corrected for multiple comparisons at a cutoff of z = 3 at a 99-voxel extent to maintain a significance probability of P < 0.05. Rendering of statistical images onto inflated cortical surfaces was achieved using CARET visualization software (http://brainmap.wustl.edu/caret; Van Essen et al. 2001).
Voxelwise Exploratory Analyses of Age-Group–Related Effects
To ensure that developmental effects were not overlooked, voxelwise (whole brain) exploratory maps were created comparing time courses of activity for each response type (separately) across paired combinations of age groups in mixed-effects ANOVA. Thus, voxels showing differential time courses of activity for correct AS trials for adults versus children, adolescents versus children, and adults versus adolescents were identified (i.e., voxels showing a significant interaction of “age-group pair” and time), as were voxels showing differential (“age-group pair” dependent) time courses for error trials. The resulting (F)z-statistical images were smoothed and corrected for multiple comparisons (as above).
For each age group, uncorrected (t)z-statistical images assessing the level of activity observed at time point 6 for each response type were also examined. Thresholds were lowered to an arbitrary minimum of z = 0.1 (i.e., P = 0.92). Time point 6 (at 7.5 s following trial onset) was selected for inspection based on its being the most representative time of peak activity across the whole brain. These (uncorrected and minimally thresholded) statistical images served to provide visual confirmation that interpreted results from other analyses were neither attributable to statistical threshold effects, nor insufficiently comprehensive.
Results
Movement Results
Consistent with prior reports (Schlaggar et al. 2002; Wenger et al. 2004), children exhibited significantly more head movement on average than did adults or adolescents, who did not differ (main effect of age group, F2,74 = 9.06, P < 0.001; t49 = 0.25, P > 0.8 for the uncorrected comparison of adults and adolescents). Nonetheless, average RMS movement for all age groups was considerably below the a priori selected cutoff of 1 at 0.26 for adults and adolescents, and 0.39 for children.
Behavioral Results
As expected, participants made significantly more AS than PS errors (0.37 vs. < 0.01, main effect of error type, F1,74 = 276.88, P < 0.0001). Over 93% of all AS errors were corrected. No significant difference in correction rates was observed across age groups (main effect of age group, F2,74 = 1.33, P = 0.27). Because uncorrected AS errors were excluded from imaging analyses, “AS errors” henceforth refers to corrected AS errors. Thus, children made significantly more AS errors than adolescents (with mean error rates of 48% vs. 32%), who made more errors than adults (22%; main effect of age group, F2,74 = 13.24, P < 0.0001 and pairwise group comparisons; t49 = 2.86, P < 0.01 for children vs. adolescents, t49 = 2.10, P < 0.05 for adolescents vs. adults). A highly significant linear relationship was observed between AS error rates and age (see Fig. 2A). Having excluded 1 adult participant who made no AS errors, no minimum number of AS errors was required to warrant a participant's inclusion in imaging analyses. Distributions of AS correct and AS error trials for each age group are provided in Supplementary Figure 1. PS performance was at ceiling (> 0.99 correct for all age groups) and hence, PS error rates did not differ by age group (interaction of error type by age group, F2,74 = 16.53, P < 0.0001; for analyses including PS trials only, main effect of age group, F2,74 = 0.96, P = 0.39).
Figure 2.
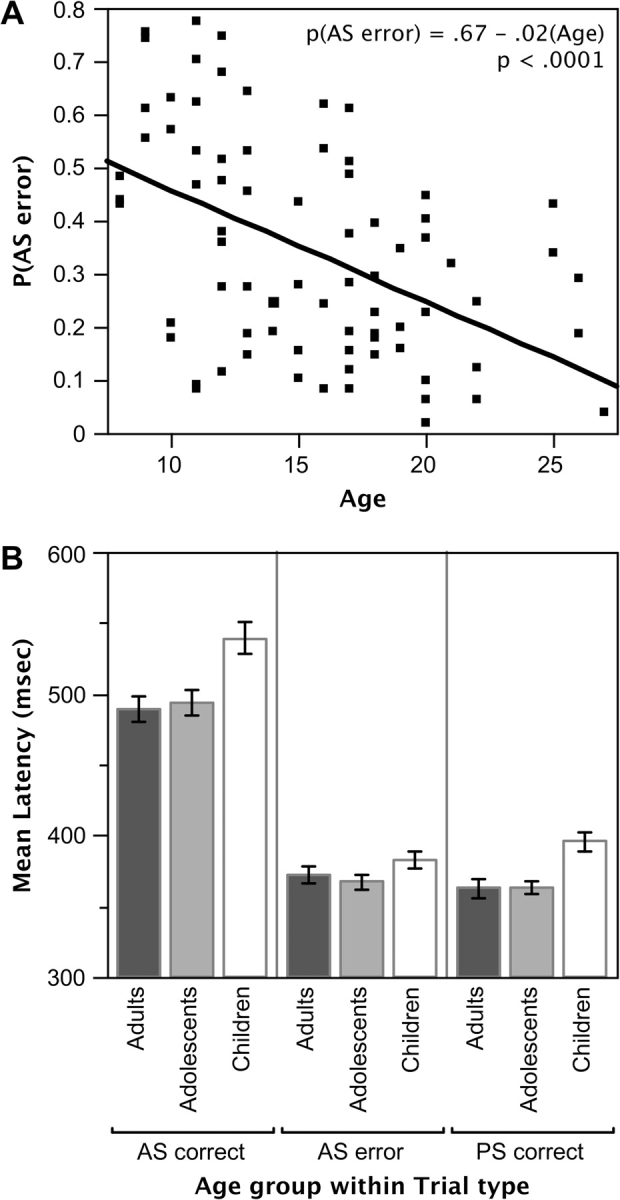
(A) Bivariate plot showing AS error rates as a function of age. (B) Mean initial saccade latencies for correctly and incorrectly performed AS trials and for correctly performed PS trials as a function of age group.
Latencies for initial saccades on correctly performed AS trials were significantly longer than for AS errors and PS trials (main effect of trial type; F2,148 = 299.31, P < 0.0001). Although AS errors and PS trials did not differ for adults and adolescents, children showed longer latencies on PS trials than on AS errors (interaction of age group and trial type, F2,151 = 6.26, P < 0.01 in comparisons of PS trials and AS errors only). Indeed, children were significantly slower overall than were older age groups (main effect of age group; F2,74 = 4.52, P < 0.05; see Fig. 2B). Nonetheless, mean latencies for all groups were within 1 repetition time (TR/whole brain image acquisition). The trial type by age-group interaction for latencies failed to reach significance (P = 0.13).
fMRI Results
Guided by previous work from our laboratory and established results in the literature, we identified 4 eye-movement-control–related ROIs in SMA/preSMA, FEF, PPC, and putamen. Regions within medFG/rACC and dACC were interrogated based on literatures describing their differing roles in correct and error trial performance. Right dlPFC was selected as an ROI based on prior findings of age-related differences during tasks requiring inhibitory control and its purported role in instantiating performance adjustments driven by ACC. Selected papers motivating interest in these regions are listed in Table 2. An ROI in primary visual cortex (V1) served as a control (Kang et al. 2003; Wenger et al. 2004).
Control Region in Primary Visual Cortex
We began by examining time courses for correct AS and AS error trials in V1. Given that the behavioral task produced different numbers and proportions of correct and error trials across age groups, this analysis served to establish that, despite these differences which might impact power, similar time courses could be obtained across age groups in a region thought to produce relatively age-invariant signal (Kang et al. 2003; Wenger et al. 2004). As anticipated, no age-related effects were observed (age group by time interaction, F28,1036 = 0.67, P = 0.91; age group by time by response type interaction, F28,1036 = 0.93, P = 0.57). Instead, error trials consistently produced greater modulation than did correct trials (response type by time interaction, F14,1036 = 5.51, P < 0.0001, modifying a main effect of time, F14,1036 = 99.35, P < 0.0001). This result, depicted in Supplementary Figure 2, is not surprising given that error trials are associated with greater visual input due to participants looking at the response stimulus then at the intended target location (versus only at the target location), and supports the veracity of our results in other regions. We do, however, acknowledge the possibility that age-invariance in V1 may simply be due to signal in this region being robust in the face of a variety of differences in subject performance. Nonetheless, we note that activity was not so robust as to preclude finding differences between correct and error trials, even in adults who produced a relatively small number of errors. Further, the observed age-invariance (here, beyond age 8) accords with findings from a number of other studies having less imbalance between conditions of interest (e.g., Kang et al. 2003; Wenger et al. 2004; Brown et al. 2005).
