

Abstract
Cross-sectional magnetic resonance imaging (MRI) studies of cortical thickness and volume have shown age effects on large areas, but there are substantial discrepancies across studies regarding the localization and magnitude of effects. These discrepancies hinder understanding of effects of aging on brain morphometry, and limit the potential usefulness of MR in research on healthy and pathological age-related brain changes. The present study was undertaken to overcome this problem by assessing the consistency of age effects on cortical thickness across 6 different samples with a total of 883 participants. A surface-based segmentation procedure (FreeSurfer) was used to calculate cortical thickness continuously across the brain surface. The results showed consistent age effects across samples in the superior, middle, and inferior frontal gyri, superior and middle temporal gyri, precuneus, inferior and superior parietal cortices, fusiform and lingual gyri, and the temporo-parietal junction. The strongest effects were seen in the superior and inferior frontal gyri, as well as superior parts of the temporal lobe. The inferior temporal lobe and anterior cingulate cortices were relatively less affected by age. The results are discussed in relation to leading theories of cognitive aging.
Keywords: aging, cortex, frontal lobes, morphometry, MRI
Introduction
In magnetic resonance imaging (MRI) studies, there is consensus that higher age is associated with reduction of brain volumes, including the cerebral cortex, as well as expansion of the ventricular system (Jernigan et al. 1991; Pfefferbaum et al. 1994; Blatter et al. 1995; Sullivan et al. 1995, 2004; Murphy et al. 1996; Raz et al. 1997; Courchesne et al. 2000; Resnick et al. 2000; Good et al. 2001; Jernigan et al. 2001; Raz, Gunning-Dixon, et al. 2004; Salat et al. 2004; Taki et al. 2004; Allen et al. 2005; Fotenos et al. 2005; Walhovd et al. 2005; Kruggel 2006). However, cumulative evidence also indicates that brain aging is not uniform, and significant heterogeneity of age effects is observed across brain regions (Raz et al. 1997, 2005; Good et al. 2001; Jernigan et al. 2001; Raz, Gunning-Dixon, et al. 2004; Salat et al. 2004). Unfortunately, interpretation of this heterogeneity is complicated by inconsistency among results (Raz and Rodrigue 2006). The aim of the present study was to address this problem by testing the consistency of age effects on cortical thickness across 6 samples from 4 different research groups.
Studies using manual drawing of regions of interest (ROIs) on MRI scans have especially shown age effects on frontal cortices, with significant but more moderate age effects in the temporal, parietal, and occipital association areas. In contrast, the primary sensory (especially visual) cortices seem largely preserved (Raz, Gunning-Dixon, et al. 2004; Allen et al. 2005; Raz and Rodrigue 2006). New automated and semiautomated segmentation techniques have enabled studies of age effects continuously across the cortical mantle without manually defining ROIs. This facilitates comparison of results across studies. In such studies, there is consensus that age effects are strong in frontal or prefrontal areas (Good et al. 2001; Sato et al. 2003; Salat et al. 2004; Taki et al. 2004; Brickman et al. 2007; Raz, Rodrigue and Haacke 2007; Abe et al. 2008; Kalpouzos et al. 2008), in line with findings from manual morphometry studies (Raz and Rodrigue 2006). However, in contrast to most manual studies, several investigations using automated/semiautomated methods find that the occipital lobes are negatively affected by age (Sato et al. 2003; Salat et al. 2004; Taki et al. 2004; Abe et al. 2008; Kalpouzos et al. 2008). Moreover, most have demonstrated age effects on parietal cortex (Good et al. 2001; Salat et al. 2004; Brickman et al. 2007; Abe et al. 2008), and around the central sulcus (Good et al. 2001; Salat et al. 2004), although the exact localization of the effects vary.
Age effects on the anterior cingulate cortex (ACC) have been found to be inconsistent between studies using automated/semiautomated techniques. In some studies (Salat et al. 2004; Abe et al. 2008), ACC thickening has been found, whereas ACC preservation or reduction with age has been found in others (Good et al. 2001; Tisserand et al. 2002; Brickman et al. 2007; Vaidya et al. 2007; Kalpouzos et al. 2008). In manual investigations, only mild or no age differences have been found in the ACC (for a review, see Raz et al. 2000). Automated/semiautomated studies have also reported discrepant results within the prefrontal cortex. Some have found sparing of the medial orbitofrontal cortex (Salat et al. 2004), whereas others have reported age effects throughout most of the anterior part of the brain (Taki et al. 2004; Abe et al. 2008). It is unclear whether such discrepancies are method or sample dependent, although studies comparing different methods have suggested that the former is likely (Tisserand et al. 2002; Kennedy et al. 2008).
The temporal lobes and especially medial-temporal regions are involved in memory processing, and have received much attention in research on normal and pathological aging (Braak and Braak 1985; Mesulam 1999). Temporal cortical thinning has been observed in mild cognitive impairment and Alzheimer's disease (AD) (Dickerson, Feczko, et al. 2008; Fjell et al. 2008). Relative sparing of temporal (Salat et al. 2004) and parahippocampal (Raz et al. 1997; Raz, Gunning-Dixon, et al. 2004; Salat et al. 2004) cortices has been found in healthy aging, and the entorhinal cortex has been found to be relatively less affected by age than other cortical regions (Good et al. 2001; Raz et al. 2005). However, other studies have found temporal cortical thinning in healthy aging also (Sato et al. 2003; Taki et al. 2004; Brickman et al. 2007; Abe et al. 2008). Volume reductions accompanied by volume increases in different parts of the temporal cortex have also been observed (Kalpouzos et al. 2008). Longitudinal age-related shrinkage of the entorhinal cortex was identified in 1 study (Du et al. 2006), whereas others have found that such atrophy was limited to persons with relatively poor, though normal cognitive performance (Raz, Rodrigue, et al. 2004; Rodrigue and Raz 2004; Raz et al. 2008). Notably, no correlations of entorhinal volume with age were found at baseline in these studies. Targeting morphometric effects in the temporal lobe with a newly developed semiautomated technique, another study found that entorhinal and perirhinal, but not posterior parahippocampal cortices, were reduced in volume with age (Dickerson, Feczko, et al. 2008). This could be attributed mainly to reduction of the surface area rather than thickness of the cerebral cortex.
Although, as reviewed above, well-established patterns of age effects exist, significant discrepancies are observed across samples (for a review, see Raz and Rodrigue 2006). Such between-study variability complicates interpretation of the findings. Multiple reasons have been invoked to explain the discrepant results, and at least 3 seem to be of great importance. First, the differences in methodology of image processing may contribute to variance in the results (Tisserand et al. 2002; Kennedy et al. 2008). Second, characteristics of participants may contribute to interstudy heterogeneity through uncontrolled differences in health and genetic endowment. Finally, although the sample sizes employed appear quite large, they still may be insufficient to detect subtle differences in age effects on specific cortical regions.
The present study was designed to address these problems. We applied a uniform semiautomated method of cortical thickness measurement yielding thickness estimates continuously across the entire cortical mantle to a large sample of healthy adults recruited from multiple populations. The data include 6 samples (3 from the United States, 2 from Norway, and 1 from Sweden) drawn from 4 research centers, with a total of 883 participants. The data were analyzed both sample-wise and pooled. The principal aim was to examine the consistency of age effects on regional cortical thickness across samples. This was done to test which cortical areas consistently underwent age-related thinning or thickening, and which areas were differentially affected by age in different samples.
For measurement of cortical thickness, FreeSurfer software, freely downloadable from http://surfer.nmr.mgh.harvard.edu/, was used (Center for Biomedical Imaging, Charlestown, MA). This approach provides reliable measures of cortical thickness continuously across the whole cortical mantle without manually defining ROIs. Although automated methods may have some undesirable features, such as resolution loss in registration of morphologically different brains to a common stereotactic space, and the need for smoothing, they have several advantages over manual methods. First, they require minimal intervention by highly trained personnel and allow processing of many brains in a reasonable time frame. Second, they are characterized by very high reliability and repeatability of measures. Third, they allow a hypothesis-free search for patterns of differences without the need to define anatomically plausible ROIs, enabling detection of differences in regions where precise anatomical definitions and placement of anatomical borders would not be feasible.
Materials and Methods
Samples
The details of each of the 6 samples are described in Table 1, where key publications and sample selection criteria are indicated, and in Supplementary Table 1. The total number of participants was 883, with an age range of 75 years (18–93 years). All samples were screened for history of neurological conditions. Even though subclinical effects of pathological processes cannot be ruled out without follow-up data, it is likely that effects can largely be attributed to the influence of nondemented aging. Twenty-two participants were excluded due to bad scan quality, including overfolding, MR artifacts, errors during data transfer or saving, converting errors, or deviant signal intensity in the MRI images. Most of these scans were possible to process, but were deemed not to be of sufficient quality to yield valid results. Included in these 22 were a small number of participants that were excluded due to white matter (WM) abnormalities. We find it unlikely that excluding this small number of problematic scans bias the data.
Table 1.
Sample characteristics
| Sample | Country | N (% f) | Age mean (range) | Education mean (range) | Key publications | Main screening instruments/inclusion criteria |
| 1 | Nor | 69 (57) | 51.3 (20–88) | 15 (7–20) | (Walhovd et al. 2005) | Health interview, MMSE > 26, BDI < 16, IQ > 85, RH only |
| 2 | Nor | 208 (71) | 46.8 (19–75) | 14 (9–22) | (Espeseth et al. 2008) | Health interview, IQ > 85 |
| 3 | Swe | 106 (32) | 41.6 (19–56) | 14 (9–22) | (Jonsson et al. 2006; Nesvag et al. 2008) | Health interview, DSM-III-R, WASI vocabulary > 16a |
| 4 | USA | 155 (65) | 44.5 (18–93) | 3.5 (1–5)b | (Marcus et al. 2007) | Health interview, CDR = 0c, MMSE > 25c, RH only |
| 5 | USA | 154 (61) | 44.4 (18–94) | 3.4 (1–5)b | Similar to sample 4 | Similar to sample 4 |
| 6 | USA | 191 (60) | 47.3 (18–81) | 15.7 (12–21) | (Raz, Gunning-Dixon, et al. 2004) | Health interview, BIMCT > 30, GDQ < 15, RH only, neuroradiology |
Note: Nor, Norway; Swe, Sweden; % f, percentage of female participants; MMSE, Mini Mental Status Exam (Folstein et al. 1975); BDI, Beck Depression Inventory (Beck 1987); BIMCT, Blessed Information–Memory–Concentration Test (Blessed et al. 1968); CDR, clinical dementia rating (Berg 1984, 1988; Morris 1993); GDQ, Geriatric Depression Questionnaire (Auer and Reisberg 1997); RH, right handed; WASI, Wechsler Abbreviated Scale of Intelligence (Wechsler 1999).
Available for 70 participants.
Available for all participants ≥60 years, and sporadically for the rest. 1: less than high school grad., 2: high school grad., 3: some college, 4: college grad., 5: beyond college.
Available for participants ≥60 years only.
MR Acquisition
All participants were scanned on 1.5T magnets, but from 2 different manufacturers (Siemens, Erlangen, Germany; General Electric CO [GE], Milwaukee, WI), and 4 different models (Siemens Symphony Quantum, Siemens Sonata, Siemens Vision, GE Signa). With the exceptions of the data from sample 4 and 5, all the samples were from different scanners. All participants within each sample were scanned on the same scanner. T1-weighted sequences were acquired (3D magnetization prepared gradient-echo (MP–RAGE) for the Siemens scanners, and 3D spoiled gradient recalled (SPGR) pulse sequences for GE. Slice thickness were between 1.25 mm (samples 4 and 5) and 1.5 mm (sample 1), with acquisition matrices of 256 × 192 (samples 1, 3, and 6) or 256 × 256 (samples 2, 4, and 5). For 4 of the samples (samples 1, 2, 4, and 5), multiple scans were acquired within the same scanning session, and averaged to increase the signal-to-noise ratio (SNR). The detailed sequences used are presented in Table 2. Examples of scan quality from each sample are presented in Figure 1.
Table 2.
MRI parameters
| Sample | MRI scanner | MRI protocol |
| Sample 1 | 1.5T Siemens Symphony Quantum | Two 3D MP–RAGE T1-weighted sequences |
| TR/TE/TI/FA = 2730 ms/4 ms/1000 ms/7° | ||
| Matrix = 192 × 256 | ||
| Scan time: 8.5 min per volume. | ||
| Each volume consisted of 128 sagittal slices (1.33 × 1 × 1 mm). | ||
| Sample 2 | 1.5T Siemens Sonata | Two 3D MP–RAGE T1-weighted sequences |
| TR/TE/TI/FA = 2730 ms/3.43 ms/1000 ms/7° | ||
| Matrix: 256 × 256 | ||
| Scan time: 8 min and 46 s per volume | ||
| Each volume consisted of 128 sagittal slices (1.33 × 1 × 1 mm) | ||
| Sample 3 | 1.5T General Electric Signa | One 3D SPGR pulse T1-weighted sequence |
| TR/TE/FA = 24 ms/6.0 ms/35°, number of excitations were 2 | ||
| Matrix: 256 × 192 | ||
| Each volume consisted of 1.5-mm coronal slices, no gap, FOV = 24 cm | ||
| Sample 4 | 1.5T Siemens Vision | Three to 4 individual T1-weighted MP–RAGE T1-weighted sequences |
| TR/TE/TI/FA = 9.7 ms/4.0 ms/20 ms/10° | ||
| Matrix = 256 × 256. | ||
| Each volume consisted of 128 sagittal slices (1.25 × 1 × 1 mm). | ||
| Sample 5 | See sample 4 | See sample 4 |
| Sample 6 | 1.5T General Electric Signa | One 3D SPGR pulse T1-weighted sequence |
| TR/TE/FA = 24 ms/5.0 ms/30° | ||
| Matrix = 256 × 192 | ||
| Each volume consisted of 124 contiguous axial slices (1.30 × 0.94 × 0.86 mm), FOV = 22 cm |
Note: FOV, field of view; FA, flip angle; TR, repetition time; TE, echo time; TI, inversion time.
Figure 1.
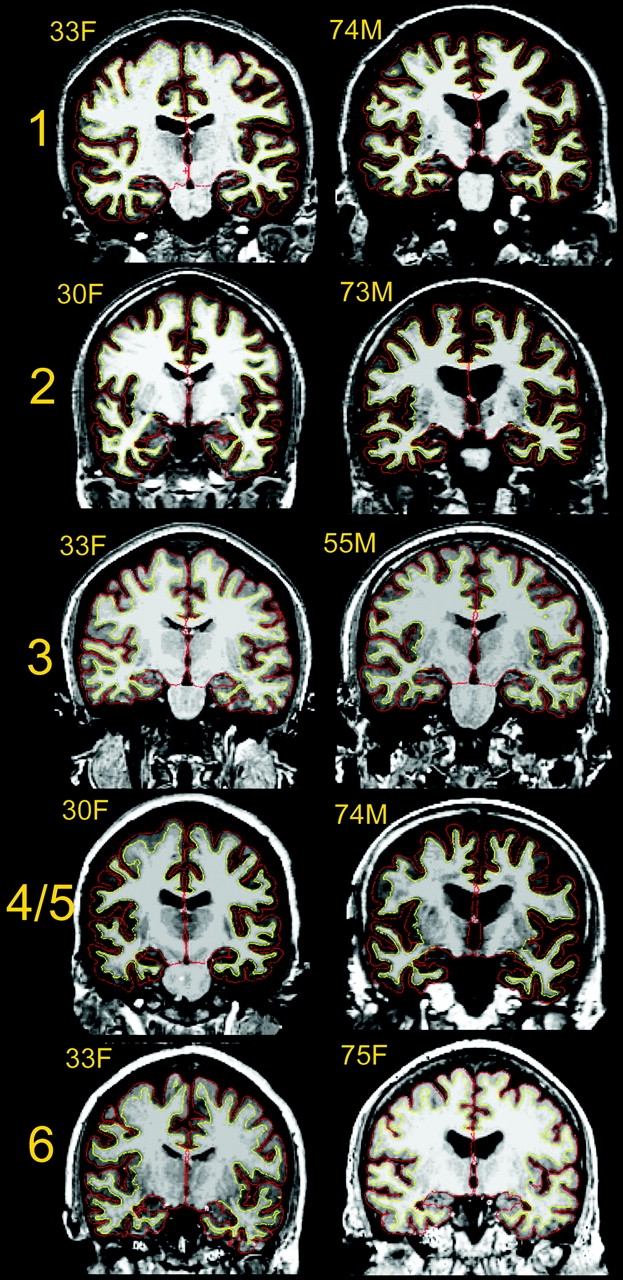
Example scans from each sample. Scans representative of image quality of 1 young and 1 elderly participant from each of the samples are shown (because sample 4 and 5 are from the same scanner, only examples from sample 4 are shown. All scans are converted from their native format to Freesurfer format. Samples 1, 2, and 4 are taken from Siemens scanners, and 2–4 acquisitions were averaged from each participant to yield high contrast and signal to noise ratio. Sample 2 and 6 are from GE scanners (Signa), with 1 acquisition. The cortex–CSF boundary (red) and the gray–white boundary (yellow) are indicated by the thin line. Anatomical differences between the scans from each sample are incidental.