Eye-Movement Control Regions
Results replicated past findings indicating that SMA/preSMA, FEF, PPC, and striatum, notably putamen, increase activity during oculomotor tasks, particularly those requiring inhibitory control (see Table 2), and additionally demonstrated that this modulation is attenuated for AS errors. Figure 3 shows voxelwise activity observed in each age group and demonstrates considerable overlap in regions of significant activity across development. Time courses of activity in each of the a priori specified canonical eye-movement regions (defined using the main effect of time image) are presented in Figure 4. In each case, a significant interaction of response type and time was observed (F14,1036 = 2.3, P < 0.01 in SMA/PreSMA, and P < 0.0001 in all other ROIs) with increased activity observed for correct AS trials versus error trials. No significant age-group–related effects were observed, suggesting early maturation of core task execution regions.
Figure 3.
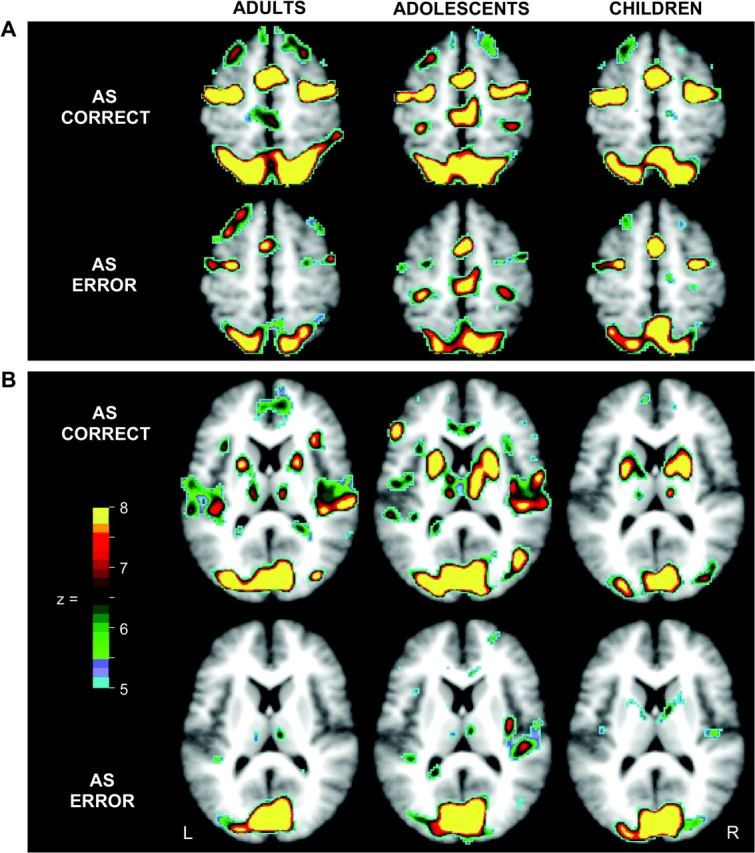
Voxels showing significant statistical activity change for correctly and incorrectly performed AS trials (i.e., correct and error trials) in adults, adolescents and children. All statistical images were corrected for multiple comparisons and sphericity (see Methods) and were then additionally thresholded at z = 5, P < 0.000001. (A) Horizontal sections at z = 54 show differing activity for correct and error trials, though similar distributions and levels of activity across age groups in SMA/preSMA, FEF, and PPC. (B) Horizontal sections at z = 12 show increased activity in putamen for correct trials relative to error trials in all age groups. Note that because maps are based on ANOVA, the direction of effects is not represented.
Figure 4.

Canonical eye-movement regions consistently showed increased activity for correct versus error trials in the absence of age-group effects. (A) A horizontal section shows the SMA/preSMA region from which time courses for adults, adolescents, and children were obtained for correct and error trials (depicted immediately to the right). (B–D) Regions in FEF, PPC, and putamen. Time courses are arranged similarly to those for (A). For (B), (C), and (D), depicted time courses are derived from circled regions, but do not differ in their (uncircled) contralateral homologues. Standard error bars are included at peaks for time courses in putamen (D) to emphasize the similarity of effects across age groups.
No significant age-related differences were observed in striatum (F28,1036 = 0.86, P > 0.67, right putamen, and F28,1036 = 0.98, P > 0.50, left putamen, for the interaction of age group and time, and, F28,1036 = 0.65, P > 0.92, right putamen, and F28,1036 = 0.55, P > 0.97, left putamen, for the interaction of age group, time, and response type) despite the apparently increased activity in younger age groups observed in exploratory maps. To more closely examine this result, maps comparing activity for correct and error trials (separately) for all pairwise combinations of groups were examined. In no case were significant voxels (representing differences between age groups) apparent in striatum, in accord with the null findings obtained in time course analyses.
medFG/rACC and dACC: Regions Implicated in Error Processing
medFG/rACC and dACC showed distinctly different patterns of activity (see Fig. 5A,B). In medFG/rACC, a significant response type by time interaction was observed (F14,1036 = 4.04. P < 0.00001). Whereas medFG/rACC showed significant negative modulation for correctly performed trials (F14,1036 = 7.22, P < 0.00001 for the main effect of time for correct trials analyzed independently), error time courses did not differ significantly from baseline (F14,1036 = 0.90, P = 0.56 (uncorrected) for error trials). No age-related effects were observed.
Figure 5.

Activity in ACC and dlPFC. (A and B) medFG/rACC and dACC showed dissociable patterns of activity. (A) A transverse section shows the medFG/rACC regions from which time courses for each age group were derived. Whereas time courses for error trials did not differ significantly from baseline, correct trials were associated with significant deactivations in all age groups. (B) A transverse section showing the dACC region. dACC showed significantly greater modulation during error versus correct trials, with adults showing greater differential activity than adolescents or children. Time points 6 (at 7.5 s following trial onset) and 7 (at 9 s following trial onset) are marked with vertical lines to distinguish early from late time points. For each age group, black asterisks mark the time point showing mean maximal peak activity for error trials. Gray asterisks mark the time point showing mean maximal differences in activity between error and correct trials. (C) Children showed increased activity in right dlPFC relative to adolescents and adults. Across age groups, correctly performed AS trials were associated with increased activity relative to AS errors. Panel layout parallels (A) and (B).
In contrast to findings in medFG/rACC, activity in dACC was positive. A response type by time interaction was again observed (F14,1036 = 6.08, P < 0.00001), as was an interaction of age group and time (F28,1036 = 1.80, P < 0.05). Error trials produced greater modulation than correct trials in all age groups (response type by time interactions; F14,350 = 1.77, P < 0.05 for children, F14,336 = 2.32, P < 0.01 for adolescents, and, F14,350 = 5.20, P < 0.00001 for adults), with children showing greater activation summed over levels of response type than adolescents (interaction of age group and time for children compared with adolescents, F14,686 = 2.30, P < 0.01) who differed from adults only for error trials (resulting in a significant interaction of age group, time, and response type for adolescents versus adults, F14,686 = 1.83, P < . 05). The full 3-way interaction of age group, response type, and time was also marginally significant (P = 0.10; but P < 0.05 uncorrected) attributable to adults showing greater differentiation between correct and error trials versus other age groups (response type by time interaction for adults, F14,150 = 5.20, P < 0.00001; for adolescents, F14,336 = 2.32, P < 0.01; for children, F14,350 = 1.77, P < 0.05). Time courses also suggested that the timing of peak activity and of peak differences between error and correct trials changed with age.
To explore these observations, the dependence of the time of peak activity for error trials and of the time of peak differences between error and correct trials on age was examined using chi-square. Although the time of peak activity for error trials (whether at time points 5 or 6 at 6–7.5 s following trial onset, or at time points 7 or 8, at 9–10.5 s following trial onset) failed to show significant dependence on age group (with 15 of 26 adults, 13 of 25 teens and 9 of 26 children showing greater activity at later versus earlier time points), such dependence was observed for the time of peak difference between error and correct trials (), with adults more likely to show a later time of peak difference (20 of 26) than adolescents (10 of 25) or children (8 of 26). We note that this effect can not be attributed to differences in behavioral response latencies across age groups, as children showed the longest response times. This result is also in accord with Polli et al.'s (2005) finding of late occurring differential activity in dACC in adults. Our data further demonstrate that this late differential recruitment of dACC is age-dependent.
In order to visualize this effect, and to confirm our replication and extension of Polli et al.'s (2005) findings, we follow their example and display the temporal evolution of voxelwise activity in medial cortex for adults, adolescents and children (see Fig. 6). Although late differential activation in dACC can be clearly observed in adults, the number of contiguous voxels in medFG/rACC showing (negative) differences between error and correct trials at each of the selected time points and in each (separate) age group was insufficient to pass stringent corrections for multiple comparisons and hence the clusters that drive our regionwise results in medFG/rACC are not displayed.
Figure 6.
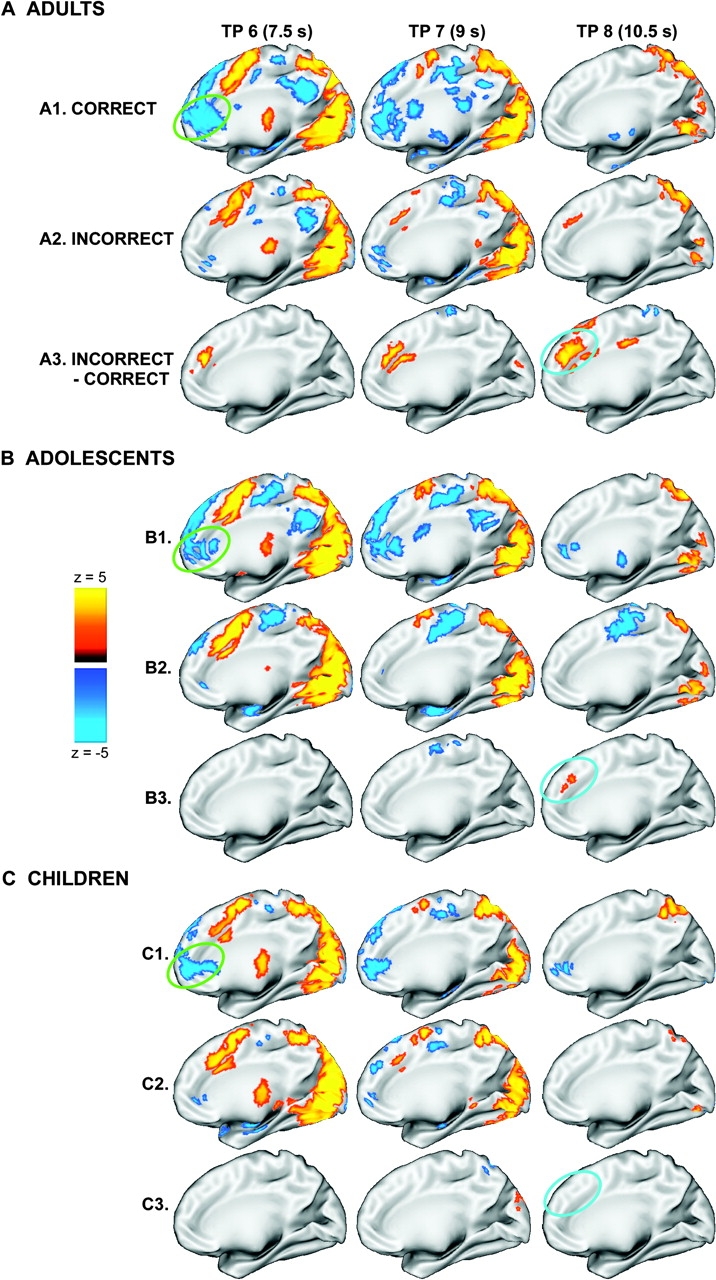
Findings of dissociable medFG/rACC and dACC activity across time for correct and error AS trials in each age group. (A) Statistical activation maps displayed on the partially inflated medial cortical surface of the right hemisphere for correctly performed AS trials (A1), AS errors (A2), and for AS errors—AS correct trials (A3) for adults at time points 6, 7, and 8. (B and C) Similarly arranged for adolescents and children. The approximate location of medFG/rACC is circled in green and dACC in blue. We note that effects in the left hemisphere paralleled those depicted in the right.
Effects in Prefrontal Cortex
A prefrontal region encompassing the dorsal extent of middle frontal gyrus near BA 9/46 and adjacent portions of BA 6 along the precentral gyrus is implicated in controlled processing and in instantiating trial-to-trial performance adjustments (see Table 2). One such region that encompassed several previously described regions of right middle dlPFC (e.g., van Veen et al. 2001; Bunge et al. 2002; Badre and Wagner 2004) was identified based on the main effect of time. Time courses of activity in this region are depicted in Figure 5C, revealing a significant interaction of time and age group (F28,1036 = 1.97, P < 0.03) attributable to children showing increased activity relative to adults and adolescents, who did not differ (age group by time interaction, F14,700 = 1.78, P = 0.05 for the direct comparison of adults versus children, and F14,686 = 3.51, P < 0.001 for adolescents versus children, but no such interaction for adults versus teens, F = 0.59, P = 0.87). The interaction of response type and time was also significant (F14,1036 = 3.90, P < 0.00001), with correct trials showing greater modulation than error trials. The full 3-way interaction of age group, response type, and time failed to reach significance (P > 0.21).
Exploratory Developmental Findings
To describe developmental changes at the systems level, we present summary findings from all regions (defined based on the main effect of time or an interaction between any factor and time, including regions defined based on the interaction of “age-group” [endpoints] and time) that showed both a significant main effect of time (indicating modulation) and an age-group–related effect (i.e., either a significant age group by time interaction or an interaction of age group, response type, and time). These are presented in Table 3. Although findings from regions derived from analyses comparing adults and children (endpoint age groups) are presented, findings from effects observed in regions defined based on the main effect of time and the interaction of response type and time are emphasized because these regions were 1) defined based on data from all participants, 2) typically of a size and location commensurate with anatomical areas, and 3) unbiased with respect to age-group effects.
Three exploratory findings are of note. First, no age-related effects were observed in canonical eye-movement regions (and hence these are not represented in Table 3). Second, prefrontal regions tended to show parallel effects for correct and error trials, such that the absolute magnitude of activation across levels of response type decreased with age, particularly in middle and superior frontal gyri where decreases stabilized in adolescence, and along precentral, gyrus where they continued to adulthood. Exceptions, however, were; 1) adjacent regions along the dorsal extent of medial frontal gyrus (extending into superior frontal gyrus) that showed increased negative activation in adults; and 2) a region in inferior frontal gyrus which showed maximal (negative) activity in adolescents that was then attenuated with age. (Beyond the bounds of prefrontal cortex, we also noted a similarly modulating swath of active regions extending from paracentral lobule along postcentral gyrus, bilaterally.) Third, with increasing age, an anterior to posterior shift was observed such that adults and adolescents showed increased recruitment of more posterior parietal and occipital regions relative to children. These observations are supported by whole-brain exploratory results depicted in Figure 7D–F. Figure 7 also indicates that there was considerable similarity in the location of activated foci across age groups, which we suggest is commensurate with the AS task permitting limited use of verbal strategies that might differ with age.
Figure 7.

Exploratory statistical images showing voxels that were differentially activated across age groups. (A1, B1, and C1) Voxels where time courses of activity for correctly performed AS trials differed for adults versus children, adolescents versus children, and adults versus adolescents, respectively. (A2, B2, and C2) Voxels where, across paired age groups, time courses differed for AS errors. Because these maps are based on ANOVA (i.e., are (F)z-statistical images), the direction of effects is not indicated. (D–F) (t)z-statistical images assessing the extent to which the estimated level of activity at time point 6 (at 7.5 s following trial onset) for correct and error trials, for each age group, differed from the baseline constant term. Time point 6 was selected as the most representative point of peak activity across the whole brain. No corrections were applied. The minimal threshold was arbitrarily set to z = ±0.1 to allow visualization of a relatively “complete” data set showing the direction of effects. (D1, E1, and F1) Effects for correct AS trials for adults, adolescents and children, respectively. (D2, E2, and F2) Effects for AS error trials. Arrows highlight activity in dlPFC.
Discussion
Performance of cognitive tasks requiring inhibitory control has frequently been shown to improve from childhood through young adulthood (Levin et al. 1991; Williams et al. 1999; Davies et al. 2004a, 2004b; Luna et al. 2004). The aim of the present study was to examine the functional anatomic correlates of such improvement in the context of an oculomotor task requiring inhibitory control. Critically, time courses of activity for correctly performed and incorrectly performed (but corrected) trials were assessed. Examination of error trials afforded an opportunity to identify regions implicated in inhibitory failure, and to specify developmental changes in activity that may signal the need for response adjustment on subsequent trial performance. In contrast to the bulk of developmental neuroimaging studies, we used an approach that required no assumptions be made about the timing or shape of the hemodynamic response and that permitted estimation of responses to task trials relative to a “nontask” (constant term) baseline. Relative to a “task” baseline, a constant term baseline is arguably less subject to inherent developmental change yet still permits task comparison. Both voxelwise and regionwise activity for correctly and incorrectly performed trials was compared across age groups, regardless of the direction of effects. Three primary results emerged from our data set: First, regions known to support the voluntary control of eye movements (e.g. trajectory programming, saccade suppression) showed greater activity during correctly performed AS trials than on error trials, but little developmental change. Second, dACC showed greater activity for error trials than for correct trials with the magnitude of that difference increasing and extending in time from childhood to adulthood—despite children showing significantly longer response latencies. Finally, children showed increased involvement of dlPFC relative to adolescents and adults, with an anterior to posterior shift evidenced with increasing age. Each of these results is discussed in turn below.