Cortical Thickness Analyses
Cortical thickness measurements were obtained by reconstructing representations of the gray/white matter boundary (Dale and Sereno 1993; Dale et al. 1999) and the cortical surface, and then calculating the distance between these surfaces at each point across the cortical mantle. This method uses both intensity and continuity information from the entire 3D MR volume in segmentation and deformation procedures to construct representations of cortical thickness. The maps are created using spatial intensity gradients across tissue classes and are therefore not simply reliant on absolute signal intensity. The maps produced are not restricted to the voxel resolution of the original data and thus are capable of detecting submillimeter differences between groups (Fischl and Dale 2000). This has been validated using histology and MR (Rosas et al. 2002; Kuperberg et al. 2003). Rosas and colleagues (Rosas et al. 2002) have shown that processing of MR images of autopsy brains gave cortical thickness estimates within ±0.25 mm of those obtained using neuropathologic methods and were statistically indistinguishable. Thickness measures may be mapped on the “inflated” or “semi-inflated” surface of each participant's reconstructed brain (Dale et al. 1999; Fischl et al. 1999), allowing visualization of data across the entire cortical surface without interference from cortical folding. Maps were smoothed using a circularly symmetric Gaussian kernel across the surface with a full width at half maximum (FWHM) of 15 mm and averaged across participants using a nonrigid high-dimensional spherical averaging method to align cortical folding patterns (Fischl et al. 1999). This procedure provides accurate matching of morphologically homologous cortical locations among participants on the basis of each individual's anatomy while minimizing metric distortion, resulting in a measure of cortical thickness for each person at each point on the reconstructed surface. Statistical comparisons of global data and surface maps were generated by computing a GLM of the effects of each variable on thickness at each vertex. In addition, by use of a validated automated labeling system (Fischl et al. 2004; Desikan et al. 2006), the cortex was divided into 33 different gyral-based areas in each hemisphere, and mean thickness in each was calculated (see Supplementary Fig. 1).
The thickness estimation procedure is automated, but requires manual checking of the accuracy of the spatial registration and the WM and gray matter (GM) segmentations. The types of errors that most often require user intervention are insufficient removal of nonbrain tissue (typically dura in superior brain areas) and inclusion of vessels adjacent to the cortex (especially, in the temporal lobes). In addition, if large field inhomogeneity exists, small parts of WM may mistakenly be misclassified as GM, thus obscuring the GM/WM boundary. These types of errors are limited in spatial extension, typically seen in a minor area of the brain in a few slices, but are nevertheless routinely corrected by manual interventions. However, some researchers argue that manual interventions are not always necessary, and have shown that cortical thickness can be estimated reliably across field strength, scanner upgrade and manufacturer without any manual intervention (Han et al. 2006), showing reliable correlations with cognitive function (Dickerson, Feczko, et al. 2008; Dickerson, Fenstermacher, et al. 2008).
Statistical Analyses
First, general linear models (GLMs) were used to test the relationship between age and cortical thickness at each vertex across the entire cortical mantle in each sample separately, with effects of sex regressed out. To handle the problem of multiple comparisons in neuroimaging data, false discovery rate (FDR) < 0.05, was applied to threshold the initial data. However, because widespread and robust effects were expected, results were also presented with higher P value thresholds (P < 10−3 to 10−9) to better visualize where the effects of age on cortical thickness were strongest. In addition, the degree of overlap between results from the different samples was calculated based on the number of samples in which each of the P value thresholds was reached for each surface vertex. This information was color coded and projected onto a template brain. To present the reader with numerical information, correlations with mean thickness in each of the 33 cortical regions (mean of left and right hemisphere) as well as the results of 1-way ANOVAs (F-statistics) are included as supplementary information.
Next, all samples were included simultaneously in 1 GLM. The effect of age, with the main effect of sample regressed out, and the age × sample interaction, were modeled. The results are presented both at FDR < 0.05, and P < 10−15 to 10−25 (uncorrected). Post hoc regression analyses from selected regions of interest (ROIs) were performed to investigate the basis of between-samples variation in age effects. Finally, to test whether nonlinear effects of age could be identified, the GLMs were repeated with both age and age2 included as regressors.
Results
Widespread age differences in cortical thickness were observed across samples. However, the magnitude of effects varied among samples and brain regions. For illustration purposes, mean thickness for each point on the cortical surface for 3 age groups (young, middle-aged, and older adults) is presented in Figure 2.
Figure 2.

Mean cortical thickness in 3 age groups. Mean thickness in each hemisphere for the age groups <40 years, 40–60 years, and >60 years are color coded and projected onto an inflated template brain for better visualization of effects buried in sulci. Note that the participants from all the samples are pooled together in each of the age groups, with no corrections for scanner or sample.
Results of the GLM analyses of age differences in cortical thickness are shown in Figure 3. When FDR < 0.05 was used as threshold, it was clear that age was associated with thinning of the cortex across almost the entire brain surface. Still, for 3 of the samples (samples 4–6), bilateral thickening in the medial frontal cortex, including anterior cingulate gyrus, was seen. The age effects were generally smaller in sample 3 than in the other samples, but even in that sample, thinning appeared most prominent in the prefrontal regions, and the overlap among the samples was generally substantial. Using a P value scale from 10−3 to 10−9 (uncorrected) allowed inspection of regionally differential effects. The frontal cortices, that is, superior and inferior frontal gyri, were among the areas most strongly affected by age across all samples. Age did generally have more moderate effects on the medial-temporal cortices (parahippocampal and entorhinal), although some thinning was seen in 4 of the samples. Lateral inferior parts of the temporal lobes were among the best preserved in 4 of the samples. Superior parts of the lateral temporal lobes were more affected by age than the inferior parts. When the higher P value threshold was used, the thickening in the medial frontal cortex was only seen in samples 4 and 5, and only for the left hemisphere. In addition to these analyses, region-based analyses are presented in Supplementary Table 1. Here, correlations between age in each subsample and for the total sample are shown, along with corresponding F values for age group based ANOVAs.
Figure 3.

Age effects on cortical thickness in each sample. The figure shows the effect of age on cortical thickness across the entire brain surface when effects of sex were regressed out. The results are color coded and projected onto a semi-inflated template brain for better visualization of effects buried in sulci. Each row represents the results from 1 sample. On the left side of the figure, the effects are thresholded at FDR < 0.05 (corrected for multiple comparisons). On the right side, the results are color coded by use of a wider P value scale.
The degree of overlap between the studies is shown in Figure 4. When effects were seen in 5 or all 6 samples for a given area, it was regarded as an area with consistent age effects. When effects were seen in none or one of the samples, it was regarded as an area of preservation. When FDR < 0.05 was used, large areas showed consistent age effects across studies, especially frontal cortices, where effects were seen in superior, middle, and inferior frontal cortices in all 6 samples. Also superior and middle temporal gyri were affected in all samples, as were also fusiform and lingual gyri, and the temporoparietal junction. The occipital lobe and entorhinal cortex were affected by age in 4 of the samples, whereas the inferior temporal lobe and anterior cingulate were generally spared. Effects in the right precuneus were found in 4 of the samples, and in all or 5 of the samples in the left. When the threshold was increased to P < 10−6 (uncorrected), the frontal lobes (superior and inferior frontal gyrus) stood out as related to age in 5 of the samples. Superior parts, especially posteriorly, of the temporal lobes were also relatively prone to effects of age, although there was some variability between samples and between different parts of the gyri.
Figure 4.

Consistency across samples. The number of samples in which a statistical effect was reached is color coded and projected onto a semi-inflated template brain. The first row depicts the results when a threshold of FDR < 0.05 was used. As can be seen, age-related thinning of the cerebral cortex is seen in all or 5 of the samples across most of the brain surface. In the second and third row, higher P value thresholds were used.
Next, the effect of age on cortical thickness was tested with all samples pooled together, and the effect of sample regressed out. A movie of the continuous reduction of thickness is presented in Supplemental Material 1, and the statistical results are presented in Figure 5. With FDR < 0.05 as threshold, negative effects of age on cortical thickness were seen across the entire cortical mantle. Thus, the scale was increased to P < 10−15 to 10−25. The pattern of effects mimicked the results from the analyses of the separate samples. The strongest age effects were found in the superior, middle, and inferior frontal gyri, and in the superior temporal gyri as well as the superior parts of the middle temporal gyri. Further, the temporoparietal junction was also very prone to the effects of age, as were the pericalcarine cortex, fusiform and lingual gyri. Medial parts of the temporal lobes, as well as precuneus, were relatively more preserved, as were the post- and precentral gyri, superior parietal gyri, and the occipital lobe. The effects appeared symmetrically distributed across hemispheres.
Figure 5.

Age effects in the total sample. The figure shows the effects of age on cortical thickness when all samples were included in the same analysis (n = 883), with main effect of sample regressed out. In the second row, a higher P value threshold was employed. Even with a P value threshold of 10−25, large effects were seen in several areas.
The interaction effect of sample × age on cortical thickness was modeled and the results are presented in Figures 6 and 7. With a threshold of FDR < 0.05, several areas showed sample × age interactions. These were mostly overlapping with areas of large age effects across studies. To investigate the basis of the interaction effects further, 5 ROIs were drawn on the inflated template brain surface. These were guided by areas of large sample age × interaction effects, and mean thickness in each ROI was calculated. As can be seen from the scatter plots (Fig. 7), the interaction effects are mainly due to different rates of estimated age-related declines across samples. None of the ROIs showed positive relationships with age. In 2 ROIs, the inferior and superior frontal gyri, thickness correlated significantly with age in all samples, whereas in a third, the middle temporal gyrus, this was found for 5 of the samples. For these areas, there was variation in the strength of the relationships. For instance, correlations ranged from −0.51 to −0.75 for the superior frontal gyrus. Thus, sample × age interactions were to a large extent caused by differences in quantity, not quality, of the age-relationships. Two exceptions to this pattern were found. First, the left medial frontal cortex showed thickening with age in 3 of the samples, whereas relative preservation was found in the other samples. Second, interaction effects could be seen in inferior lateral temporal areas as well as the entorhinal cortex, which were due to thinning in some of the samples (i.e., samples 1, 2, and 5), with relative sparing or only small effects in the other samples. The same was true for the occipital lobe.
Figure 6.

Sample × age interaction effects. The figure shows which areas of the cerebral cortex that were affected differently by age across samples. The color-coded areas represent significant age × sample interaction effects. Note that the areas which display age × sample interactions are the ones where the strongest age effects were found.
Figure 7.

Scatter plots of mean thickness in selected cortical areas. Manual ROIs were drawn on the inflated template brain surface. The areas in which age and sample interacted were used to guide the manual drawing of ROIs. Mean thickness in different cortical areas were calculated, and plotted against age. The ROIs are shown in the upper row. The scatter plots are shown in the middle row. The participants from each sample are coded in different colors. The last row depicts the Pearson correlation coefficients between age and mean thickness in each of the ROIs. The coefficients are given above each bar if P ≤ 0.05 (uncorrected), and not given if not significant (P > 0.05).
Finally, we tested for presence of nonlinear (quadratic) relationships by repeating the GLM analyses with age and age2 as simultaneous predictors. This analysis was run in each of the samples separately. In no case did age2 yield significant effects (FDR < 0.05) on cortical thickness.
Discussion
The present study investigated age effects on the cerebral cortex across multiple large samples, providing a unique opportunity to assess degree of consistency. Because the same preprocessing procedures were performed for all brains, a significant component of interstudy variability was eliminated. Several conclusions can be drawn from the findings. First, advanced age was associated with widespread thinning of the cerebral cortex. When a commonly employed statistical threshold (FDR < 0.05) was used, thinning was observed across most of the cortical surface. Second, the magnitude of age differences in thickness varied across cortical regions. Age effects were strongest in the prefrontal cortex, especially in superior, lateral, and medial regions. The superior temporal gyri at the lower bank of the Sylvian fissure were also heavily affected. In contrast, inferior temporal, parahippocampal, and entorhinal gyri, anterior cingulate, paracentral gyrus and the precuneus, were relatively more preserved. Third, although there was generally good agreement among the different samples, some discrepancies were also observed. For example, in one of the samples (sample 3), the effects of age were weaker than in the other 5. Still, also in that sample, clear age-related thinning was found in the prefrontal cortex. A puzzling finding was thickening of an area in medial prefrontal cortex that was observed in the 3 American but not in the 3 Scandinavian samples. Unstable results were also found in medial-temporal areas (parahippocampal and entorhinal gyri) and occipital cortices. Still, large areas were consistently affected across all or 5 of the 6 samples, that is, the superior, middle, and inferior frontal cortices, superior and middle temporal cortices, temporoparietal junction and pre- and postcentral gyri. Finally, nonlinear age effects were not observed within any of the samples.
Consistency of Effects of Age on Cortical Thickness
The degree of consistency of age-related differences in cortical thickness across samples depended on spatial location. Some regions, such as most of the lateral prefrontal as well as superior parts of the medial prefrontal cortices, showed age-related thinning in virtually all samples, whereas others, such as the occipital cortices, were affected by age in some samples only. Further, inferior temporal cortices, including the entorhinal and parahippocampal cortices, were not consistently related to age across samples. However, the superior and middle temporal gyri showed consistent age effects. Motor cortex was related to age in 4–6 of the samples, depending on region. Age effects in these regions were also reported by Salat et al. (2004). The ACC was not consistently prone to thinning with age, and showed increased thickness in some of the samples. Finally, parts of the medial orbitofrontal cortex showed preservation with age, both in American and Scandinavian samples. In addition to the inconsistencies evident from the P-thresholded statistics, interaction effects of sample × age on cortical thickness were also found in areas of consistent thinning (e.g., superior frontal gyrus). These effects were due to differences between samples in the magnitude of age effects, as illustrated in Figure 7.
When all the samples were pooled together, age effects were seen across almost the entire brain surface. This indicates that if the number of participants gets high enough, global age effects are detected. Further, it indicates that data from different studies and scanners may increase the sensitivity to age effects, even in areas where consistency across samples is not impressive. For instance, age effects in inferior temporal areas (parahippocampal, entorhinal, inferior temporal gyri) were not seen consistently across samples, but thickness in these areas showed up as related to age in the analysis involving all the samples. Thus, it may be beneficial to pool data from different studies, samples, and scanners to increase the sensitivity of the tests employed.