The Constancy of Canonical Oculomotor Regions
Single unit recording, lesion and imaging studies have revealed contributions from several cortical and subcortical regions to the generation of voluntary saccades. In particular, previous studies have indicated that saccadic eye-movements are controlled by a cortical network that includes FEF, SMA/preSMA, (lateral) PPC and striatum (Pierrot-Deseilligny et al. 1995; Pierrot-Deseilligny et al. 2004; Brown et al. 2006; Ettinger et al. 2007). In the present study, each of these regions was found to show greater activity for correct versus error trials, and to do so similarly across age groups. We interpret this finding as reflecting early maturation of core oculomotor control regions that contribute to correct trial performance.
Few studies have used functional imaging to examine the development of oculomotor control. In the single prior study using the AS task to examine the development of oculomotor control, Luna et al. (2001) observed progressive increases in activity in canonical eye-movement regions from childhood through adulthood. This study, however, used blocked methods, which confound activity associated with correctly and incorrectly performed trials. Given that the probability of correct inhibitory responses increases with age, and that correct trials associate with increased activity relative to error trials in these regions, the present data effectively predict such blocked results. In turn, these observations speak to the desirability of performance matching across age groups when blocked designs are used, and reaffirm the caution required in their interpretation. Primarily, however, our results demonstrate that when eliciting the same response, that is, a correct or incorrect inhibitory response, the same regions known to support cognitive control of eye movements are used across development.
Results in striatum provide a second case in point. The striatum (caudate nucleus, putamen and nucleus accumbens) receives inputs from all cortical areas, and projects principally to frontal regions concerned with motor planning, notably prefrontal and premotor cortex and SMA, via the thalamus (Evarts and Wise 1984; Alexander et al. 1986; Herrero et al. 2002). The striatum is known to have an important regulatory influence on cortex, and putamen and caudate, in particular, are thought to be involved in motor and eye-movement initiation and control, as well as playing a role the shifting of attentional sets and spatial working memory (Middleton and Strick 2000; Ravizza and Ciranni 2002; Monchi et al. 2006). In the present study, as hypothesized, robust activity was observed in striatum, largely localized to putamen. This result is in accord with prior reports (e.g., Petit et al. 1993; O'Driscoll et al. 1995), and is consistent with their characterization of putamen as a primary striatal target of cortical projections from the oculomotor circuit. Again, activity was greater for correct than for error trials, but no age-group effects were observed.
Prior developmental studies examining age-related changes in striatal activity during inhibitory task performance have produced mixed results (cf. Bunge et al. 2002; Durston et al. 2002; Booth et al. 2003; Rubia et al. 2006). In Luna et al.'s (2001) study of developmental change using the AS task, activity in a large predefined region of striatum including the head of the caudate, putamen and globus pallidus, was greater in adults and adolescents as compared with children. Again, however, this was in conjunction with significant performance differences across age groups. Other studies examining the development of inhibitory control have typically used go/no-go tasks or tasks heavily dependent on resistance to interference, such as Stroop and flanker tasks. Several of these studies have used event-related methods. Those using go/no-go tasks typically identify activity in the caudate body (rather than putamen) that increases from childhood to adulthood (e.g., Durston et al. 2002). Bunge et al.'s (2002) seminal event-related developmental study, however, reported robust activity in putamen for adults (but not children) during correct performance of incongruent versus neutral trials during flanker task performance. Yet, the extent to which results such as these are directly comparable to our own is uncertain, given that performance of the AS task requires not simply suppression of a response that is more frequently required in the experimental context, as for no-go trials, but suppression of a biologically given prepotent response in concert with purposefully enacting a response that is incompatible with its eliciting cue. Flanker and Stroop tasks also differ from the present task in that they emphasize the ability to inhibit distracting information, but require responses that are concordant with a given relevant cue or cue dimension. We contend, therefore, that prior eye-movement studies are the most satisfactory basis for comparison and that our data provide compelling preliminary evidence for the early maturation of both cortical and striatal regions implicated in successful eye-movement control.
Age-Related Changes in ACC
If activity in core oculomotor regions shows early maturation, what is the basis for improved performance with age? Our data support the hypothesis that improvements in error regulation subserved by dACC, play an important role in age-related performance enhancement.
ACC and related medial frontal regions have long been the central focus of research into error and conflict monitoring and performance adjustment (Carter et al. 1998; Bush et al. 2000; Taylor et al. 2007). In particular, dACC has been implicated in performance evaluation and error-related feedback processing (Carter et al. 1998; Gehring and Fencsik 2001). Several authors have argued that dorsal and rostral ACC are functionally dissociable (Devinsky et al. 1995; Polli et al. 2005; Margulies et al. 2007). Rostral portions of the ACC adjoining and overlapping with medFG are the proposed source of the error positivity in electrophysiological work (van Veen and Carter 2002), and are included in the default mode network in theories derived from fMRI and positron emission tomography research (Gusnard et al. 2001; Raichle et al. 2001; Greicius et al. 2003). Indeed, in their study of AS performance in young adults, Polli et al. (2005) demonstrated that dACC and rACC show temporally and directionally dissociable responses to correct and error trials. Specifically, these authors demonstrated that early bilateral deactivation of rACC was associated with correct trial performance, whereas AS errors were associated with late activation of dACC (as were a subset of voxels in rACC), consistent with dACC's putative role in performance evaluation and adjustment.
The present data replicate Polli et al.'s (2005) findings in adults, and extend them in developmental analyses. In particular, medFG/rACC showed task induced deactivation at early stages following trial onset for correct trials, but not for error trials, and did so similarly in all age groups. This result is consistent with the notion that deactivation in default mode regions is a requirement for accurate performance on demanding tasks. The absence of age-related effects suggests also that task induced deactivations may be associated with core cognitive processes that are in place early in development.
In contrast to findings in medFG/rACC, dACC showed increased activity for error trials versus correct trials, consistent with Polli et al. (2005) and with an extensive prior literature (Table 2). What is also demonstrated here is that differences between error and correct trials increase with age, and further, peak differences occur at later time points following trial onset in adults versus younger age groups and versus peaks observed in canonical oculomotor regions. Polli et al. and others have argued that such late recruitment in adults reflects dACC's contribution to the evaluation of error responses. Although neither the design employed by Polli et al. nor our own can definitively place late differential BOLD activity in dACC for error versus correct trials as occurring postresponse, our mutual conjecture that this is the case is supported by other recent studies. For example, Brown et al. (2006) used a compound trial paradigm permitting separation of activity associated with preparatory cue presentation and response cue presentation in the AS task. These authors, assuming separate modeled hemodynamic response functions for each trial phase, found increased activity in a region of ACC that shows considerable overlap with our own dACC region following response cue but not preparatory cue presentation on correct AS trials. (Here, incorrectly performed trials were excluded from analysis). Chevrier et al. (2007) more recently used a novel analytic technique to demonstrate that error-related activity in dACC during a stop signal task followed inhibitory failures, in accord with characterization of the region as being involved in postresponse monitoring, evaluation, and adjustment signaling (see also, Ploran et al. 2007). In the present data set, however, children and adolescents, while detecting errors at adult rates, failed to show fully mature responses of this sort. Indeed, our results are in accord with proposals emphasizing the monitoring and evaluation functions of ACC (e.g., Kerns et al. 2004; Suchan et al. 2007), rather than error or conflict “detection” per se. Although children and adolescents appear fully mature in their ability to recognize when they have made an error (as indicated by their correction rates), activity in dACC in these age groups differed from that in adults. To the extent that dACC provides a signal that can inform subsequent task performance, as a growing body of work indicates it does (for a review, see Ridderinkhof et al. 2004), our data suggest that the performance of children and adolescents receives less support from such feedback signaling, and hence implicates immature error regulation and error-feedback utilization as a source of performance decrements in younger age groups.
Activity in Frontal Cortex is Age Dependent
A considerable source of debate is the relative involvement of prefrontal cortex across development in controlled task performance. Several developmental studies have reported increased frontal activation in adults compared with children during tasks requiring inhibitory control (e.g., Rubia et al. 2000; Adleman et al. 2002; Bunge et al. 2002; Marsh et al. 2006; Rubia et al. 2007). These observations support a maturational hypothesis of brain function where brain regions thought to mature late are also brain regions that increase in function with age. Yet, other developmental imaging studies of inhibitory control have found increased frontal activation in children and adolescents compared with adults, interpreted as reflecting more diffuse and less specialized prefrontal function in younger age groups (e.g., Casey et al. 1997; Durston et al. 2002; Booth et al. 2003). Still other studies have identified different developmental effects in multiple prefrontal regions, such as decreased activity with increasing age in inferior frontal gyrus coupled with increased activity in middle frontal gyrus during tests of inhibitory control (Tamm et al. 2002). Further, studies examining different age ranges have revealed nonlinear developmental changes, such as increases from childhood to adolescence in middle frontal gyrus, followed by decreases to adulthood (Luna et al. 2001).