As reviewed, previous studies have yielded partly inconsistent results. We found that when using identical approaches to data processing and analysis, there was generally good consistency across samples. Even though inconsistent results were found for some cortical regions, these inconsistencies were much smaller than what would have been expected from previous literature. A substantial part of the variability in previous reports may be due to one or more of at least 5 factors: 1) Different statistical analyses performed: The exact results obtained depend on the analytic strategy used, for example, which variables are corrected for. Differences in strategy may lead to inconsistent results. 2) Different demarcation criteria for ROI analyses: Automated segmentation approaches allow statistics to be done without predefined ROIs. This reduces the problem of different definitions of brain areas between studies. 3) Image quality: The image quality differs between studies, leading to different SNR and contrast-to-noise ratio (CNR). In the present study, relatively consistent results, at least in some brain areas, were found across samples from different scanners and sequences. 4) Different segmentation procedures: The tools used to segment the MR scans into tissue classes, and the procedures used to spatially normalize the data, may have a huge impact on the results (Tisserand et al. 2002). Of the studies using automated/semiautomated techniques reviewed in the introduction, the majority were based on voxel-based morphometry (VBM). In the present study, a surface-based approach was used, similar to the basic methods of Salat and colleagues (Salat et al. 2004). We believe that this approach has advantages compared with the traditional VBM method. FreeSurfer uses surface geometry in intersubject registration, which in our experience results in a better matching of homologous cortical regions. The target used for registration is the white matter surface geometry, which makes the registration invariant to morphometric changes in GM. Hence, the registration is the same even in case of GM atrophy, which is important when studying groups where this is expected, such as in aging and dementia. However, this approach will not be invariant to WM changes that correlate with changes in GM. The approach used also allows separation of the 2 components of volume (thickness and surface area). These do not necessarily follow each other (Dickerson, Feczko, et al. 2008), and to be able to study each in isolation may increase both accuracy and sensitivity to brain changes. 5) Sample characteristics: Differences in sample characteristics may be an important source of discrepancies. For example, between-sample inconsistency observed with regards to age differences in occipital (pericalcarine) and medial-temporal (entorhinal) cortices may stem from sample-specific contribution of unmeasured vascular risk factors. A longitudinal study showed that unlike healthy adults, persons with known vascular risk, for example, hypertension, show shrinkage of the pericalcarine regions as well as proliferation of occipital white matter abnormalities (Raz, Rodrigue, Kennedy, et al. 2007).
In the present study, factors 1, 2, and 4 were eliminated. Thus, the inconsistencies that were observed may be attributed to differences in scan quality between the samples or differences in sample characteristics. Even though all samples were well screened, it is certainly possible that unmeasured differences in somatic or psychological factors have influence on discrepancies between the samples. Also, the age range sampled may have affected the results. For instance, the age range in sample 3 is narrower than in the other samples, which may have contributed to the somewhat lower age correlations in this study. One could speculate that the 3 Nordic samples were more alike regarding culturally dependent factors, such as nutrition, education, and medical care, because health care and education are free in Norway and Sweden. However, this does not appear to be a major factor of influence here, because the findings for samples 1 and 2 (Norway) was very similar to the findings from samples 4 and 5 (United States). Further, level of education was fairly similar across the samples. Regarding scan quality, all samples but samples 3 and 6 were based on at least 2 T1 runs that were averaged. Averaging of multiple runs may enhance the SNR/CNR, contributing to more valid and thickness estimates. This could contribute to the somewhat higher age correlations in these samples. However, it needs to be stressed that even though interaction effects were found, the general pattern of effects was highly replicable across the different samples.
Comparing automated and manual methods was not a goal of the study, but the regional distribution of the observed effects of age on cortical thickness corresponded quite well to the age differences reported in a previous study with manual volume measures. One dataset used in this study (sample 6) almost completely overlapped with the sample that has previously been used in a manual morphometry study (Raz, Gunning-Dixon, et al. 2004), where volume was calculated in 9 different cortical ROIs. The main results from that investigation are in good agreement with the findings from the present study although the overlap is not complete. Corresponding age effects were found for lateral prefrontal and orbitofrontal cortices, precentral gyrus, postcentral gyrus and fusiform gyrus, and no age differences in the anterior cingulate gyrus and the visual cortex were observed with either method. However, the previously reported age effect on parahippocampal gyrus and inferior temporal cortex was not found in the present analysis, and the present but not the previous study found an effect in the inferior parietal lobule. Still, an agreement this close is remarkable in the light of the manifest differences in segmentation approach, the ROI versus vertex-based statistics, the volume vs. thickness measures, and the fact that adjustments for body height/intracranial volume differed between the studies.
Effects of Age versus Aging
A limitation of the present study is that the estimates of age-related changes are based on cross-sectional data. Thus, the observed effects reflect the influences associated with chronological age, not necessarily the process of aging. To assess individual variability in change requires a longitudinal design. Unfortunately, only a handful of longitudinal studies of regional brain changes have been conducted so far. Comparison of cross-sectional and longitudinal effects indicates that the latter sometimes reveal aging trends that would not be observed with a cross-sectional design alone (Du et al. 2006). This means that cross-sectional studies, possibly due to the large inter-individual variability in initial brain volumes, may underestimate real age changes. However, longitudinal data have also been found to largely agree with cross-sectional estimates (Resnick et al. 2003; Fotenos et al. 2005). Longitudinal studies have methodological problems as well, e.g. selective drop out. Participants who are less healthy, less socioeconomically endowed, and prone to greater emotional distress are less likely to volunteer for multiple measurements, thus biasing estimates of change. Also, it is extremely difficult to collect longitudinal data comparable to the 6 decade range covered in most cross-sectional studies. Even relatively short follow-ups of 5–6 years are difficult to complete due to scanner changes and upgrades, which may affect the volumetric estimates (Han et al. 2006). Thus, both cross-sectional and longitudinal designs remain necessary for studying the aging brain.
Implication of the Findings for Cognitive Aging: The Frontal Lobe Hypothesis
Although investigations of brain aging patterns are interesting in their own right, their primary importance is for advancing our understanding of the mechanisms underpinning adult cognitive development and aging. Previous studies have provided some evidence of associations between neuroanatomical age-related differences in specific regions and performance on specific age-sensitive cognitive tasks. For instance, smaller prefrontal cortices seem to predict poorer performance on age-sensitive tests of executive functions (Raz et al. 1998; Gunning-Dixon and Raz 2003) and other cognitive operations dependent on executive control (Head et al. 2008). This is in line with neuropsychological studies showing that executive functions, which heavily depend on frontal neural circuits (e.g., fronto-striatal circuits), are among the cognitive functions to decline most with advancing age (Connelly et al. 1991; Schretlen et al. 2000). Previous research, as well as the present findings, support the selective vulnerability of the prefrontal cortex in aging. This fits the so-called “last in, first out” hypothesis, according to which the brain areas that are latest to develop phylogenetically and ontogenetically are the first to be affected by aging. An important aspect in the frontal lobe theory of aging is the assumed reduced efficiency of the executive function inhibition (Connelly et al. 1991), for example, as assessed by the Stroop task (but see Rabbitt et al. 2001). In functional MRI studies, the center of activation in such tasks is typically within the anterior cingulate gyrus (Bush et al. 2000). However, the present results indicate that this area of the prefrontal cortex is relatively spared. Thus, there is no simple correspondence between morphometric age-related changes in prefrontal cortex and reduced inhibition in aging.
Relatively weaker effects were seen in parahippocampal and entorhinal gyri, which are affected early in the AD disease process (Braak and Braak 1991). Taken together with the findings of frontal morphometric reductions, this fits with a proposal of a double dissociation between AD and healthy aging. According to this theory, early AD selectively attacks the medial-temporal lobes and adjacent cortical areas, leading to memory problems, whereas an anterior to posterior gradient exists in healthy aging, causing executive problems (Head et al. 2005). Still, reduced memory function is seen also in healthy aging (Craik and Jennings 1992), and memory complaints are common in elderly, reported by up to 50% of people aged 64 and over (Reid and Maclullich 2006). Thus, because recollective memory also depends on frontal cortical structures (Craik and Grady 2002), it is possible that reduced memory function in healthy aging is related to the morphometric changes observed in these brain areas (West 1996).
Limitations
An important limitation of MRI studies of aging is the maximum CNR that can be achieved. Lower CNR will decrease the accuracy of the thickness estimation, and will probably vary across the cortical surface (Han et al. 2006), for example, due to some areas with higher degree of myelination than other, as in visual areas (Braitenberg and Schuz 1991). Also, the narrow separation between putamen or hippocampus and the adjacent cortical GM may cause a problem in finding the GM/WM surface around the insular and entorhinal cortical regions, which may again increase the variability of the thickness estimates (Han et al. 2006). If this is not systematically related to age, it will probably reduce the age-relationships observed. If it is systematically related to age, it may falsely enhance or create age-relationships. Future research efforts should try to estimate to what extent regional differences in age correlations can be attributed to regional differences in CNR. The problem of varying CNR across the cortex affects all segmentation techniques. Another question regards the need for smoothing of data, which can reduce noise and thus improve reliability of the thickness estimates (Han et al. 2006), at the cost of lower spatial resolution. Han and colleagues have shown that thickness measurement variability becomes smaller as smoothing level increases (Han et al. 2006). The current smoothing level of 15-mm FWHM will be sufficient to reduce thickness measurement variability by at least 50%. By use of manual methods, smoothing would not be necessary. However, thickness cannot be reliably measured by manual methods, because both the localization and the orientation of the white and pial surfaces must be known for proper measurements to be obtained (Fischl and Dale 2000).
Conclusion
The present study shows that age-related differences in cortical thickness are widespread and robust, and are especially strong in the superior and inferior frontal gyri and in the superior temporal cortices. These findings were obtained from data from 6 different samples, from 3 countries, and with use of scanners from 2 manufacturers. The automated approach employed in this study appears a reliable method to boost the statistical power of the studies of brain aging and to improve generalizability of the findings across various populations. It is concluded that consistent effects of age are likely to be found when the segmentation techniques employed are standardized across samples and studies.
Supplementary Material
Supplementary material can be found at: http://www.cercor.oxfordjournals.org/
Funding
The Norwegian Research Council (177404/W50) to K.B.W., (175066/D15) to A.M.F., (154313/V50) to I.R., (177458/V50) to T.E.; University of Oslo to K.B.W. and A.M.F.; the National Institutes of Health (R01-NS39581, R37-AG11230, and R01-RR13609); the Mental Illness and Neuroscience Discovery Institute; The Wallenberg Foundation and the Swedish Medical Research Council (K2004-21X-15078-01A 45, K2007-62X-15077-04-1, and K2007-62X-15078-04-3). The National Center for Research Resources (P41-RR14075, R01 RR16594-01A1 and the NCRR BIRN Morphometric Project BIRN002, U24 RR021382); the National Institute for Biomedical Imaging and Bioengineering (R01 EB001550, R01EB006758); the National Institute for Neurological Disorders and Stroke (R01 NS052585-01); as well as the Mental Illness and Neuroscience Discovery Institute; and is part of the National Alliance for Medical Image Computing (NAMIC), funded by the National Institutes of Health through the NIH Roadmap for Medical Research (grant U54 EB005149); additional support was provided by The Autism & Dyslexia Project funded by the Ellison Medical Foundation.
Acknowledgments
We thank Vivi Agnete Larsen for assistance in processing of the data. We thank the developers of the OASIS (Open Access Series of Imaging Studies) database for access to MRI data constituting samples 4 and 5 of the present work. Conflict of Interest: Anders M. Dale is a founder and holds equity in CorTechs Labs, Inc., and also serves on the Scientific Advisory Board.
References
- Abe O, Yamasue H, Aoki S, Suga M, Yamada H, Kasai K, Masutani Y, Kato N, Kato N, Ohtomo K. Aging in the CNS: comparison of gray/white matter volume and diffusion tensor data. Neurobiol Aging. 2008;29:102–116. doi: 10.1016/j.neurobiolaging.2006.09.003. [DOI] [PubMed] [Google Scholar]
- Allen JS, Bruss J, Brown CK, Damasio H. Normal neuroanatomical variation due to age: the major lobes and a parcellation of the temporal region. Neurobiol Aging. 2005;26:1245–1260. doi: 10.1016/j.neurobiolaging.2005.05.023. discussion 1279–1282. [DOI] [PubMed] [Google Scholar]
- Auer S, Reisberg B. The GDS/FAST staging system. Int Psychogeriatr. 1997;9(Suppl. 1):167–171. doi: 10.1017/s1041610297004869. [DOI] [PubMed] [Google Scholar]
- Beck AaSR. Beck Depression Inventory scoring manual. New York: The Psychological Corporation; 1987. [Google Scholar]
- Berg L. Clinical Dementia Rating. Br J Psychiatry. 1984;145:339. [PubMed] [Google Scholar]
- Berg L. Clinical Dementia Rating (CDR) Psychopharmacol Bull. 1988;24:637–639. [PubMed] [Google Scholar]
- Blatter DD, Bigler ED, Gale SD, Johnson SC, Anderson CV, Burnett BM, Parker N, Kurth S, Horn SD. Quantitative volumetric analysis of brain MR: normative database spanning 5 decades of life. AJNR Am J Neuroradiol. 1995;16:241–251. [PMC free article] [PubMed] [Google Scholar]
- Blessed G, Tomlinson BE, Roth M. The association between quantitative measures of dementia and of senile change in the cerebral grey matter of elderly subjects. Br J Psychiatry. 1968;114:797–811. doi: 10.1192/bjp.114.512.797. [DOI] [PubMed] [Google Scholar]
- Braak H, Braak E. On areas of transition between entorhinal allocortex and temporal isocortex in the human brain. Normal morphology and lamina-specific pathology in Alzheimer's disease. Acta Neuropathol Berl. 1985;68:325–332. doi: 10.1007/BF00690836. [DOI] [PubMed] [Google Scholar]
- Braak H, Braak E. Neuropathological staging of Alzheimer-related changes. Acta Neuropathol. 1991;82:239–259. doi: 10.1007/BF00308809. [DOI] [PubMed] [Google Scholar]
- Braitenberg V, Schuz A. Anatomy of the cortex. Berlin: Springer; 1991. [Google Scholar]
- Brickman AM, Habeck C, Zarahn E, Flynn J, Stern Y. Structural MRI covariance patterns associated with normal aging and neuropsychological functioning. Neurobiol Aging. 2007;28:284–295. doi: 10.1016/j.neurobiolaging.2005.12.016. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci. 2000;4:215–222. doi: 10.1016/s1364-6613(00)01483-2. [DOI] [PubMed] [Google Scholar]
- Connelly SL, Hasher L, Zacks RT. Age and reading: the impact of distraction. Psychol Aging. 1991;6:533–541. doi: 10.1037//0882-7974.6.4.533. [DOI] [PubMed] [Google Scholar]
- Courchesne E, Chisum HJ, Townsend J, Cowles A, Covington J, Egaas B, Harwood M, Hinds S, Press GA. Normal brain development and aging: quantitative analysis at in vivo MR imaging in healthy volunteers. Radiology. 2000;216:672–682. doi: 10.1148/radiology.216.3.r00au37672. [DOI] [PubMed] [Google Scholar]
- Craik FIM, Grady CL. Aging, memory, and frontal lobe functioning. In: Stuss DT, Knight RT, editors. Principles of frontal lobe function. Oxford: Oxford University Press; 2002. pp. 528–540. [Google Scholar]
- Craik FIM, Jennings JJ. Human memory. In: Craik FIM, Salthouse TA, editors. The handbook of aging and cognition. Hillsdale (NJ): Lawrence Erlbaum Associates; 1992. pp. 51–110. [Google Scholar]
- Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage. 1999;9:179–194. doi: 10.1006/nimg.1998.0395. [DOI] [PubMed] [Google Scholar]
- Dale AM, Sereno MI. Improved localization of cortical activity by combining EEG and MEG with MRI cortical surface reconstruction: a linear approach. J Cogn Neurosci. 1993;5:162–176. doi: 10.1162/jocn.1993.5.2.162. [DOI] [PubMed] [Google Scholar]
- Desikan RS, Segonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM, Maguire RP, Hyman BT, et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. 2006;31:968–980. doi: 10.1016/j.neuroimage.2006.01.021. [DOI] [PubMed] [Google Scholar]