In the present study, activity observed in regions of prefrontal cortex was very broadly in accord with a “diffuse to specialized” hypothesis, though nonlinear trajectories and differing developmental patterns across regions were also readily evident (e.g., in [superior] medial frontal and inferior frontal cortex). However, basing our comments on effects observed in the a priori selected region of right dlPFC and in regions that were unbiased with respect to age-group effects, the absolute magnitude of BOLD responses in prefrontal cortex was typically increased in children relative to adolescents and adults. (Here we exclude canonical oculomotor regions and medFG/rACC). Further, this association most often held for both correct and error trials. Similar to adult studies showing increased activity in prefrontal cortex with increased task difficulty (e.g., Velanova et al. 2003; Wheeler and Buckner 2003; Geier et al. 2007), the present results could reflect the additional recruitment in younger subjects of task-general frontal control systems that permit, for example, improved task focus or that reflect additional processing required to manage task performance independent of trial accuracy. In particular we suggest that such additional processing in dlPFC might reflect “compensatory” control exerted to overcome inefficiency in the biasing of response pathways suitable for inhibiting a prepotent response.
Maturational Shift
Our results show that rather than using prefrontal cortex, adults increasingly engage regions that are supplemental to core regions known to support cognitive control. Although adults showed relatively decreased prefrontal activity, particularly in dorsolateral regions, they showed increased activity in occipital and parietal regions beyond postcentral gyrus (see Table 3 and Fig. 7, lower panels). This anterior to posterior shift with age implies a transition from reliance on task-general frontal systems to reliance on a more widely distributed circuitry that includes posterior attentional systems. These results accord with evidence of brain maturational events that continue into adolescence including synaptic pruning (Huttenlocher 1990; Huttenlocher and Dabholkar 1997) and myelination (Jernigan et al. 1991; Pfefferbaum et al. 1994) which both enhance computational abilities in local circuits and permit the integration of distributed neural systems. Developmental reductions in gray matter also continue through adolescence in cortical association areas, notably in frontal and temporal regions (Gogtay et al. 2004; Toga et al. 2006) and improvements in white matter integrity extend from frontal to parietal regions (Olesen et al. 2003). Finally these results are in accord with recent resting state functional connectivity results (Fair et al. 2007) demonstrating increases in long-range functional connectivity from childhood through adulthood particularly from dlPFC to PPC, simultaneous with decreases in short range connections, notably from anterior prefrontal cortex to dlPFC, that effectively segregated 2 putative control networks; 1 dominated by dACC and frontal operculum, and the other a frontoparietal network.
Thus, taken together our results support the hypothesis that developmental improvements in inhibitory control are supported both by enhancement of ACC's error-regulatory functions and by the recruitment of a more widely distributed circuitry that enables attentional and sensory regions, in addition to prefrontal executive regions, to assist with inhibitory control.
Supplementary Material
Supplementary material can be found at: http://www.cercor.oxfordjournals.org/
Funding
National Institutes of Mental Health (NIMH RO1 MH067924).
Acknowledgments
We thank Mark McAvoy and Abraham Snyder for support and development of functional data analysis procedures. Enami Yasui provided assistance with data collection.
Conflict of Interest: None declared.
References
- Adleman NE, Menon V, Blasey CM, White CD, Warsofsky IS, Glover GH, Reiss AL. A developmental fMRI study of the Stroop color-word task. Neuroimage. 2002;16:61–75. doi: 10.1006/nimg.2001.1046. [DOI] [PubMed] [Google Scholar]
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. doi: 10.1146/annurev.ne.09.030186.002041. [DOI] [PubMed] [Google Scholar]
- Annett M. The binomial distribution of right, mixed and left handedness. Q J Exp Psychol. 1967;19:327–333. doi: 10.1080/14640746708400109. [DOI] [PubMed] [Google Scholar]
- Asato MR, Sweeney JA, Luna B. Cognitive processes in the development of TOL performance. Neuropsychologia. 2006;44:2259–2269. doi: 10.1016/j.neuropsychologia.2006.05.010. [DOI] [PubMed] [Google Scholar]
- Badre D, Wagner AD. Selection, integration, and conflict monitoring; assessing the nature and generality of prefrontal cognitive control mechanisms. Neuron. 2004;41:473–487. doi: 10.1016/s0896-6273(03)00851-1. [DOI] [PubMed] [Google Scholar]
- Benes FM, Vincent SL, Molloy R, Khan Y. Increased interaction of dopamine-immunoreactive varicosities with GABA neurons of rat medial prefrontal cortex occurs during the postweanling period. Synapse. 1996;23:237–245. doi: 10.1002/(SICI)1098-2396(199608)23:4<237::AID-SYN1>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Lei Z, Trommer BL, Davenport ND, Li W, Parrish TB, Gitelman DR, Mesulam MM. Neural development of selective attention and response inhibition. Neuroimage. 2003;20:737–751. doi: 10.1016/S1053-8119(03)00404-X. [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychol Rev. 2001;108:624–652. doi: 10.1037/0033-295x.108.3.624. [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Cohen JD, Carter CS. Conflict monitoring and anterior cingulate cortex: an update. Trends Cogn Sci. 2004;8:539–546. doi: 10.1016/j.tics.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Brown JW, Braver TS. Learned predictions of error likelihood in the anterior cingulate cortex. Science. 2005;307:1118–1121. doi: 10.1126/science.1105783. [DOI] [PubMed] [Google Scholar]
- Brown MR, Goltz HC, Vilis T, Ford KA, Everling S. Inhibition and generation of saccades: rapid event-related fMRI of prosaccades, antisaccades, and nogo trials. Neuroimage. 2006;33:644–659. doi: 10.1016/j.neuroimage.2006.07.002. [DOI] [PubMed] [Google Scholar]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Developmental changes in human cerebral functional organization for word generation. Cereb Cortex. 2005;15:275–290. doi: 10.1093/cercor/bhh129. [DOI] [PubMed] [Google Scholar]
- Bruce CJ, Goldberg ME. Primate frontal eye fields. I. Single neurons discharging before saccades. J Neurophysiol. 1985;53:603–635. doi: 10.1152/jn.1985.53.3.603. [DOI] [PubMed] [Google Scholar]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Immature frontal lobe contributions to cognitive control in children: evidence from fMRI. Neuron. 2002;33:301–311. doi: 10.1016/s0896-6273(01)00583-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgund ED, Lugar HM, Miezin FM, Schlaggar BL, Petersen SE. The development of sustained and transient neural activity. Neuroimage. 2006;29:812–821. doi: 10.1016/j.neuroimage.2005.08.056. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci. 2000;4:215–222. doi: 10.1016/s1364-6613(00)01483-2. [DOI] [PubMed] [Google Scholar]
- Carter CS, Botvinick MM, Cohen JD. The contribution of the anterior cingulate cortex to executive processes in cognition. Rev Neurosci. 1999;10:49–57. doi: 10.1515/revneuro.1999.10.1.49. [DOI] [PubMed] [Google Scholar]
- Carter CS, Braver TS, Barch DM, Botvinick MM, Noll D, Cohen JD. Anterior cingulate cortex, error detection, and the online monitoring of performance. Science. 1998;280:747–749. doi: 10.1126/science.280.5364.747. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, Castellanos FX, Haxby JV, Noll DC, Cohen JD, et al. A developmental functional MRI study of prefrontal activation during performance of a Go-No-Go task. J Cogn Neurosci. 1997;9:835–847. doi: 10.1162/jocn.1997.9.6.835. [DOI] [PubMed] [Google Scholar]
- Chawla D, Rees G, Friston KJ. The physiological basis of attentional modulation in extrastriate visual areas. Nat Neurosci. 1999;2:671–676. doi: 10.1038/10230. [DOI] [PubMed] [Google Scholar]