- Dickerson BC, Feczko E, Augustinack JC, Pacheco J, Morris JC, Fischl B, Buckner RL 2008. Differential effects of aging and Alzheimer's disease on medial temporal lobe cortical thickness and surface area. Neurobiol Aging. doi: 10.1016/j.neurobiolaging.2007.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickerson BC, Fenstermacher E, Salat DH, Wolk DA, Maguire RP, Desikan R, Pacheo J, Quinn BT, Van der Kouwe A, Greve DN, et al. Detection of cortical thickness correlates of cognitive performance: Reliability across MRI scan sessions, scanners and field strength. Neuroimage. 2008;39:10–18. doi: 10.1016/j.neuroimage.2007.08.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du AT, Schuff N, Chao LL, Kornak J, Jagust WJ, Kramer JH, Reed BR, Miller BL, Norman D, Chui HC, et al. Age effects on atrophy rates of entorhinal cortex and hippocampus. Neurobiol Aging. 2006;27:733–740. doi: 10.1016/j.neurobiolaging.2005.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espeseth T, Westlye LT, Fjell AM, Walhovd KB, Rootwelt H, Reinvang I. Accelerated age-related cortical thinning in healthy carriers of apolipoprotein E varepsilon4. Neurobiol Aging. 2008;29:329–340. doi: 10.1016/j.neurobiolaging.2006.10.030. [DOI] [PubMed] [Google Scholar]
- Fischl B, Dale AM. Measuring the thickness of the human cerebral cortex from magnetic resonance images. Proc Natl Acad Sci USA. 2000;97:11050–11055. doi: 10.1073/pnas.200033797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis. 2: Inflation, flattening, and a surface-based coordinate system. Neuroimage. 1999;9:195–207. doi: 10.1006/nimg.1998.0396. [DOI] [PubMed] [Google Scholar]
- Fischl B, van der Kouwe A, Destrieux C, Halgren E, Segonne F, Salat DH, Busa E, Seidman LJ, Goldstein J, Kennedy D, et al. Automatically parcellating the human cerebral cortex. Cereb Cortex. 2004;14:11–22. doi: 10.1093/cercor/bhg087. [DOI] [PubMed] [Google Scholar]
- Fjell AM, Walhovd KB, Amlien I, Bjørnerud A, Reinvang I, Gjerstad L, Cappelen T, Willoch F, Due-Tønnessen P, Grambaite R, et al. Morphometric changes in the episodic memory network and tau pathologic features correlate with memory performance in patients with mild cognitive impairment. Am J Neuroradiol. 2008;29:1–7. doi: 10.3174/ajnr.A1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folstein MF, Folstein SE, McHugh PR. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res. 1975;12:189–198. doi: 10.1016/0022-3956(75)90026-6. [DOI] [PubMed] [Google Scholar]
- Fotenos AF, Snyder AZ, Girton LE, Morris JC, Buckner RL. Normative estimates of cross-sectional and longitudinal brain volume decline in aging and AD. Neurology. 2005;64:1032–1039. doi: 10.1212/01.WNL.0000154530.72969.11. [DOI] [PubMed] [Google Scholar]
- Good CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ, Frackowiak RS. A voxel-based morphometric study of ageing in 465 normal adult human brains. Neuroimage. 2001;14:21–36. doi: 10.1006/nimg.2001.0786. [DOI] [PubMed] [Google Scholar]
- Gunning-Dixon FM, Raz N. Neuroanatomical correlates of selected executive functions in middle-aged and older adults: a prospective MRI study. Neuropsychologia. 2003;41:1929–1941. doi: 10.1016/s0028-3932(03)00129-5. [DOI] [PubMed] [Google Scholar]
- Han X, Jovicich J, Salat D, van der Kouwe A, Quinn B, Czanner S, Busa E, Pacheco J, Albert M, Killiany R, et al. Reliability of MRI-derived measurements of human cerebral cortical thickness: the effects of field strength, scanner upgrade and manufacturer. Neuroimage. 2006;32:180–194. doi: 10.1016/j.neuroimage.2006.02.051. [DOI] [PubMed] [Google Scholar]
- Head D, Rodrigue KM, Kennedy KM, Raz N. Neuroanatomical and cognitive mediators of age-related differences in episodic memory. Neuropsychology. 2008;22:491–507. doi: 10.1037/0894-4105.22.4.491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Head D, Snyder AZ, Girton LE, Morris JC, Buckner RL. Frontal-hippocampal double dissociation between normal aging and Alzheimer's disease. Cereb Cortex. 2005;15:732–739. doi: 10.1093/cercor/bhh174. [DOI] [PubMed] [Google Scholar]
- Jernigan TL, Archibald SL, Berhow MT, Sowell ER, Foster DS, Hesselink JR. Cerebral structure on MRI, Part I: Localization of age-related changes. Biol Psychiatry. 1991;29:55–67. doi: 10.1016/0006-3223(91)90210-d. [DOI] [PubMed] [Google Scholar]
- Jernigan TL, Archibald SL, Fennema-Notestine C, Gamst AC, Stout JC, Bonner J, Hesselink JR. Effects of age on tissues and regions of the cerebrum and cerebellum. Neurobiol Aging. 2001;22:581–594. doi: 10.1016/s0197-4580(01)00217-2. [DOI] [PubMed] [Google Scholar]
- Jonsson EG, Edman-Ahlbom B, Sillen A, Gunnar A, Kulle B, Frigessi A, Vares M, Ekholm B, Wode-Helgodt B, Schumacher J, et al. Brain-derived neurotrophic factor gene. BDNF. Variants and schizophrenia: an association study. Prog Neuropsychopharmacol Biol Psychiatry. 2006;30:924–933. doi: 10.1016/j.pnpbp.2006.02.008. [DOI] [PubMed] [Google Scholar]
- Kalpouzos G, Chetelat G, Baron JC, Landeau B, Mevel K, Godeau C, Barre L, Constans JM, Viader F, Eustache F, et al. Voxel-based mapping of brain gray matter volume and glucose metabolism profiles in normal aging. Neurobiol Aging. 2008;30:112–124. doi: 10.1016/j.neurobiolaging.2007.05.019. [DOI] [PubMed] [Google Scholar]
- Kennedy KM, Erickson KI, Rodrigue KM, Voss MW, Colcombe SJ, Kramer AF, Acker JD, Raz N. Age-related differences in regional brain volumes: a comparison of optimized voxel-based morphometry to manual volumetry. Neurobiol Aging. 2008 doi: 10.1016/j.neurobiolaging.2007.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruggel F. MRI-based volumetry of head compartments: normative values of healthy adults. Neuroimage. 2006;30:1–11. doi: 10.1016/j.neuroimage.2005.09.063. [DOI] [PubMed] [Google Scholar]
- Kuperberg GR, Broome MR, McGuire PK, David AS, Eddy M, Ozawa F, Goff D, West WC, Williams SC, van der Kouwe AJ, Salat DH, Dale AM, Fischl B. Regionally localized thinning of the cerebral cortex in schizophrenia. Arch Gen Psychiatry. 2003;60:878–888. doi: 10.1001/archpsyc.60.9.878. [DOI] [PubMed] [Google Scholar]
- Marcus DS, Wang TH, Parker J, Csernansky JG, Morris JC, Buckner RL. Open Access Series of Imaging Studies (OASIS): cross-sectional MRI data in young, middle aged, nondemented, and demented older adults. J Cogn Neurosci. 2007;19:1498–1507. doi: 10.1162/jocn.2007.19.9.1498. [DOI] [PubMed] [Google Scholar]
- Mesulam MM. Neuroplasticity failure in Alzheimer's disease: bridging the gap between plaques and tangles. Neuron. 1999;24:521–529. doi: 10.1016/s0896-6273(00)81109-5. [DOI] [PubMed] [Google Scholar]
- Morris JC. The Clinical Dementia Rating (CDR): current version and scoring rules. Neurology. 1993;43:2412–2414. doi: 10.1212/wnl.43.11.2412-a. [DOI] [PubMed] [Google Scholar]
- Murphy DG, DeCarli C, McIntosh AR, Daly E, Mentis MJ, Pietrini P, Szczepanik J, Schapiro MB, Grady CL, Horwitz B, et al. Sex differences in human brain morphometry and metabolism: an in vivo quantitative magnetic resonance imaging and positron emission tomography study on the effect of aging. Arch Gen Psychiatry. 1996;53:585–594. doi: 10.1001/archpsyc.1996.01830070031007. [DOI] [PubMed] [Google Scholar]
- Nesvag R, Lawyer G, Varnas K, Fjell AM, Walhovd KB, Frigessi A, Jonsson EG, Agartz I. Regional thinning of the cerebral cortex in schizophrenia: effects of diagnosis, age and antipsychotic medication. Schizophr Res. 2008;98:16–28. doi: 10.1016/j.schres.2007.09.015. [DOI] [PubMed] [Google Scholar]