- Chevrier AD, Noseworthy MD, Schachar R. Dissociation of response inhibition and performance monitoring in the stop signal task using event-related fMRI. Hum Brain Mapp. 2007;28:1347–1358. doi: 10.1002/hbm.20355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cole MW, Schneider W. The cognitive control network: integrated cortical regions with dissociable functions. Neuroimage. 2007;37:343–360. doi: 10.1016/j.neuroimage.2007.03.071. [DOI] [PubMed] [Google Scholar]
- Curtis CE, D'Esposito M. Success and failure suppressing reflexive behavior. J Cogn Neurosci. 2003;15:409–418. doi: 10.1162/089892903321593126. [DOI] [PubMed] [Google Scholar]
- Cunningham MG, Bhattacharyya S, Benes FM. Amygdalo-cortical sprouting continues into early adulthood: implications for the development of normal and abnormal function during adolescence. J Comp Neurol. 2002;453:116–130. doi: 10.1002/cne.10376. [DOI] [PubMed] [Google Scholar]
- Dale AM. Optimal experimental design for event-related fMRI. Hum Brain Mapp. 1999;8:109–114. doi: 10.1002/(SICI)1097-0193(1999)8:2/3<109::AID-HBM7>3.0.CO;2-W. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale AM, Buckner RL. Selective averaging of rapidly presented individual trials using fMRI. Hum Brain Mapp. 1997;5:329–340. doi: 10.1002/(SICI)1097-0193(1997)5:5<329::AID-HBM1>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- Davies PL, Segalowitz SJ, Gavin WJ. Development of error-monitoring event-related potentials in adolescents. Ann N Y Acad Sci. 2004a;1021:324–328. doi: 10.1196/annals.1308.039. [DOI] [PubMed] [Google Scholar]
- Davies PL, Segalowitz SJ, Gavin WJ. Development of response-monitoring ERPs in 7- to 25-year-olds. Dev Neuropsychol. 2004b;25:355–376. doi: 10.1207/s15326942dn2503_6. [DOI] [PubMed] [Google Scholar]
- Devinsky O, Morrell MJ, Vogt BA. Contributions of anterior cingulate cortex to behaviour. Brain. 1995;118(Pt 1):279–306. doi: 10.1093/brain/118.1.279. [DOI] [PubMed] [Google Scholar]
- Diamond A, Goldman-Rakic PS. Comparison of human infants and rhesus monkeys on Piaget's AB task: evidence for dependence on dorsolateral prefrontal cortex. Exp Brain Res. 1989;74:24–40. doi: 10.1007/BF00248277. [DOI] [PubMed] [Google Scholar]
- Donaldson DI, Petersen SE, Ollinger JM, Buckner RL. Dissociating state and item components of recognition memory using fMRI. Neuroimage. 2001;13:129–142. doi: 10.1006/nimg.2000.0664. [DOI] [PubMed] [Google Scholar]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella JA, Casey BJ. A shift from diffuse to focal cortical activity with development. Dev Sci. 2006;9:1–8. doi: 10.1111/j.1467-7687.2005.00454.x. [DOI] [PubMed] [Google Scholar]
- Durston S, Thomas KM, Worden MS, Yang Y, Casey BJ. The effect of preceding context on inhibition: an event-related fMRI study. Neuroimage. 2002;16:449–453. doi: 10.1006/nimg.2002.1074. [DOI] [PubMed] [Google Scholar]
- Ettinger U, Ffytche DH, Kumari V, Kathmann N, Reuter B, Zelaya F, Williams SC. Decomposing the neural correlates of antisaccade eye movements using event-related fMRI. Cereb Cortex. 2007 doi: 10.1093/cercor/bhm147. doi:10.1093/cercor/bhm147. [DOI] [PubMed] [Google Scholar]
- Evarts EV, Wise SP. Basal ganglia outputs and motor control. Ciba Found Symp. 1984;107:83–102. doi: 10.1002/9780470720882.ch6. [DOI] [PubMed] [Google Scholar]
- Fair DA, Brown TT, Petersen SE, Schlaggar BL. A comparison of analysis of variance and correlation methods for investigating cognitive development with functional magnetic resonance imaging. Dev Neuropsychol. 2006;30:531–546. doi: 10.1207/s15326942dn3001_2. [DOI] [PubMed] [Google Scholar]
- Fair DA, Dosenbach NU, Church JA, Cohen AL, Brahmbhatt S, Miezin FM, Barch DM, Raichle ME, Petersen SE, Schlaggar BL. Development of distinct control networks through segregation and integration. Proc Natl Acad Sci USA. 2007;104:13507–13512. doi: 10.1073/pnas.0705843104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer B, Biscaldi M, Gezeck S. On the development of voluntary and reflexive components in human saccade generation. Brain Res. 1997;754:285–297. doi: 10.1016/s0006-8993(97)00094-2. [DOI] [PubMed] [Google Scholar]
- Fischer B, Weber H. Effects of stimulus conditions on the performance of antisaccades in man. Exp Brain Res. 1997;116:191–200. doi: 10.1007/pl00005749. [DOI] [PubMed] [Google Scholar]
- Ford KA, Goltz HC, Brown MR, Everling S. Neural processes associated with antisaccade task performance investigated with event-related FMRI. J Neurophysiol. 2005;94:429–440. doi: 10.1152/jn.00471.2004. [DOI] [PubMed] [Google Scholar]
- Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA, Noll DC. Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): use of a cluster-size threshold. Magn Reson Med. 1995;33:636–647. doi: 10.1002/mrm.1910330508. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Holmes AP, Poline JB, Grasby PJ, Williams SC, Frackowiak RS, Turner R. Analysis of fMRI time-series revisited. Neuroimage. 1995;2:45–53. doi: 10.1006/nimg.1995.1007. [DOI] [PubMed] [Google Scholar]
- Gehring WJ, Fencsik DE. Functions of the medial frontal cortex in the processing of conflict and errors. J Neurosci. 2001;21:9430–9437. doi: 10.1523/JNEUROSCI.21-23-09430.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gehring WJ, Willoughby AR. The medial frontal cortex and the rapid processing of monetary gains and losses. Science. 2002;295:2279–2282. doi: 10.1126/science.1066893. [DOI] [PubMed] [Google Scholar]
- Geier CF, Garver KE, Luna B. Circuitry underlying temporally extended spatial working memory. Neuroimage. 2007;35:904–915. doi: 10.1016/j.neuroimage.2006.12.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gitelman DR. ILAB: a program for postexperimental eye movement analysis. Behav Res Methods Instrum Comput. 2002;34:605–612. doi: 10.3758/bf03195488. [DOI] [PubMed] [Google Scholar]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, Nugent TF, 3rd, Herman DH, Clasen LS, Toga AW, et al. Dynamic mapping of human cortical development during childhood through early adulthood. Proc Natl Acad Sci USA. 2004;101:8174–8179. doi: 10.1073/pnas.0402680101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius MD, Krasnow B, Reiss AL, Menon V. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis. Proc Natl Acad Sci USA. 2003;100:253–258. doi: 10.1073/pnas.0135058100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gusnard DA, Akbudak E, Shulman GL, Raichle ME. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proc Natl Acad Sci USA. 2001;98:4259–4264. doi: 10.1073/pnas.071043098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallett PE. Primary and secondary saccades to goals defined by instructions. Vision Res. 1978;18:1279–1296. doi: 10.1016/0042-6989(78)90218-3. [DOI] [PubMed] [Google Scholar]
- Herrero MT, Barcia C, Navarro JM. Functional anatomy of thalamus and basal ganglia. Childs Nerv Syst. 2002;18:386–404. doi: 10.1007/s00381-002-0604-1. [DOI] [PubMed] [Google Scholar]
- Holroyd CB, Coles MG. The neural basis of human error processing: reinforcement learning, dopamine, and the error-related negativity. Psychol Rev. 2002;109:679–709. doi: 10.1037/0033-295X.109.4.679. [DOI] [PubMed] [Google Scholar]
- Holroyd CB, Nieuwenhuis S, Yeung N, Nystrom L, Mars RB, Coles MG, Cohen JD. Dorsal anterior cingulate cortex shows fMRI response to internal and external error signals. Nat Neurosci. 2004;7:497–498. doi: 10.1038/nn1238. [DOI] [PubMed] [Google Scholar]
- Huttenlocher PR. Morphometric study of human cerebral cortex development. Neuropsychologia. 1990;28:517–527. doi: 10.1016/0028-3932(90)90031-i. [DOI] [PubMed] [Google Scholar]
- Huttenlocher PR, Dabholkar AS. Regional differences in synaptogenesis in human cerebral cortex. J Comp Neurol. 1997;387:167–178. doi: 10.1002/(sici)1096-9861(19971020)387:2<167::aid-cne1>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- Ito S, Stuphorn V, Brown JW, Schall JD. Performance monitoring by the anterior cingulate cortex during saccade countermanding. Science. 2003;302:120–122. doi: 10.1126/science.1087847. [DOI] [PubMed] [Google Scholar]
- Jernigan TL, Trauner DA, Hesselink JR, Tallal PA. Maturation of human cerebrum observed in vivo during adolescence. Brain. 1991;114(Pt 5):2037–2049. doi: 10.1093/brain/114.5.2037. [DOI] [PubMed] [Google Scholar]
- Kang HC, Burgund ED, Lugar HM, Petersen SE, Schlaggar BL. Comparison of functional activation foci in children and adults using a common stereotactic space. Neuroimage. 2003;19:16–28. doi: 10.1016/s1053-8119(03)00038-7. [DOI] [PubMed] [Google Scholar]
- Kerns JG. Anterior cingulate and prefrontal cortex activity in an FMRI study of trial-to-trial adjustments on the Simon task. Neuroimage. 2006;33:399–405. doi: 10.1016/j.neuroimage.2006.06.012. [DOI] [PubMed] [Google Scholar]
- Kerns JG, Cohen JD, MacDonald AW, 3rd, Cho RY, Stenger VA, Carter CS. Anterior cingulate conflict monitoring and adjustments in control. Science. 2004;303:1023–1026. doi: 10.1126/science.1089910. [DOI] [PubMed] [Google Scholar]
- Klein C, Foerster F. Development of prosaccade and antisaccade task performance in participants aged 6 to 26 years. Psychophysiology. 2001;38:179–189. [PubMed] [Google Scholar]
- Koechlin E, Ody C, Kouneiher F. The architecture of cognitive control in the human prefrontal cortex. Science. 2003;302:1181–1185. doi: 10.1126/science.1088545. [DOI] [PubMed] [Google Scholar]
- Kramer AF, de Sather JC, Cassavaugh ND. Development of attentional and oculomotor control. Dev Psychol. 2005;41:760–772. doi: 10.1037/0012-1649.41.5.760. [DOI] [PubMed] [Google Scholar]
- Kwong KK, Belliveau JW, Chesler DA, Goldberg IE, Weisskoff RM, Poncelet BP, Kennedy DN, Hoppel BE, Cohen MS, Turner R, et al. Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc Natl Acad Sci USA. 1992;89:5675–5679. doi: 10.1073/pnas.89.12.5675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladouceur CD, Dahl RE, Carter CS. ERP correlates of action monitoring in adolescence. Ann N Y Acad Sci. 2004;1021:329–336. doi: 10.1196/annals.1308.040. [DOI] [PubMed] [Google Scholar]
- Lambe EK, Krimer LS, Goldman-Rakic PS. Differential postnatal development of catecholamine and serotonin inputs to identified neurons in prefrontal cortex of rhesus monkey. J Neurosci. 2000;20:8780–8787. doi: 10.1523/JNEUROSCI.20-23-08780.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin HS, Culhane KA, Hartmann J, Evankovich K, Mattson AJ. Developmental changes in performance on tests of purported frontal lobe functioning. Dev Neuropsychol. 1991;7:377–395. [Google Scholar]
- Luna B, Garver KE, Urban TA, Lazar NA, Sweeney JA. Maturation of cognitive processes from late childhood to adulthood. Child Dev. 2004;75:1357–1372. doi: 10.1111/j.1467-8624.2004.00745.x. [DOI] [PubMed] [Google Scholar]
- Luna B, Sweeney JA. Cognitive functional magnetic resonance imaging at very-high-field: eye movement control. Top Magn Reson Imaging. 1999;10:3–15. doi: 10.1097/00002142-199902000-00002. [DOI] [PubMed] [Google Scholar]
- Luna B, Sweeney JA. The emergence of collaborative brain function: FMRI studies of the development of response inhibition. Ann N Y Acad Sci. 2004;1021:296–309. doi: 10.1196/annals.1308.035. [DOI] [PubMed] [Google Scholar]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, Keshavan MS, Genovese CR, Eddy WF, Sweeney JA. Maturation of widely distributed brain function subserves cognitive development. Neuroimage. 2001;13:786–793. doi: 10.1006/nimg.2000.0743. [DOI] [PubMed] [Google Scholar]
- Maccotta L, Zacks JM, Buckner RL. Rapid self-paced event-related functional MRI: feasibility and implications of stimulus- versus response-locked timing. Neuroimage. 2001;14:1105–1121. doi: 10.1006/nimg.2001.0912. [DOI] [PubMed] [Google Scholar]
- MacDonald AW, 3rd, Cohen JD, Stenger VA, Carter CS. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science. 2000;288:1835–1838. doi: 10.1126/science.288.5472.1835. [DOI] [PubMed] [Google Scholar]
- Margulies DS, Kelly AM, Uddin LQ, Biswal BB, Castellanos FX, Milham MP. Mapping the functional connectivity of anterior cingulate cortex. Neuroimage. 2007;37:579–588. doi: 10.1016/j.neuroimage.2007.05.019. [DOI] [PubMed] [Google Scholar]
- Marsh R, Zhu H, Schultz RT, Quackenbush G, Royal J, Skudlarski P, Peterson BS. A developmental fMRI study of self-regulatory control. Hum Brain Mapp. 2006;27:848–863. doi: 10.1002/hbm.20225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuda T, Matsuura M, Ohkubo T, Ohkubo H, Matsushima E, Inoue K, Taira M, Kojima T. Functional MRI mapping of brain activation during visually guided saccades and antisaccades: cortical and subcortical networks. Psychiatry Res. 2004;131:147–155. doi: 10.1016/j.pscychresns.2003.12.007. [DOI] [PubMed] [Google Scholar]
- McAvoy MP, Ollinger JM, Buckner RL. Cluster size thresholds for assessment of significant activation in fMRI. Neuroimage. 2001;11:735–739. [Google Scholar]
- Michelon P, Snyder AZ, Buckner RL, McAvoy M, Zacks JM. Neural correlates of incongruous visual information. An event-related fMRI study. Neuroimage. 2003;19:1612–1626. doi: 10.1016/s1053-8119(03)00111-3. [DOI] [PubMed] [Google Scholar]
- Middleton FA, Strick PL. Basal ganglia output and cognition: evidence from anatomical, behavioral, and clinical studies. Brain Cogn. 2000;42:183–200. doi: 10.1006/brcg.1999.1099. [DOI] [PubMed] [Google Scholar]
- Miezin FM, Maccotta L, Ollinger JM, Petersen SE, Buckner RL. Characterizing the hemodynamic response: effects of presentation rate, sampling procedure, and the possibility of ordering brain activity based on relative timing. Neuroimage. 2000;11:735–759. doi: 10.1006/nimg.2000.0568. [DOI] [PubMed] [Google Scholar]
- Milham MP, Banich MT, Barad V. Competition for priority in processing increases prefrontal cortex's involvement in top-down control: an event-related fMRI study of the Stroop task. Brain Res Cogn Brain Res. 2003;17:212–222. doi: 10.1016/s0926-6410(03)00108-3. [DOI] [PubMed] [Google Scholar]
- Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annu Rev Neurosci. 2001;24:167–202. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- Miller LM, Sun FT, Curtis CE, D'Esposito M. Functional interactions between oculomotor regions during prosaccades and antisaccades. Hum Brain Mapp. 2005;26:119–127. doi: 10.1002/hbm.20146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monchi O, Petrides M, Strafella AP, Worsley KJ, Doyon J. Functional role of the basal ganglia in the planning and execution of actions. Ann Neurol. 2006;59:257–264. doi: 10.1002/ana.20742. [DOI] [PubMed] [Google Scholar]
- Munoz DP, Broughton JR, Goldring JE, Armstrong IT. Age-related performance of human subjects on saccadic eye movement tasks. Exp Brain Res. 1998;121:391–400. doi: 10.1007/s002210050473. [DOI] [PubMed] [Google Scholar]
- Muri RM, Iba-Zizen MT, Derosier C, Cabanis EA, Pierrot-Deseilligny C. Location of the human posterior eye field with functional magnetic resonance imaging. J Neurol Neurosurg Psychiatry. 1996;60:445–448. doi: 10.1136/jnnp.60.4.445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieuwenhuis S, Ridderinkhof KR, Blom J, Band GP, Kok A. Error-related brain potentials are differentially related to awareness of response errors: evidence from an antisaccade task. Psychophysiology. 2001;38:752–760. [PubMed] [Google Scholar]
- O'Driscoll GA, Alpert NM, Matthysse SW, Levy DL, Rauch SL, Holzman PS. Functional neuroanatomy of antisaccade eye movements investigated with positron emission tomography. Proc Natl Acad Sci USA. 1995;92:925–929. doi: 10.1073/pnas.92.3.925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogawa S, Tank DW, Menon R, Ellermann JM, Kim SG, Merkle H, Ugurbil K. Intrinsic signal changes accompanying sensory stimulation: functional brain mapping with magnetic resonance imaging. Proc Natl Acad Sci USA. 1992;89:5951–5955. doi: 10.1073/pnas.89.13.5951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ojemann JG, Akbudak E, Snyder AZ, McKinstry RC, Raichle ME, Conturo TE. Anatomic localization and quantitative analysis of gradient refocused echo-planar fMRI susceptibility artifacts. Neuroimage. 1997;6:156–167. doi: 10.1006/nimg.1997.0289. [DOI] [PubMed] [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Olesen PJ, Nagy Z, Westerberg H, Klingberg T. Combined analysis of DTI and fMRI data reveals a joint maturation of white and grey matter in a fronto-parietal network. Brain Res Cogn Brain Res. 2003;18:48–57. doi: 10.1016/j.cogbrainres.2003.09.003. [DOI] [PubMed] [Google Scholar]
- Ollinger JM, Corbetta M, Shulman GL. Separating processes within a trial in event-related functional MRI. Neuroimage. 2001;13:218–229. doi: 10.1006/nimg.2000.0711. [DOI] [PubMed] [Google Scholar]