- Pfefferbaum A, Mathalon DH, Sullivan EV, Rawles JM, Zipursky RB, Lim KO. A quantitative magnetic resonance imaging study of changes in brain morphology from infancy to late adulthood. Arch Neurol. 1994;51:874–887. doi: 10.1001/archneur.1994.00540210046012. [DOI] [PubMed] [Google Scholar]
- Rabbitt P, Lowe C, Shilling V. Frontal tests and models for cognitive ageing. Eur J Cogn Psychol. 2001;13:5–28. [Google Scholar]
- Raz N, Gunning FM, Head D, Dupuis JH, McQuain J, Briggs SD, Loken WJ, Thornton AE, Acker JD. Selective aging of the human cerebral cortex observed in vivo: differential vulnerability of the prefrontal gray matter. Cereb Cortex. 1997;7:268–282. doi: 10.1093/cercor/7.3.268. [DOI] [PubMed] [Google Scholar]
- Raz N, Gunning-Dixon FM, Head D, Dupuis JH, Acker JD. Neuroanatomical correlates of cognitive aging: evidence from structural magnetic resonance imaging. Neuropsychology. 1998;12:95–114. doi: 10.1037//0894-4105.12.1.95. [DOI] [PubMed] [Google Scholar]
- Raz N, Gunning-Dixon F, Head D, Rodrigue KM, Williamson A, Acker JD. Aging, sexual dimorphism, and hemispheric asymmetry of the cerebral cortex: replicability of regional differences in volume. Neurobiol Aging. 2004;25:377–396. doi: 10.1016/S0197-4580(03)00118-0. [DOI] [PubMed] [Google Scholar]
- Raz N, Lindenberger U, Ghisletta P, Rodrigue KM, Kennedy KM, Acker JD. Neuroanatomical correlates of fluid intelligence in healthy adults and persons with vascular risk factors. Cereb Cortex. 2008;18:718–726. doi: 10.1093/cercor/bhm108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D, Acker JD. Regional brain changes in aging healthy adults: general trends, individual differences and modifiers. Cereb Cortex. 2005;15:1676–1689. doi: 10.1093/cercor/bhi044. [DOI] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM. Differential aging of the brain: Patterns, cognitive correlates and modifiers. Neurosci Biobehav Rev. 2006;30:730. doi: 10.1016/j.neubiorev.2006.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM, Haacke EM. Brain aging and its modifiers: insights from in vivo neuromorphometry and susceptibility weighted imaging. Ann N Y Acad Sci. 2007;1097:84–93. doi: 10.1196/annals.1379.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM, Head D, Kennedy KM, Acker JD. Differential aging of the medial temporal lobe: a study of a five-year change. Neurology. 2004;62:433–438. doi: 10.1212/01.wnl.0000106466.09835.46. [DOI] [PubMed] [Google Scholar]
- Raz N, Rodrigue KM, Kennedy KM, Acker JD. Vascular health and longitudinal changes in brain and cognition in middle-aged and older adults. Neuropsychology. 2007;21:149–157. doi: 10.1037/0894-4105.21.2.149. [DOI] [PubMed] [Google Scholar]
- Raz N, Williamson A, Gunning-Dixon F, Head D, Acker JD. Neuroanatomical and cognitive correlates of adult age differences in acquisition of a perceptual-motor skill. Microsc Res Tech. 2000;51:85–93. doi: 10.1002/1097-0029(20001001)51:1<85::AID-JEMT9>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- Reid LM, Maclullich AM. Subjective memory complaints and cognitive impairment in older people. Dement Geriatr Cogn Disord. 2006;22:471–485. doi: 10.1159/000096295. [DOI] [PubMed] [Google Scholar]
- Resnick SM, Goldszal AF, Davatzikos C, Golski S, Kraut MA, Metter EJ, Bryan RN, Zonderman AB. One-year age changes in MRI brain volumes in older adults. Cereb Cortex. 2000;10:464–472. doi: 10.1093/cercor/10.5.464. [DOI] [PubMed] [Google Scholar]
- Resnick SM, Pham DL, Kraut MA, Zonderman AB, Davatzikos C. Longitudinal magnetic resonance imaging studies of older adults: a shrinking brain. J Neurosci. 2003;23:3295–3301. doi: 10.1523/JNEUROSCI.23-08-03295.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigue KM, Raz N. Shrinkage of the entorhinal cortex over five years predicts memory performance in healthy adults. J Neurosci. 2004;24:956–963. doi: 10.1523/JNEUROSCI.4166-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosas HD, Liu AK, Hersch S, Glessner M, Ferrante RJ, Salat DH, van der Kouwe A, Jenkins BG, Dale AM, Fischl B. Regional and progressive thinning of the cortical ribbon in Huntington's disease. Neurology. 2002;58:695–701. doi: 10.1212/wnl.58.5.695. [DOI] [PubMed] [Google Scholar]
- Salat DH, Buckner RL, Snyder AZ, Greve DN, Desikan RS, Busa E, Morris JC, Dale AM, Fischl B. Thinning of the cerebral cortex in aging. Cereb Cortex. 2004;14:721–730. doi: 10.1093/cercor/bhh032. [DOI] [PubMed] [Google Scholar]
- Sato K, Taki Y, Fukuda H, Kawashima R. Neuroanatomical database of normal Japanese brains. Neural Netw. 2003;16:1301–1310. doi: 10.1016/j.neunet.2003.06.004. [DOI] [PubMed] [Google Scholar]
- Schretlen D, Pearlson GD, Anthony JC, Aylward EH, Augustine AM, Davis A, Barta P. Elucidating the contributions of processing speed, executive ability, and frontal lobe volume to normal age-related differences in fluid intelligence. J Int Neuropsychol Soc. 2000;6:52–61. doi: 10.1017/s1355617700611062. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Marsh L, Mathalon DH, Lim KO, Pfefferbaum A. Age-related decline in MRI volumes of temporal lobe gray matter but not hippocampus. Neurobiol Aging. 1995;16:591–606. doi: 10.1016/0197-4580(95)00074-o. [DOI] [PubMed] [Google Scholar]
- Sullivan EV, Rosenbloom M, Serventi KL, Pfefferbaum A. Effects of age and sex on volumes of the thalamus, pons, and cortex. Neurobiol Aging. 2004;25:185–192. doi: 10.1016/s0197-4580(03)00044-7. [DOI] [PubMed] [Google Scholar]
- Taki Y, Goto R, Evans A, Zijdenbos A, Neelin P, Lerch J, Sato K, Ono S, Kinomura S, Nakagawa M, et al. Voxel-based morphometry of human brain with age and cerebrovascular risk factors. Neurobiol Aging. 2004;25:455–463. doi: 10.1016/j.neurobiolaging.2003.09.002. [DOI] [PubMed] [Google Scholar]
- Tisserand DJ, Pruessner JC, Sanz Arigita EJ, van Boxtel MP, Evans AC, Jolles J, Uylings HB. Regional frontal cortical volumes decrease differentially in aging: an MRI study to compare volumetric approaches and voxel-based morphometry. Neuroimage. 2002;17:657–669. [PubMed] [Google Scholar]
- Vaidya JG, Paradiso S, Boles Ponto LL, McCormick LM, Robinson RG. Aging, grey matter, and blood flow in the anterior cingulate cortex. Neuroimage. 2007;37:1346–1353. doi: 10.1016/j.neuroimage.2007.06.015. [DOI] [PubMed] [Google Scholar]
- Walhovd KB, Fjell AM, Reinvang I, Lundervold A, Dale AM, Eilertsen DE, Quinn BT, Salat D, Makris N, Fischl B. Effects of age on volumes of cortex, white matter and subcortical structures. Neurobiol Aging. 2005;26:1261–1270. doi: 10.1016/j.neurobiolaging.2005.05.020. discussion 1275–1268. [DOI] [PubMed] [Google Scholar]
- Wechsler D. Wechsler Abbreviated Scale of Intelligence. San Antonio (TX): Psychological Corporation; 1999. [Google Scholar]
- West RL. An application of prefrontal cortex function theory to cognitive aging. Psychol Bull. 1996;120:272–292. doi: 10.1037/0033-2909.120.2.272. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.