- Ollinger JM, Shulman GL, Corbetta M. Separating processes within a trial in event-related functional MRI. Neuroimage. 2001;13:210–217. doi: 10.1006/nimg.2000.0710. [DOI] [PubMed] [Google Scholar]
- Petit L, Orssaud C, Tzourio N, Salamon G, Mazoyer B, Berthoz A. PET study of voluntary saccadic eye movements in humans: basal ganglia-thalamocortical system and cingulate cortex involvement. J Neurophysiol. 1993;69:1009–1017. doi: 10.1152/jn.1993.69.4.1009. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Mathalon DH, Sullivan EV, Rawles JM, Zipursky RB, Lim KO. A quantitative magnetic resonance imaging study of changes in brain morphology from infancy to late adulthood. Arch Neurol. 1994;51:874–887. doi: 10.1001/archneur.1994.00540210046012. [DOI] [PubMed] [Google Scholar]
- Pierrot-Deseilligny C, Milea D, Muri RM. Eye movement control by the cerebral cortex. Curr Opin Neurol. 2004;17:17–25. doi: 10.1097/00019052-200402000-00005. [DOI] [PubMed] [Google Scholar]
- Pierrot-Deseilligny C, Muri RM, Nyffeler T, Milea D. The role of the human dorsolateral prefrontal cortex in ocular motor behavior. Ann N Y Acad Sci. 2005;1039:239–251. doi: 10.1196/annals.1325.023. [DOI] [PubMed] [Google Scholar]
- Pierrot-Deseilligny C, Rivaud S, Gaymard B, Muri R, Vermersch AI. Cortical control of saccades. Ann Neurol. 1995;37:557–567. doi: 10.1002/ana.410370504. [DOI] [PubMed] [Google Scholar]
- Ploran EJ, Nelson SM, Velanova K, Donaldson DI, Petersen SE, Wheeler ME. Evidence accumulation and the moment of recognition: dissociating perceptual recognition processes using fMRI. J Neurosci. 2007;27:11912–11924. doi: 10.1523/JNEUROSCI.3522-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polli FE, Barton JJ, Cain MS, Thakkar KN, Rauch SL, Manoach DS. Rostral and dorsal anterior cingulate cortex make dissociable contributions during antisaccade error commission. Proc Natl Acad Sci USA. 2005;102:15700–15705. doi: 10.1073/pnas.0503657102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci USA. 2001;98:676–682. doi: 10.1073/pnas.98.2.676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravizza SM, Ciranni MA. Contributions of the prefrontal cortex and basal ganglia to set shifting. J Cogn Neurosci. 2002;14:472–483. doi: 10.1162/089892902317361985. [DOI] [PubMed] [Google Scholar]
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science. 2004;306:443–447. doi: 10.1126/science.1100301. [DOI] [PubMed] [Google Scholar]
- Robinson DL, Goldberg ME. Sensory and behavioral properties of neurons in posterior parietal cortex of the awake, trained monkey. Fed Proc. 1978;37:2258–2261. [PubMed] [Google Scholar]
- Rosenberg DR, Sweeney JA, Gillen JS, Kim J, Varanelli MJ, O'Hearn KM, Erb PA, Davis D, Thulborn KR. Magnetic resonance imaging of children without sedation: preparation with simulation. J Am Acad Child Adolesc Psychiatry. 1997;36:853–859. doi: 10.1097/00004583-199706000-00024. [DOI] [PubMed] [Google Scholar]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, Andrew C, Bullmore ET. Functional frontalisation with age: mapping neurodevelopmental trajectories with fMRI. Neurosci Biobehav Rev. 2000;24:13–19. doi: 10.1016/s0149-7634(99)00055-x. [DOI] [PubMed] [Google Scholar]
- Rubia K, Smith AB, Taylor E, Brammer M. Linear age-correlated functional development of right inferior fronto-striato-cerebellar networks during response inhibition and anterior cingulate during error-related processes. Hum Brain Mapp. 2007;28:1163–1177. doi: 10.1002/hbm.20347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubia K, Smith AB, Woolley J, Nosarti C, Heyman I, Taylor E, Brammer M. Progressive increase of frontostriatal brain activation from childhood to adulthood during event-related tasks of cognitive control. Hum Brain Mapp. 2006;27:973–993. doi: 10.1002/hbm.20237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Functional neuroanatomical differences between adults and school-age children in the processing of single words. Science. 2002;296:1476–1479. doi: 10.1126/science.1069464. [DOI] [PubMed] [Google Scholar]
- Shima K, Tanji J. Role for cingulate motor area cells in voluntary movement selection based on reward. Science. 1998;282:1335–1338. doi: 10.1126/science.282.5392.1335. [DOI] [PubMed] [Google Scholar]
- Snyder AZ. Difference image versus ratio image error function forms in PET–PET realignment. In: Bayley D, Jones T, editors. Quantification of brain function using PET. San Diego (CA): Academic Press; 1996. pp. 131–137. [Google Scholar]
- Suchan B, Jokisch D, Skotara N, Daum I. Evaluation-related frontocentral negativity evoked by correct responses and errors. Behav Brain Res. 2007;183:206–212. doi: 10.1016/j.bbr.2007.06.013. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. Co-planar stereotaxic atlas of the brain. New York: Thieme; 1988. [Google Scholar]
- Tamm L, Menon V, Reiss AL. Maturation of brain function associated with response inhibition. J Am Acad Child Adolesc Psychiatry. 2002;41:1231–1238. doi: 10.1097/00004583-200210000-00013. [DOI] [PubMed] [Google Scholar]
- Taylor SF, Stern ER, Gehring WJ. Neural systems for error monitoring: recent findings and theoretical perspectives. Neuroscientist. 2007;13:160–172. doi: 10.1177/1073858406298184. [DOI] [PubMed] [Google Scholar]
- The Psychological Corporation. Wechsler Abbreviated Scale of Intelligence (WASI) manual. San Antonio (TX): The Psychological Corporation; 1999. [Google Scholar]
- Toga AW, Thompson PM, Sowell ER. Mapping brain maturation. Trends Neurosci. 2006;29:148–159. doi: 10.1016/j.tins.2006.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Essen DC, Drury HA, Dickson J, Harwell J, Hanlon D, Anderson CH. An integrated software suite for surface-based analyses of cerebral cortex. JAMA. 2001;8:443–459. doi: 10.1136/jamia.2001.0080443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Veen V, Carter CS. The timing of action-monitoring processes in the anterior cingulate cortex. J Cogn Neurosci. 2002;14:593–602. doi: 10.1162/08989290260045837. [DOI] [PubMed] [Google Scholar]
- van Veen V, Cohen JD, Botvinick MM, Stenger VA, Carter CS. Anterior cingulate cortex, conflict monitoring, and levels of processing. Neuroimage. 2001;14:1302–1308. doi: 10.1006/nimg.2001.0923. [DOI] [PubMed] [Google Scholar]
- Velanova K, Jacoby LL, Wheeler ME, McAvoy MP, Petersen SE, Buckner RL. Functional-anatomic correlates of sustained and transient processing components engaged during controlled retrieval. J Neurosci. 2003;23:8460–8470. doi: 10.1523/JNEUROSCI.23-24-08460.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velanova K, Lustig C, Jacoby LL, Buckner RL. Evidence for frontally mediated controlled processing differences in older adults. Cereb Cortex. 2007;17:1033–1046. doi: 10.1093/cercor/bhl013. [DOI] [PubMed] [Google Scholar]
- Visscher KM, Miezin FM, Kelly JE, Buckner RL, Donaldson DI, McAvoy MP, Bhalodia VM, Petersen SE. Mixed blocked/event-related designs separate transient and sustained activity in fMRI. Neuroimage. 2003;19:1694–1708. doi: 10.1016/s1053-8119(03)00178-2. [DOI] [PubMed] [Google Scholar]
- Wenger KK, Visscher KM, Miezin FM, Petersen SE, Schlaggar BL. Comparison of sustained and transient activity in children and adults using a mixed blocked/event-related fMRI design. Neuroimage. 2004;22:975–985. doi: 10.1016/j.neuroimage.2004.02.028. [DOI] [PubMed] [Google Scholar]
- Wheeler ME, Buckner RL. Functional dissociation among components of remembering: control, perceived oldness, and content. J Neurosci. 2003;23:3869–3880. doi: 10.1523/JNEUROSCI.23-09-03869.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams BR, Ponesse JS, Schachar RJ, Logan GD, Tannock R. Development of inhibitory control across the life span. Dev Psychol. 1999;35:205–213. doi: 10.1037//0012-1649.35.1.205. [DOI] [PubMed] [Google Scholar]
- Worsley KJ, Friston KJ. Analysis of fMRI time-series revisited—again. Neuroimage. 1995;2:173–181. doi: 10.1006/nimg.1995.1023. [DOI] [PubMed] [Google Scholar]
- Zarahn E, Aguirre GK, D'Esposito M. Empirical analyses of BOLD fMRI statistics. I. Spatially unsmoothed data collected under null-hypothesis conditions. Neuroimage. 1997;5:179–197. doi: 10.1006/nimg.1997.0263. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.